


Abstract
A decreased central dopaminergic and/or noradrenergic transmission is believed to be involved in the pathophysiology of depression. It is known that dopamine (DA) neurons in the ventral tegmental area (VTA) and norepinephrine (NE) neurons in the locus ceruleus (LC) are autoregulated by somatodendritic D2-like and α2-adrenoceptors, respectively. Complementing these autoreceptor-mediated inhibitory feedbacks, anatomical and functional studies have established a role for noradrenergic inputs in regulating dopaminergic activity, and reciprocally. In the present study, a microiontophoretic approach was used to characterize the postsynaptic catecholamine heteroreceptors involved in such regulations. In the VTA, the application of DA and NE significantly reduced the firing activity of DA neurons. In addition to a role for D2-like receptors in the inhibitory effects of both catecholamines, it was demonstrated that the α2-adrenoceptor antagonist idazoxan dampened the DA- and NE-induced attenuations of DA neuronal activity, indicating that both of these receptors are involved in the responsiveness of VTA DA neurons to catecholamines. In the LC, the effectiveness of iontophoretically applied NE and DA to suppress NE neuronal firing was blocked by idazoxan but not by the D2-like receptor antagonist raclopride, which suggested that only α2-adrenoceptors were involved. In the dorsal hippocampus, a forebrain region having a sparse dopaminergic innervation but receiving a dense noradrenergic input, the suppressant effects of DA and NE on pyramidal neurons were attenuated by idazoxan but not by raclopride. The suppressant effect of DA was prolonged by administration of the selective NE reuptake inhibitor desipramine and, to lesser extent, of the selective DA reuptake inhibitor 1-(2-[bis(4-fluorophenyl)methoxy]ethyl)-4-(3-phenylpropyl)-piperazine (GBR12909), suggesting that both the NE and DA transporters were involved in DA uptake in the hippocampus. These findings might help in designing new antidepressant strategies aimed at enhancing DA and NE neurotransmission.
The catecholamines neurotransmitters dopamine (DA) and norepinephrine (NE) are believed to be involved in psychiatric disorders, and a better knowledge of the reciprocal interactions between these two systems should improve our understanding of the pathophysiology and treatment of mood disorders. Anatomical evidence indicates that noradrenergic neurons of the locus ceruleus (LC) send projections to the ventral tegmental area (VTA) in the vicinity of DA neuron cell bodies (Simon et al., 1979). Several subtypes of α-adrenergic receptors have been identified in the VTA (Lee et al., 1998), raising the possibility that NE inputs play a role in modulating DA neuronal activity. Consistent with this assumption, it was recently demonstrated that a selective lesion of LC NE neurons increases the mean firing activity of DA neurons by 70% and their burst activity by almost 50%, thus revealing a net inhibitory effect of NE in the VTA (Guiard et al., 2008). In contrast, it had been reported that the systemic administration of the selective NE reuptake inhibitor reboxetine enhanced the burst firing activity of DA neurons in the VTA, but without any apparent effect on their mean firing rate (Linnér et al., 2001). In earlier attempts at characterizing the mechanism of action of NE in the VTA, it has also been shown that the systemic administration of the α1-adrenoceptor antagonist prazosin dose-dependently decreased the burst firing of DA neurons (Grenhoff and Svensson, 1993), apparently supporting an excitatory action of NE in the VTA. However, the selective α2-adrenoceptor antagonist idazoxan increased both the firing rate and bursting activity of VTA DA neurons (Grenhoff and Svensson, 1993), thereby suggesting a complex regulation of DA neuronal activity by NE inputs. As for the local application of NE in the VTA, it usually reduced the spontaneous firing of DA neurons (Aghajanian and Bunney, 1977a,b; White and Wang, 1984a,b), suggesting that a postsynaptic inhibitory element sensitive to NE does exist in the VTA. However, whether NE exerts its effects in the VTA via α-adrenergic and/or dopaminergic receptors remained an unresolved issue.
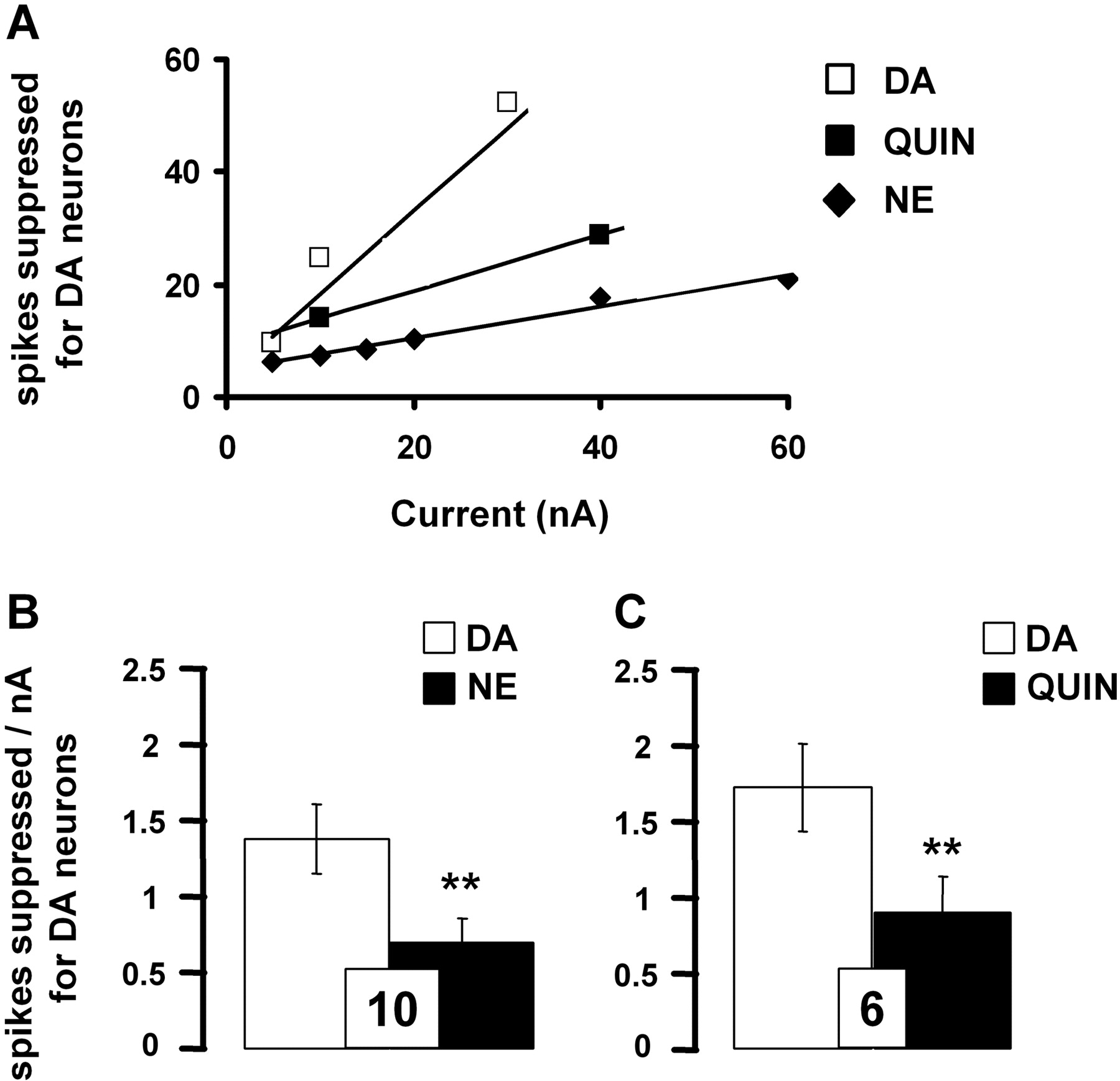
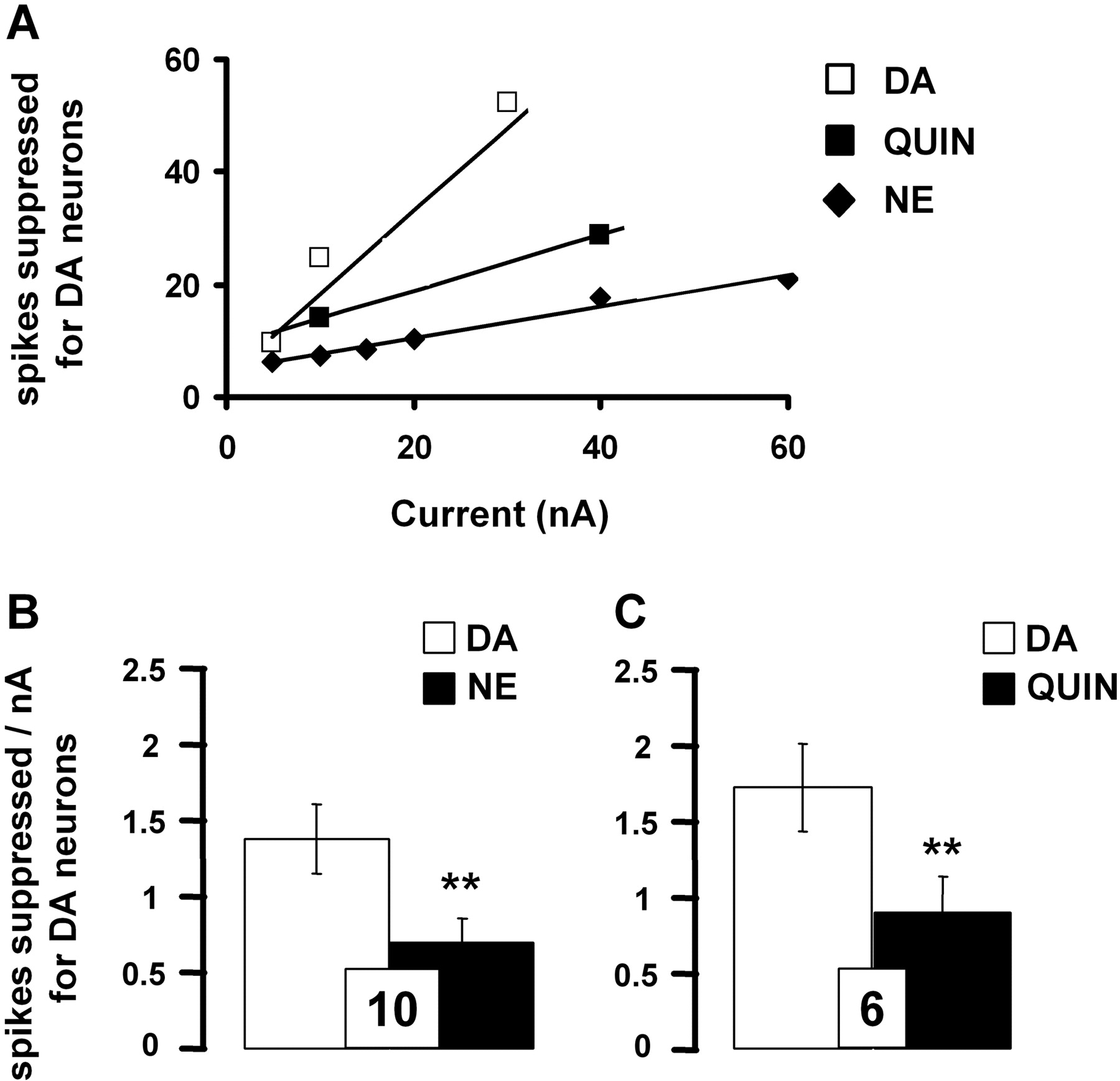
Comparative effects of iontophoretically applied DA, NE, and quinpirole on the firing rate of VTA DA neurons. A, data are expressed as the means of the number of spikes suppressed for DA neurons by DA (0.1 M), NE (0.01 M), and quinpirole (Quin; 0.05 M). B and C, responsiveness of VTA DA neurons to iontophoretic applications of DA, NE, and quinpirole. Data are expressed as mean ± S.E.M. of the number of spikes suppressed by nanoamperes for DA neurons. The number of neurons tested is indicated in each histogram. **, P < 0.01, significantly different from the effect of DA alone by two-tailed Student's t test.
