

Oscillatory activity is a universal design feature of olfactory systems (Tank et al., 1994), having been demonstrated by Lord Adrian in the olfactory bulb of the hedgehog >60 years ago (Adrian, 1942). Hints as to the computational role of olfactory oscillations have only recently begun to emerge (Stopfer et al., 1997; Teyke and Gelperin, 1999; Nusser et al., 2001) (but see Fletcher et al., 2005). The effort to understand the role of olfactory oscillations is aided by insights from computational models of olfactory networks incorporating realistic dynamics at the cellular and network levels (Linster and Cleland, 2001; Davison et al., 2003; Ermentrout et al., 2004; Sivan and Kopell, 2004; Bazhenov et al., 2005; Galan et al., 2005; Migliore et al., 2005). I will review some of the major issues we face in attempts to understand the computational and behavioral roles of olfactory oscillations (Gelperin, 1999), in the context of more general efforts to understand cortical oscillations in a variety of sensory processing and motor control pathways (Salinas and Sejnowski, 2001; Buzsaki and Draguhn, 2004; Borgers et al., 2005; Mann and Paulsen, 2005; Schnitzler and Gross, 2005).
What is being measured?
Coherent oscillatory activity in olfactory networks is expressed in both periodic transmembrane potential fluctuations (Margrie and Schaefer, 2003; Balu et al., 2004; Hayar et al., 2004a) and local field potential (LFP) measurements (Gelperin and Tank, 1990; Dorries and Kauer, 2000), if permitted by the geometrical and electrotonic properties of the neurons experiencing rhythmic transmembrane events (Hubbard et al., 1969; Harris et al., 2000). A major contributor to the local field potential fluctuations is coherent synaptic activity within a local population of neurons. In the case of the olfactory bulb, the reciprocal synaptic connections between mitral and granule cells give rise to oscillatory local field potentials in the 40–100 Hz (gamma) range mainly because of synaptic currents in granule cells (Neville and Haberly, 2003). It is now clear that the gamma band is composed of two distinct components that predominate in different behavioral states (Kay, 2003). Local field potential oscillations in the beta frequency range (15–40 Hz) result from reciprocal synaptic interactions between the olfactory bulb and the piriform cortex (Neville and Haberly, 2003). The relative prominence of beta and gamma oscillations is dramatically altered during the progress of learning new meanings for an odor cue (Ravel et al., 2003). These studies also make clear the critical importance of measurements made in the awake behaving animal during odor-guided decision making as an essential adjunct to studies in anesthetized animals and olfactory bulb slice preparations.
A comparative perspective
A comparative perspective on olfactory information processing has identified a set of 14 design features and circuit properties found across wide swaths of the phylogenetic tree (Hildebrand and Shepherd, 1997; Gelperin, 1999). These include a critical role for local inhibitory interneurons in shaping network dynamics (Gelperin and Tank, 1990; Lagier et al., 2004; Murphy et al., 2005; Saghatelyan et al., 2005), spontaneous or odor-induced oscillatory activity (Adrian, 1942; Tank et al., 1994; Delaney and Hall, 1996; Nikonov et al., 2002; Lam et al., 2003; Ravel et al., 2003), changes in odor representations attributable to learning in the first stage of central odor processing (Freeman and Schneider, 1982; Kimura et al., 1998; Sandoz et al., 2003; Daly et al., 2004; Martin et al., 2004; Wilson et al., 2004; Kirino et al., 2005), addition of new circuit elements during postpartum odor experience (Mair et al., 1982; Zakharov et al., 1998; Beltz and Sandeman, 2003; Saghatelyan et al., 2005), evidence for spatial segregation of related odor representations (Friedrich and Korsching, 1997; Kimura et al., 1998; Mori et al., 1999; Galizia and Menzel, 2000; Luo and Katz, 2001; Wang et al., 2003; Johnson et al., 2005), temporal evolution of odor representations (Laurent et al., 2001; Lei et