

Abstract
Telencephalic organization in birds is so unusual that many homologies between avian and mammalian telencephalic areas remain controversial. Particularly contested is the avian “neostriatum,” which has historically been homologized to either mammalian striatum, lateral neocortex, or endopiriform claustrum. Because homologies between these adult structures have been so difficult to resolve, we have begun to examine how telencephalic development diverges between birds and other vertebrates. To this end, biotinylated dextran was injected into the lateral telencephalon of chick embryos at 3 d of incubation, and the distribution of labeled cells was examined up to 14 d later. The data show that a definite boundary to cellular migration develops just ventral to the neostriatum between 5 and 8 d of incubation. Labeled polyclones within the neostriatum stretch from the ventricular zone to the brain surface and exhibit an increasingly rostrocaudal orientation as development proceeds. Individual polyclones contribute cells to several of the distinct auditory, visual, somatosensory, and olfactory regions within the neostriatum. A comparative analysis suggests that the avian neostriatum develops from a precursor region that in other vertebrates gives rise to olfactory cortex and, when present, to other components of the piriform lobe, such as the endopiriform claustrum and basolateral amygdala. Conclusions about lateral pallial homologies between birds and mammals remain uncertain, however, primarily because so little is known about the development of the lateral pallium in mammals. This lacuna might be filled by applying to mammals the novel fate-mapping method described in the present paper.
The avian telencephalon is characterized by a prominent intraventricular ridge, a major portion of which is referred to as “neostriatum” because it was once believed to be homologous to the mammalian caudate and putamen (Ariëns Kappers et al., 1936). This hypothesis became untenable when modern histochemical methods revealed that only an area immediately ventral to the neostriatum stained heavily for several markers typically associated with mammalian striatum (Juorio and Vogt, 1967; Karten, 1969). When modern connectional studies then revealed that the neostriatum receives major ascending sensory inputs from the dorsal thalamus, the neostriatum became widely regarded as homologous to part of mammalian neocortex (Karten, 1969; but see Lohman and Smeets, 1990). Classical embryological studies supported the hypothesis that the neostriatum develops as part of the telencephalic roof, or pallium (Kuhlenbeck, 1938; Källén, 1962), but also suggested that it develops as a thickening of the most lateral pallial wall, which gives rise to piriform cortex in most vertebrates and may give rise also to the endopiriform claustrum and the basolateral amygdala in mammals (Holmgren, 1922, 1925). Developmental data have therefore been used to argue that the neostriatum is homologous to part of what is sometimes referred to as the piriform lobe in mammals (Striedter, 1997). Little is known about the embryology of the lateral telencephalon in both birds and mammals, however, and the available data are rife with contradictions.
Thus, some studies in mammals demonstrated a distinct boundary of gene expression and cell migration just dorsolateral to the lateral ganglionic eminence (LGE), which gives rise to the striatum (Bulfone et al., 1993; Fishell et al., 1993; Krushell et al., 1993). Another study showed, however, that at least some cells migrate from the LGE into neocortex (Anderson et al., 1997). Furthermore, mammalian piriform cortex has been claimed to develop from the lateral pallial wall, the LGE, or both (Smart, 1985; Bayer and Altman, 1991; De Carlos et al., 1996). Similarly confusing are the data for birds, in which tritiated-thymidine data suggested the presence of a distinct pallial–subpallial boundary (Tsai et al., 1981a,b), whereas chimeric transplantation indicated extensive cell mixing between dorsal and ventral regions of the telencephalon (Balaban et al., 1988). Retroviral lineage tracing, finally, revealed that neurons in both birds and mammals may migrate orthogonally to the radial glia, which calls into question any developmental scenarios based on radial glial data (Walsh and Cepko, 1992; Kornack and Rakic, 1995; Rakic, 1995; Tan et al., 1995; Szele and Cepko, 1996; Striedter and Beydler, 1997). Whether these data are really contradictory is unclear, however, because the various studies relied on very different, and often highly indirect, methods of investigation.
In the present study, we are able to clarify several aspects of avian telencephalic development by injecting biotinylated dextrans into a small patch of the telencephalic ventricular zone (VZ) and then following the fate of labeled cells for up to 2 weeks, when an adult-like pattern of telencephalic organization has become established (Striedter and Beydler, 1997). Biotinylated tracers should generally be more sensitive than their fluorescently labeled counterparts because biotinylation enables the use of powerful signal amplification techniques (e.g., Ding and Elberger, 1995), and this increased sensitivity should ameliorate the problem of tracer dilution with cell division that is inherent in many lineage tracing studies. In accordance with this expectation, the use of biotinylated dextrans enabled us to detect labeled cells at significantly later developmental stages than had been possible in a similar previous study that used fluorescent lineage tracers (Montgomery, 1996).
