


Abstract
The transient receptor potential type V1 channel (vanilloid receptor 1, TRPV1) is a Ca2+-permeable nonspecific cation channel activated by various painful stimuli including ischemia. We hypothesized that TRPV1 is expressed in the arterioles and is involved in the regulation of microvascular tone. We found that TRPV1 stimulation by capsaicin (intra-arterial administration) of the isolated, perfused right hind limb of the rat increased vascular resistance (by 98 ± 21 mm Hg at 10 μg) in association with decreased skeletal muscle perfusion and elevation of skin perfusion (detected by dual-channel laser Doppler flowmetry). Denervation of the hind limb did not affect capsaicin-evoked changes in vascular resistance and tissue perfusion in the hind limb but reduced the elevation of perfusion in the skin. In isolated, pressurized skeletal (musculus gracilis) muscle arterioles (diameter, 147 ± 35 μm), capsaicin had biphasic effects: at lower concentrations, capsaicin (up to 10 nM) evoked dilations (maximum, 32 ± 13%), whereas higher concentrations (0.1-1 μM) elicited substantial constrictions (maximum, 66 ± 7%). Endothelium removal or inhibition of nitric-oxide synthase abolished capsaicin-induced dilations but did not affect arteriolar constriction. Expression of TRPV1 was detected by reverse transcriptase-polymerase chain reaction in the aorta and in cultured rat aortic vascular smooth muscle cells (A7r5). Immunohistochemistry revealed expression primarily in the smooth muscle layers of the gracilis arteriole. These data demonstrate the functional expression of TRPV1 in vascular smooth muscle cells mediating vasoconstriction of the resistance arteries. Because of the dual effects of TRPV1 stimulation on the arteriolar diameter (dilation in skin, constriction in skeletal muscle), we propose that TRPV1 ligands represent drug candidates for tissue-specific modulation of blood distribution.
The transient receptor potential type V1 channel (vanilloid receptor-1, TRPV1) is a nonselective cation channel, structurally belonging to the transient receptor potential family of ion channels. TRPV1 is found in sensory C and A-δ fibers (Caterina et al., 1997) and functions as a ligand-, proton-, and heat-activated molecular integrator of nociceptive stimuli in the periphery (Szallasi and Blumberg, 1999; Di Marzo et al., 2002a,b; Ross, 2003). Activation of TRPV1 leads to central (pain) as well as to local “sensory-efferent” effects (Szolcsányi, 1988).
It is well established that the sensory-efferent effects of TRPV1 stimulation include the release of neuropeptides such as calcitonin gene-related peptide (CGRP) and substance P (SP) from the sensory nerve terminals. These peptides cause vasodilatation in different vascular beds such as mesenteric, hepatic, basilar, dural, and meningeal arterioles (Zygmunt et al., 1999; Ralevic et al., 2001; Harris et al., 2002; Dux et al., 2003; Akerman et al., 2004; O'Sullivan et al., 2004). It is interesting that TRPV1-induced release of SP from sensory neurons has been implicated recently in mediating pressure-induced myogenic constriction (Scotland et al., 2004). Likewise, previous studies have also proposed that in certain circumstances, TRPV1 activation may lead to vasoconstriction in mesenteric (Pórszász et al., 2002), coronary (Szolcsányi et al., 2001), skeletal muscle (Lizanecz et al., 2006), and dural vessels (Dux et al., 2003), although the underlying mechanism remained obscure.
In the present study, the possible mechanisms of TRPV1-mediated vascular effects were investigated. It was found that TRPV1 stimulation results in opposite effects in different arterial beds from the same hind limb of the rat in vivo, namely vasodilation in the skin and vasoconstriction in the skeletal muscle. Moreover, investigation of the possible mechanisms of TRPV1-mediated responses confirmed TRPV1 expression in vascular smooth muscle cells and suggested cell type-specific differences in the capsaicin responsiveness.
Materials and Methods
Animals, Anesthesia, and General Preparation in the in Vivo Experiments. The experiments were performed on male Wistar rats weighing 250 to 450 g raised on a standard laboratory food and water ad libitum. Anesthesia was performed with 100 mg/kg i.p. thiobutobarbital (Inactin-Byk). The right common carotid artery and the left internal jugular vein were cannulated with polyethylene tubing for continuous measurement of arterial blood pressure and for administration of drugs, respectively. Respiratory movements were measured by means of a Statham transducer connected to one side of a Y-shaped cannula introduced into the trachea. The body core temperature was maintained at approximately 37°C with a temperature-controlled infrared heating lamp. Recordings were displayed on a polyphysiograph. All procedures used in this study are in agreement with the rules of the Ethics Committee on Animal Research.
Hind Limb Autoperfusion and Recording of Perfusion Pressure. Isolated hind limb autoperfusion and perfusion pressure recording were performed as described previously (Colquhoun et al., 1988). In brief, after administration of heparin sodium (1000 U/kg, i.v.), the right hind limb was perfused by means of a peristaltic pump (Masterflex; Cole-Parmer Instrument Co., Vernon Hills, IL) with blood taken from a catheter inserted in the left common carotid artery. The outlet side of the perfusion circuit (Masterflex Silicon tube platinum) was connected to the common iliac artery, which was approached through a midline abdominal incision. Perfusion pressure, measured from a side arm in the perfusion circuit between the pump and the iliac artery, was recorded using a Spectramed P23XL pressure transducer and displayed on a recorder. During the whole surgical procedure, the ischemic period of the leg never lasted longer than 3 to 5 min. The peristaltic pump was set to produce a constant flow rate (3 ml/min). This arrangement enabled us to record reproducible responses over 2 to 3 h without swelling of the paw. Capsaicin (8-methyl-N-vanillyl-trans-6-nonenamide), norepinephrine [(±)-4-(2-amino-1-hydroxyethyl)-1,2-benzenediol hydrochloride], and oxytocin (α-hypophamine) were administered into the perfusion cannula near to the iliac artery catheterization site. For administration, a Hamilton syringe was used, and the injected volumes varied between 10 and 100 μl. Acute cross-section of femoral, genitofemoral, and sciatic nerves were carried out in the right hind limb to prevent nociceptive reflexes from the perfused leg.
