

Abstract
Each eye of the mollusc Hermissenda consists of five photoreceptors, two type A and three type B cells. Type A cells are quiescent, whereas B cells are spontaneously active in the dark. Differences in the intrinsic membrane properties of type A and B photoreceptors were studied using voltage- and current-clamp techniques. The current density of a Ni2+-sensitive, low-voltage activated Ca2+ current was similar in the two cell types. However, type B cells express an inward rectifier current (Ih ) that has different permeation and pharmacological properties from the inward rectifier current in type A cells. The current in the B cells was time-dependent and was blocked by Cs+. Na+ and K+ were the charge carriers forIh . The inward rectifier current in A cells (IK1) was time-independent, was selectively permeable to K+, and was blocked by Ba2+. Ni2+ reduced the spontaneous spike activities of type A and B cells, whereas Cs+ produced membrane hyperpolarization and reduced the spike activities of dark-adapted B cells. The application of both Cs+ and Ni2+ completely blocked dark-adapted spontaneous activities of B cells. Moreover, Ba2+ increased the excitability of type A cells but not B cells. Hence, differential expression of the two distinct inward rectifiers found in type A and B cells contributes to differences in their intrinsic membrane properties. Because changes in the excitability of the two cell types are correlates of conditioning inHermissenda, modulation of these underlying currents may play a major role during conditioning-induced plasticity.
Alteration of membrane excitability of neurons is an example of cellular plasticity associated with learning in nervous systems of vertebrates and invertebrates (Abrams, 1985; Jester et al., 1995). The molluscHermissenda has a simple eye that serves not only as a site for photo transduction but also as a locus for cellular plasticity (Crow, 1988; Farley et al., 1990; Alkon et al., 1992; Matzel and Rogers, 1993). The eye consists of five photoreceptors that are classified as type A and B cells, based on anatomical and electrophysiological differences (Alkon and Fuortes, 1972). Classical conditioning of Hermissenda produces enhanced excitability of the B photoreceptors that is expressed as an increase in input resistance (Crow and Alkon, 1980; Crow, 1988; Matzel and Rogers, 1993) and a reduction of both the transient K+ current (IA ) and the calcium-activated K+ current (IK,Ca) (Alkon, 1979; Alkon et al., 1985). In contrast, type A cells undergo a reduction in dark-adapted input resistance (Farley and Alkon, 1982; Farley et al., 1990). Differences between the two cell types are not only expressed in conditioned animals but can be seen in naı̈ve animals as well.
Type A cells are quiescent in the dark, and type B cells undergo rhythmic spike activities in darkness. Light elicits depolarizing generator potentials in both cell types. At the end of a light stimulus, the A cells remain electrically silent after a few seconds of dark adaptation, but the B cells remain tonically active (Alkon and Fuortes, 1972). The underlying mechanisms for the different electrical properties between the two cell types are unknown. Synaptic inhibition between the photoreceptors has been used, in part, to explain differences in the electrical properties of A and B cells (Alkon and Fuortes, 1972). However, such an explanation may not suffice to explain the properties of the two cell types. For instance, the A cells are relatively hyperpolarized in the dark-adapted state, whereas the B cells are more depolarized and remain spontaneously active; yet the inhibitory inputs at the B cells are stronger than are those at the A cells (Alkon and Fuortes, 1972).
Differential expression of different ionic currents in type A and B cells may contribute to the apparent electrophysiological differences between the two cell types. To examine this possibility, we studied ionic currents and electrical properties of the photoreceptors using the whole-cell voltage-clamp technique and conventional intracellular recordings. The properties of three distinct inward currents are presented. Type A and B cells express a transient Ca2+ current (ICat) at similar densities. But the A cells predominantly express a time-independent K+-permeable inward rectifier current (IK1), which tends to hyperpolarize the cells and prevent tonic activity in darkness. In contrast, type B cells exhibit a time-dependent Na+/K+-permeable inward rectifier current (Ih ).Ih promotes membrane depolarization. The combined effects of ICat andIh support spontaneous activities of type B photoreceptors in the dark.
