

Abstract
It is thought that galanin, a 29 amino acid neuropeptide, is involved in various neuronal functions, including the regulation of food intake and hormone release. Consistent with this idea, galanin receptors have been demonstrated throughout the brain, with high levels being observed in the hypothalamus. However, little is known about the mechanisms by which galanin elicits its actions in the brain. Therefore, we studied the effects of galanin and its analogs on synaptic transmission using an in vitro slice preparation of rat hypothalamus. In arcuate nucleus neurons, application of galanin resulted in an inhibition of evoked glutamatergic EPSCs and a decrease in paired-pulse depression, indicating a presynaptic action. The fragments galanin 1–16 and 1–15 produced a robust depression of synaptic transmission, whereas the fragment 3–29 produced a lesser degree of depression. The chimeric peptides C7, M15, M32, and M40, which have been reported to antagonize some actions of galanin, all produced varying degrees of depression of evoked EPSCs. In a minority of cases, C7, M15, and M40 antagonized the actions of galanin. Analysis of mEPSCs in the presence of TTX and Cd2+, or after application of α-latrotoxin, indicated a site of action for galanin downstream of Ca2+ entry. Thus, our data suggest that galanin acts via several subtypes of presynaptic receptors to depress synaptic transmission in the rat arcuate nucleus.
Galanin, a 29 amino acid neuroregulatory peptide, has been implicated in numerous neuronal functions including feeding behavior (Crawley et al., 1990; Kyrkouli et al., 1990) and regulation of hormone release (Bauer et al., 1986;Bjorkstrand et al., 1993; Kondo et al., 1993; Sahu et al., 1994;Rossmanith et al., 1996). These two effects of galanin are thought to result from its actions within the hypothalamus. Galanin receptor binding has been demonstrated to be high within the hypothalamus, particularly within the arcuate and paraventricular nuclei (Skofitsch and Jacobowitz, 1985; Melander et al., 1986; Vrontakis et al., 1991;Merchenthaler et al., 1993). Thus, galanin very likely plays a significant role in hypothalamic function, although the mechanisms underlying its actions are unclear.
Two galanin receptors, GALR1 and GALR2, have been cloned to date (Habert-Ortoli et al., 1994; Howard et al., 1997). Binding and other studies using galanin analogs have indicated that additional subtypes of galanin receptors are likely to exist. Chimeric peptides C7, M40, M32, and M15 have been show to act as galanin antagonists in certain systems, including feeding behavior (Bartfai et al., 1993; Corwin et al., 1993; Crawley et al., 1993), stimulation (Pramanik andÖgren, 1993) and inhibition (Bartfai et al., 1991, 1993; Lindskog et al., 1992) of acetylcholine release in the striatum and hippocampus, respectively, inhibition of glutamate and aspartate release in the hippocampus (Zini et al., 1993), and stimulation of luteinizing hormone release (Sahu et al., 1994). However, in other systems, such as galanin-mediated inhibition of cAMP accumulation in RINm5F cells (Bartfai et al., 1993), inhibition of glucose-induced insulin release (Bartfai et al., 1993), facilitation of the spinal cord reflex (Bartfai et al., 1993), mobilization of intracellular Ca2+stores (Fridolf and Ahrén, 1993), and hyperpolarization of magnocellular neurosecretory cells (Papas and Bourque, 1997), these same compounds are inactive or act as partial or full agonists. In addition, the galanin fragments 1–15 and 3–29 have been shown to behave as full agonists in some preparations (Wynick et al., 1993;Hedlund et al., 1994; Lorinet et al., 1994), whereas they have no effect in others (Wynick et al., 1993; Lorinet et al., 1994), again suggesting the existence of additional galanin receptor subtypes.
Recent work has identified various cellular effects, typical of G-protein-linked receptors, through which galanin might regulate neuronal function. Galanin is capable of opening K+channels and hyperpolarizing neurons (Ahrén et al., 1989; Dunne et al., 1989; Konopka et al., 1989; Parsons and Merriam, 1992; Papas and Bourque, 1997), inhibiting adenylate cyclase activity (Nishibori et al., 1988; Chen et al., 1992), inhibiting voltage-gated Ca2+ channels (Merriam and Parsons, 1995), inhibiting phosphoinositide turnover (Palazzi et al., 1988), and regulating the release of dopamine (Nordström et al., 1987), noradrenaline (Tsuda et al., 1989), acetylcholine (Fisone et al., 1987;Ögren and Pramanik, 1991; Ögren et al., 1993), and glutamate (Zini et al., 1993). The regulation of transmitter release is an important mechanism by which galanin could potentially modulate neuronal function. However, few studies to date have directly investigated these actions of galanin. We have now investigated the ability of galanin to modulate synaptic transmission within the arcuate nucleus of the hypothalamus. Our results indicate that galanin acts through multiple presynaptic receptors to inhibit the release process.
MATERIALS AND METHODS
Preparation of brain slices. The methods for the preparation of thin hypothalamic brain slices were similar to those described previously (Glaum et al., 1994). Experiments were conducted on Wistar rats of either sex, age 10–20 d postnatal. Animals were anesthetized with ether by inhalation, and killed by decapitation using a guillotine. The brain was removed rapidly by dissection and placed in chilled (0–6°C) extracellular solution of the following composition (in mm): NaCl 126, KCl 3, CaCl2 2.5, MgSO4 1.3, NaH2PO4 1.25, NaHCO3 26, and d-glucose 10 (gassed with 95% O2/5% CO2, pH 7.4; osmolarity = 310 mOsm). Thin (200- to 220-μm-thick) coronal slices of the arcuate nucleus of the hypothalamus were cut using a vibrating tissue chopper (Vibratome). Slices were maintained at 30–32°C until needed for recording.