Measurement of Skin and Striated Muscle Blood Flow. Laser-Doppler recordings of microvascular blood flow changes were made in the middle of the paw covered by thin glabrous skin and from the flexor muscles of the thigh using a dual-channel laser Doppler flowmeter (MBF3D; Moor Instruments, Axminster, Devon, UK). The time constant was set to 1 s. During the experiment, the exposed skeletal muscle was kept moist by a wet chamber placed around the probe. Blood flow changes were recorded continuously throughout the experiment and were expressed as arbitrary units of flux (Escott and Brain, 1993; Pórszász and Szolcsányi, 1994). The zero level was verified at the end of the experiment. Disturbances caused by direct light were excluded by means of a piece of cotton wool placed onto the right hind limb. The peak of the changes was used to calculate the effect of drugs.
Long-Term Denervation of Hind Limb. In a group of animals (n = 15), the nervous supply of the right hind leg was denervated 7 to 20 days before the experiment under sodium pentobarbital [5-ethyl-5-(1-methylbutyl)-2,4,6-trioxohexahydropyrimidine] anesthesia (40 mg/kg, i.p.), as described previously (Sántha and Jancso, 2003). In brief, an incision was performed on the abdomen, and the right femoral, genitofemoral, and sciatic nerves were cross-sected transperitoneally. Then hemostasis was confirmed, and the wound was sutured. During recovery from the anesthesia, the animals were placed under an infrared heating lamp. Animals showing autotomy were not included in the experiments.
Isolation of Arterioles and Experimental Protocols. The isolation of the skeletal (gracilis) muscle arterioles of the rat and diameter measurement of the arterioles were performed as described earlier (Lizanecz et al., 2006). In brief, after spontaneous tone developed in response to intraluminal pressure of 80 mm Hg, arteriolar responses were obtained in maximal response to cumulative doses of the TRPV1 agonist, capsaicin (0.1 nmol/l to 1 μM). Capsaicin-induced responses were also observed after endothelium removal (Koller and Bagi, 2004) or after inhibition of NO synthase with Nω-nitro-l-arginine methyl ester (Koller and Bagi, 2004). In separate experiments, intraluminal pressure was changed from 20 to 120 mm Hg, and changes in diameter were measured before and after capsaicin treatment (1 μM for 20 min followed by a 40-min regeneration period) (Scotland et al., 2004).
Immunohistochemistry. The immunohistochemical experiments were performed as described by Lizanecz et al. (2006) with minor modifications. In short, musculus gracilis, skin, and small mesenteric tissues were dissected from Wistar rats and were embedded in Tissue-Tek OCT compound (Electron Microscopy Sciences, Hatfield PA). Cryostat sections (thickness, 10 μm) were placed on adhesive slides and fixed in acetone for 10 min. The slices were blocked with normal goat sera (1.5% in phosphate-buffered saline; Sigma, St. Louis, MO) for 20 min and stained with anticapsaicin receptor antibodies [AB 5370P (rabbit) and AB 5566 (guinea pig); Millipore Bioscience Research Reagents, Temecula, CA; PC 547 (rabbit); Calbiochem, San Diego, CA; and RA 10110 (rabbit) and GP 14100 (guinea pig); Neuromics, Edina, MN] at a 1:500 dilution (for all TRPV1-specific antibodies), with smooth muscle actin antibody (NCL-SMA, dilution, 1:20; Novocastra Laboratories, New Castle, UK) or with a neurofilament-specific antibody (dilution, 1:100; Sigma) in the blocking buffer. Then the slices were incubated with anti-rabbit, mouse, and guinea pig antibodies conjugated with Texas red or Cy2. The pictures were captured by a Scion Corporation (Frederick, MA) digital camera attached to a Nikon Eclipse 80i fluorescent microscope (Nikon, Tokyo, Japan).
Detection of TRPV1 mRNA. Total RNA was isolated from rat aorta and A7r5 cells (obtained from American Type Culture Collection, Manassas, VA) (LGC Promochem, Wesel, Germany) and maintained in 10% fetal bovine serum containing Dulbecco's modified Eagle's medium, both were from Invitrogen (Carlsbad, CA) with RNeasy RNS isolation kit (Qiagen GmbH, Hilden, Germany) and cDNA was synthesized by a RevertAid H Minus kit (Fermentas UAB, Vilnius, Lithuania), according to the manufacturer's instruction. The RT-PCR was performed by a sense (5′-CTACCTGGAACACCAATGTGGG-3′) and an antisense primer (5′-GCTGGGTGGCATGTCTATCTCG-3′) designed to produce a 596-bp fragment from DNA and a 149-bp fragment from RNA. Glyceraldehyde-3-phosphate dehydrogenase was used as control. Polymerase chain reaction was performed in a volume of 25 μl consisting of 1 μl of cDNA, 1 μM primer, 200 μM dNTP, 6 mM MgCl2, 50 mM KCl, 10 mM Tris-HCl, pH 8.3, and 2.5 U Long PCR Enzyme Mix (Fermentas UAB). The protocol was 94°C 5 min followed by 35 cycles of 20 s at 94°C, 20 s at 58°C, and 40 s at 72°C.
Materials and Solutions. Oxytocin (α-hypophamine), capsaicin (8-methyl-N-vanillyl-trans-6-nonenamide; Sigma), and resiniferatoxin [RTX; 6,7-deepoxy-6,7-didehydro-5-deoxy-21-dephenyl-21-(phenylmethyl)daphnetoxin 20-(3-hydroxy-5-methoxybenzeneacetate] (from Sigma or from LC Laboratories, Woburn, MA) were dissolved in ethanol/Tween 80/physiological saline in the ratio 1:1:8 for a 10 mg/ml capsaicin and a 100 μg/ml RTX stock solution, and further dilutions were made with saline. Norepinephrine and oxytocin from Gedeon Richter (Budapest, Hungary) were dissolved in physiological saline.