MATERIALS AND METHODS
Cell isolation. Specimens of Hermissendawere obtained from Sea Life Supply (Sand City, CA) and housed in artificial seawater (ASW) tanks at 14–16°C. Animals were fed scallops daily and maintained on a 12 hr light/dark cycle. The photoreceptors of Hermissenda were freshly isolated from the nervous system, and the type A and B cells were identified according to methods described previously (Yamoah and Crow, 1994, 1996). The nervous systems were dissected and incubated in ASW (composition in mm is 420 NaCl, 10 KCl, 10 CaCl2, 22.9 MgCl2, 25.5 MgSO4, and 15 HEPES, buffered to pH 7.8 with 1N NaOH) at 4°C for 10 min and then were exposed to an enzyme solution made of 7 mg/ml dispase grade II (Boehringer Mannheim, Mannheim, Germany) and 1 mg/ml protease (Sigma, St. Louis, MO) in ASW for 10 min at 4°C. The preparation was transferred to room temperature (20°C) for another 7–10 min and then was washed several times with ASW at 4°C. After incubation for 20 min at 4°C, the eyes were isolated. Type A and B cells were identified on the basis of their position relative to the lens and the optic nerve. Therefore, isolated eyes without a lens and the stump of the optic nerve were discarded. We did not further classify isolated photoreceptors as either medial or lateral (Alkon and Fuortes, 1972). Using a fire-polished pipette, we isolated individual photoreceptors, plated them into 35 mm sterile culture dishes (Falcon, 1008; Fisher Scientific, Houston, TX) containing 2 ml of gentamycin-treated ASW (50 mg/ml), and stored the dishes at 4°C (Yamoah et al., 1994). The solution was aspirated and replaced with recording medium 5 min before voltage-clamp experiments were performed.
Voltage clamp. Macroscopic whole-cell inward rectifier and calcium currents were recorded at room temperature (20°C). All chemicals were obtained from Sigma unless otherwise noted. The compositions of the bath solutions for recording calcium and inward rectifier currents were as follows (in mm): for the Ca2+ current, 400 tetraethylammonium (TEA) acetate or 400 N-methyl-d-glucamine (NMG) and 200/250 TEA-chloride, 10 CaCl2, 5 MgSO4, 5 4-aminopyridine, and 15 HEPES, pH 7.7, titrated with TEA-hydroxide (TEA-OH); and for the inward rectifier currents, 200 NMG, 300 choline chloride, 2–50 KCl, 0–2 CaCl2, 50 MgSO4, and 15 HEPES, pH 7.7 (with KOH). Experimental agents were applied by bath superfusion at a rate of 1–1.5 ml/min using gravity and an aquarium pump for suction (Penn-Pax Plastic, Garden City, NY) in a 1.2 ml experimental chamber. The pipette solution contained (in mm): for the Ca2+ current, 300 NMG, 200 CsCl, 50 HEPES, 2–5 EGTA, 20 NaCl, 2 MgSO4, 20 TEA, 5 MgATP, 1 Na2GTP, and 10 reduced glutathione, pH 7.4 (with TEA-OH); and for the inward rectifier currents, 0–20 NaCl, 0–2 MgSO4, 50 HEPES, 0.1–5 EGTA, 50–250 KCl, 250–350 NMG, 5 MgATP, 1 Na2GTP, and 10 reduced glutathione, pH 7.4 (with HCl).
Voltage clamp in the whole-cell configuration (Hamill et al., 1981) was used to measure the inward rectifier and Ca2+currents using the Axopatch 200A amplifier (Axon Instruments, Foster City, CA). Recording pipettes were pulled and fire-polished with a horizontal Flaming-Brown microelectrode puller (Sutter Instrument Company, San Rafael, CA). To reduce the pipette capacitance, we coated regions close to the tip of the pipette with Sylgard 184 (Dow Coring, Midland, MI). Pipettes had a final resistance of 0.7–1.6 MΩ. Series resistance (1.2–4.5 MΩ) was compensated (nominally 50–60%). Records were filtered at 5 kHz with a low-pass Bessel filter and were digitized at 10 kHz with a Digidata interface controlled by the pClamp software version 5.7.1 (Axon Instruments). Data were stored on a personal computer (Gateway, Sioux City, SD). Leakage current was canceled on-line with the p/−5 protocol. Cell capacitance was measured as described (Yamoah et al., 1994). After the capacitance of the electrode had been canceled electronically, cells were held at −60 mV and stepped to −80 mV (ΔV = 20 mV). The time constant of relaxation and the area above the transient that represents the charge (Q) were measured. The capacitance (Cm ) was calculated from the equationCm = Q/ΔV. Analyses of the data were performed using Clampfit (Axon Instruments), Sigma plot (Jandel Scientific, San Rafael, CA), and Microcal Origin (Microcal Software, Northampton, MA) programs. Values are expressed as mean ± SD.