For recording, slices were transferred to a submersion chamber mounted on the stage of an upright microscope (Leitz Laborlux) and viewed with a Zeiss 40× water immersion objective with Hoffman Contrast Optics. The slices were perfused continuously throughout the experiment with extracellular solution at room temperature (20–25°C). All recordings were made from visually identified neurons located in the arcuate nucleus. The arcuate nucleus is rich in galanin-positive neuronal cell bodies and may be an important structure in the regulation of reproduction and food intake (Landry et al., 1995).
Patch-clamp recording, synaptic stimulation, and data analysis. Patch-clamp recording pipettes were made from thin-walled borosilicate glass capillaries (DC resistance = 5–10 MΩ when filled with internal solution) using a Flaming-Brown horizontal pipette puller (Sutter Instruments). In all experiments, electrodes were filled with internal solution of the following composition (in mm): potassium gluconate 145, MgCl2 2, K2ATP 5, EGTA 1.1, CaCl20.1, and HEPES 5, pH 7.2; osmolarity adjusted to 280–290 mOsm. Patch recording pipettes were mounted in the headstage attached to a stage-mounted three-way hydraulic micromanipulator (Narashige) and positioned over the somas of neurons under visual control. Conventional methods for obtaining whole-cell recordings from thin slices (Hamill et al., 1981; Edwards et al., 1989) were used. After the attainment of cell access, transmembrane voltage and current were recorded using an Axoclamp 2B (Axon Instruments, Foster City, CA) amplifier in the discontinuous voltage-clamp mode (filtered at 5 kHz and acquired at 20 kHz), stored on a Pentium computer (Quantex) and via chart recorder (Gould), and analyzed using pClamp 6.0.1 (Axon Instruments).
Bipolar tungsten stimulating electrodes were placed lateral to the arcuate nucleus to activate inputs to arcuate nucleus neurons. In all experiments, stimuli of between 50 and 500 μsec were used to elicit synaptic responses and were maintained at 10 sec intervals to record the time-dependent effects of drug perfusion. Paired-pulse stimuli were recorded at a 20 sec interval using a 30 msec interpulse interval. In normal saline, the paired-pulse ratio was a random mixture of both paired-pulse facilitation and depression that likely resulted from the polysynaptic inputs recorded in arcuate neurons. Therefore, studies of paired stimuli were recorded in high Ca (5 mm) and low Mg (0.5 mm) saline solution, which increases the probability of neurotransmitter release in response to the initial stimulus and favors paired-pulse depression (Khazipov et al., 1995; Dobrunz and Stevens, 1997). These recording conditions significantly improved the reproducibility of the paired-pulse ratio recorded from arcuate neurons. For the purposes of data analysis, 2–3 min of evoked EPSCs were averaged (12–18 EPSCs), and the peak of the averaged EPSC was measured. Cells that responded to drug application with a 15% or greater reduction (a change greater than the 99% confidence limits of the control window mean) were considered to have responded positively. Data are expressed in mean ± SEM.
Recording and analysis of mEPSCs. Miniature EPSCs (mEPSCs) were recorded from arcuate nucleus neurons at a holding potential of −60 to −70 mV in the presence of 1 μm TTX, 40 μm 7-chlorokynurenic acid, 10 μmbicuculline, and, unless noted otherwise, 100 μm cadmium chloride (Cd2+). Currents were filtered at 1–2 kHz, sampled at 8 kHz, and acquired to disk using pClamp software. Cells were monitored periodically for changes in access resistance, and cells that exhibited any significant (>15%) changes during the recording period were rejected.
mEPSCs were analyzed using pClamp software or software written by Drs. Andreas Kyrozis, Romain Girod, and Daniel McGehee in Axobasic specifically for the analysis of mEPSCs. This program allows for the automatic screening of events based on amplitude or 10–90% rise time or both. All events were examined visually and accepted or rejected on the basis of subjective visual criteria as well as the objective criteria of amplitude, rise time, and decay time. Events that had an amplitude of >4 pA, rise times of between 300 μsec and 3 msec, and decay times of between 3 and 30 msec were included in the analysis.
Analyzed data from a 3–5 min recording period (100–1000 events) were examined and further analyzed using Prism (Graphpad) and Statmost (Datamost). Cumulative probability plots were constructed to visually examine the effects of galanin on the amplitude and interval distributions of mEPSCs, whereas amplitude and interval distributions were compared statistically using a Kolmogorov–Smirnov test or a Student’s unpaired t test with Welch’s correction. Differences in distributions were considered significant ifp < 0.05. Data are expressed in mean ± SEM.