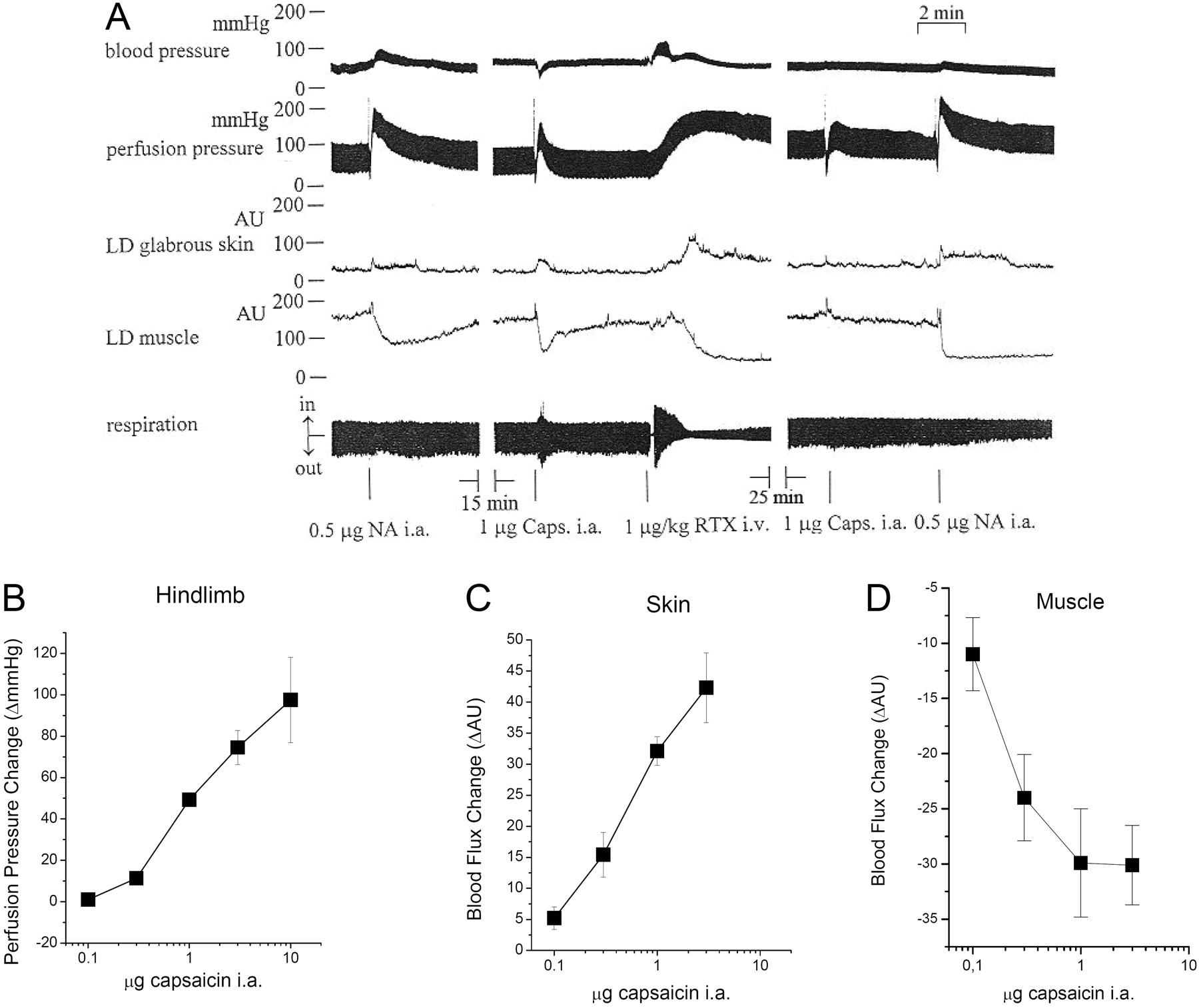
Effects of TRPV1 stimulation in perfused hind limb of the rat. The left common carotid artery and the common iliac artery were cannulated for blood perfusion of the right hind limb of the rat (3 ml/min). Systemic (arterial blood pressure and respiration) and local (perfusion pressure, blood flow in the skin and skeletal muscle) effects of noradrenaline (NA, 0.5 μg i.a.), capsaicin (Caps, 1 μg i.a.), and RTX (1 μg/kg i.v.) were recorded on the same preparation. A representative of the in vivo experiments performed is shown in A. The dose-response of capsaicin is shown on the perfusion pressure (B), on the blood flow of the skin (C), and skeletal muscle (D) on the right hind limb. Values are average ± S.E.M.
Statistical Analysis. The peak increase or decrease in skin or muscle blood flow (arbitrary units of flux) was determined after drug administration for the assessment of the blood flow. The change of the evoked effect was related to the values measured before drug administration and expressed as percentages (mean ± S.E.M.). Changes in arteriolar diameter were expressed as the percentage change to baseline diameter (constrictions) or in the case of the arteriolar dilations, the percentage change of the diameter as was related to the maximal dilation, determined at 80 mm Hg intraluminal pressure in a Ca2+-free medium. After the normality test, the statistical analysis was made by one-way analysis of variance, and pair-wise multiple comparisons were performed by Student-Newman-Keuls method. The diagrams were plotted using the computer program MicroCal Origin 5.0 (OriginLab Corp., Northampton, MA).