On the other hand, DA can influence the neuronal activity of LC NE neurons. Anatomical data support this possibility because descending pathways from the VTA innervate the LC (Ornstein et al., 1987). Moreover, in rodents, although only a small percentage of its VTA-derived afferents seem to be dopaminergic (Swanson, 1982), DA receptors mainly of the D2-like subtype have been identified in the LC (Yokoyama et al., 1994). Lesion experiments have shed some light on the nature of the effects of DA in the LC. The selective lesion of VTA DA neurons increased by 33% the mean firing rate of LC NE neurons discharging in a single spike mode and by almost 60% that of LC neurons displaying both single spike and busting activities (Guiard et al., 2008), revealing an inhibitory influence of DA inputs. In agreement with these findings, pharmacological studies have indicated that the systemic administration of the nonselective DA receptor agonist (+)-3-PP dose-dependently reduced the firing rate of LC NE neurons, whereas this effect was partially antagonized by the α2-adrenoceptor antagonist yohimbine but not the D2-like receptor antagonist haloperidol (Elam et al., 1986). These results stand in contrast with data showing that the systemic administration of haloperidol enhances the firing rate of LC NE neurons, indicating the existence of a tonically active dopamine input that modulates the firing pattern of LC NE neurons (Piercey et al., 1994; Nilsson et al., 2005). The iontophoretic application of DA in the LC has been shown to inhibit the electrical activity of NE neurons (Cedarbaum and Aghajanian, 1977; Elam et al., 1986). However, the lack or weak effect of the systemic administration of D2/D3 receptor agonists, such as pramipexole or apomorphine, on the firing rate of LC NE neurons (Cedarbaum and Aghajanian, 1977; Chernoloz et al., 2008) has not allowed the identification of the postsynaptic receptor(s) that mediate DA effects in the LC.
In rats, the dorsal hippocampus receives a dense noradrenergic and sparse dopaminergic innervation arising from the LC (Jones and Moore, 1977) and the VTA (Swanson and Hartman, 1975; Scatton et al., 1980), respectively. In addition, because a significant decrease in hippocampal DA levels had been reported when most of the noradrenergic neurons were lesioned (Bischoff et al., 1979), it is possible that in this brain region, DA not only subserves a neurotransmitter role in dopaminergic neurons but is also present as the precursor of NE in noradrenergic neurons. Indeed, it had been proposed that approximately 40% of hippocampal DA is confined in this population of neurons (Bischoff et al., 1979). Radioligand binding studies have revealed the presence of α1-, α2-, β1-, and β2-adrenoceptors (Crutcher and Davis, 1980) and in D2-like receptors (Bischoff et al., 1986; Bruinink and Bischoff, 1993) in the hippocampus, suggesting a role of both catecholamines in the modulation of CA3 pyramidal neurons. Therefore, in vivo electrophysiological evidence demonstrated that NE generally decreased pyramidal neuronal activity, but both excitation and biphasic responses have also been observed. Whereas the inhibitory effects of iontophoretic application of NE on rat dorsal hippocampus result from the activation of α2-adrenoceptors (Curet and de Montigny, 1988a,b), it had been shown that the excitatory effects are mediated by β-adrenoceptors (Mueller et al., 1982; Curet and de Montigny, 1988). The existence of DA-sensitive receptor sites in the hippocampus was first suggested from observations that the local application of DA decreases the firing rate of CA3 pyramidal neurons (Segal et al., 1973; Benardo and Prince, 1982). Despite these results, little is known about the role of DA in the hippocampus and the nature of the receptors mediating its electrophysiological effects.
The present study was therefore aimed at characterizing the effects of microiontophoretically applied NE and DA in the VTA, LC, and the dorsal hippocampus. This technique has the advantage of limiting the zone of possible drug-receptor interactions to the discrete area of application and, as a result, to allow the identification of postsynaptic receptor(s) mediating the response to catecholamines.
Materials and Methods
Animals. Male Sprague-Dawley rats (Charles River Canada, Montreal, QC, Canada) weighing 250 to 300 g were used in all experiments. They were housed individually and kept under standard laboratory conditions (12-h light/dark cycle with free access to food and water). All animals were handled according to the guidelines of the Canadian Council on Animal Care, and the protocols were approved by the local Animal Care Committee (Institute of Mental Health Research, Ottawa, ON, Canada).
In Vivo Microiontophoresis. Rats were anesthetized with chloral hydrate (400 mg/kg i.p.) and placed into a stereotaxic frame. The extracellular recordings of the DA, NE, and pyramidal neurons in the VTA, LC, and CA3 region of the hippocampus, respectively, were carried out using multibarreled glass micropipettes (ASI Instruments, Warren, MI). Five-barreled glass micropipettes preloaded with fiberglass strands to promote capillary filling were pulled in the conventional manner. The central barrel used for extracellular unitary recording was filled with 2 M NaCl solution. One side barrel, filled with 2 M NaCl solution, was used for automatic current balancing. The other side barrels were filled with three of the following drug solutions: dopamine (DA-HCl, 0.1 M in 0.2 M NaCl, pH 4), norepinephrine (l-NE-HCl, 0.01 M in 0.2 M NaCl, pH 4), quinpirole-HCl (0.05 M in 0.2 M NaCl, pH 4), raclopride-tartrate (0.025 M in 0.2 M NaCl, pH 4), idazoxan-HCl (0.05 M in 0.2 M NaCl, pH 4), and quisqualate (0.0015 M in 0.2 M NaCl, pH 8). All drugs were ejected as cations and retained with currents of -10 to -8 nA, except for quisqualate, which was ejected as an anion and retained with a current of +5 nA. The impedances of the central barrel were 2 to 5 MΩ in the LC and hippocampus and 6 to 8 MΩ in the VTA. The impedances of the balance barrel and side barrels were 20 to 30 and 50 to 100 MΩ, respectively. Haloperidol (200 μg/kg), desipramine (2, 4, and 6 mg/kg), and GBR12909 (7.5 mg/kg i.v.) were administered intravenously. All drugs were purchased from Sigma (St. Louis, MO).