Application of drugs. Drugs were dissolved in distilled water or DMSO (≤0.1% final concentration in artificial CSF) and applied by bath perfusion. The following compounds were used: bicuculline methobromide (Sigma, St. Louis, MO), 7-chlorokynurenic acid (Tocris Cookson), d,l-2-amino-5-phosphonpentanoic acid (d,l-AP5) (Tocris Cookson), TTX (Sigma), α-latrotoxin (Alomone Labs), (rat) galanin (American Peptides), galanin 1–16 (Bachem, Torrance, CA), galanin 1–15, 3–29, and M32 (gifts of Mary Walker, Synaptic Pharmaceuticals), galantide (M15) (American Peptides), M40 (American Peptides), and C7 (American Peptides). All drugs, with the exception of AMPA and α-latrotoxin, were applied for between 5 and 10 min to obtain a steady-state bath concentration. AMPA was applied 2–3 times in succession (25 μm for a period of 40 sec at 10 min intervals) to achieve a series of rapid inward currents (−60 to −150 pA), which were then averaged. In the case of α-latrotoxin, it was found that the time required for action of the toxin in an in vitro slice preparation was unfeasible; thus, slices were incubated for 2 hr in 2 nm α-latrotoxin and then transferred to the recording chamber, where the slice was then perfused with standard extracellular Ringer’s solution. The effects of α-latrotoxin are likely irreversible within the time course of this experiment (Wanke et al., 1986; Capogna et al., 1996a,b), and thus there should be no significant “washout” of its effects during the recording period. Such extensive exposure to α-latrotoxin could result in some depletion of transmitter stores (Tzeng et al., 1978;Hurlbut et al., 1990; McMahon et al., 1990; Storchak et al., 1994), as well as some loss of synaptic morphology (Tzeng et al., 1978). Under these conditions, however, a much greater percentage of cells (∼80%) was observed than under control conditions (∼25%), which displayed an mEPSC frequency adequate for analysis (>0.5 Hz) (see Results).
RESULTS
Whole-cell patch recordings were obtained from 110 neurons in 65 preparations of 200–220 μm thick slices of rat arcuate nucleus. Unless noted otherwise, all experiments were performed at a cell holding potential of between −60 and −70 mV in the presence of bicuculline (10 μm), 7-chlorokynurenic acid (40 μm), and 1.5 mm external Mg2+ to pharmacologically isolate the AMPA receptor-mediated EPSCs. The mean cellular input resistance was 1200 ± 74 MΩ (n = 71). The mean 10–90% rise time was 2.14 ± 0.09 msec (n = 50), and the mean 10–90% decay was 12.77 ± 0.49 msec (n = 50). No significant differences in values were observed between control and drug application, so the values were pooled. The figure for the 10–90% rise time indicates that many of the synaptic responses recorded were the result of multiple asynchronous inputs.
Galanin receptor-mediated synaptic depression
To investigate the effects of galanin on synaptic transmission in the hypothalamus, a stimulating electrode was placed ventrolateral to the arcuate nucleus, and synaptic inputs were stimulated at a frequency of 0.1 Hz while recording from arcuate neurons in the discontinuous voltage-clamp mode. In the presence of 7-chlorokynurenic acid and bicuculline, stimulation of inputs resulted in exclusively AMPA receptor-mediated EPSCs (Glaum et al., 1996). Under such conditions, application of galanin (galanin 1–29, 100 nm) caused a strong reduction of synaptic transmission in 19 of 26 cases examined (55.54 ± 4.03%; n = 19) (Figs.1A,2). In cases in which a sufficient washout period was recorded, there was some variability in the degree of recovery observed. In some cells the effect of galanin readily reversed during washout (Fig. 1A). However, in six cells there was a slow and incomplete recovery observed after a long (+40 min) washout period (data not shown). Such variability was observed with galanin 1–15 and galanin 1–16 as well (see below). These observations are consistent with previous reports of long-lasting actions of other peptides in the hypothalamus (Van den Pol et al., 1996; Rhim et al., 1997)