Results
It is well established that vanilloid receptor-1 (TRPV1)-mediated vasodilatation involves the activation of TRPV1 in sensory neuronal terminals and the consequent release of neurotransmitters evoking endothelial synthesis of NO (Zygmunt et al., 1999). The effects of TRPV1 stimulation by capsaicin on vascular resistance were tested in the isolated, perfused right hind limb of the rat in vivo. Changes in the systemic and perfusion pressure in the local blood flow in skeletal muscle and in skin of the same hind limb were measured simultaneously together with the respiration of the rat by pressure transducers and dual-channel laser-Doppler flowmetry (Fig. 1A). First, the responsiveness of the perfused rat hind limb preparations was tested by norepinephrine. Intra-arterial injection of norepinephrine (0.5 μg) resulted in an increase of both systemic and tissue blood pressure and a decrease in the blood flow in the perfused skeletal muscle, whereas no responses were detected in skin perfusion and respiration at this dose (Fig. 1). After a 15-min regeneration period, the effects of TRPV1 stimulations were tested. Injection of capsaicin (1 μg) into the arterial perfusion cannula of the blood-perfused right hind leg evoked a decrease in the systemic blood pressure, an increase in the perfusion pressure, a parallel increase in the cutaneous blood flow, and a decrease in blood perfusion of the muscle, whereas respiration was not affected by this dose. These responses were mimicked by the application of the ultrapotent TRPV1 agonist resiniferatoxin (1 μg/kg i.v.). In addition, the application of resiniferatoxin resulted in alterations in the respiration and desensitized vanilloid receptors, as shown by the unresponsiveness to repeated capsaicin stimulation after 25 min, whereas the noradrenaline responses were unaltered.

Effects of denervation on the perfusion pressure changes of the rat right hind limb in vivo. In a group of animals (n = 15), the nerves innervating the right hind leg were transected 7 to 20 days before the measurement. The effects of capsaicin (1 μg i.a.) and oxytocin (0.5 I.U.) were tested on the perfusion pressure in control and in denervated hind limbs. Values are average ± S.E.M.
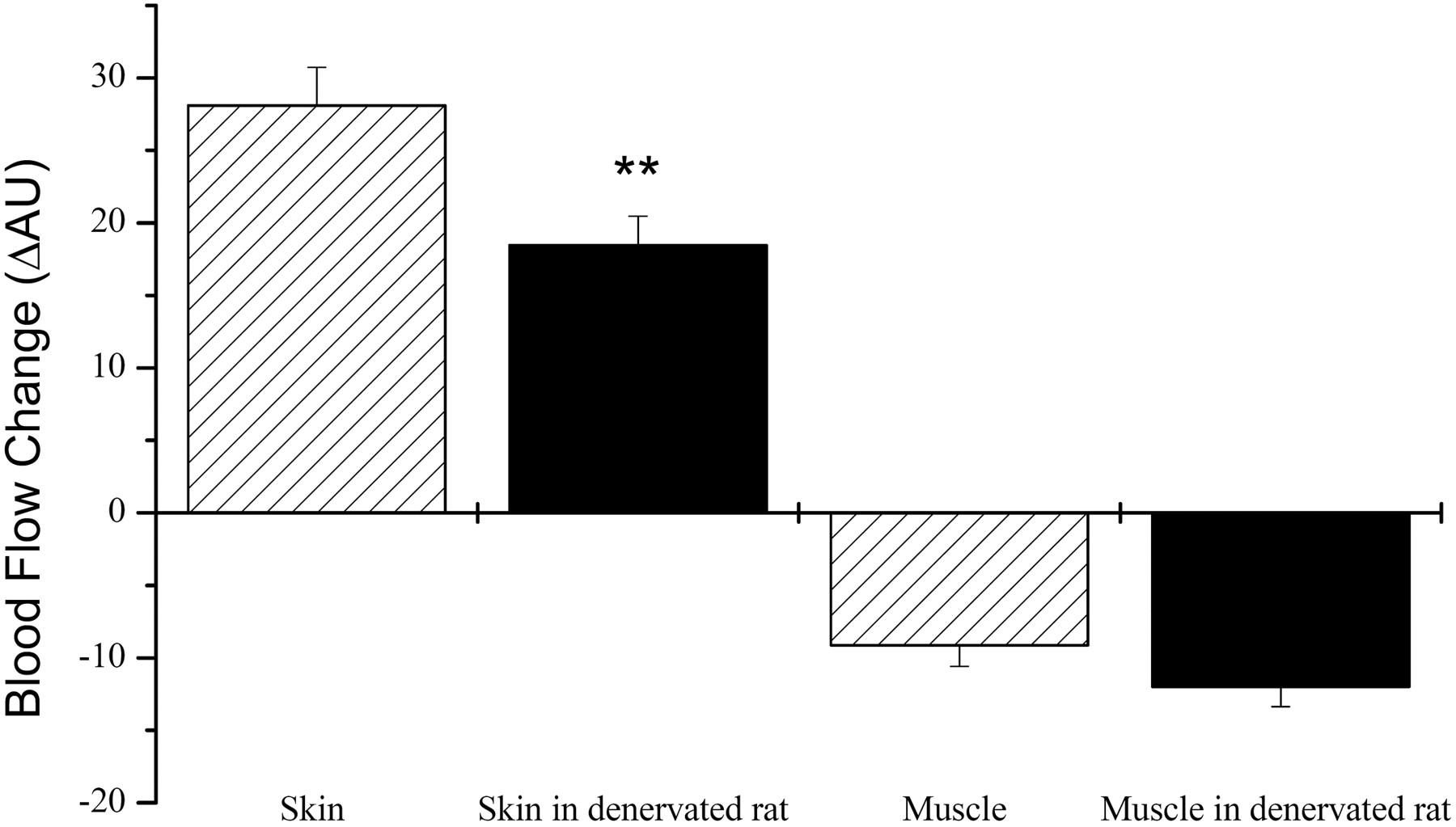
Effects of denervation on the blood flow changes of the rat right hind limb in vivo. Denervation was performed as mentioned earlier. The effects of capsaicin (1 μg i.a.) on the blood flow of skin and skeletal muscle were tested in control and denervated hind limbs. Values are average ± S.E.M., n = 7.