Our data indicate that (1) the neostriatum develops just dorsal to a cell migration boundary that is probably homologous to the pallial–subpallial boundary in other vertebrates and (2) the various subdivisions of the “neostriatal” complex develop from overlapping portions of the VZ. From a combined evolutionary and developmental perspective, these data suggest that the avian neostriatum develops from the embryonic lateral pallium, which in other vertebrates probably gives rise not to neocortex but to olfactory cortex and, when present, additional components of the piriform lobe.
MATERIALS AND METHODS
Fertile eggs of White Leghorn chickens were incubated and windowed after ∼3 d (72 hr) of incubation, which corresponds to developmental stages 17–18 as described by Hamburger and Hamilton (1951). These ages were chosen for injections because the chick telencephalon at 3 d of incubation (1) is as large as it will be before it is covered, and made relatively inaccessible, by the chorioallantois and (2) consists only of a pseudostratified neurepithelium covered by a layer of mesenchyme (Tsai, 1977). Postmitotic neurons first appear in the telencephalon on the fourth day of incubation and then form an incipient mantle layer just superficial to the ventricular zone (Tsai, 1977).
All embryos (n = 187) were exposed by making a slit in the vitelline membrane, through which one or two drops of 0.001% fast green in ampicillin (50 mg/ml) were applied to enhance visualization of the embryo. Glass micropipettes were pulled on a horizontal pipette puller to an internal tip diameter of 1.5 μm. These electrodes were sharp enough to penetrate the mesenchyme but blunt enough to resist penetration into the ventricular lumen. The pipettes were backfilled with 5% biotinylated dextran (10,000 molecular weight; Molecular Probes, Eugene, OR) in 1.1 m KCl and 0.05% Triton X-100 and advanced into the tissue overlying the lateral telencephalon with a micromanipulator. The depth of penetration was difficult to judge because of tissue dimpling and resilience but probably ranged from 200 to 400 μm. Visualization of the pipette tip was facilitated by dipping filled pipettes into an aqueous suspension of drawing ink. Tracer was ejected with +10 μA of current for 10 min (5 min for some of our first injections). After injection, the eggs were closed with adhesive tape and returned to the incubator for up to 14 more days.
The location of each injection site was mapped onto standardized drawings of embryos at the same stage of development (Fig.1). The accuracy of this mapping is limited by slight differences in telencephalic development between embryos, by the paucity of definite landmarks available for the alignment of telencephala across embryos, and by the fact that electrode tips could not be visualized directly once they had penetrated into the mesenchyme. Although our map of injection sites is therefore limited in precision, the resultant fate map (Fig. 1) is in good general agreement with the more comprehensive fate map reported byMontgomery (1996) for slightly younger chick embryos. Without reference to such diagrammatic fate maps, it is difficult to verbalize where an injection site is located because the telencephalon at these early ages is not visibly divided into distinct regions and even the course of the rostrocaudal axis is, because of the bending of the neural tube, a subject of controversy (Puelles and Rubenstein, 1993; Nieuwenhuys et al., 1998). We therefore refer to injections into regions that eventually give rise to the neostriatum as injections into the “presumptive neostriatum.” Injections into regions that give rise to the paleostriatum, which is widely acknowledged to be the avian homolog of mammalian striatum (Reiner et al., 1984; Medina and Reiner, 1995; Veenman et al., 1995), are similarly referred to as injections into the “presumptive paleostriatum.”

Schematic lateral view of the anterior portion of the head in a chick embryo at 3 d of incubation. Injections sites are indicated by circles, and the subsequently observed pattern of labeled cells is coded by the shading patternas indicated in the box legend. The orientation of the “true” rostrocaudal axis in 3 d embryos remains controversial (see text), but injection sites toward the right edge of the diagram label more rostral structures at the later, adult-like, stages than do injection sites toward the left edge of the diagram. HV, Ventral hyperstriatum;Paleo., paleostriatum; Tel, telencephalon.
At the end of the incubation period, embryos were cooled on ice and fixed by immersion or transcardial perfusion with 2% paraformaldehyde and 0.2% glutaraldehyde in 0.1 m phosphate buffer (PB). After 6–20 hr of post-fixation, the brains were dissected out and cryoprotected in 30% sucrose in 0.1 m phosphate buffer. Brains were embedded in 10% gelatin and 30% sucrose, fixed overnight, and sectioned frozen at 40 μm. Free-floating sections were pretreated for 2 hr in 0.3% H2O2 in PB to bleach out the remaining blood and endogenous peroxidases. They were then incubated for 2 hr in a Vectastain ABC Elite standard kit solution (Vector Laboratories, Burlingame, CA) and reacted with a tetramethylbenzidine-sodium tungstate procedure (Ding and Elberger, 1995), followed by a nickel and cobalt-intensified diaminobenzidine reaction (Adams, 1981). Mounted sections were stained with Giemsa dye at room temperature (Iñiguez et al., 1985). Three-day-old embryos were bisected, reacted as free-floating whole mounts, embedded, sectioned, and stained using the methods described above.