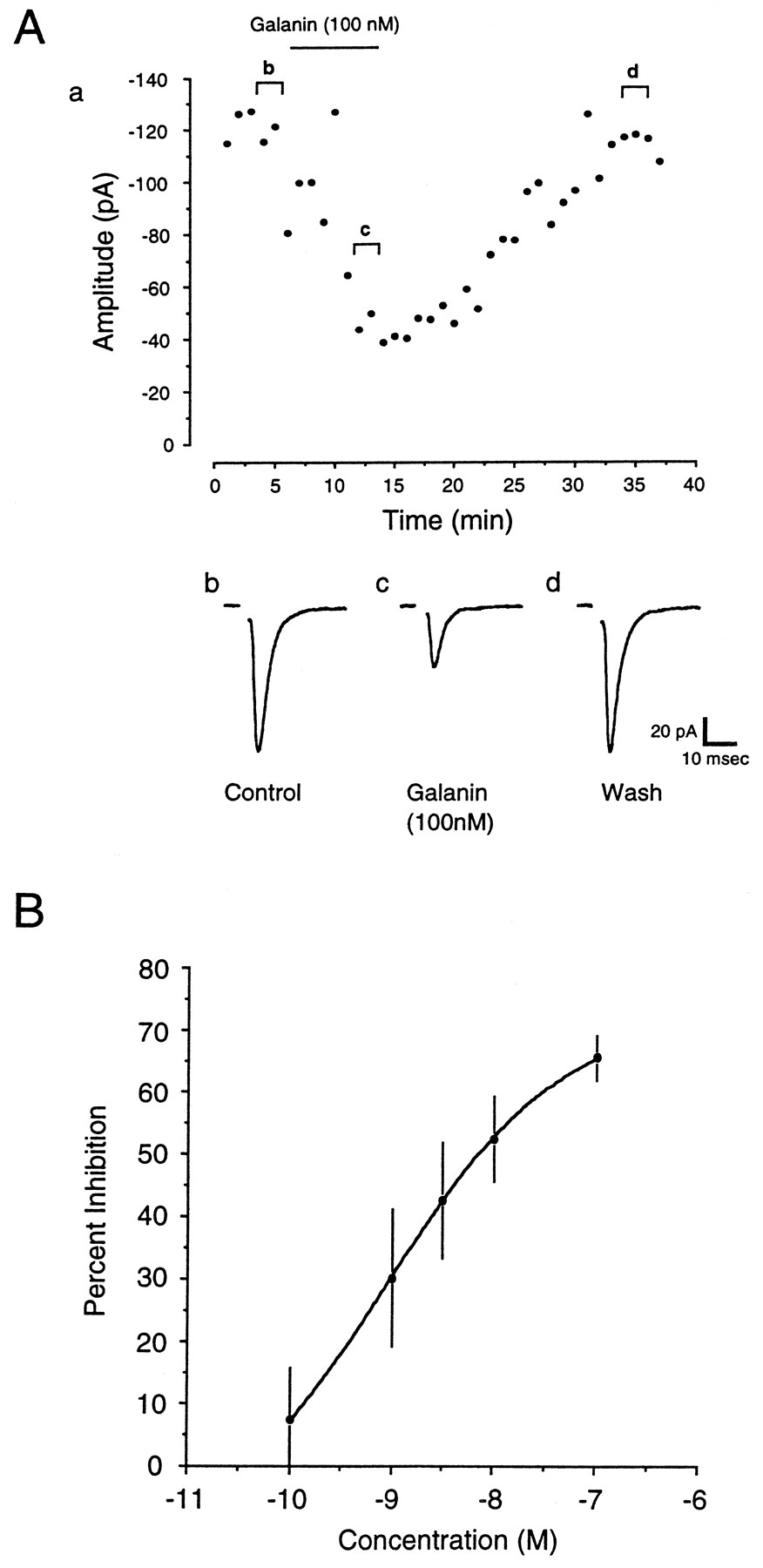
Application of galanin caused a depression of AMPA receptor-mediated synaptic transmission in the arcuate nucleus.A, Application of 100 nm galanin caused a depression in the evoked EPSC, as recorded from an arcuate nucleus neuron. After washout of the drug, the EPSC recovered to control levels within 20 min. For A, a represents the time course of the effects of galanin on synaptic transmission, whereasb–d are averaged EPSCs taken from the respective cells showing EPSCs before drug application (b), during drug application (c), and after washout of the drug (d). B, Dose–response curve for galanin. Application of progressively higher doses of galanin (0.1–100 nm) resulted in a dose-dependent decrease in the amplitude of evoked synaptic currents (n = 5), as recorded from arcuate nucleus neurons. Data are mean ± SEM.

Histogram representing the degree of inhibition of the evoked EPSC by the different galanin analogs. Vertical bars represent the absolute magnitude of inhibition produced by each of the compounds tested in this study, as marked below each bar. The numbers of observations for each compound are as follows (number of positive trials per total number of trials): galanin (19 of 26), galanin 1–16 (7 of 9), galanin 1–15 (4 of 10), galanin 3–29 (4 of 11), C7 (7 of 15), M40 (10 of 15), M15 (9 of 12), M32 (3 of 3). Data are mean ± SEM.
The dose–response relationship for the effect of galanin on synaptic transmission was investigated by bath applying the peptide in increasing doses to slices while evoked EPSCs were recorded. The concentration producing a maximal effect was determined to be ∼100 nm, with an EC50 of 0.85 nm(n = 5) (Fig. 1B).
To investigate the types of galanin receptors mediating inhibition of the EPSC, various galanin analogs were tested. Galanin (1–15) and (1–16) are N-terminal fragments of galanin, whereas galanin 3–29 is a galanin fragment truncated at the N terminus. Application of both galanin 1–15 (100 nm) and galanin 1–16 (100 nm) resulted in a depression of evoked EPSCs (Figs. 2,3A,B). Although both fragments caused a similar degree of inhibition (1–15: 56.56 ± 7.28%,n = 4; 1–16: 51.84 ± 7.60%, n = 7), galanin 1–16 acted on a larger group of cells (seven of nine) than galanin 1–15 (4/10). Strikingly, galanin 1–16 was effective on some cells in which galanin 1–15 had no effect (four of five cells) (Fig.3B). Galanin 3–29 (100 nm to 1 μm) was much less potent, causing a moderate inhibition in only 4 of 11 cells (21.28 ± 1.37%) (Figs. 2, 3C). In one cell on which galanin was also applied, the degree of inhibition mediated by galanin was much greater. In addition, galanin caused inhibition in each of the cells that did not respond to galanin 3–29 (Fig. 3C).

Application of galanin 1–15, 1–16, and 3–29 caused a depression of AMPA receptor-mediated synaptic transmission in the arcuate nucleus. A, Application of 100 nm galanin 1–15 caused a depression in the evoked EPSC, as recorded from an arcuate nucleus neuron. After washout of the drug, the EPSC partially recovered. Application of 100 nm galanin 1–16 after washout resulted in a nearly complete depression of the synaptic response. B, Application of 100 nmgalanin 1–15 to another cell failed to elicit a response. However, application of galanin 1–16 after washout resulted in a significant depression of transmission. C, In another cell, application of 100 nm galanin 3–29 produced only a small effect on transmission. After washout of this peptide fragment, application of 100 nm galanin resulted in a significantly larger depression of transmission. For A–C,a–e are the same as in Figure1A.