Next, the capsaicin-mediated responses were investigated in detail. Intra-arterial application of capsaicin (0.1-10 μg) resulted in a dose-dependent increase of perfusion pressure (98 ± 21 mm Hg increase at 10 μg, Fig. 1B), an increase in the blood flow in the skin (42 ± 5 AU increase at 3 μg; Fig. 1C) and a decrease in the blood flow in the skeletal muscle (30 ± 4 AU decrease at 3 μg; Fig. 1D).
To determine the role of neural elements in the capsaicin-induced responses, long-term denervation was performed. In a group of animals (n = 15), the genitofemoral, femoral, and sciatic nerves were cross-sected 7 to 20 days before the commencement of the experiment (see Materials and Methods). Denervation was not able to abolish capsaicin (1 μg, intra-arterial application)-evoked increase in the perfusion pressure (Fig. 2, increase in the perfusion pressure is 49 ± 3 mm Hg in control and 32 ± 3 mm Hg in denervated hind limbs), nor was it able to affect oxytocin (0.5 I.U.)-mediated elevations (49 ± 7 mm Hg increase in control and 56 ± 8 mm Hg in denervated hind limbs).
The effect of denervation was also tested on the local blood flow in the skin and in the skeletal muscle (Fig. 3). Capsaicin-evoked increases in the blood flow were decreased in the skin (28 ± 3 AU increase in the case of control versus 18 ± 2AU in the case of denervated right hind limb; P < 0.01, n = 7), but the decrease in the blood flow in the skeletal muscle was not affected (9 ± 1.5 AU decrease in the case of control, versus 12 ± 1.5 AU in the case of denervated; P = 0.25, n = 7).
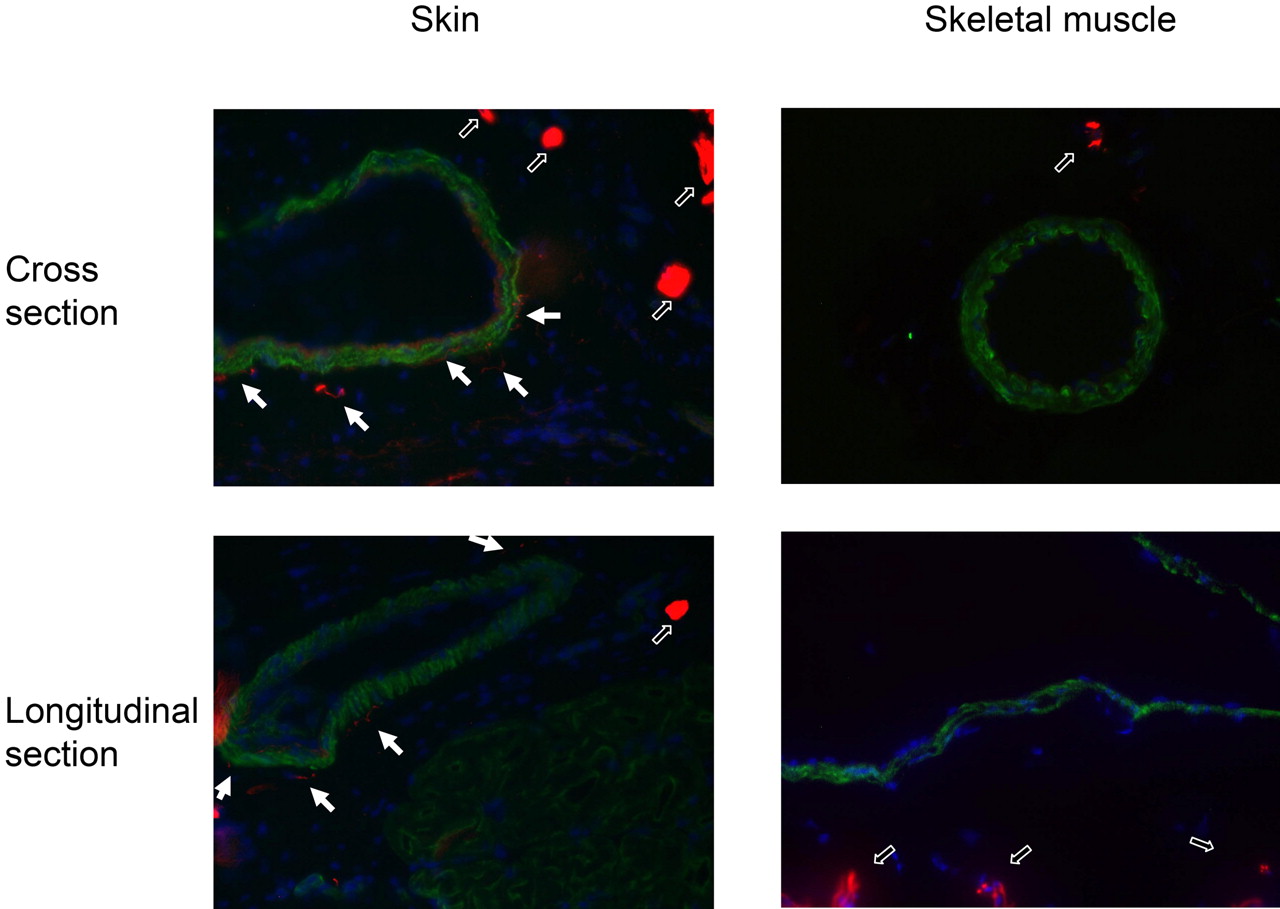
The presence of neurogenic innervation of the arteries in the skin and in the skeletal muscle was tested by immunohistochemistry (Fig. 4). Dense innervation of arteries was found in the skin (filled arrows in Fig. 4), but neurofilament-positive nerve terminals were not detected in the skeletal muscle arteries. In contrast, thicker nerves farther away from the vessels were detected in both skin and skeletal muscle tissue samples (labeled by open arrows in Fig. 4).