Intracellular recording. Isolated nervous systems were incubated in a protease solution (P-8038, 2 mg/ml in ASW; Sigma) at room temperature for 7 min to facilitate microelectrode penetration. Photoreceptors were impaled using 30–45 MΩ microelectrodes filled with potassium acetate. Current-clamp experiments were accomplished via an Axoclamp 2A (Axon Instruments). Data were fed into a tape recorder and later digitized at 1 kHz or obtained as recordings from a Gould Brush recorder.
RESULTS
We investigated the membrane properties of Hermissendaphotoreceptors to understand the underlying mechanisms for spontaneous activity and the response of type A and B photoreceptors to light and darkness. The kinetic and pharmacological properties of a low-voltage activated transient calcium current (ICat) and two inward rectifier currents were examined in both cell types. Type A and B cells expressedICat at similar densities. Two types of inward rectifier currents are described from both type A and B cells, time-dependent and time-independent currents. Both cell types expressed the two currents, but the A cells predominantly expressed the time-independent current (∼90%), whereas the B cells exhibited the time-dependent current (∼77%).
Light response and dark adaptation of photoreceptors
Figure 1 illustrates the basic differences in the electrical properties of type A and B photoreceptors. Figure 1 (top) shows the light response of a typical type A cell during a 5 sec light pulse. The A cells are relatively hyperpolarized at rest compared with type B cells (Alkon and Fuortes, 1972) and are quiescent after dark adaptation. In contrast, as shown in Figure 1 (bottom), B cells are tonically active even after dark adaptation. Inspection of the records in Figure 1 suggests that type A and B photoreceptors differ both in their tonic and phasic responses to light. We speculated that the inward currents expressed by the two cell types may differ, rendering the B cells spontaneously active while the A cells remain electrically silent after dark adaptation. Three inward currents were found to be crucial for the dark-response properties of the A and B cells.

Differences in light- and dark-response properties of Hermissenda photoreceptors.Top, Voltage response obtained in a type A cell with a 5 sec light pulse (solid bar) is shown. The nervous system was bathed in artificial seawater, and the photoreceptor was impaled with an electrode containing a 3 m potassium acetate solution. After attenuation of the light stimuli, the membrane potential of A cells typically drops to the resting potential and remains silent in darkness. Inhibitory synaptic potentials induced from adjacent photoreceptors can be recorded. Bottom, Type B cells continue to be tonically active in darkness. A 5 sec light flash induces the characteristic membrane depolarization shown here. In contrast to A cells, B cells remain spontaneously active in the dark-adapted state.
Transient calcium current
Transient low-voltage activated Ca2+currents have been reported to contribute to the initial depolarization phase of action potentials (Llinas and Yarom, 1981). The transient Ca2+ current in the soma of the photoreceptors has been studied previously (Yamoah and Crow, 1994), but the functions of the current are unknown. Experiments were performed in solutions expected to suppress all ionic currents except Ca2+currents (see Materials and Methods). Shown in Figure2A are representativetraces of the transient component of the Ca2+ current recorded from a type B cell at a holding potential of −90 mV. Data of the current density plot from types A and B cells are represented in Figure 2B. In both cell types, the current was activated at potentials close to −50 mV and peaked at approximately −10 to 0 mV. There were no significant differences between the two cell types in the current densities at all recorded step potentials. The examples shown represent cells that predominantly express the transient component of the Ca2+ current. Typically, we isolated the transient current by suppressing the sustained Ca2+ current with bath application of nitrendipine (5 μm) (Yamoah and Crow, 1994). Steady-state activation and inactivation ofICat revealed a window current close to the resting potential of the photoreceptors (−60 to −40 mV) (Fig.2C). A previous report showed that both type A and B cells express sustained and transient Ca2+ currents (Yamoah and Crow, 1994, 1996). In an A cell in which both Ca2+ current subtypes were expressed, Ni2+ blocked the transient component but left the sustained current unblocked (Fig. 2D). Typicaltraces of the effects of Ni2+ onICat in an A cell are shown in Figure2E. The records show that the transient Ca2+ current was reduced by Ni2+in a concentration-dependent manner with a half-blocking concentration of 0.4 ± 0.21 mm (n = 6). The time course and the concentration dependence of the blocking action of Ni2+ on the transient current are demonstrated in Figure 2F. The effect of Ni2+ was reversed after ∼50 sec of washout.