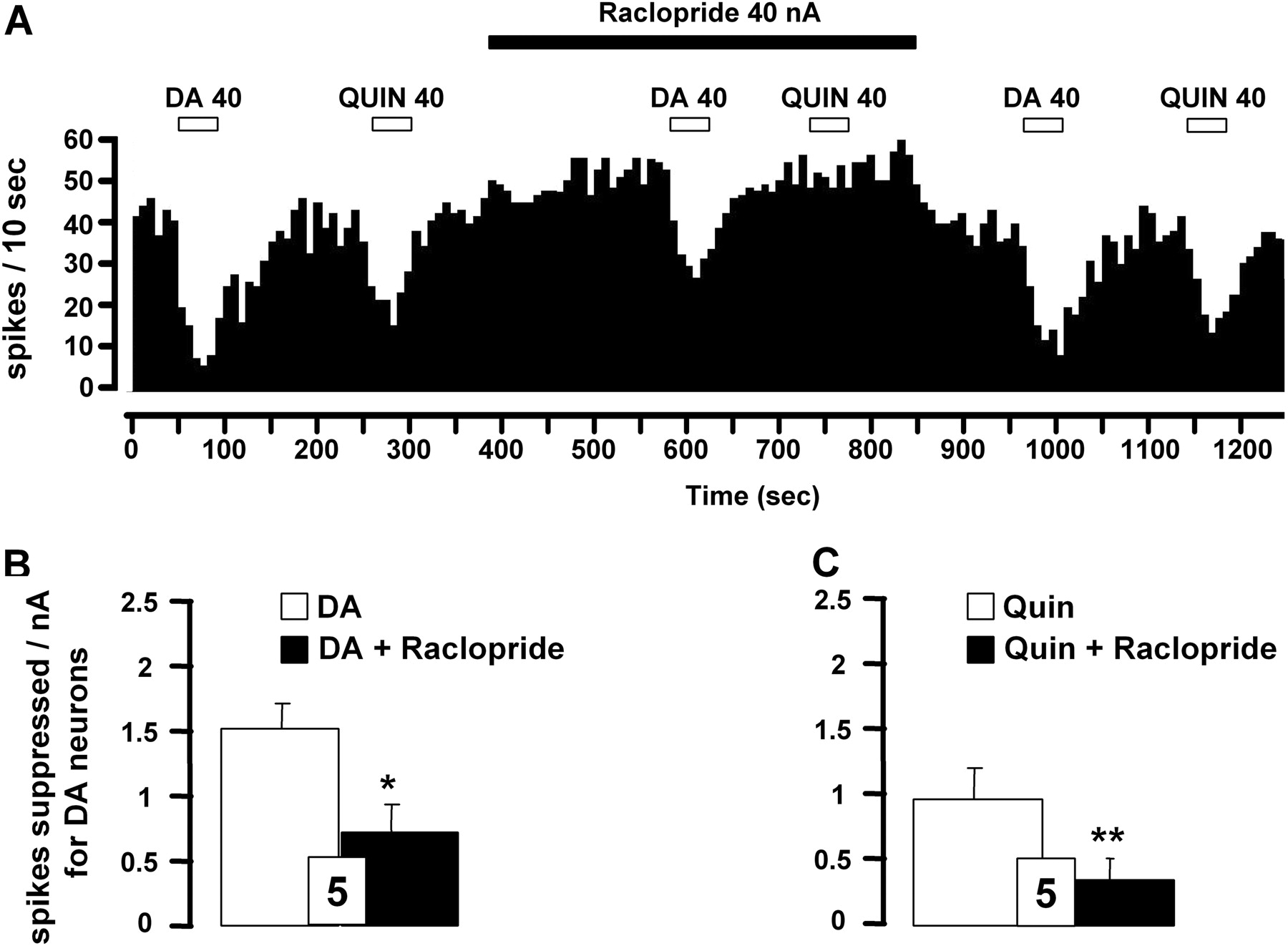
Effect of D2-like receptor antagonist raclopride on iontophoretically applied DA- or quinpirole-induced inhibition of VTA DA neurons. A, integrated firing rate histograms illustrating the effects of raclopride on DA- or quinpirole-induced decrease in VTA DA neuronal activity. Horizontal bars indicate the duration of iontophoretic ejection and current values in nanoamperes. B and C, responsiveness of VTA DA neurons to iontophoretic applications of DA (B) or quinpirole (C) in the presence or absence of raclopride (0.025 M). Data are expressed as means ± S.E.M. of the number of spikes suppressed by nanoamperes for DA neurons. The number of neurons tested is indicated in each histogram. *, P < 0.05, and **, P < 0.01, significantly different from the effect of DA alone by two-tailed Student's t test.
Recording of VTA DA Neurons. The five-barreled glass micropipettes were positioned using the following coordinates (from bregma): AP, -6 to -5.4 mm; L, 1 to 0.6 mm; and V, 7 to 9 mm. The presumed DA neurons were identified according to the well established electrophysiological properties in vivo: a typical triphasic action potential with a marked negative deflection; a characteristic long duration (> 2.5 ms) often with an inflection or “notch” on the rising phase; and a slow spontaneous firing rate (2-10 Hz) with an irregular single spiking pattern and slow bursting activity (characterized by spike-amplitude decrement).
Recording of LC NE Neurons. The five-barreled glass micropipettes were positioned using the following coordinates (from lambda): AP, -1.0 to -1.2 mm; L, 1.0 to 1.3 mm; and V, 5 to 7 mm. Spontaneously active NE neurons were identified using the following criteria: regular firing rate (0.5-5.0 Hz) and positive action potential of long duration (0.8-1.2 ms) exhibiting a brisk excitatory response to a nociceptive pinch of the contralateral hind paw. The mesencephalic fifth nucleus neurons were first located by a response to lower jaw depression, and then the electrode was lowered medially to record LC NE neurons.
Recording of Pyramidal Neurons in the CA3 Region of Dorsal Hippocampus. The five-barreled glass micropipettes were positioned using the following coordinates (from bregma): AP, 3.8 to 4.5 mm; L, 4 to 4.2 mm; and V, 3 to 4.5 mm. Pyramidal neurons stimulated with quisqualate were identified by their high amplitude (0.5-1.2 mV), high frequency (13-15 Hz) and long duration (0.6-1.0 ms) action potential, and their characteristic “complex spike” discharge.
Assessment of Neuronal Responsiveness. In each brain region (i.e., the VTA, LC, and dorsal hippocampus), a current-response curve was obtained by determining for each current the number of spikes suppressed during the 50-s drug ejection period. Two others parameters were used to assess neuronal responsiveness to microiontophoretic application: 1) the number of spikes suppressed per nanoampere, obtained by dividing the number of spikes suppressed from the beginning of the ejection period by the current used (in nanoamperes); and 2) the RT50 value (in seconds), which represents the time required for the firing activity to recover by 50% from the cessation of the microiontophoretic application. In the present study, the RT50 value was used to provide an index of the capacity of NE terminals in the dorsal hippocampus to remove NE or DA from the synaptic cleft in presence or not of intravenous cumulative doses of the selective NE reuptake inhibitor desipramine (from 2 to 6 mg/kg by adding 2 mg/kg after each injection) or of the selective DA reuptake inhibitor GBR12909 (7.5 mg/kg). The doses of desipramine and GBR12909 were chosen on the basis of their capacity to significantly block the NE and DA transporters, respectively (Curet and de Montigny, 1988; Einhorn et al., 1988).