These results suggested non-neuronal vasoconstriction (decrease in the blood flow) upon TRPV1 stimulation in the skeletal muscle arterioles. To test this hypothesis, skeletal muscle (musculus gracilis) arteries were isolated and cannulated to directly measure the vasoactive effects of capsaicin. Effects of TRPV1 stimulations were tested after the spontaneous development of the myogenic tone in response to 80 mm Hg intraluminal pressure (spontaneous tone, 31 ± 4% of the maximal diameter). TRPV1 activation with capsaicin resulted in a biphasic effect on these arterioles. Low nanomolar concentrations (0.1-10 nM) of capsaicin resulted in substantial arteriolar dilation (maximum at 10 nM, 32 ± 13%, n = 12), which was abolished by the removal of endothelium (Fig. 5A) or by NO synthase inhibition with Nω-nitro-l-arginine methyl ester (0.2 mM) (7 ± 5% dilation, n = 8, or 1 ± 2% constriction, n = 4, respectively). In contrast, higher concentrations of capsaicin (0.1-1 μM) elicited a significant vasoconstriction (Fig. 5A; apparent maximum at 1 μM, 66 ± 7%, n = 12), which was not affected by endothelium removal (Fig. 5A; apparent maximum at 1 μM, 68 ± 4%, n = 8 after endothelium removal). We have found that capsaicin-induced arteriolar constriction was transient (Fig. 5B), reaching its maximum at approximately 90 s (maximal constriction: 59 ± 10%, n = 5) and returning to the baseline diameter at the end of the 20-min treatment (3 ± 3% dilation, n = 5).
The role of the endogenous activation of TRPV1 on the determination of arteriolar diameter was also tested. Application of the TRPV1 antagonist capsazepine (10 μM) resulted in a significant vasodilation (28 ± 7%, n = 5) in isolated skeletal muscle arteries.
Based on these functional and immunohistochemical observations, we hypothesized that TRPV1 expression is not restricted to sensory neurons in the vasculature. The vascular expression of TRPV1 in the musculus gracilis arterioles was evaluated by immunohistochemistry. In these arterioles, TRPV1-expressing cells were costained with smooth muscle α-actin (Fig. 6B), demonstrating the expression of TRPV1 in the smooth muscle cells. To further confirm the specificity of the TRPV1 staining, arterioles were costained with different TRPV1 antibodies developed against different TRPV1 epitopes in different species, which resulted in overlapping staining patterns (data not shown). the presence of TRPV1 mRNA in aorta and in cultured A7r5 vascular smooth muscle cells was also confirmed by RT-PCR (Fig. 6A).
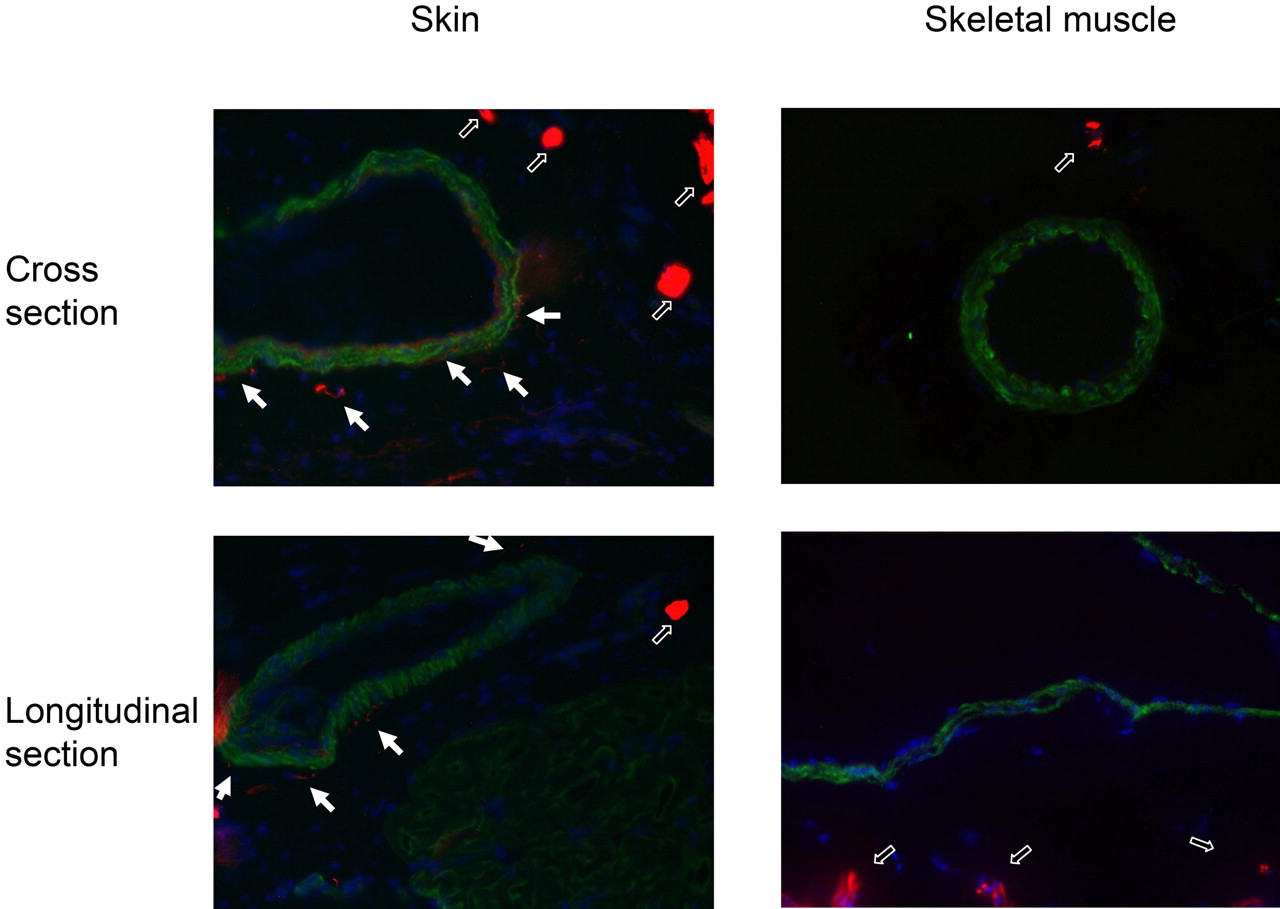
Innervation of arteries in the skin and skeletal muscle of the rat hind limb. The skin (paw) and skeletal muscle (gracilis) of the hind limb were sectioned in a cryostate (thickness, 10 μm) and fixed in acetone. The presence of neuronal elements (innervation) was tested by a neurofilament specific antibody (dilution: 1:100; Sigma) and visualized by a secondary antibody conjugated with Texas red (red on the figures). For the staining of the arteries, a smooth muscle-specific antibody (dilution: 1:20; Novocastra) and a secondary antibody conjugated with Cy-2 (green on the figures) were used. Localization of these elements were visualized in cross and longitudinal sections. Thick nerves in the tissue sections are represented by open arrows, whereas thin neurits innervating the arteries are shown by closed arrows.