Because our injections are extracellular and therefore label multiple precursors cells, labeled cells at subsequent stages constitute polyclones. The percentage of injections that resulted in labeled polyclones varied widely (0–80%), as details of the injection procedure were varied to improve the method. The present paper is based on a detailed analysis of 44 successful cases, which were fixed 1–3 hr after injection (n = 8) or at 5 d (n = 4), 8 d (n = 3), 10 d (n = 7), 12 d (n = 7), 14 d (n = 9), or 17 d (n = 6) of embryonic age. Labeled cells were identified through the microscope and mapped onto a low-resolution video image of the section by means of stage transducers and commercial software (Transtek, San Diego, CA). Cells were classified as labeled only if they were stained solidly black and/or exhibited dendritic processes.
Because it was important to know whether labeled polyclones extend into piriform cortex, which in birds cannot be identified solely from cytoarchitecture (Reiner and Karten, 1985), olfactory bulb projections were examined in several embryos. Chick embryos were perfused with 4% buffered paraformaldehyde at 10 d (n = 5), 12 d (n = 4), 14 d (n = 2), and 17 d (n = 1) of incubation. After several days of post-fixation, skull and meninges over the olfactory bulbs were removed, and small crystals of 1,1′-dioctadecyl-3,3,3′,3′-tetramethylindocarbocyanine perchlorate (DiI; Molecular Probes) were pushed into one of the olfactory bulbs. Application sites were covered with agar to prevent the crystals from dislodging, and brains were stored at 40°C for 4–6 weeks to let the dye diffuse along the axon membranes. Brains were then dissected out, embedded in gelatin–sucrose, fixed again, and cut at 50 μm on a vibratome. The sections were examined under epifluorescent illumination.
RESULTS
In embryos killed shortly after tracer injection (i.e., at 3 d of incubation) and reacted as whole mounts, labeled cells were invariably observed in the mesenchyme overlying the telencephalon (Fig.2A), but labeled VZ cells were observed in only two of the eight cases (Fig.2B). Moreover, individual VZ cells varied in the degree to which they were labeled. These data indicate that our injections generally involved leakage of tracer into the mesenchyme. Some of the labeled mesenchymal cells probably give rise to the vascular endothelial cells that were sometimes observed (Fig.3A) in the lateral telencephalon, but there is no evidence to indicate that mesenchymal precursors give rise to any telencephalic neurons (see Noden, 1987, on other fates of cephalic mesenchyme). The data also indicate that our injection sites had ill-defined boundaries and varied in size. Analysis of embryos at 3–8 d of incubation (Figs. 2, 3) does indicate, however, that labeled VZ cells were always tightly clustered together and that our injection sites were probably <50 μm in diameter. Control injections into the ventricular lumen (n = 6) produced no labeled VZ cells, confirming that accidental spillage of tracer into the ventricle could not have labeled VZ cells far away from the injection site. Finally, because different cells within an injection site take up varying amounts of tracer, the relative labeling intensity of cells at later ages cannot be used as a direct indicator of when the labeled cells became postmitotic. However, the size and degree of differentiation of labeled neurons probably are at least approximate indirect indicators of cellular birth date (see below).

A, B, Dextran injection into a 3 d embryo reacted as a whole mount (A) and sectioned (B) to show labeled cells in the ventricular zone (VZ) and overlying mesenchyme (Mes) is shown. Tel, Telencephalon. C, D, By 5 d, a mantle zone has developed, containing radial glial processes and young neurons, some of which have migrated ventrally (arrows).Mtl, Mantle layer. Scale bars: A, 100 μm; B–D, 50 μm.

At 8 d of incubation, labeled polyclones extend in a clearly radial manner from the ventricular zone (VZ in A) through the neostriatum (N) to the brain surface (arrowsin B). Asterisks indicate labeled endothelial cells. Scale bar: A, B, 50 μm.
At 5 d of incubation, numerous cells have migrated out of the VZ and form an incipient mantle layer (Fig. 2C,D). Labeled cells within the VZ form radial columns and exhibit little tangential dispersion. Labeled cells in the mantle layer are primarily superficial to the labeled VZ patch and commingle with numerous labeled processes that extend from the VZ to the brain surface and probably emanate from radial glial cells. In several cases, tangentially migrated cells were observed in the mantle zone ventral to the injection site (Fig. 2C,D,arrows).
At 8 d, the VZ has become dramatically reduced in thickness, and labeled polyclones extend from the VZ to the brain surface in a clearly radial manner (Fig. 3), although their long axes are slightly tilted ventrally and rostrally away from the VZ origin. Labeled cells near the brain surface are generally larger, possess more and longer labeled processes, and exhibit greater tangential dispersion than do cells near the VZ. The dorsal medullary lamina (LMD) (Fig.4) becomes apparent at this age, separating the developing neostriatum from the paleostriatum. Injections into the presumptive neostriatum (Fig. 1; see Materials and Methods) labeled cells only dorsal to the LMD, whereas injections into the presumptive paleostriatum labeled cells only ventral to the LMD.