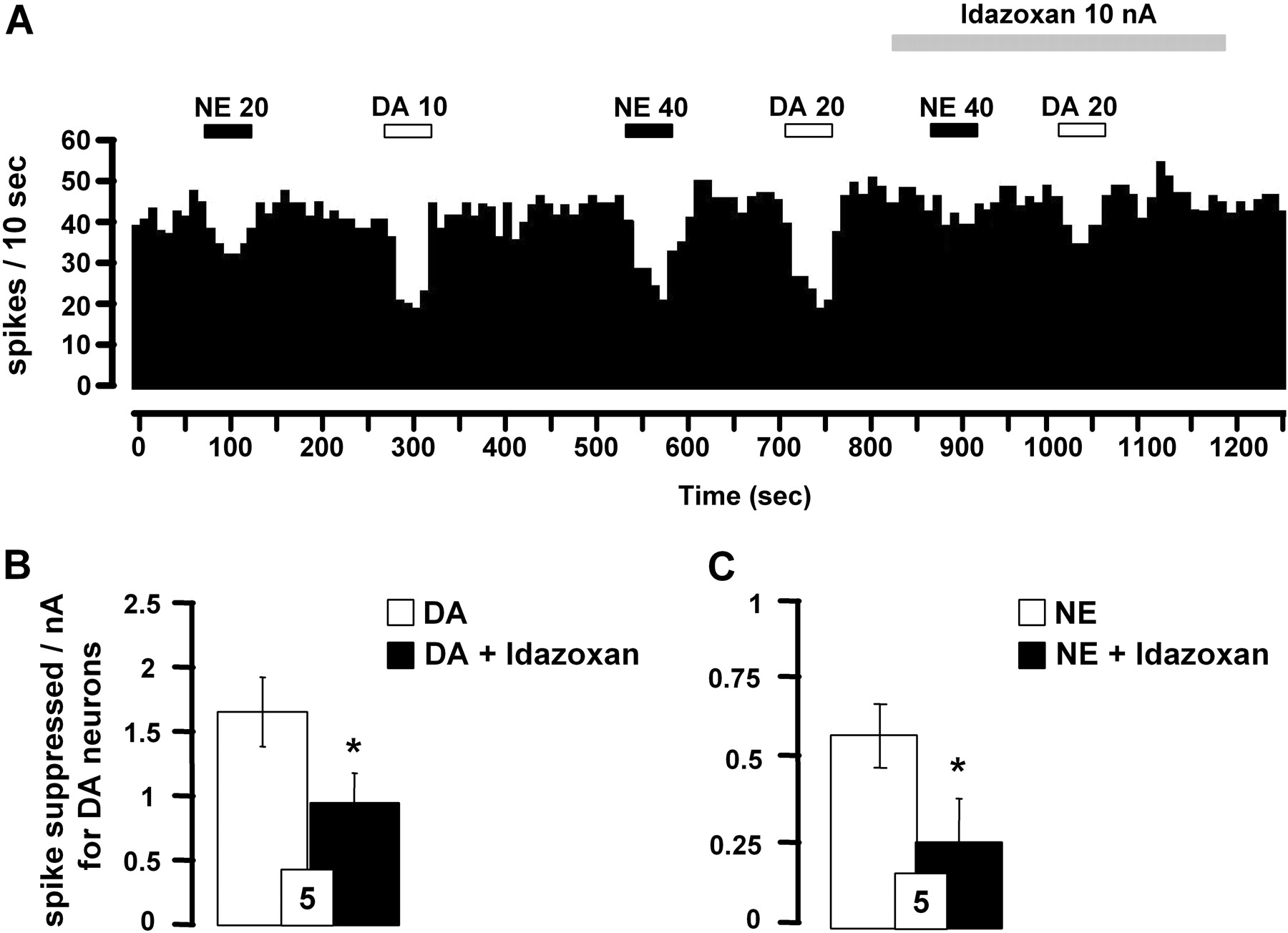
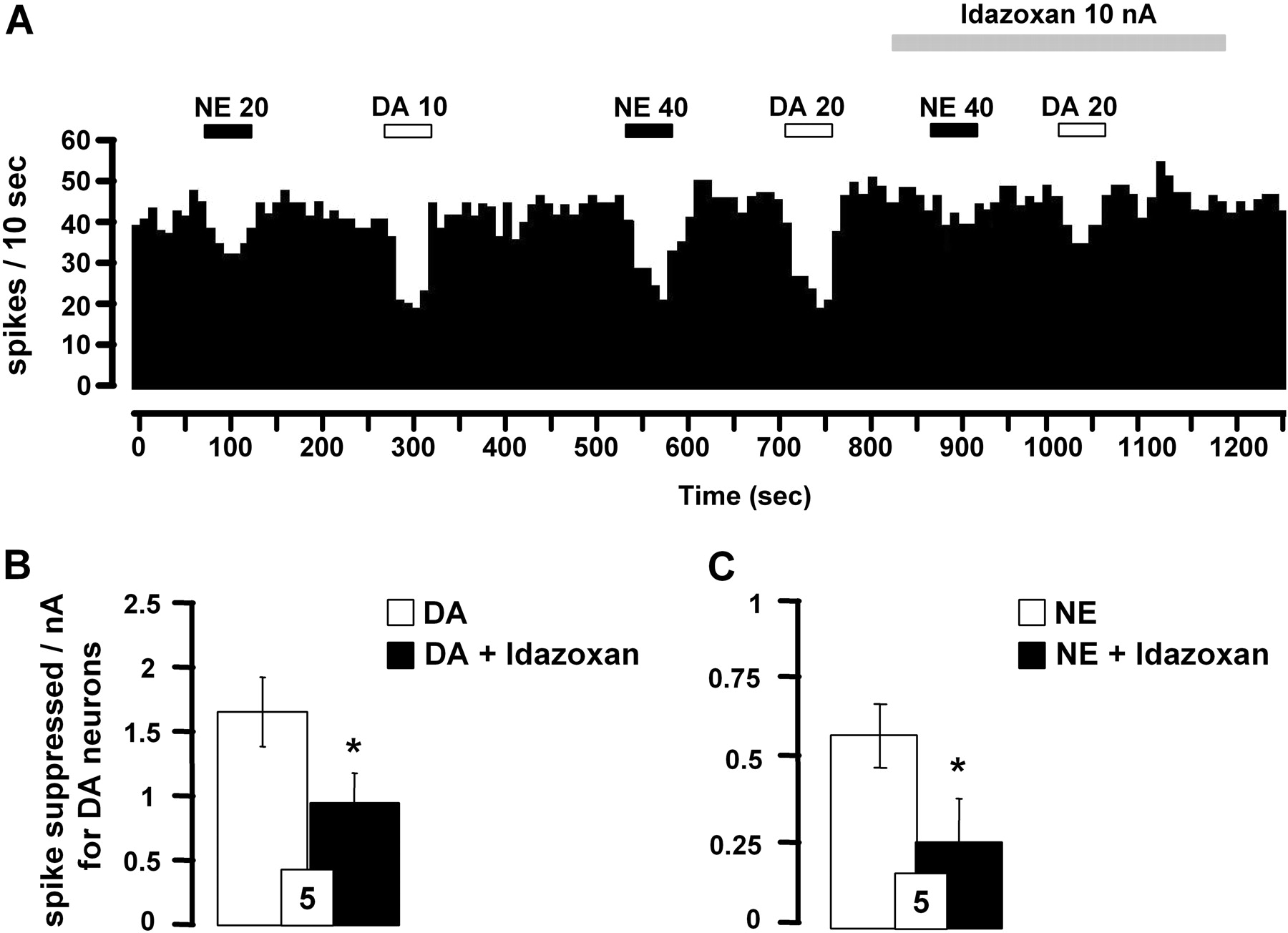
Effect of α2-adrenoceptor antagonist idazoxan on iontophoretically applied DA- and NE-induced inhibition of VTA DA neurons. A, integrated firing rate histograms illustrating the effects of idazoxan on DA- or NE-induced decrease in VTA DA neuronal activity. Horizontal bars indicate the duration of iontophoretic ejection and current values in nanoamperes. B and C, responsiveness of VTA DA neurons to iontophoretic applications of DA (B) or NE (C) alone and in the presence of idazoxan (0.05 M). Data are expressed as means ± S.E.M. of the number of spikes suppressed by nanoamperes for DA neurons. The number of neurons tested is indicated in each histogram. *, P < 0.05, significantly different from DA or NE effects alone by paired two-tailed Student's t test.
Statistical Analysis. Electrophysiological data were expressed as means ± S.E.M of number of spike suppressed per nanoampere, spontaneous firing rate, or RT50 values. The paired Student's t test was used to assess the statistical significance of the variations of a parameter measured from the same neurons under two conditions. When more than two groups were compared (e.g., RT50 values after the various doses of desipramine), a one-way analysis of variance with repeated measures and treatment as main factor, followed by a Fisher's protected least-significance difference post hoc test, was used.
Results
Pharmacological Characterization of the Effects of Iontophoretically Applied DA and NE on DA Neurons in the VTA. In the VTA, DA neurons displayed spontaneous electrical activity in a range similar to that described previously (i.e., 4.2 ± 0.4 Hz, n = 26). DA neurons typically were inhibited in a current-dependent manner to iontophoretically applied DA (5-30 nA) and NE (5-60 nA; Fig. 1A). However, as found by other investigators (White and Wang, 1984a), even at high ejection currents, a complete suppression of VTA-DA neuronal activity was usually not observed.
The iontophoretic application of the D2-like receptor agonist quinpirole also reduced the firing activity of all VTA DA neurons recorded in a current-dependent manner (10-40 nA; Fig. 1A), consistent with the well documented inhibitory role of D2-like receptor subtype on this population of neurons. It is noteworthy that the inhibition of the firing rate of VTA DA neurons was more pronounced after the iontophoretic application of DA than NE or quinpirole (Fig. 1, B and C). Complementing the latter results, it was observed that the iontophoretic application of the D2-like receptor antagonist raclopride (40 nA) blocked the suppressant effects of both DA and quinpirole on VTA DA neuronal activity (Fig. 2, A-C). For some neurons, the systemic administration of the D2-like receptor antagonist haloperidol also prevented the suppressant effect of DA and NE (n = 3). It is noteworthy that raclopride applied by microiontophoresis increased the spontaneous firing rate of VTA DA neurons (4.7 ± 0.7 versus 6.3 ± 0.6 Hz before and after its ejection, respectively; n = 5, P < 0.05). It thus seems that the inhibitory effect of DA was mediated by D2-like receptors. However, the iontophoretic application of raclopride did not allow the complete blockade of the electrophysiological effects of DA, raising the possibility that other(s) receptor(s) might be involved in the inhibitory action of both pharmacological agents.
The iontophoretic application of the α2-adrenoceptor antagonist idazoxan was used to test whether the effects of DA could be mediated, at least in part, by a nondopaminergic mechanism. The suppressant effect of DA was partially blocked by idazoxan (Fig. 3, A and B), whereas this drug per se did not affect the spontaneous firing of VTA DA neurons (3.9 ± 0.7 versus 4.1 ± 0.8 Hz, before and after its ejection; n = 5, P > 0.05). The suppressant effect of NE was also blocked by idazoxan in the VTA (Fig. 3, A and C).