Various putative galanin antagonists have been reported (Bartfai et al., 1991, 1993; Lindskog et al., 1992; Pramanik and Ögren, 1992;Corwin et al., 1993; Crawley et al., 1993; Zini et al., 1993; Sahu et al., 1994). C7, M40, M15, and M32 all proved to have agonist effects in our experiments. Application of C7 (100 nm) caused a reduction (32.27 ± 3.52%) in synaptic transmission in 7 of 15 cases examined, as did M40 (100 nm) (35.8 ± 6.15%; 10 of 15), M15 (100 nm) (33.22 ± 4.47%; 9 of 12), and M32 (100–500 nm) (35.59 ± 9.34%;n = 3) (Fig. 2). Thus, in the majority of experiments, application of these “antagonist” peptides resulted in a depression of synaptic transmission, although the magnitude of the depression was less than that observed with galanin. This may indicate partial agonist activity or selective activation of limited subtypes of galanin receptors. In many cases, application of galanin after a response to one of the antagonist peptides resulted in a further depression of synaptic transmission. The application of galanin after C7 produced a 46.07 ± 9.12% depression (relative to C7; four of five) (Fig.4A), and similar effects were seen with M15 (39.88 ± 8.06%; four of six) (Fig.4B), and M40 (37.00 ± 17.09%; three of five) (data not shown).

C7 and M15 have partial agonist–antagonist actions on galanin receptors within the arcuate nucleus.A, Application of 100 nm C7 caused a reduction in synaptic transmission. Application of galanin (100 nm) resulted in a further reduction in transmission. During washout of C7, there was no further depression of transmission induced by galanin. B, Application of M15 to another cell also caused a moderate depression in synaptic transmission. Concurrent application of galanin resulted in a further depression of the response. C, Application of C7 had no effect on synaptic transmission. Galanin applied in the presence of C7 was also ineffective. After washout of C7, however, galanin caused a significant reduction in the synaptically evoked current. For A–C,a–e are the same as in Figure1A.
Finally, in a minority of experiments, there was convincing evidence for some antagonist activity associated with the putative galanin antagonists. In three of seven cases, washout of C7 while in the presence of galanin resulted in a reduction in the evoked current (43.33 ± 10.09%) (Fig. 4C). M15 and M40 were less active as antagonists, similarly antagonizing the actions of galanin in only one of three and one of five cases, respectively (36.1% and 18.5%) (data not shown). In some experiments, application of galanin in the presence of one of the antagonist peptides produced no additional effect. It is unclear, however, whether this was caused by occlusion or antagonism of the galanin response.
Mechanisms of action of galanin
A series of experiments was performed to determine the mechanism by which galanin produced synaptic inhibition in the arcuate nucleus. Activation of K+ channels in the presynaptic terminal could serve to shunt the incoming action potential, reducing Ca2+ channel activation and Ca2+influx, thereby reducing transmitter release (Segev, 1990; Gage, 1992;Graham and Redman, 1994; Miller, 1997). Although no outward currents were observed in any of the cells from which we recorded, it is possible that K+ channel activation in the presynaptic terminal might still occur. However, application of galanin in the presence of 100 μm Ba2+, which blocks G-protein-activated K+ channels (Ransom and Sontheimer, 1995; Wang and McKinnon, 1996; Rhim et al., 1997), still depressed synaptic transmission to the same extent (n = 4; data not shown), indicating that K+ channel activation was not involved in the actions of galanin in this case. The effects of galanin were also examined using a paired-pulse stimulus protocol (Baskys and Malenka, 1991). Application of galanin (100 nm) was found to depress the initial evoked EPSC, P1, in 6 of 11 cells examined (57.5 ± 4.7%; n = 6) (Fig.6A). In cases in which a depression of the initial EPSC was observed, the paired-pulse ratio was strongly increased in galanin (P2/P1: 1.27 ± 0.17;n = 6) versus control (P2/P1: 0.61 ± 0.05;n = 6) (Fig. 5). In contrast, in the cells in which no inhibition of the initial EPSC was observed, the paired-pulse ratio in galanin (100 nm) (P2/P1: 1.10 ± 0.25) was not significantly different from the control period (1.01 ± 0.07;p = 0.740, unpaired t test;n = 5; data not shown). Finally, direct postsynaptic modulation of the receptor was examined using a series of short bath applications of AMPA (25 μm for 40 sec). Simultaneous application of galanin had no effect on the whole-cell currents recorded under these conditions (107.7 ± 6.5% of control;n = 6) (data not shown). These effects of galanin support a presynaptic mechanism of action.

Application of galanin reduced paired-pulse depression in the arcuate nucleus. A, Application of 100 nm galanin produced a reversible decrease in the amplitude of the initial first evoked EPSC, P1, and an increase in the paired-pulse ratio P2/P1. A,a represents the time course of the effects of galanin on the ratio of the amplitude of the evoked EPSCs, P2/P1. ForA, b–d are the same as in Figure1A. Interpulse interval was 30 msec.B, Histogram representing the effect of galanin (100 nm) on the mean paired-pulse depression from six cells. The pooled data show a significant increase in the paired-pulse ratio (p = 0.01; unpaired ttest).