Two 10 d cases resulting from injections into the presumptive neostriatum in which caudalmost (left) and rostralmost (right) sections are separated by 1440 μm (A) and 1200 μm (B). Labeled cells remain dorsal to the dorsal medullary lamina (LMD) boundary except in B, where some labeled cells are found in the superficial paleostriatum.HV, Ventral hyperstriatum; LH, hyperstriatal lamina; N, neostriatum; P, paleostriatum; W, Wulst. Scale bars, 1 mm.
At 10 d, the telencephalon has thickened further and the hyperstriatal lamina (LH), separating the neostriatum from the ventral hyperstriatum (HV), can be identified (Fig. 4). Labeled polyclones again extend from the VZ to the brain surface, but the rostrocaudal extent of these polyclones has increased (from <30 to >35% of telencephalic length) as the neostriatal VZ was displaced caudally relative to the anterolateral brain surface. The polyclones also exhibit greater tangential dispersion than they did at earlier ages, particularly within the anterior neostriatum, where some cells appear to be migrating dorsally toward HV (Fig. 4B). One polyclone at this age, resulting from an injection into the caudal presumptive neostriatum, extends from the ventral hyperstriatum across LH into the neostriatum, which indicates that LH does not represent a lineage restriction boundary at least at this caudal level of the telencephalon. The LMD, in contrast, does appear to be a lineage boundary, because injection sites into the presumptive neostriatum consistently label cells only dorsal to the LMD (Figs. 4,5A,B). The only exception to this rule is that a few cells (0–1.4%) are labeled within the most superficial paleostriatum (Fig. 5C). Even in cases in which the injection sites were in the boundary region between presumptive neostriatum and presumptive paleostriatum (Fig. 1;n = 2 at this age), large clusters of labeled cells in the neostriatum are pushed up against the LMD in a manner suggesting that they encountered a barrier to cell migration. An injection into the presumptive paleostriatum labeled cells only ventral to the LMD (Fig. 5D). In all cases, labeled cells near the brain surface tend to be larger and better differentiated than are neurons near the VZ, and axonal processes emanate from some neurons in the anterolateral neostriatum (Fig. 5C).

At 10 d of incubation, a polyclone extends from the neostriatal ventricular zone (A), through the neostriatum (N) just dorsal to the dorsal medullary lamina (LMD) (B), to the most superficial neostriatum (C). A labeled cell is also apparent in the most superficial paleostriatum (arrow inC). An injection into the presumptive paleostriatum, in contrast, labels cells only ventral to theLMD (D). P, Paleostriatum. Scale bars: A–C, 50 μm;D, 100 μm.
Between 12 and 17 d, an adult-like pattern of telencephalic organization becomes apparent as subdivisions of the neostriatum can be discerned in Nissl-stained sections. Labeled polyclones at these later ages are generally similar to those at earlier stages but can now be observed to extend through several of the neostriatal subdivisions, including the auditory field L, the visual ectostriatum, the somatosensory nucleus basalis, and the frontoarchistriatal tract at the brain surface (Figs. 6,7). Data from these later ages also confirm that injections into the caudal presumptive neostriatum labeled polyclones that extend not only through the neostriatum but also across LH into the ventral hyperstriatum (n = 4; Figs. 1, 6). The ventral hyperstriatum therefore develops at least in part from the same precursor region that also gives rise to much of the neostriatum. Tangential dispersion is pronounced within the anterior neostriatum and near field L. As observed in the younger cases, injections into the presumptive neostriatum labeled cells only dorsal to the LMD, except in some cases in which a few cells were labeled also in the superficial paleostriatum. Labeled axons of neurons in the anterior neostriatum projected to the overlying ventral hyperstriatum (Fig. 7C), the underlying paleostriatum, and the archistriatum. By 14 d of incubation, the neostriatal VZ was in most places reduced to a thin layer of possibly nonmitotic cells, making it more difficult to identify labeled injection sites, but labeled polyclones were similar to those observed earlier (Fig. 8).

A, One 12 d case resulting from an injection into the caudal presumptive neostriatum in which caudalmost (left) and rostralmost (right) sections are separated by 1440 μm. B, A similar 14 d case with caudalmost and rostralmost sections separated by 1200 μm. B, Nucleus basalis; E, ectostriatum; FA, frontoarchistriatal tract;HV, ventral hyperstriatum; L, field L;LH, hyperstriatal lamina; LMD, dorsal medullary lamina; N, neostriatum; P, paleostriatum; W, Wulst. Scale bars, 1 mm.