Effect of D2-like receptor antagonist raclopride on iontophoretically applied NE- or DA-induced inhibition of LC NE neurons. A, comparative effects of microiontophoretically applied DA, NE, and quinpirole on the firing rates of LC NE neurons. Data are expressed as means ± S.E.M. of the number of spikes suppressed for NE neurons by DA (0.1 M), NE (0.01 M), and quinpirole (0.05 M). B, integrated firing rate histograms illustrating the effects of raclopride on DA- or NE-induced decrease in LC NE neuronal activity. Horizontal bars indicate the duration of iontophoretic ejection and current values in nanoamperes. C and D, responsiveness of LC NE neurons to iontophoretic applications of DA (C) or NE (D) alone and in the presence of raclopride (0.025 M). Data are expressed as mean ± S.E.M. of the number of spikes suppressed by nanoamperes for NE neurons. The number of neurons tested is indicated in each histogram.
Pharmacological Characterization of the Effects of Iontophoretically Applied DA on Noradrenergic Neurons in the LC. In the LC, the mean spontaneous electrical activity of NE neurons was 2.1 ± 0.2 Hz (n = 18). These neurons were inhibited in a current-dependent manner by iontophoretically applied NE and DA (10-15 nA), whereas the D2-like receptor agonist quinpirole produced a relatively weak and current-independent inhibition of LC NE neuronal activity (10-20 nA; Fig. 4A). The latter results suggest that D2-like receptors are not involved in the regulation of LC NE neurons. This inference was supported by the observation that the iontophoretic application of raclopride failed to significantly attenuate the suppressant effects of DA on LC NE neuronal activity, as well as that of NE (Fig. 4, B-D). It is noteworthy that raclopride alone did not modify the mean spontaneous firing activity of NE neurons (2.1 ± 0.5 versus 1.8 ± 0.4 Hz, before and after its application, respectively; n = 6, P > 0.05).

Effect of α2-adrenoceptor antagonist idazoxan on iontophoretically applied DA- or NE-induced inhibition of LC NE neurons. A, integrated firing rate histograms illustrating the effects of raclopride on NE- or DA-induced decrease in LC NE neuronal activity. Horizontal bars indicate the duration of iontophoretic ejection and current values in nanoamperes. B and C, responsiveness of LC NE neurons to iontophoretic applications of NE (B) or DA (C) in the presence or not of idazoxan (0.05 M). Data are expressed as the mean ± S.E.M. of the number of spikes suppressed by nanoamperes for NE neurons. The number of neurons tested is indicated in each histogram. ***, P < 0.001, significantly different from NE or DA alone by paired two-tailed Student's t test.
In agreement with the presence of α2-adrenergic autoreceptors in the LC, the iontophoretic application of idazoxan blocked the suppressant effect of NE on NE neuronal activity (Fig. 5, A and B). The fact that similar results were obtained with DA suggests that DA may also activate α2-adrenergic autoreceptors to exert an inhibitory effect on LC NE neurons (Fig. 5, A and C). Despite its blocking activity on both NE and DA, idazoxan did not affect by itself the spontaneous firing of NE neurons (2.1 ± 0.3 versus 2.3 ± 0.4 Hz, before and after its local application, respectively; n = 7, P > 0.05).
Pharmacological Characterization of the Effects of Iontophoretically Applied NE and DA on Pyramidal Neurons of the CA3 Region of the Dorsal Hippocampus. The CA3 region of the dorsal hippocampus was chosen to further establish the possibility that DA could act via α-adrenergic receptors because of it has sparse DA projections. The firing activity of dorsal hippocampus pyramidal CA3 neurons was activated by quisqualate to 14.3 ± 0.7 Hz (n = 29). All CA3 neurons responded to a marked and current-dependent inhibition to iontophoretically applied DA (5-80 nA) and NE (5-30 nA), whereas the D2-like receptor agonist quinpirole (5-80 nA) had no effect (Fig. 6A). The suppressant effect of DA on these pyramidal CA3 neurons was not blocked by the iontophoretic application of raclopride (Fig. 6, B and D) or by the systemic injection administration of haloperidol (Figs. 6E) but was significantly attenuated by idazoxan (Fig. 7, A and B).
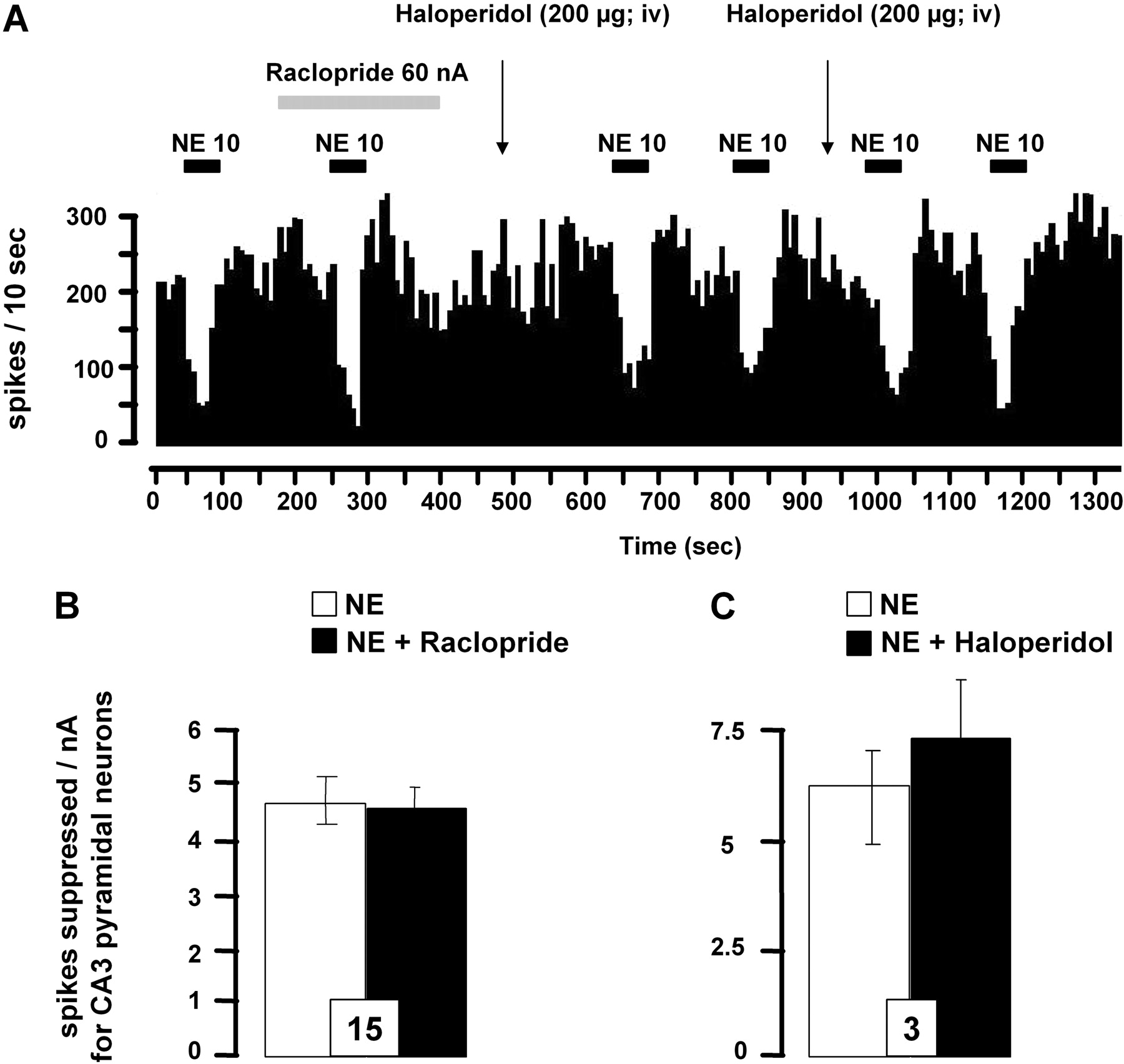
As demonstrated previously, NE suppressed the firing activity of CA3 pyramidal neurons (Curet and de Montigny, 1988). Although previous studies showed that the inhibitory action of NE was mediated by α2-adrenoceptors, the putative involvement of D2-like receptor in this response was examined. No modification of the suppressant effect of microiontophoretically applied NE was detected in presence of raclopride or haloperidol, thereby confirming the lack of regulation of pyramidal CA3 neurons by D2 receptors in the hippocampus (Fig. 8, A-C). Finally, the spontaneous firing activity of CA3 pyramidal neurons remained unchanged after the local application of raclopride (17.3 ± 2.1 versus 13.4 ± 2.4 Hz, before and after its application, respectively; n = 8, P > 0.05) or of idazoxan (13.4 ± 4.2 versus 13.7 ± 4.9 Hz, before and after its application, respectively; n = 5, P > 0.05).