Transient Ca2+ current.A, Examples of transient Ca2+ currenttraces generated by depolarizing voltage steps in a B cell in ASW with 10 mm Ca2+ are shown. Voltage steps were delivered from the holding potential of −90 mV to potentials ranging from −80 to 60 mV in 10 mV increments. Sometraces have been removed for clarity. B, The I–V relations of current densities plotted as a function of voltage from type A (•) and B (○) cells are shown. Cell capacitance was measured as described in Materials and Methods.C, In a type B cell that predominantly expressed the transient Ca2+ current, steady-state activation (○) and inactivation (•) curves were generated and fitted to a Boltzmann function. The fits are plotted withsolid and dotted lines. Half-activation and inactivation (assuming complete voltage-dependent inactivation) were −40 and −48 mV, respectively. The maximum slopes for the activation and inactivation curves were 10 and 11 mV, respectively. Overlap of the steady-state activation and inactivation curves at a voltage range of −60 to −40 mV suggests that the Ca2+ current is active at the resting potentials of the photoreceptors (−55 to −45 mV). D, Ni2+ at 0.5 to 2 mm selectively blocked the transient Ca2+ current. The currenttraces were elicited from a holding potential of −90 mV and stepped to −20 mV from a type A cell. Only the sustained Ca2+ current remained after application of 0.8 mm Ni2+. The difference current (Ni2+-sensitive current; trace shown with dotted line) is the transient component.E, Changes in the transient Ca2+current magnitude as [Ni2+] was gradually increased from 0 to 0.8 mm are shown. F, The effect of Ni2+ was reversible after washout. The time course and the effect of different concentrations of Ni2+ on the transient Ca2+current are shown.
Time-dependent inward rectifier current
In Figure 3A are representative traces of a time-dependent, hyperpolarization-activated current recorded from a type B cell with a holding potential of −45 mV. A previous report of inward rectification in the photoreceptors identified the current asIir (Acosta-Urquidi and Crow, 1993). In the initial report, only one form of inward rectification in the B photoreceptors was examined. As shown below, there are two distinct types of inward rectification in type A and B cells. Because of the similarities of the inward rectifier current in the B cells to the time-dependent hyperpolarization-activated currents in photoreceptors (Hestrin, 1987) and other systems (Angstadt and Calabrese, 1989;Holt and Eatock, 1995), the current will be denotedIh . Tail current traces from the same cell are shown (Fig. 3B). The current–voltage relationship is shown in Figure 3C. The permeation property of the channel was assessed by alteration of bath [K+], [Na+], and [Cl−]. Alteration of bath or pipette [K+] shifted the reversal potential (Erev) of the current (Fig.3C). An increase in bath [Na+] shifted the Erev to more positive voltages, whereas the reverse was the case when [Na+] was reduced (Fig.3D). In contrast, changes in [Cl−] did not alter the Erev of the current. A small but significant Cl− current is present in the photoreceptors (Yamoah and Kuzirian, 1994); however, this current is predominantly outward rectifying. Therefore, Cl−current contributes very little to the total inward-rectifying current in the photoreceptors.