At 12 d of incubation, a labeled polyclone resulting from an injection into the caudal presumptive neostriatum labels cells in the ventricular zone adjacent to the ventral hyperstriatum (HV) but extends into field L (L) (A) and to the superficial neostriatum (B). Labeled neurons in the anterior neostriatum have axons that terminate in HV(C) and extend into the frontoarchistriatal tract (FA) (D), wherein they course toward the archistriatum. LH, Hyperstriatal lamina;LMD, dorsal medullary lamina; N, neostriatum; P, paleostriatum. Scale bar:A–D, 50 μm.

A, By 14 d of incubation, the neostriatal ventricular zone is nearly exhausted (between thearrows). B, Labeled cells at this age abut, but do not cross, the dorsal medullary lamina (LMD). CP, Choroid plexus;Hip, hippocampal formation; N, neostriatum; P, paleostriatum. Scale bars, 50 μm.
Injections into the presumptive paleostriatum between 12 and 17 d of incubation (n = 4) labeled cells almost exclusively ventral to the LMD (Fig. 9). A few labeled cells were seen within the LMD or just dorsal to it, but the great majority of cells was clearly ventral to the LMD. Labeled cells near the brain surface were generally larger and more highly differentiated than were those near the VZ (Fig. 5D). Labeled polyclones were widely dispersed throughout the paleostriatum, including most of the augmented paleostriatum and some portions of the primitive paleostriatum (see Fig. 9), but our data do not permit a detailed analysis of paleostriatal development.

An injection into the presumptive paleostriatum yielded labeled cells (small filled circles) throughout most of the paleostriatum (PA andPP) in a case that was fixed at 14 d of incubation. Sections are arranged from caudal to rostral in aclockwise manner. A, Archistriatum;HV, ventral hyperstriatum; N, neostriatum; PA, augmented paleostriatum;PP, primitive paleostriatum; W, Wulst. Scale bar, 1 mm.
Application of DiI to the olfactory bulb yielded results consistent with previous data from adult pigeons (Reiner and Karten, 1985) and therefore not described in detail. Briefly, labeled axons course caudally from the olfactory bulb, distribute to the olfactory tubercle and parts of the septum, and by day 12, invade the superficial neostriatum (Fig. 10). Some of these fibers course within the frontoarchistriatal tract. Secondary olfactory fibers are therefore present in the vicinity of the most superficial neostriatal cells labeled in the lineage tracing experiments described above.

Charting of a case in which DiI was applied to one olfactory bulb (OB) in a 14 d embryo. A densely labeled fiber tract is shaded. Labeled fibers (dashes) course from there into the superficial paleostriatum, neostriatum (N), and archistriatum (A), as well as into some portions of the septum. FA, Frontoarchistriatal tract;HV, ventral hyperstriatum; P, paleostriatum; W Wulst. Scale bar, 1 mm.
DISCUSSION
Fluorescent dextrans are widely used for tracing the fate of embryonic precursor regions (e.g., Gimlich and Braun, 1985; Fraser et al., 1990), and fluorescent carbocyanine dyes have been used previously to study cell migration in the telencephalon of chick embryos (Montgomery, 1996). However, Montgomery (1996) reported that fluorescent tracer injected at 3 d of incubation could no longer be detected in embryos older than 6–7 d of incubation, presumably because of tracer dilution with cell division. Because this is well before the age at which the principal adult regions of the telencephalon become identifiable (Striedter and Beydler, 1997), we sought to develop a more sensitive fate-mapping method and chose biotinylated dextrans because biotinylation enables the use of highly sensitive signal amplification procedures to detect the tracer (see Materials and Methods). As described above, this method yielded satisfactory labeling of polyclones for at least 2 weeks after the initial tracer injection.
The present method is preferable to retrovirus-based methods for tracing cell lineage (e.g., Szele and Cepko, 1996) because the dextrans are not incorporated randomly into the VZ and can therefore be targeted at specific precursor regions. Compared with the chimeric transplantation method for mapping the fate of embryonic precursor regions (e.g., leDouarin et al., 1996), the present method is advantageous in that it permits the labeling of relatively small precursor regions at relatively late developmental stages, when vascularization complicates transplantation. Another useful feature of the present method is that labeled cells frequently remain detectable within the VZ long after most of the postmitotic neurons have migrated into more superficial regions, thereby enabling rather direct visualization of which VZ regions gave rise to which regions of the adult-like telencephalon. As the VZ becomes exhausted during development, fewer and fewer cells remain labeled in the VZ, but even 11 d after injection, the VZ often contained a cluster of labeled cells (Fig. 8), most of which are probably nondividing epithelial cells (Alvarez-Buylla et al., 1998).
Disadvantages of the present method are primarily that (1) the initial location of the injection sites cannot be visualized directly, (2) some descendant cells may escape detection because of tracer dilution, and (3) multiple precursor cells are generally labeled. Below, we present our model of neostriatal development, followed by a discussion of data obtained with other methods and, finally, an analysis of how telencephalic development in birds diverges from that in other vertebrates.