Effects of D2-like receptor antagonist raclopride on iontophoretically applied DA-induced inhibition of dorsal hippocampus CA3 pyramidal neurons. A, comparative effects of microiontophoretically applied DA, NE, and quinpirole on the firing rates of CA3 pyramidal neurons. Data are expressed as means ± S.E.M. of the number of spikes suppressed by DA (0.1 M) or NE (0.01 M) for CA3 pyramidal neurons. B, integrated firing rate histograms illustrating the effects of raclopride on DA-induced decrease in CA3 pyramidal neuronal activity. Horizontal bars indicate the duration of iontophoretic ejection and current values in nanoamperes. (C, D, and E, responsiveness of CA3 pyramidal neurons to iontophoretic applications of DA or quinpirole (C) or DA in the presence of raclopride (0.025 M) (D) or haloperidol (200 μg/kg i.v.) (E). Data are expressed as mean ± S.E.M. of the number of spikes suppressed by nanoamperes for CA3 pyramidal neurons. The number of neurons tested is indicated in each histogram.
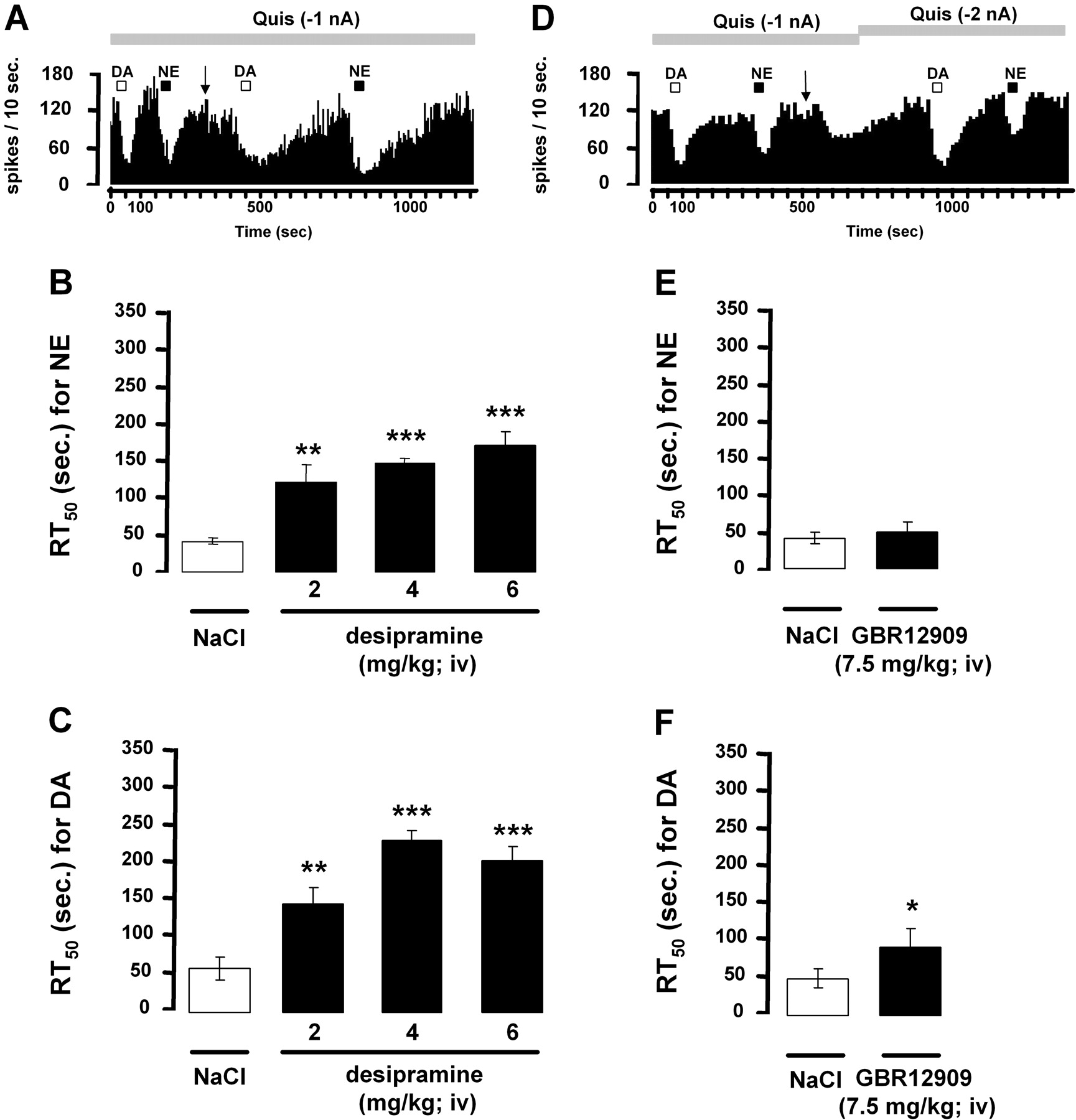
Inhibitory Action of Iontophoretically Applied DA on CA3 Pyramidal Neurons in the Presence of the Selective NE Reuptake Inhibitor Desipramine or the Selective DA Reuptake Inhibitor GBR12909. The recovery time, from the suppression of hippocampus pyramidal neuron firing activity after microiontophoretic application of NE or DA, was assessed by determining RT50 values before and after the short-term intravenous administration of desipramine or GBR12909. In agreement with previous data, cumulative doses of desipramine (2, 4, and 6 mg/kg i.v.) did not significantly modify the firing activity of CA3 pyramidal neurons. After the administration of desipramine (2 mg/kg i.v.), the RT50 values were significantly increased after the application of NE and DA (Fig. 9, A-C). Cumulative injections of desipramine (4 and 6 mg/kg) further increased RT50 values for both monoamines, whereas GBR12909 (7.5 mg/kg i.v.) decreased the firing activity of pyramidal neurons by 42%. A small but significant increase in the RT50 value of DA but not NE was obtained after the short-term administration of GBR12909 (Fig. 9, D-F).

Effect of α2-adrenoceptor antagonist idazoxan on iontophoretically applied DA-induced inhibition of dorsal hippocampus CA3 pyramidal neurons. A, integrated firing rate histograms illustrating the effects of idazoxan on DA-induced decrease in CA3 pyramidal neuronal activity. Horizontal bars indicate the duration of iontophoretic ejection and current values in nanoamperes. B, responsiveness of CA3 pyramidal neurons to iontophoretic applications of DA alone and in the presence of idazoxan (0.05 M). Data are expressed as mean ± S.E.M. of the number of spikes suppressed by nanoamperes for CA3 pyramidal neurons. The number of neurons tested is indicated in each histogram. *, P < 0.05, significantly different from DA alone by two-tailed Student's t test.
Discussion
The present electrophysiological data show that the microiontophoretic application of DA and NE inhibits the spontaneous firing activity of VTA DA, LC NE, and hippocampal CA3 pyramidal neurons. In the VTA, the suppressant effects of DA and NE were blocked not only by the D2-like receptor antagonist raclopride, but also by the α2-adrenoceptor antagonist idazoxan. In the LC and dorsal hippocampus, the suppressant effect of both catecholamines was only attenuated by idazoxan.
In the VTA, the suppressant effect of both DA and quinpirole on DA neurons and their blockade by the D2-like receptor antagonist raclopride further support the involvement of D2 receptor in the DA response. The observation that piperoxane, a nonselective α-adrenoceptor antagonist, also attenuates the inhibitory effects of DA (Freedman and Aghajanian, 1984) suggested actions on this class of receptors also. Indeed, the present study showed that the local application of the selective α2-adrenoceptor antagonist idazoxan attenuated the suppressant effect of DA on VTA DA neurons. Such an effect could not be attributed to a nonselective action of idazoxan because this compound has no affinity for DA receptors. Competition experiments in the rat cortex have shown that idazoxan has at least 1000-fold lower affinity for D2 receptors than for α2-adrenoceptors (Ki values, >10 μM and 8 nM, respectively) (Doxey et al., 1983; Neve et al., 1990). It thus seems that in addition to the stimulation of D2 receptors, DA also inhibits the firing activity of DA neurons by acting upon α2-adrenoceptors. Although functional studies previously emphasized such an unselective property of DA in the central nervous system (Cornil et al., 2008), the role of α2-adrenoceptors in a direct regulation of DA neurons themselves remained debatable, especially because, in rat brain, the affinity of DA for α2-adrenoceptors is approximately 3- to 7-fold lower than that of NE (Boyajian et al., 1987). Moreover, despite the expression of α2-adrenoceptors on DA neurons in the VTA (Lee et al., 1998), it was reported that the microiontophoretic application of clonidine (20-120 nA) has no depressant effect on VTA DA neuronal activity (White and Wang, 1984a,b; Aghajanian and Bunney, 1977a,b). The apparent discrepancy between the microiontophoretic effects of clonidine and DA may be explained by the capacity of DA to bind and activate both D2 and α2-adrenoceptors. Indeed, the stimulation of both types of receptors by DA could be important for generating a robust inhibitory effect in the VTA. It is interesting that the systemic administration of low doses of clonidine, which probably activate presynaptic α2-adrenoceptors, does not modify the mean firing rate of VTA DA neurons but decreases their bursting activity (Gobbi et al., 2001; Georges and Aston-Jones, 2003). In contrast, higher doses of clonidine increase both the firing rate and bursting activity of VTA DA neurons (Gobbi et al., 2001). These findings are paradoxical in the light of the present data, but it is possible that systemic administration of adrenergic agonists involves long-loop feedback mechanisms that are not activated with local application. Another possibility is a blunting of the effect of locally applied clonidine by its concomitant binding to postsynaptic α1-adrenoceptors (Anden et al., 1976), whose activation stimulates VTA DA neurons (Grenhoff and Svensson, 1993; Grenhoff et al., 1993, 1995).