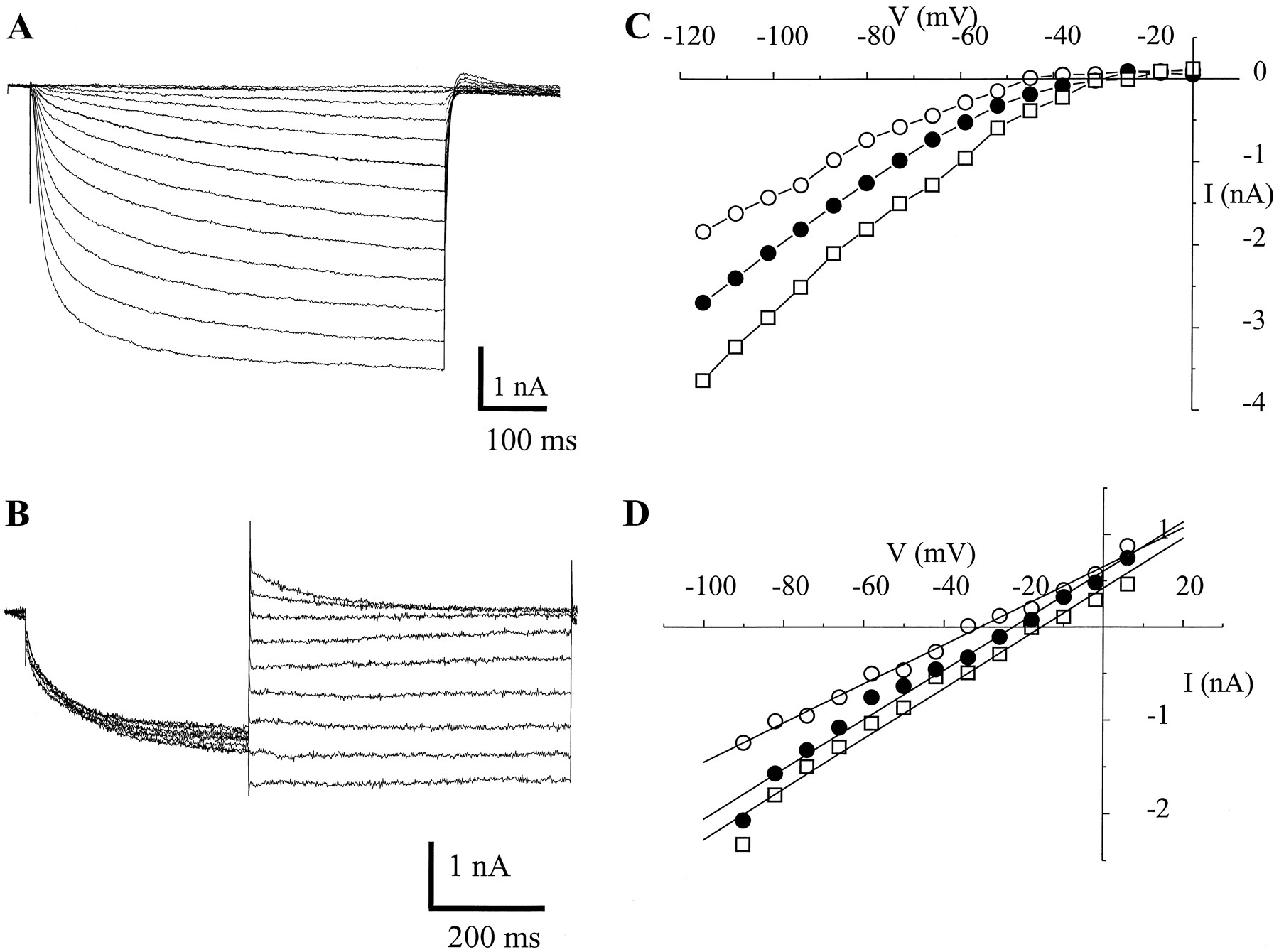
Time-dependent inward rectifier current (Ih ). A, A type B photoreceptor was held at −45 mV and stepped to positive and negative voltages relative to the holding potential (ΔV = 7 mV). A time-dependent inward current that exhibited rectification at potentials positive to the apparent reversal potential was elicited.Traces shown were collected from an average of five runs. B, Shown as well are tail currenttraces from the same cell. C, The corresponding I–V plot is shown. An apparent reversal potential of −36 mV was calculated from the regression line generated from instantaneous current traces (○). Alteration of external K+ concentration ([K+]o) shifted the apparent reversal potential of Ih . Pipette [K+]i was 250 mm, and the altered [K+]o was 2.5, 10, or 30 mm, shown on the corresponding I–V plots as ○, •, or ■, respectively. D, Changes in [Na+]o also altered the apparent reversal potential (Erev) ofIh . Alteration ofErev by changing [Na+]o suggested that theIh channel is permeable to Na+ as well. Shown are representative plots of the instantaneous I–V relationship generated from tail current records with varying [Na+]o. The [Na+]o, the correspondingErev, and the conductances are 20 mm, −30.9 mV, 21.1 nS; 70 mm, −22.8 mV, 22.8 nS; and 100 mm, −15.6 mV, 27.0 nS, shown as ○, •, and ■, respectively. Using the Goldman–Hodgkin–Katz equation (Hille, 1992), the permeability ratio for K+ versus Na+(PK/PNa) = 0.7 for this channel.