Mechanisms underlying modulation of synaptic transmission can also be studied by recording the spontaneous release of vesicles from the presynaptic terminal. Alterations in the frequency or amplitude distribution of miniature events are indicative of a presynaptic or postsynaptic site of action, respectively. mEPSCs were recorded from arcuate nucleus neurons in the presence of 7-chlorokynurenic acid (40 μm) or d,l-AP5 (100 μm), bicuculline (10 μm), TTX (1 μm), and Cd2+ (100 μm) to isolate Ca2+ entry-independent spontaneous AMPA receptor-mediated events. To confirm that Cd2+ was adequately blocking Ca2+ entry through voltage-gated Ca2+ channels, 100 μmCd2+ was applied to the preparation while evoked EPSCs were recorded. In four of four cases, application of 100 μm Cd2+ completely abolished synaptic transmission (data not shown). Under these conditions, spontaneous mEPSCs were observed in approximately one of four cells recorded from at a frequency adequate for analysis (>0.5 Hz). Application of galanin (100 nm) resulted in a 54.07 ± 3.45% reduction in the frequency of recorded mEPSCs but had no effect on the amplitude distribution (95.83 ± 2.06%) (Fig.6A–F) (n = 7).
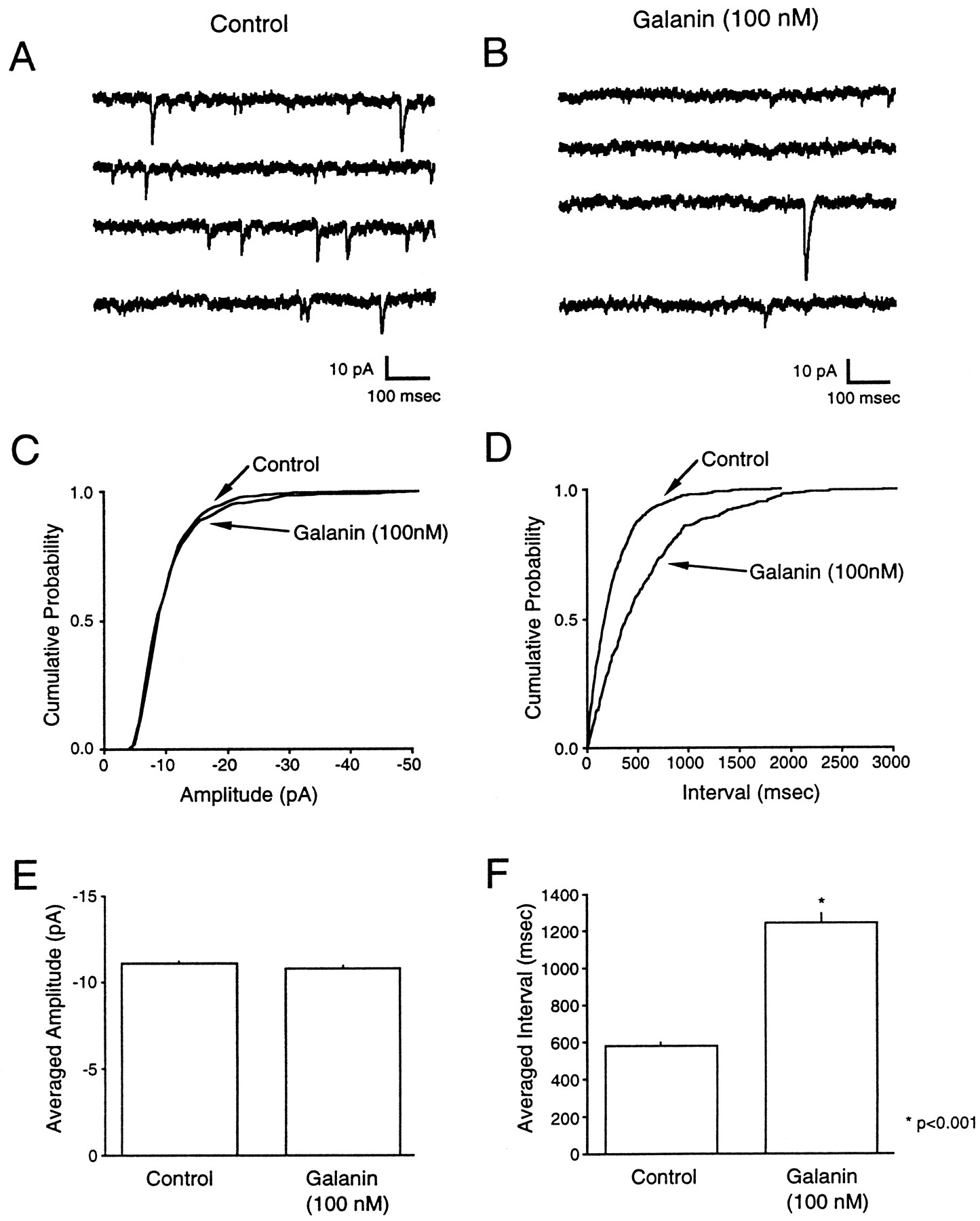
Galanin caused a reduction in the frequency of mEPSCs with no effect on the amplitude distribution. A, B, Samples of mEPSCs recorded from arcuate nucleus neurons. In both A and B, traces represent a continuous recording period of ∼3 sec for control (A) and in the presence of 100 nmgalanin (B). C, D, Cumulative probability plots of the distribution of mEPSC amplitudes (C) and intervals (D) for a single cell, showing a significant change in the interval distribution (p < 0.0001; unpaired t test with Welch’s correction). E, F, Effect of galanin on the mean amplitude (E) and mean interval (F) for data pooled from six cells. The pooled data showed a significant change in the interval (p < 0.0001; unpaired t test with Welch’s correction), whereas no significant change in the mean amplitude was observed. All data were acquired in the presence of 1 μm TTX, 40 μm 7-chlorokynurenic acid, 10 μm bicuculline, and 100 μmCd2+.