Development of the neostriatum in birds
Our data indicate that the avian neostriatum initially develops as a relatively straightforward thickening of the lateral pallial wall. Young neurons in this region generally accumulate deep to older, more highly differentiated, neurons, and their movements tend to be constrained along the long axes of radial glia (Striedter and Beydler, 1997). The initially slight rostrocaudal inclination of radial glia and polyclones increases between 8 and 10 d of incubation as the neostriatum bulges caudomedially and pulls the associated VZ with it. Polyclones at later stages therefore course diagonally from caudomedial origins in the VZ to rostrolateral terminations at the brain surface.
Tangential migration plays a relatively minor role in neostriatal development and is most apparent at fairly early and relatively late stages of development. Early tangential migration occurs within the mantle zone before 5 d of incubation, when cells often migrate ventrally away from the injection sites (see also Montgomery, 1996). The cellular dispersion produced by such early tangential migrations is amplified by the subsequent general expansion of the telencephalon. Thus, the early ventrally migrated cells probably correspond to the widely dispersed cells sometimes labeled in the most superficial paleostriatum at later stages of development. Late tangential migration increases as radial glia disappear (Striedter and Beydler, 1997), but migrating cells do not cross a boundary that emerges just ventral to the neostriatum between 5 and 8 d of incubation. This boundary develops in the same location and at approximately the same time as the LMD.
Although the LMD forms along a migration boundary, other cytoarchitectural boundaries within the lateral telencephalon do not coincide with lineage restriction boundaries. Thus, labeled polyclones generally extend from the caudal HV into the neostriatum and across several of the major neostriatal sensory regions, including the olfactory sensory zone in the most superficial neostriatum. These subdivisions do not, therefore, develop from unique patches of the VZ, and their parcellation must be attributable to factors other than lineage. Moreover, the data suggest that the term “ventral hyperstriatum” is misleading because at least the caudal portion of HV develops in close association with the neostriatum.
Some important questions that remain unresolved by our analysis are (1) whether some neostriatal cells might originate from regions dorsal to what we have called the presumptive neostriatum, (2) whether some sites in the caudal presumptive neostriatum give rise only to neostriatum and not to any part of the HV, and (3) which sites in the embryonic telencephalon give rise to the anterior portion of the HV. In addition, we did not make enough injections into the presumptive paleostriatum to be confident about the details of paleostriatal development. These issues will need to be addressed by more complete and detailed fate-mapping studies.
Comparison to data obtained with other methods
Previous studies based on purely descriptive material had led to a variety of disparate conclusions but generally showed that the neostriatum develops as a thickening of the lateral telencephalic wall (Holmgren, 1925). Tritiated-thymidine birth dating subsequently indicated that the developing neostriatum thickens primarily by the accumulation of young cells deep to older cells (Tsai et al., 1981a,b). Descriptive studies further indicated that the neostriatum develops just dorsal to a cytoarchitectural boundary that coincides with the dorsal limit of Cash-1 expression at early stages of development and later becomes the LMD (Montgomery, 1996).
Tritiated-thymidine data suggested that this cytoarchitectural boundary is a barrier to cell migration because isochrone zones were discontinuous across the boundary (Tsai et al., 1981b). This hypothesis was supported by lineage tracing with fluorescent tracers, showing that patterns of early cell migration differed across the boundary (Montgomery, 1996). It was called into question by chimeric transplantation experiments, however, that revealed extensive cell mixing between dorsal and ventral regions of the telencephalon (Balaban et al., 1988). Our data can be reconciled with the transplantation data if (1) some of the cell mixing in chimeric embryos occurred before 3 d of incubation, (2) the graft boundaries were not coincident with the migration boundary described here, and/or (3) some of the cell mixing is attributable to tangential migration of nonradial glia, which are rarely stained in our material (probably because they are born primarily after 10 d of incubation and have little cytoplasm).
Previous descriptive studies generally recognized HV, ectostriatum, field L, and nucleus basalis as brain divisions separate from the neostriatum proper. Radial glial data suggested that these areas might all be part of a large neohyperstriatal complex that becomes parcellated along lines other than lineage restriction (Striedter and Beydler, 1997). The present study strongly supports this hypothesis and further indicates that parts of HV, field L, neostriatum proper, ectostriatum, and nucleus basalis all develop from the same small patch of VZ. This was surprising, because field L, ectostriatum, and nucleus basalis are found at very different rostrocaudal levels of the telencephalon, whereas radial glia had been reported to course primarily radially (Striedter and Beydler, 1997). This discrepancy is probably caused by (1) an underestimate of the rostrocaudal inclination of radial glia in our previous analysis and (2) the increase in the rostrocaudal inclination of polyclones at later stages of development, when the radial glia have begun to disappear.