al., 2004; Szyszka et al., 2005; Zochowski and Cohen, 2005), role for both chemical and electrical coupling between network elements (Friedman and Strowbridge, 2003; Zhang and Restrepo, 2003; Ermentrout et al., 2004; Christie et al., 2005), control of network dynamics and plasticity by nitric oxide (NO) (Kendrick et al., 1997; Gelperin et al., 2000; Collmann et al., 2004; Fujie et al., 2005; Korneev et al., 2005), synchronization of odor-responsive output neurons (Laurent, 2002; Brody and Hopfield, 2003; Christensen et al., 2003; Friedrich et al., 2004; Hayar et al., 2004b), use of coupled burster neurons to enhance network responses (Ermentrout et al., 2001; Hayar et al., 2004b), continual turnover of receptor cells (Chase and Rieling, 1986; Farbman, 1994), receptor neurons map to glomeruli based on receptor gene expression (Vosshall et al., 2000; Wang et al., 2003; Buck, 2004; Zou et al., 2004), afferents interact with relay neurons in glomeruli (Chase and Tolloczko, 1986; Galizia et al., 1999; Wachowiak et al., 2004), and expression of speed–accuracy tradeoff (Ditzen et al., 2003; Uchida and Mainen, 2003; Abraham et al., 2004; Khan and Sobel, 2004; Friedrich, 2005). This set of features may be incomplete but nevertheless serves to highlight the functional analogies found in a very diverse set of olfactory information processing systems.
The comparative approach reflected above takes seriously the notion that not only olfactory information processing but also human diseases, learning, and other cognitive functions can be fruitfully studied in both mammalian and nonmammalian species (Kazemi-Esfarjani and Benzer, 2002; Barco et al., 2003; Bonini and Fortini, 2003; Greenspan and van Swinderen, 2004). Given this perspective, it is now useful to look at a particular set of molluscan olfactory systems and see in more detail how they illuminate general issues of the functions of olfactory oscillations.
Molluscan model systems
“For many problems there is an animal on which it can be most conveniently studied.” —August Krogh
The terrestrial pulmonate mollusks epitomize Krogh’s principle (Krebs, 1975) for the study of olfactory information processing and in particular the computational role of cellular and network oscillations in olfaction (Gelperin, 1999; Chase, 2002; Kirino et al., 2005). The terrestrial slugs and snails are dominated by olfaction for orientation, nutrition, and reproduction. This macrosmatic lifestyle is reflected in the dedication of most of the neurons in the CNS (∼104) to the processing of olfactory information (Chase, 2000). There are two major olfactory processing structures: the digitate ganglion at the superior tentacle tip right behind the nose (Ito et al., 2000) and the procerebral (PC) lobe in the CNS. The PC lobe receives both first-order input from olfactory receptors and second-order input from the digitate ganglion (Chase and Kamil, 1983; Murakami et al., 2004).
Oscillatory LFPs are recorded from both the digitate ganglion (Ito et al., 2004) and the PC lobe (Gelperin and Tank, 1990; Kawahara et al., 1997; Nikitin and Balaban, 2000). In the PC lobe, activity waves are initiated at the apical pole and propagate to the base at 1.1 mm/s (Delaney et al., 1994; Toda et al., 2000; Watanabe et al., 2004). At a stationary LFP recording site, the recorded oscillation (0.7 Hz in vitro) corresponds to the passing front of the activity wave. Recordings of PC lobe activity in intact behaving slugs implanted with fine-wire electrodes show periods of 0.7 Hz oscillation interspersed with periods of complex multicomponent activity (Cooke and Gelperin, 2001). Optical recordings of the PC lobe wave activity during odor stimulation of the nose show a momentary collapse of the activity wave (Kleinfeld et al., 1994), which is hypothesized to allow nearest-neighbor interactions suppressed by the traveling wave activity (Ermentrout and Kleinfeld, 2001). Activity waves have also been recorded from an in vitro preparation of the ferret thalamus (Kim et al., 1995) and from turtle visual cortex (Prechtl et al., 2000; Robbins and Senseman, 2004).