α-latrotoxin binds to at least two types of presynaptic receptors and enhances transmitter release in both a Ca2+-dependent and -independent manner (Misler and Hurlbut, 1979; Grasso et al., 1980; Nicholls et al., 1982; Meldolesi et al., 1984; Rosenthal et al., 1990; Capogna et al., 1996a; Krasnoperov et al., 1997). When galanin was applied to slices that had been incubated in 2 nm α-latrotoxin, there was a 47.76 ± 6.33% reduction in the frequency of mEPSCs (Fig.7A–F) (n = 7). The amplitude distribution remained unchanged (97.78 ± 2.70% of control; n = 7) (Fig.7A–F). These results suggest that galanin can inhibit transmitter release at a site “downstream” of Ca2+ entry through voltage-dependent Ca2+ channels and may act at a site very close to the final step in the release process.

Galanin caused a reduction in the frequency of mEPSCs with no effect on the amplitude distribution in slices exposed to α-latrotoxin. A, B, Samples of mEPSCs recorded from arcuate nucleus neurons in slices exposed to α-latrotoxin for >2 hr. In both A and B, traces represent a continuous recording period of ∼4 sec total for control (A) and in the presence of 100 nmgalanin (B). C, D, Cumulative probability plots of the distribution of mEPSC amplitudes (A) and intervals (B) for a single cell, showing a significant change in the interval distribution (p < 0.0001; unpaired t test with Welch’s correction). C, D, Effect of galanin on the mean amplitude (C) and mean interval (D) for data pooled from seven cells. The pooled data showed a significant change in the interval (p < 0.0001; unpaired t test with Welch’s correction), whereas no significant change in the mean amplitude was observed. All data were acquired in the presence of 1 μm TTX, 100 μmd,l-AP5, 10 μm bicuculline, and 100 μmCd2+ in slices exposed to 2 nmα-latrotoxin for 2 hr before recording.
DISCUSSION
Galanin acts in the hypothalamus to stimulate feeding behavior (Crawley et al., 1990; Kyrkouli et al., 1990; Leibowitz and Kim, 1992;Corwin et al., 1993) and to regulate hormone secretion (Bauer et al., 1986; Bjorkstrand et al., 1993; Kondo et al., 1993; Landry et al., 1995) and the reproductive cycle (Grafstein-Dunn et al., 1994; Sahu et al., 1994; Rossmanith et al., 1996). In addition, galanin has been shown to exert numerous effects elsewhere in the brain, including effects on learning and memory (Sundström et al., 1988;Ögren and Pramanik, 1991; Crawley et al., 1993).
How does galanin produce its effects? Cloning studies have indicated that the actions of galanin are mediated through at least two subtypes of galanin receptors, GALR1 and GALR2 (Habert-Ortoli et al., 1994;Howard et al., 1997). In addition, the existence of other galanin receptors has been inferred from binding and functional studies (Wynick et al., 1993; Gu et al., 1995; Valkna et al., 1995). The effects of galanin we have observed would be expected to involve at least two different receptors. Both GALR1 and GALR2 have been shown to be present within the hypothalamus (Habert-Ortoli et al., 1994; Gustafson et al., 1996; Howard et al., 1997; Wang et al., 1997) and might be responsible for much of the activity observed here using galanin agonists. Furthermore, these receptors also interact with the putative galanin antagonists we have used. The majority of our observations are consistent with the idea that these peptides act as partial agonists, as has been found in several other investigations (Fridolf and Ahrén, 1993; Mulvaney et al., 1995; Papas and Bourque, 1997). Additionally, we have made at least two other interesting observations. The first of these concerns the activity of galanin 3–29. This analog does not interact significantly with either GALR1 or GALR2 (Habert-Ortoli et al., 1994; Parker et al., 1995; Howard et al., 1997;Sullivan et al., 1997). However, Wynick et al. (1993) reported that the anterior pituitary and hypothalamus do contain a receptor that is activated by N-terminal truncated analogs such as galanin 3–29. The modest effects we have observed may reflect actions at such a receptor. Second, it is striking that we have observed groups of cells that responded to both galanin 1–16 and 1–15 or only to the former peptide. It is clear that GALR1 and GALR2 both respond to galanin 1–16 (Habert-Ortoli et al., 1994; Parker et al., 1995; Howard et al., 1997;Sullivan et al., 1997). However, previous studies have generally indicated that the potency of galanin analogs decreases substantially after the removal of the amino acid in the 16th position (Lagny-Pourmir et al., 1989; Land et al., 1991). Thus, the robust activity of galanin 1–15 observed in some cells is somewhat surprising. However, there is a report of a galanin 1–15 selective receptor in the hippocampus (Hedlund et al., 1994), and intracerebroventricular injection of galanin 1–15 but not 1–29 modulates baroreflex sensitivity in the rat (Diaz et al., 1996). Thus, it is conceivable that this receptor also occurs in the hypothalamus.