Activation of the current was voltage- and time-dependent. A normalized, steady-state activation curve forIh is presented in Figure4A. The Boltzmann distribution fit for the curve had a half-activation voltage (V1/2) of −74 ± 5 mV and a maximum slope (Km) of 15.5 ± 2.3 mV (n = 9). A single time constant provided a best fit for the activation and deactivation profiles of the current. A plot of the time constants of activation and deactivation versus membrane potentials is shown in Figure 4B.

Steady-state activation ofIh and the time constant of activation.A, The relative current plotted against the step potentials. Error bars represent SD (n = 9 cells). The continuous curve was generated from the Boltzmann function (I/Imax = [1+exp(V1/2-V)/Km]−1), where V1/2 is the half-activation voltage and Km is the maximum slope; V1/2 of −73.6 ± 5.1 mV andKm of 15.5 ± 2.3 mV were estimated. B, Time constants of activation and deactivation fitted with a single exponential and plotted against the corresponding voltages. Representative data from five cells are plotted with scattered points. C, Block ofIh by Cs+. The time-dependent inward rectifier current was blocked in the presence of 0.5 mm Cs+. Cs+induced a rapid decay of the current. D, Current–voltage relationship of control (▪) and after application of 0.5 mm Cs+ (○) (n= 6 type B cells). In the presence of 2 mmCs+, most of the inward-rectifying currents were blocked; however, there remained a time-independent current (data not shown). In five type B cells with a peak current of −1.95 ± 0.98 nA, the remaining time-independent peak current after application of 2 mm Cs+ was −0.20 ± 0.14 nA. Therefore it is estimated that only 10% of inward rectification in the type B cells is time-independent. E, Estimation of the binding site for Cs+. The voltage dependence of Cs+ blockade of the pore was determined assuming a binding model as described previously (Woodhull, 1973). The model assumes a single site for Cs+ binding whose availability but not intrinsic affinity changes with voltage. Cs+ is assumed to be an impermeant blocker that enters and exits the pore from the same side. A linear transformation of the voltage dependence of the ratio of the unblocked current (peak current amplitude) and blocked current (steady-state current amplitude) is as follows: ln{(Ic /Ib )−1} = ln(KA ) × [Cs+]−δ(ZeV/RT), where Ic is the unblocked andIb is the blocked macroscopic current amplitudes, KA is the association constant, δ is the fractional electrical distance from the outer face of the membrane, and Z, e,R, and T have their usual meanings. The slope of the linear plot gives the δ. The data were fit by least squares linear regression. The calculated membrane electric field (zδ) sensed by Cs+ from the linear plot was 0.8.
The sensitivity to block of Ih by Cs+ was tested (Angstadt and Calabrese, 1989; Bond et al., 1994). Cs+ blockedIh in a voltage- and concentration-dependent manner. At large negative pulses, the current turned on rapidly and then decayed with time. This decay became pronounced in the presence of Cs+ (Fig.4C). The current–voltage relationships for controltraces and after application of 0.5 mmCs+ were obtained from six B cells (Fig.4D). The apparent fraction of the membrane electric field (zδ) sensed by Cs+ was calculated to be 0.8 ± 0.1 (n = 5) (Fig.4E; Woodhull, 1973). Sensitivity of the current to Cs+ is similar to the inwardly rectifying conductances observed in vertebrate CNS neurons (Yamaguchi et al., 1990), optic nerve, and cone photoreceptors (Eng et al., 1990;Maricq and Korenbrot, 1990), as well as in invertebrate neurons (Angstadt and Calabrese, 1989). However, the hyperpolarization-activated current in Hermissendaphotoreceptors is dissimilar to those reported from retinal glial cells (Newman, 1993), cholinergic neurons in rat brain (Yamaguchi et al., 1990), and Aplysia buccal muscles (Brezina et al., 1994) in that the current showed no sensitivity to Ba2+. In fact, in the presence of Ba2+ or Ca2+, the magnitude of the current increased (data not shown). This may reflect contamination ofIh by a background Ca2+current in the photoreceptors. To avoid the masking effects of a possible hyperpolarizing-activated Ca2+ conductance (Preston et al., 1992), we made recordings ofIh at reduced [Ca2+] (see Materials and Methods).