Effect of D2-like receptor antagonists on the responsiveness of dorsal hippocampus CA3 pyramidal neurons to iontophoretically applied NE. A, integrated firing rate histograms illustrating the effects of raclopride or haloperidol on NE-induced decreases in CA3 pyramidal neuronal activity. Horizontal bars indicate the duration of iontophoretic ejection and current values in nanoamperes. B and C, responsiveness of CA3 pyramidal neurons to iontophoretic applications of NE in presence or not of idazoxan (0.05 M, 10 nA) (B) or haloperidol (200 μg/kg i.v.) (C). Data are expressed as mean ± S.E.M. of the number of spikes suppressed by nA for CA3 pyramidal neurons. The number of neurons tested is indicated in each histogram.
The iontophoretic application of NE also reduced the firing activity of DA neurons. Previous reports have indicated that the inhibition of VTA DA neuronal activity induced by local application of NE may be prevented by the D2-like receptor antagonist sulpiride (Aghajanian and Bunney, 1977a,b; White and Wang, 1984a,b), but we show here that idazoxan also has the capacity of preventing the effects of NE. Taken together, these results indicate that NE and DA can activate D2 and α2-receptors in the VTA to inhibit DA neuronal activity.

Effect of the selective NE reuptake inhibitor desipramine or the selective DA reuptake inhibitor GBR12909 on RT50 values from iontophoretic applications of NE and DA on dorsal hippocampus CA3 pyramidal neurons. RT50 values (means ± S.E.M.) represent the time (in seconds) required by the neuron recorded to recover 50% of its firing activity from the end of the iontophoretic application of NE or DA. RT50 values have been determined before and after the short-term intravenous administration of desipramine (A-C) or GBR12909 (D-F). In A, the arrow indicate the injection of desipramine (2 mg/kg i.v.), and in D, the injection of GBR12909 (7.5 mg/kg i.v.). In the later experiment, because an inhibition of the firing activity was produced by GBR12909, the current of quisqualate had to be increased to restore a firing rate similar to that before the injection. The number of neurons tested is indicated in each histogram. *, P < 0.05, **, P < 0.01, ***, P < 0.001, significantly different from the preinjection value by a one-way analysis of variance followed by a protected least-significance difference post hoc test (desipramine experiments) or a two-tailed Student's t test (GBR12909 experiments).
In the LC, prior results have already suggested a higher sensitivity for NE than DA, because a similar inhibition of NE neurons was detected after the ejection of both NE and DA but with a 10-fold lower concentration of NE than DA. The earlier observation that iontophoretically applied NE-induced inhibition of LC NE neurons is blocked by piperoxane (Cedarbaum and Aghajanian, 1977) was compatible with an involvement of α-adrenoceptors. Consistent with these results, the inhibitory effects of both NE and DA here reported were antagonized by idazoxan. Although the inhibitory influence of NE through α2-adrenoceptor on LC NE neurons has been extensively studied (Svensson et al., 1975; Cedarbaum and Aghajanian, 1977), evidence that DA stimulates α2-adrenoceptors to reduce, at least in part, LC NE neuronal firing, was not clearly documented.
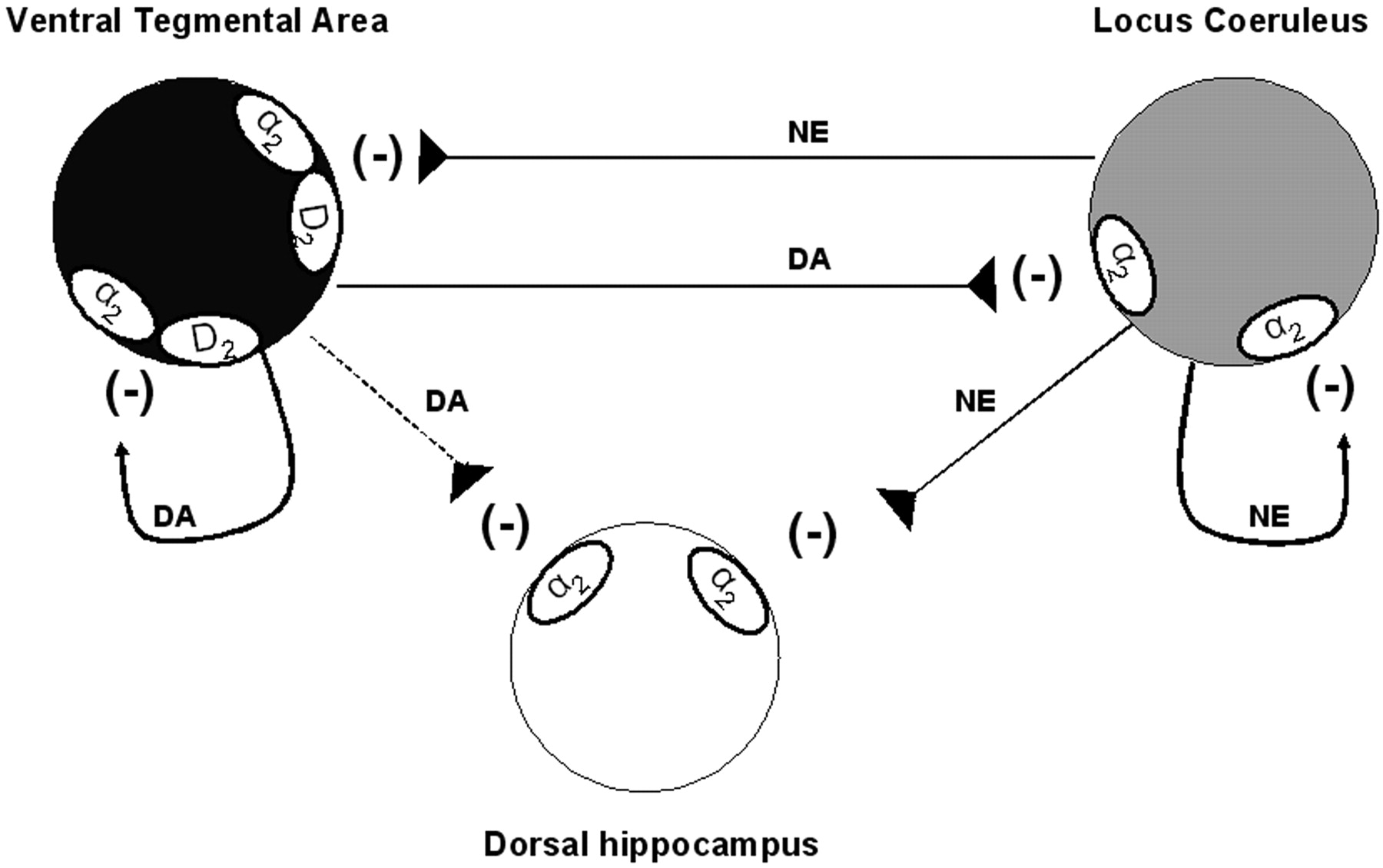
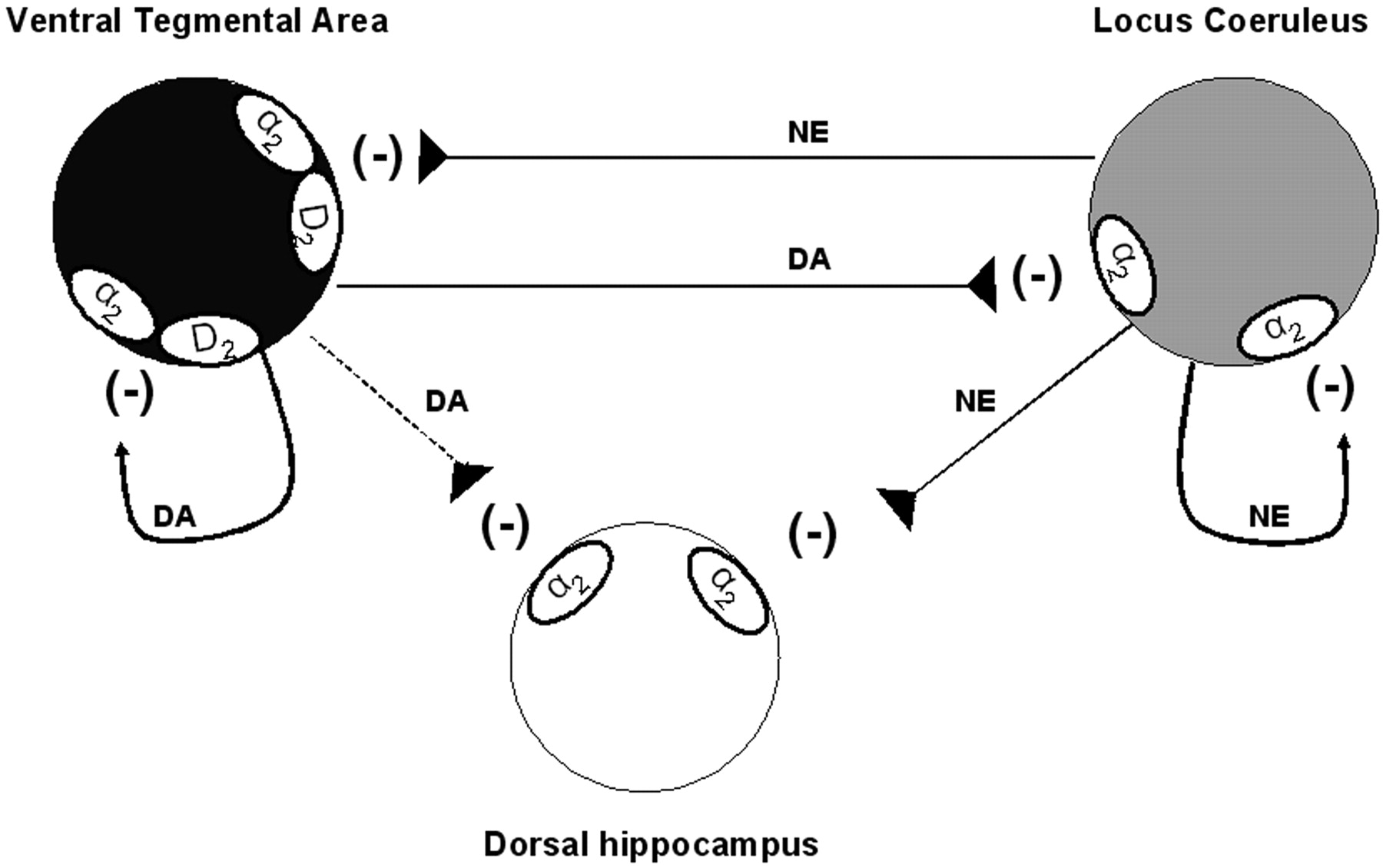
Model of the regulation of neuronal activities of ventral tegmental area, locus ceruleus, and dorsal hippocampus by DA and NE at the cell body level. In the VTA, DA, and NE act on both D2 and α2 receptor types to inhibit the neuronal firing of DA neurons. In the LC, despite the presence of D2 receptors, it seems that DA and NE exclusively stimulate α2 adrenoceptor thus inhibiting NE neuronal firing. The capacity of DA to bind and activate α2 adrenoceptor was also observed in the dorsal hippocampus. Although the release of DA from dopaminergic terminals in the dorsal hippocampus has not yet been demonstrated, the present results indicate that exogenously applied DA can be removed from the extracellular space by the NE transporter.