Although our data suggest that radial migration along radial glia dominates neostriatal development, evidence for long rostrocaudal migration has been presented in several previous studies. Thus, a late tangential migration from a rostroventral portion of the VZ to field L had been postulated on the basis of silver staining and tritiated-thymidine data (Jones and Levi-Montalcini, 1958; Tsai et al., 1981a). The present data cannot rule out such a migration but show that at least part of field L develops from a more posterior portion of the VZ near the HV. Retroviral data indicative of long tangential migrations within the avian telencephalon (Szele and Cepko, 1996) are more difficult to evaluate because only the rostrocaudal subset of clones has been described. Most of the neostriatal retroviral clones are consistent with the model described above, however, and additional studies will be needed to resolve the few apparent discrepancies.
Comparison to other species
Holmgren (1922, 1925) first argued forcefully that distinct pallial and subpallial divisions of the telencephalon exist within the telencephalon of all vertebrates, at least during early stages of development. Holmgren also recognized medial (hippocampal), dorsal (general), and lateral (piriform) divisions within the pallium of most vertebrates. These three pallial divisions have been identified as distinct cytoarchitectural entities in many adult anamniotes (i.e., fishes and amphibians), in which the telencephalon does not become distorted by extensive cellular proliferation and migration (Northcutt, 1995). Connectional studies further showed that the lateral pallium in anamniotes receives prominent inputs from the olfactory bulb and that the dorsal pallium receives at least some inputs from the dorsal thalamus. In consequence, the lateral pallium of anamniotes has generally been interpreted as the homolog of mammalian piriform cortex, whereas the anamniote dorsal pallium is generally considered homologous to mammalian neocortex.
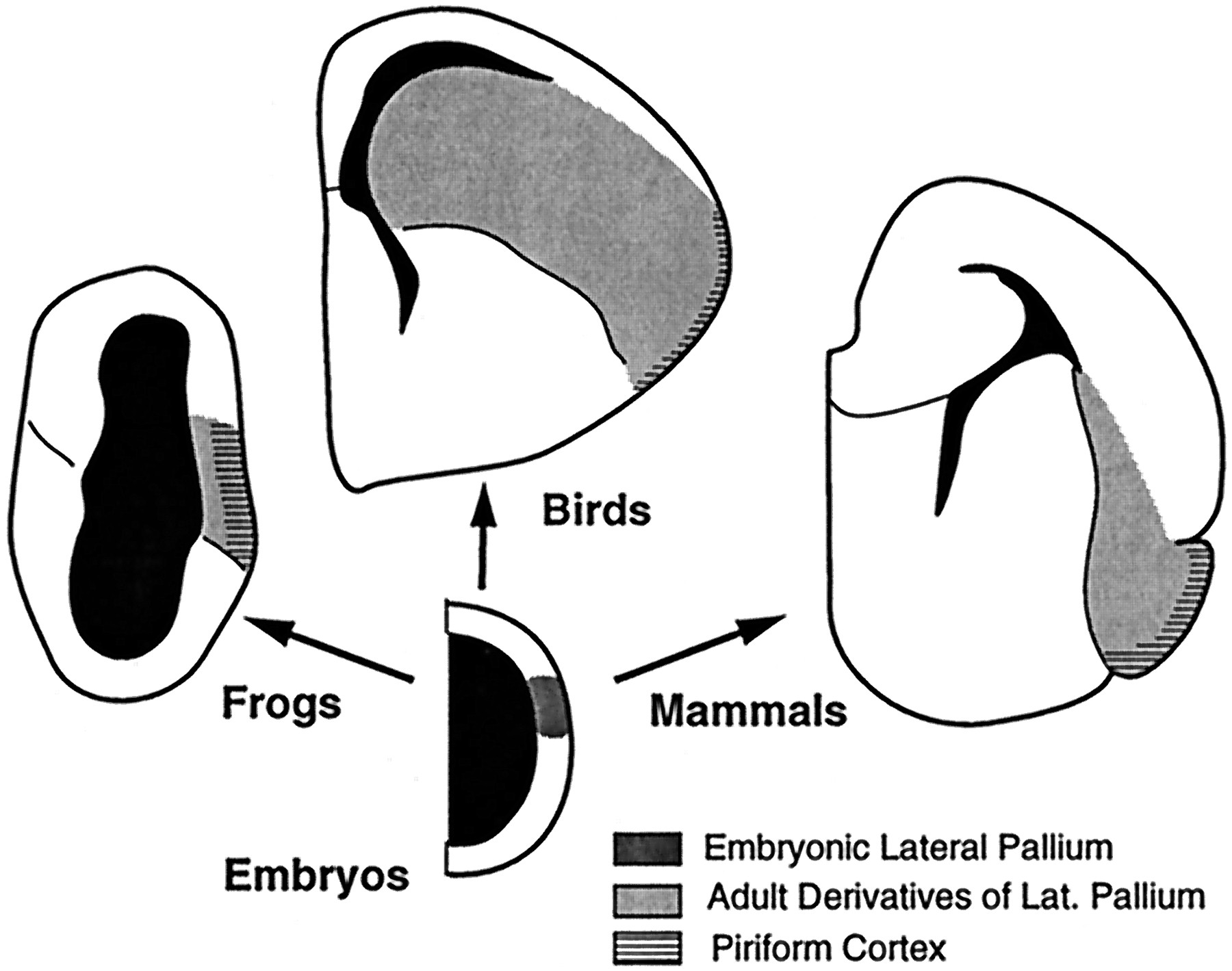
The situation is less straightforward in sauropsids (birds and reptiles), in which the lateral telencephalon thickens and differentiates further (Fig. 11). Most significantly, the secondary olfactory projections in sauropsids are restricted to the superficial portion of the thickened lateral pallial wall, whereas the deeper portions (i.e., the neostriatum in birds) receive major inputs from the dorsal thalamus. These connectional data have been used to argue that the thalamorecipient deep portions of the lateral pallial wall in sauropsids are homologous to part of mammalian neocortex (Karten, 1969; Reiner, 1991). This hypothesis implies that the avian neostriatum is homologous to part of the dorsal pallium in anamniotes. The present developmental data clearly indicate, however, that the neostriatum develops just dorsal to the pallial–subpallial boundary, in the position where the lateral pallium develops in anamniotes. This suggests that the neostriatum is instead homologous to the anamniote lateral pallium and has greatly elaborated its inputs from the dorsal thalamus (Northcutt and Kaas, 1995; Striedter, 1997). This hypothesis, in turn, raises the question of what the avian neostriatum might be homologous to in mammals.

Hypothesized relationship between ontogeny and phylogeny for the telencephalon of tetrapods. Amniote embryos possess a lateral pallial patch of VZ that develops into the lateral pallium, which is characterized by secondary olfactory projections to its most superficial portion. Whether a lateral pallium of this kind exists in mammals remains speculative (see text).