Effects of TRPV1 stimulation in isolated arterioles. A, dose-response of capsaicin on isolated pressurized skeletal muscle arterioles (control, n = 12, and endothelium-denuded, n = 8). B, the effect of 20-min continuous application of 1 μM capsaicin. Values are average ± S.E.M.

Expression of TRPV1 in vascular smooth muscle cells. RT-PCR analysis was performed using RNA isolated from aorta and cultured vascular smooth muscle cells (A). TRPV1-specific primers were designed to yield a 149-bp fragment from mRNA (shown by the arrow) and a 596-bp fragment from DNA (not detected). Glyceraldehyde-3-phosphate dehydrogenase was used as control. To investigate the localization of TRPV1, immunohistochemistry was performed in the skeletal muscle (gracilis) tissue sections of the rat (B). TRPV1- (rabbit, 1:500 dilution, green; Calbiochem) and smooth muscle actin (monoclonal, 1:20 dilution, red; Novocastra)-specific staining was visualized by a fluorescent microscope. Slides were also processed in mounting media containing 4,6-diamidino-2-phenylindole for staining of nuclei (blue). The artery used in the in vitro functional studies is identified (L, lumen; A, artery).
Discussion
The main findings of the present study are that 1) in the hind limb, TRPV1 stimulation resulted in an increase of vascular resistance and reduction of tissue perfusion (Fig. 1), independently of the innervation (Fig. 2 and 3); 2) in the isolated skeletal muscle arterioles, capsaicin elicited vasoconstriction, which was not affected by endothelium removal (Fig. 5); 3) TRPV1 mRNA is detectable in both aorta and cultured smooth muscle cells (Fig. 6); and 4) TRPV1 is expressed in vascular smooth muscle cells of the skeletal muscle arterioles (Fig. 6). These data suggest functional expression of TRPV1 in the vascular smooth muscle cells of the skeletal muscle arterioles.
Our present findings also indicate that TRPV1 stimulation results in diverse vascular effects in arterial beds of the rat hind limb. We propose that TRPV1 regulates arteriolar diameter primarily by two mechanisms (Fig. 7). In the case of skin, TRPV1 activation resulted in a neuronal vasodilation (Fig. 1 and 3), probably mediated by a sequence of events such as the activation of sensory neurons, the subsequent local release of sensory neurotransmitters (like CGRP or SP), stimulation of endothelial cells by these neurotransmitters, activation of endothelial NO synthesis, and NO-mediated relaxation of smooth muscle cells, as proposed by Zygmunt et al. (1999). In contrast, TRPV1 stimulation by capsaicin had biphasic effects in the isolated skeletal muscle resistance arterioles (vasodilation at lower concentrations and vasoconstriction at higher concentrations; Fig. 5), suggesting a dual regulation of vascular tone. The dilatative effects of capsaicin were endothelium-dependent (Fig. 5) and nitric oxide-mediated. In addition, inhibition of TRPV1 in isolated, pressurized skeletal muscle arteries resulted in a vasodilation, indicating a physiological role of TRPV1 in the regulation of vascular diameter. It should also be noted that capsaicin evoked opposite effects in different vascular beds under in vivo conditions (Fig. 1A). The increase of pressure of the isovolumetric (3 ml/min) perfusion in the hind limb suggests higher local resistance in the vasculature of the hind limb (at least partly as a result of skeletal muscle arteriolar constriction, Fig. 5), whereas the simultaneous decrease in systemic blood pressure indicates a somewhat higher overall efficiency of vasodilatative receptors (probably mediated by mesenteric, dural, skin, pulmonary, or coronary arteries) in the whole vasculature.

Proposed mechanism of tissue-specific regulation of vascular diameter by TRPV1. Our data support the well known sensory neuronal dilation in the skin arteries involving the following events: 1) activation of sensory neuronal TRPV1; 2) elevation of intracellular Ca2+ in the neuronal terminals; 3) release of sensory neurotransmitters, including CGRP and SP; 4) activation of endothelial receptors of these neurotransmitters; 5) increase of endothelial NO synthesis; and 6) NO diffusion and smooth muscle relaxation. The same mechanism was found to be responsible for capsaicin-mediated dilation in skeletal muscle arteries. In contrast, functional expression of TRPV1 was also identified in vascular smooth muscle cells of gracilis artery. The activation of these smooth muscle-located receptors led to vasoconstriction. We propose that the balance of activities of sensory neuronal and smooth muscle-located TRPV1-mediated pathways determines the vasoactive effects of TRPV1 stimulation.