Because D2 receptors have been identified in the LC (Yokoyama et al., 1994), their direct activation on NE neurons after the iontophoretic application of NE or DA could not be excluded. However, the present study showed that raclopride has no effect on NE- or DA-induced inhibition of LC NE neurons at concentrations and currents found to exert a potent inhibition of VTA DA neurons (Fig. 2C). These results are consistent with the previous demonstration that the nonselective DA receptor antagonist trifluoperazine is ineffective in blocking the inhibition of LC neurons induced by both DA and NE (Cedarbaum and Aghajanian, 1977) and that the capacity of clonidine to inhibit NE neuronal firing is unaltered by haloperidol (Piercey et al., 1994). The putative lack of regulation of LC neurons by D2-like receptors is also evidenced by the observation that these NE neurons are insensitive to the local application of D2-like receptor agonists, such as (+)-3-PP, apomorphine (Aghajanian and Bunney, 1977; Elam et al., 1986), or quinpirole as shown in the present study. Altogether, these observations converge to show that only α2-adrenoceptors are involved in the inhibitory control of LC NE exerted by NE and DA, even if the systemic administration of haloperidol has been reported to enhance the firing rate of LC NE neurons (Piercey et al., 1994; Nilsson et al., 2005). Such an effect could not be related to an antagonistic activity of haloperidol on α2-adrenoceptor (U'Prichard et al., 1977), because the systemic administration of haloperidol in the present study modified neither the spontaneous firing activity of LC NE neurons nor the inhibitory actions of NE or DA. Other studies suggest an excitatory effect of haloperidol on NE neurons that may initially be driven through the local release of glutamate in the LC (Nilsson et al., 2005).
In the dorsal hippocampus, a possible role for DA had to be considered because the iontophoretic application of DA and NE inhibited the firing activity of CA3 pyramidal neurons. However, this DA-induced inhibition of firing was only partial, as if some of the DA effects were counterbalanced by an excitatory component. Given that β-adrenoceptors exert an excitatory action on hippocampus pyramidal neurons (Curet and de Montigny, 1988b), it is possible that the net biological response to DA results from opposite effects exerted on various adrenoceptors. In keeping with a weak expression of DA receptors and raclopride binding sites in the dorsal hippocampus (Dubois et al., 1986; Delis et al., 2004), it was reported that neither the iontophoretic application of raclopride nor the systemic injection of haloperidol blocked DA- or NE-induced inhibition in the dorsal hippocampus. Although haloperidol is known to bind and antagonize both D2 and α1-receptors (Cohen and Lipinski, 1986), the possibility that it blocked the adrenoceptors in the present study can be excluded, because the administration of the selective α1-adrenoceptor antagonist prazosin reduced the effect of NE in a dose-dependent manner (Curet and de Montigny, 1988a) and was here devoid of antagonistic activity. In contrast, the inhibitory action of DA was partially blocked by idazoxan at concentrations that usually antagonize the inhibitory effects of NE or clonidine on CA3 pyramidal neurons (Curet and de Montigny, 1988a). The involvement of postsynaptic α2-adrenoceptors in the electrophysiological response to DA, as described above in VTA and LC, was strongly supported by the latter results. To better understand the physiological importance of such a property, the possibility that NE neurons themselves could be the main source of DA in the hippocampus was addressed. The observation that the selective NE reuptake inhibitor desipramine prolonged the inhibitory effects of microiontophoretic applied DA strongly suggested that the clearance of DA in the hippocampus is mediated, at least in part, by the NE transporter (NET). This is consistent with previous data showing that DA has a greater affinity for the NET than the DA transporter (DAT) itself (Giros et al., 1994) and that DA reuptake by NE terminals occurs in brain regions in which DAT expression is minimal (e.g., the frontal cortex), intermediate, or maximal (e.g., nucleus accumbens shell and the bed nucleus, respectively) (Bymaster et al., 2002; Morón et al., 2002; Carboni and Silvagni, 2004). Recent experiments have proposed that DA may be coreleased with NE from noradrenergic terminals in several cortical areas (Devoto et al., 2004). Although it is not clear whether this feature might be related to a previous nonspecific uptake of DA by NE terminals, it is proposed here that DA is taken up by the NET in the hippocampus (Fig. 9), as reported previously in the frontal cortex (Bymaster et al., 2002). It is interesting that the selective DA reuptake inhibitor GBR12909 also produced a small but significant increase in the RT50 value of iontophoretically applied DA, which could hardly be ascribed to a nonselective binding of GBR12909 to the NET because the RT50 value of NE was not altered in this experiment. Thus, in the dorsal hippocampus, DA uptake could be mediated by distinct transport systems. The involvement of DAT was somewhat unexpected because in brain regions in which DAT is weakly expressed, as in the hippocampus (Delis et al., 2004; Dahlin et al., 2007), a limited role of DAT in removing DA from the extracellular space has been reported (Morón et al., 2002). An alternative mechanism could be that residual DA uptake was mediated by the recently cloned and characterized polyspecific cation-monoamine transporters organic cation transporters (2 and 3) or plasma membrane monoamine transporter. Indeed, these transporters are expressed in the rat hippocampus (Dahlin et al., 2007), but high concentrations of GBR12909 are necessary to block their activity (Morón et al., 2002). In view of these results and of the high ratio NET/DAT in hippocampus, it seems likely that, in this brain region, DA is captured by NET before it reaches other transport sites.
In summary, the present findings showed that, depending on the brain structure studied, DA and NE activate D2 and/or α2-adrenoceptors to exert inhibitory postsynaptic effects (Fig. 10). This is in agreement with previous studies showing the in vivo and in vitro stereoselective interactions of DA with α2-adrenoceptors (Zhang et al., 2004; Cornil et al., 2008) and NE with D2-like receptors (Newman-Tancredi et al., 1997; Wedemeyer et al., 2007). The fact that DA can act in concert with the NE system to strengthen the intensity of postsynaptic noradrenergic responses may have important implications in the treatment of mood disorders.
Footnotes
-
This study was supported by the Canadian Institutes for Health Research grant (77838) and salary support from the University of Ottawa Institute of Mental Health Research (to B.P.G. and M.E.), as well as a Canada Research Chair in Psychopharmacology from the Canadian Government, and an Endowed Chair from the University of Ottawa Institute of Mental Health Research (to P.B.).
-
ABBREVIATIONS: DA, dopamine; CA, field of the hippocampus; DAT, dopamine transporter; GBR12909, 1-(2-[bis(4-fluorophenyl)methoxy]ethyl)-4-(3-phenylpropyl)piperazine; NE, norepinephrine; NET, norepinephrine transporter; LC, locus ceruleus; RT, recovery time; VTA, ventral tegmental area; (+)-3-PP, 3-(3-hydroxyphenyl)-N-n-propylpiperidine.
- Received April 18, 2008.
- Accepted August 14, 2008.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}