The LFP oscillations and activity wave in the Limax PC lobe arise from intrinsic neuronal properties and network connections among a population of bursting inhibitory interneurons, called B cells (Fig. 1) (Watanabe et al., 1998; Wang et al., 2001). These cells are connected by both chemical and electrical synapses (Ermentrout et al., 2004) and show a gradient of excitability with apical B cells most excitable (higher burst rate), accounting for normal initiation of the activity wave at the apical end in the in vitro preparation (Ermentrout et al., 1998). The gradient of B-cell excitability is attributable to a spatial gradient in the amplitude of chloride-dependent depolarizations in B cells (Watanabe et al., 2003). Activity waves occupy the full width of the PC lobe, most likely attributable to the rapid lateral conduction speed of activity in varicose B cells compared with the apical–basal conduction speed of activity in smooth B cells (Wang et al., 2001). The band-like conduction of activity along the apical–basal axis of the PC lobe is related to the band-like storage of learned odor representations in the PC lobe (Kimura et al., 1998; Teyke et al., 2000). Modification of PC lobe cell properties by learning reflects the general finding that learning affects odor representations in the earliest stages of odor processing (Kay and Laurent, 1999; Wilson et al., 2004).
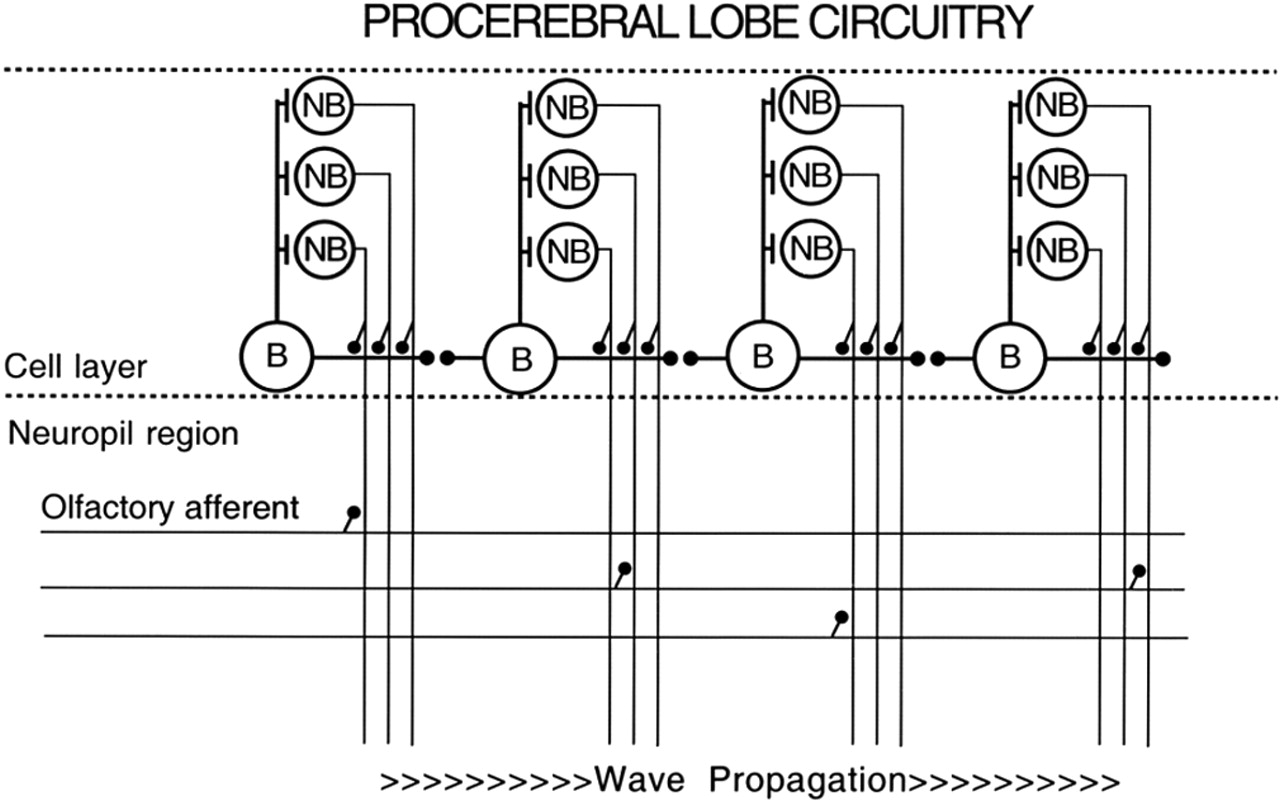
Diagram of the intrinsic circuitry of the Limax PC lobe. The intrinsic excitability of the B cells is highest on the left and decreases moving to the right. The B cells are connected by both chemical and electrical synapses. Each B cell connects with and inhibits ∼100 nonbursting (NB) cells, only three of which are shown in the diagram. The processes of the B cells are confined to the cell body layer of the PC lobe, whereas the neurites of the NB cells project into the neuropil layer in which they receive synapses from olfactory afferents and second-order neurons in the digitate ganglion (data not shown). The NB cells also synapse in the neuropil with the neurites of output neurons whose somata are located in the pedal and buccal ganglia (Gelperin and Flores, 1997) and in the metacerebrum (Ratté and Chase, 2000; Shimozono et al., 2001).