The simplest explanation of the findings is that TRPV1 is functionally expressed in vascular smooth muscle cells. Although findings based on immunohistochemical data in general should be interpreted with caution, this proposal is also supported by RT-PCR results and the vasoconstrictive effect of TRPV1 stimulation in intact or endothelial denuded isolated skeletal muscle arteries. According to these data, we hypothesize that activation of TRPV1 in skeletal muscle arteries occurs both in sensory neurons and in vascular smooth muscle cells, leading to Ca2+ influx into both cell types. The elevated intracellular Ca2+ concentration in the smooth muscle directly results in vasoconstriction, whereas in the sensory nerves, it triggers neurotransmitter release and concomitant endothelial-dependent vasodilation (Fig. 7).
Capsaicin-evoked in vivo vasoconstriction of various arterial beds was discovered decades ago (Molnár and Gyorgy, 1967; Toda et al., 1972; Donnerer and Lembeck, 1982; Duckles, 1986; Edvinsson et al., 1990). In these initial and follow-up studies, TRPV1-mediated vasoconstriction was found in dog mesenteric (Toda et al., 1972; Pórszász et al., 2002), renal, and carotid artery (Toda et al., 1972); in cat middle cerebral (Duckles, 1986; Edvinsson et al., 1990), pial (Edvinsson et al., 1990), and pulmonary (Molnár and Gyorgy, 1967) arteries; in rat heart (Szolcsányi et al., 2001), small mesenteric (Scotland et al., 2004), dural (Dux et al., 2003), and skeletal muscle arteries (Lizanecz et al., 2006); and in mouse knee joint (Keeble and Brain, 2006). Multiple mechanisms leading to TRPV1-mediated vasoconstriction were suggested, including endothelin-1 (Szolcsányi et al., 2001) or SP (Scotland et al., 2004) release from sensory neurons, and yet uncharacterized smooth muscle-mediated effects were observed (Pórszász et al., 2002; Dux et al., 2003; Keeble and Brain, 2006). In addition to these possibilities, our data provide evidence for TRPV1 expression in vascular smooth muscle cells, suggesting a direct link between TRPV1 activation and smooth muscle contraction. Nevertheless, these data indicate that vasoconstrictive effects of TRPV1 stimulation are not restricted to a specific blood vessel or to a single species.
It is interesting that in some of these cases, like in the case of rat mesenteric arteries, both vasoconstriction (Scotland et al., 2004) and vasodilation (Ralevic et al., 2000) were observed upon capsaicin stimulation. It suggests that there are two pools of TRPV1 in these systems, but one of the receptor types is down-regulated under specific circumstances, and the physiological effect of capsaicin stimulation is dominated by the active receptor population. In accordance with this idea, dose-dependent biphasic effects were also noted in some studies: low dose capsaicin evoked dilation, whereas higher concentrations resulted in constriction (Edvinsson et al., 1990; Dux et al., 2003), similar to our findings.
Several mechanisms have been suggested to regulate TRPV1 activity, besides to the expressional regulation. These include protein kinase C (Bhave et al., 2003) or protein kinase A (Bhave et al., 2002)-mediated phosphorylation, calcineurin-mediated dephosphorylation (Docherty et al., 1996), interaction with calmodulin (Numazaki et al., 2003) or with phosphoinositides (Liu et al., 2005; Lukacs et al., 2007), besides others. As a matter of TRPV1 mediating skeletal muscle vasoconstriction, phosphorylation seems to be the most likely candidate (Lizanecz et al., 2006).
Some of the findings of this study suggest pharmacological differences in the TRPV1 pools mediating constriction and dilation, namely 1) higher sensitivity/effectivity of dilatative responses (dilation in the case of low capsaicin concentrations), with a profound constrictive responses at maximal stimulation; and 2) vasodilation evoked by short-term inhibition of TRPV1. In addition, earlier data suggest that the TRPV1 receptors mediating vasodilation can be easily desensitized by neonatal capsaicin treatments, whereas the capsaicin response of receptors mediating vasoconstriction remains intact or augmented (Donnerer and Lembeck, 1982). As a therapeutical consequence of these observations, it seems to be possible to design TRPV1 ligands preferably acting on receptors mediating constrictive or dilatative responses. Although there is no shortage of drug candidates regulating TRPV1 activity (Szallasi et al., 2007), their development was concentrated on their effects on sensory neuronal functions (mostly pain). One of the examples to emphasize the feasibility of such drug development is that it was possible to design an antagonist selective to the plasma membrane located TRPV1 over to the intracellular membrane-located receptors (Tóth et al., 2004). The drugs selective to receptors mediating dilation or constriction may be useful to regulate blood distribution in various pathophysiological conditions associated with ischemia. For example, sensory neuronal TRPV1 was found to be activated upon myocardial ischemia (Zahner et al., 2003; Pan and Chen, 2004) and beneficial in postischemic recovery (Wang and Wang, 2005), suggesting that selective activation of sensory neuronal TRPV1 may be beneficial in myocardial infarction.
Taken together, we report here that TRPV1 (a nonspecific Ca2+ channel) is expressed in smooth muscle cells, and its activation leads to vasoconstriction in skeletal muscle resistance arterioles. We propose that TRPV1 has a potential physiological/pharmacological role in the regulation of arteriolar tone in skeletal muscle (apparently in the range of 40% dilation to 60% constriction), which represents a promising new therapeutic strategy to control tissue-specific blood distribution.
Footnotes
- Received November 8, 2007.
- Accepted February 5, 2008.
The study was supported by the Hungarian Academy of Sciences OTKA (F048873 and K68077 to A.T., T046244 to R.P., and F-048837 to Z.B.) and by the Hungarian Ministry of Health (ETT 430/2006 to A.T., ETT 493/2006 to R.P., ETT-454-2006 to I.E.) grants. Z.B., Z.P., R.P., and A.T. were supported by Bolyai Fellowship of the Hungarian Academy of Sciences.
R.P. and A.T. served as cosenior authors.
ABBREVIATIONS: TRPV1, transient receptor potential type V1 channel (vanilloid receptor 1); CGRP, calcitonin gene-related peptide; SP, substance P; RT-PCR, reverse transcriptase-polymerase chain reaction; bp, base pair(s); RTX, resiniferatoxin; AU, arbitrary units.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}