Unfortunately, the development of the lateral telencephalon in mammals remains poorly understood. Most controversially, some authors have argued that the mammalian LGE gives rise not only to striatum but also to piriform cortex (Smart, 1985; DeCarlos et al., 1996). Because the striatum is generally considered to be part of the subpallium, and piriform cortex part of the lateral pallium, this interpretation implies that the pallial–subpallial boundary in mammals is not a lineage restriction boundary and that mammals may not possess a lateral pallium comparable with that in other vertebrates. Alternatively, the mammalian telencephalon does contain a lateral pallium, which is initially located dorsal to the subpallium (Alvarez-Bollado and Swanson, 1996; Striedter, 1997; Nieuwenhuys et al., 1998). According to this model, the pallial–subpallial boundary in adult mammals corresponds to the zone between striatum and the endopiriform claustrum, where the VZ gradually disappears or recedes as the LGE expands dorsally. Areas immediately dorsolateral to this boundary, i.e., piriform cortex, endopiriform claustrum, and basolateral amygdala, would therefore constitute the lateral pallium in adult mammals (Fig. 11). This second hypothesis is attractive from a comparative perspective, because it requires less dramatic evolutionary changes in telencephalic development. Experimental evidence can at present be adduced in favor of both hypotheses, however, and remains less than decisive.
Nonetheless, the present study indicates that the avian neostriatum develops dorsal to a major lineage restriction boundary that is probably homologous to the lateral pallial–subpallial boundary described in other vertebrates. As a derivative of the most ventrolateral pallium, the neostriatum is unlikely to be homologous to either striatum (subpallium) or neocortex (dorsal pallium) in other vertebrates. Its homolog should therefore be sought among derivatives of the lateral pallium. This does not imply that the neostriatum, or subdivisions of it, must have strict homologs within the lateral pallium of other taxa, because evolutionary divergences in development may lead to truly novel (nonhomologous) adult characters in different species (Striedter, 1998). We propose only that the presumptive neostriatum patch of VZ injected with dextrans in the present study is probably homologous, as an embryonic lateral pallium, to embryonic lateral pallia in other vertebrates. The extent to which adult structures within the lateral pallium can be homologized across taxa will need to be determined by subsequent studies.
Footnotes
This research was supported by an Alfred P. Sloan Research Fellowship and a National Science Foundation grant to G.S. We are indebted to John Montgomery and Scott Fraser for first showing us how to inject tracers into chick embryos. We thank Evan Balaban, Glenn Northcutt, and Luis Puelles for stimulating discussions and constructive comments on the manuscript, Raju Metherate for access to his electrode puller, and Linda Do for help on the DiI experiments.
Correspondence should be addressed to Dr. Georg F. Striedter, Department of Psychobiology and Center for the Neurobiology of Learning and Memory, University of California at Irvine, Irvine, CA 92697.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}