The excitability of B cells is modulated by NO (Gelperin, 1994). Suppression of NO synthesis shuts down the LFP oscillation and wave propagation in the PC lobe (Gelperin et al., 2000). Suppression of NO synthesis in the behaving snail (Teyke, 1996) or slug (Sakura et al., 2004) blocks fine odor discrimination or odor learning. These observations of degraded odor processing and odor learning after inhibition of NO synthesis are paralleled by findings in ewes (Kendrick et al., 1997), honeybees (Müller, 1996; Hosler and Smith, 2000), the predatory snail Euglandina (Clifford et al., 2003), mice (Okere et al., 1996), and rats (Samama and Boehm, 1999). The relationship between nitric oxide and synaptic plasticity has been reviewed recently (Susswein et al., 2004).
Computational implications of olfactory oscillations
Recent work has explored the computational utility of synchronous activity in a group of neurons coding stimulus identity, with direct relevance to olfaction (Brody and Hopfield, 2003; Hopfield and Brody, 2004). Part of the mechanism to enhance synchronous activity is a shared membrane potential oscillation (Margrie and Schaefer, 2003), which may result in an oscillation of LFP, depending on geometrical and electrotonic properties of the cells showing the coherent changes in membrane potential. Olfactory interneurons can show odor-elicited synchronous activity not phased to the oscillating LFP (Christensen et al., 2003).
The temporal evolution of activity in the group of synchronously active interneurons can result in distinct temporal patterns within the odor-responsive population (Stopfer and Laurent, 1999; Christensen et al., 2003). As the pattern of activity evolves in time, sparsening of the representation can reduce the overlap of patterns set up by closely related odors (Friedrich et al., 2004; Szyszka et al., 2005). The amplitude and regularity of the LFP oscillation also shows a temporal evolution after odor onset, typically becoming larger and more regular in the first several hundred milliseconds after stimulus onset. There are also history-dependent effects such that a series of closely spaced odor stimuli result in successive epochs of oscillatory responses, with sharper LFP oscillation patterns triggered at shorter latency over the first three odor stimuli (Stopfer and Laurent, 1999). These events provide an opportunity to determine whether the latency for odor identification shows a parallel decrease in latency over the first three odor pulses, as measured by latency to emit a conditioned response (Daly et al., 2004; Yu et al., 2004).