Further evidence for the presence multiple subtypes of galanin receptors in the arcuate nucleus was obtained in experiments with the chimeric peptides M40, M15, M32, and C7. In the majority of cases, these compounds behaved as agonists. However, the degree of inhibition was notably less than that caused by galanin alone. This may reflect partial agonist actions of these compounds. Alternatively, galanin may act simultaneously at several different subtypes of receptors, whereas the chimeric peptides may be a more selective group of full agonists. Antagonism by the chimeric peptides was observed only in a minority of cases examined. This contrasts with other reports on the actions of these peptides in which they have been shown to block galanin-induced feeding behavior (Leibowitz and Kim, 1992; Corwin et al., 1993; Crawley et al., 1993), stimulation of ACh release (Pramanik and Ögren, 1993), and the actions of galanin in the hippocampus, locus coeruleus, and spinal cord (Bartfai et al., 1991, 1993). However, other reports have suggested partial or full agonist actions of these compounds, including on the mobilization of intracellular Ca2+stores (Fridolf and Ahrén, 1993), the activation of K+ currents in magnocellular neurosecretory cells (Papas and Bourque, 1997), and in the inhibition of voltage-dependent Ba2+ currents (Mulvaney et al., 1995). It appears then that the actions of these compounds are not restricted to antagonist effects and that they may even prove to be selective agonists at specific subtypes of galanin receptors. In summary, therefore, our results suggest the existence of multiple types of galanin receptors within the arcuate nucleus. Nevertheless, it is possible that some of the galanin analogs that we are using interact with other receptor populations as well. Additional pharmacological characterization is required before any definitive statements can be made regarding the actions of these compounds.
The actions of galanin on fast glutamatergic transmission observed in this study are analogous to those shown in other reports, such as its effects on slow cholinergic transmission in the hippocampus (Dutar et al., 1989). However, no studies have investigated directly the mechanisms underlying such actions. Galanin inhibited evokedl-glutamate release in the hippocampus, an effect that was blocked by glibenclamide, an ATP-sensitive K+channel blocker (Zini et al., 1993). In many other systems, galanin activates a K+ conductance and hyperpolarizes neurons (Ahrén et al., 1989; Dunne et al., 1989; Konopka et al., 1989; Seutin et al., 1989; Papas and Bourque, 1997), an effect that could produce inhibition of transmitter release (Segev, 1990; Gage, 1992; Graham and Redman, 1994; Miller, 1997). However, in our studies no outward currents were observed with galanin. Furthermore, application of 100 μm Ba2+ had no effect on galanin receptor-mediated synaptic depression (Scanziani et al., 1995; Miller, 1997).
There are also reports of galanin inhibiting voltage-gated Ca2+ channels in neurons (Merriam and Parsons, 1995;Mulvaney et al., 1995), and such an action could contribute to the effects on synaptic transmission that we have observed. We have clearly demonstrated that galanin acts presynaptically to directly decrease transmitter release independent of Ca2+ entry through voltage-sensitive Ca2+ channels. This mechanism of presynaptic inhibition has now been widely reported in slices and in culture (Scholz and Miller, 1992; Scanziani et al., 1995;Tyler and Lovinger, 1995; Trudeau et al., 1996; Dittman et al., 1996;), although its underlying molecular mechanisms are unclear. It is conceivable, however, that the mechanism of release for mEPSCs versus evoked currents is different, and consequently the mechanism of action of galanin may not be the same for these two processes (but see Capogna et al., 1996a). Interestingly, the magnitude of the effect of galanin on mEPSCs in the presence of Cd2+ (54.07 ± 3.45% reduction in frequency) was similar to the effect of galanin on synaptic transmission (55.54 ± 4.03% reduction in amplitude), indicative of a common mechanism. Significant, however, was the observation that galanin decreased the magnitude of paired-pulse depression, often reversing it to paired-pulse facilitation, indicative of a presynaptic action on evoked release (Baskys and Malenka, 1991). It is still possible, however, that galanin is acting on a Ca2+-dependent mechanism. Residual Ca2+ inside the cell, or Ca2+leak into the cell, might contribute to mEPSC frequency even in the presence of Cd2+. However, α-latrotoxin acts at the presynaptic terminal to induce a partially Ca2+-independent release of transmitter (Misler and Hurlbut, 1979; Meldolesi et al., 1984; Rosenthal et al., 1990; Capogna et al., 1996a; Krasnoperov et al., 1997). Exposure of the slices to α-latrotoxin would activate this mechanism, and thus a significant portion of the resultant mEPSCs would be Ca2+-independent. Under these conditions, application of galanin resulted in inhibition of mEPSCs of a magnitude comparable to that observed in the absence of α-latrotoxin (47.76 ± 6.33% reduction in frequency). These results are similar to those reported for the actions of GABA in the hippocampus in the presence of α-latrotoxin (Capogna et al., 1996a) and suggest that galanin has a powerful depressant effect on the release process at a site close to the final vesicle-release step. There are numerous potential sites within the presynaptic machinery at which it could act.Krasnoperov et al. (1997) reported recently that α-latrotoxin stimulates Ca2+-independent exocytosis through its interaction with a G-protein-coupled receptor (the CIRL-receptor), and that this receptor copurifies with syntaxin. Furthermore, Linial et al. (1997) reported that presynaptic muscarinic receptors could also directly interact with syntaxin. Thus, the galanin receptor might interact with, or signal to, a site within the release machinery and so directly reduce exocytosis.
Footnotes
This work was supported by Public Health Service Grants DA02121, DA02575, MH40165, NS33502, DK42086, and DK44840. We thank Drs. Daniel McGehee and Aaron Fox for their technical expertise.
Correspondence should be addressed to Dr. Richard J. Miller, Department of Pharmacological and Physiological Sciences, 947 E. 58th Street, MC 0926, Chicago, IL 60637.
Dr. Kinney’s present address: Department of Physiology and Biophysics, Veterans Affairs Medical Center, Seattle, WA 98108-1597.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}