The Limax olfactory system allows the causal link between the LFP oscillation in the PC lobe and odorant-induced behavior to be explored using an in vitro nose–brain preparation (Teyke and Gelperin, 1999; Inoue et al., 2004). Recent work by Inoue et al. (2004) uses the neural substrate of a response to aversive odors, firing of an identified mantle motor neuron, to read out the behavioral decision made by the isolated brain in response to odor stimulation. Previous conditioning of the animal creates an aversive response to a previously attractive odor (Sahley et al., 1981), which survives in the isolated nose–brain preparation (Teyke et al., 2000). Application of an aversively conditioned odor to the superior nose of the in vitro nose–brain preparation causes an increase in LFP oscillation frequency in the PC lobe and firing of the identified mantle motor neuron (Inoue et al., 2004). Directly increasing LFP oscillation frequency in the PC lobe does not activate the mantle motor neuron. It may be that the motor pathway for the aversive mantle reflex is activated in parallel with an increase in PC lobe LFP frequency as learning-related changes are engaged in the PC lobe (Ermentrout et al., 2004). If the naive nose–brain preparation can be trained in vitro while the pharmacology of the PC lobe is selectively manipulated, the role of the PC lobe LFP oscillation and activity wave in odor recognition and odor learning can be directly tested. This possibility is encouraged by the demonstrated learning ability of the isolated Limax lip–brain preparation (Chang and Gelperin, 1980) along with other isolated CNS preparations (Muhlethaler et al., 1993; Kemenes et al., 1997; Mokin and Keifer, 2005).
Multisite network measurements
Tests of computational models of synchronous activity require simultaneous monitoring of activity in dozens or hundreds of olfactory interneurons to provide an adequate sample of neurons responding to a given odor. Arrays of silicon microprobes (Christensen et al., 2000), tetrodes (Egana et al., 2005), and multiple single-unit electrodes (D. Rinberg, A. Koulakov, A. Gelperin, unpublished observations) are beginning to provide simultaneous access to sufficient numbers of mitral cells or projection neurons (insect analog of mitral cells) so that spatial and temporal evolution of odor-responsive cell populations can be analyzed in detail in the awake behaving animal. One or more electrodes in the array are devoted to measuring LFPs so that correlations between responding interneurons can also be indexed to a common field potential (Wang et al., 2003).
Optical recordings in the awake head-fixed animal (Margrie et al., 2002) have the potential to permit a behavioral readout of the results of olfactory processing, whereas voltage- or calcium-sensitive dyes or genetically encoded markers (Bozza et al., 2004) sample large populations of olfactory interneurons. It will be interesting to make these population measurements in animals with substantially reduced olfactory processing networks that spare significant olfactory ability (de Belle and Heisenberg, 1994; Slotnick et al., 2004; Komischke et al., 2005).
Network perturbations to probe causality
The critical need is to test the computational role of synchronous activity and network oscillations using selective perturbations of central olfactory circuits while measuring odor identification and discrimination. The experiments of Stopfer et al. (1997) suggest that pharmacologically induced desynchronization of first-order olfactory interneurons degrades discrimination of similar odors but not dissimilar odors. A similar result was obtained relating oscillations in the Limax PC lobe to odor discriminations, as tested using an in vitro lip–brain preparation (Teyke and Gelperin, 1999). The Limax experiment will be even more informative using a newly developed in vitro nose–brain preparation, also using motor neuron activation to index the attractive or repellent nature of the applied odor (Inoue et al., 2004). Other perturbations used to explore the causal link between olfactory circuit function and odor-guided behavior include lesions of the olfactory bulbs (Slotnick et al., 2004), genetic lesions of specific neurotransmitter receptor subtypes (Nusser et al., 2001) or ionic channel subunits (Fadool et al., 2004) in the olfactory bulb, pharmacological manipulations of the olfactory bulbs during odor-guided behavioral tasks (Ravel et al., 1994; Kendrick et al., 1997), and direct electrical stimulation of the olfactory bulb in awake behaving animals to create “electric odors” (Jirsa and Radil, 1997; Mouly and Gervais, 2002; Roman et al., 2004). Creative use of these and other methods for selective perturbation of olfactory networks during their participation in olfactory computations and the readout of the effects of these perturbations in odor-guided behavior is critical to fully clarify the computational role of olfactory oscillations and synchronous cellular activity during odor processing.
Footnotes
This work was supported by the Army Research Office, the Whitehall Foundation, and National Institutes of Health Grant MH56090.
- Correspondence should be addressed to Dr. Alan Gelperin, Monell Chemical Senses Center, 3500 Market Street, Philadelphia, PA 19104. Email: agelperin{at}monell.org





{kind=link}