Time-independent inward rectifier current
Type A cells expressed a time-dependent and time-independent inwardly rectifying current. However, the time-independent current was predominant (∼77%; n = 6; Fig.5) in the A cells. We refer to the time-independent inward rectifier current asIK1, consistent with the terminology used for a similar current in cardiac cells (Kurachi, 1985). Figure5A shows current traces recorded in a type A cell superfused with a Na+-free recording solution during hyperpolarizing voltage steps from the −50 mV holding potential. Hyperpolarization to potentials more negative than −70 mV induced an inward current. The current activated rapidly and exhibited little or no decay. As the bath [K+]o was increased, the reversal potential of the current shifted to more positive potentials, in a manner predicted by the Nernst potential for a K+-selective conductance (Fig. 5C). Raising [K+]o resulted in a steeper slope conductance as well and produced a shift in the activation voltage range of the current in the positive direction (data not shown). In contrast to Ih ,IK1 was blocked by Ba2+ (Fig.5A,B). Ba2+produced a voltage-dependent block ofIK1, and the fractional electrical distance between the external orifice of the channels and the Ba2+ binding site was calculated to be 0.6 ± 0.1 (n = 4: see Fig.5D,E). Application of Na+-containing solution induced an apparent inactivation of the current that is reminiscent of the extracellular Na+ block of inward rectifier currents observed in tunicate egg cells (Ohmori, 1978), skeletal muscles (Standen and Stanfield, 1979), and endothelial cells (Silver and DeCoursey, 1990).

The inward rectifier current in type A photoreceptors was carried by K+. A, Type A cells also exhibited inward rectification; however this current was time-independent, and the apparent Erevwas more negative (−70 to −80 mV) than were currents recorded from type B cells (−30 to −40 mV). B, This inward rectifier current can be blocked by Ba2+. The remaining current after application of Ba2+(inset) exhibited a time dependence of activation and can be blocked by Cs+. The effects of Ba2+ were reversible (data not shown).C, By varying the external concentration of K+ and maintaining a constant pipette K+ concentration of 250 mm, we obtained the reversal potentials of the inward rectifier current; these were plotted against the external K+ concentration. Data were fit with a linear regression with a slope of 56 mV per log unit (r = 0.989), consistent with a K+-selective pore. D, The current–voltage relations of the inward rectifier current at baseline (○; n = 7) and after application of 0.25 mm (•) and 2.5 mm (■) Ba2+ are illustrated. This current was more sensitive to block by Ba2+ than by Cs+ (data not shown). Cumulative data from six type A cells (control peak current, 1.59 ± 0.54 nA; Ba2+-insensitive time-dependent current, 0.35 ± 0.21 nA) show that type A cells expressed ∼77% ofIK1 and 23% of Ih.E, The binding site of Ba2+ within the pore is estimated. The slope yields a δ of 0.6 from the outer face of the membrane (see Fig. 4E).
Effects of Cs+, Ba2+, and Ni2+ on the excitability of A and B cells
Alkon (1979) reported that Ni2+ altered spontaneous activity of B photoreceptors. Although the mechanisms for the effects of Ni2+ were not investigated, it is possible that Ni2+ block of theICat may be the underlying mechanism for Ni2+-induced alteration of spontaneous activity of the B cells. It has been suggested that inward rectification is responsible for a voltage “sag” observed in the B cells after the injection of hyperpolarizing currents (Acosta-Urquidi and Crow, 1995). The voltage sag is followed by a rebound spike activity. A combination of the inward currents may partly explain the differences in the properties of A and B photoreceptors. This idea was examined further by blocking the transient Ca2+ and inward rectifier currents with bath application of Ni2+, Cs+, and Ba2+.
To account for diffusional barriers in intact preparations, we applied relatively high concentrations of the blockers. Figure6 illustrates the effects of Ni2+ and Cs+ on type B cells. Block of the transient Ca2+ current by Ni2+ induced a burst pattern that was characteristically phasic, unlike the nonphasic, spontaneous activity of the B cells in vivo. In Figure 6A, 1.5 mm Ni2+ reduced the frequency of spontaneous activity. Spontaneous activity of the photoreceptors was reduced further after the application of 2 mmCs+. Both Ni2+ and Cs+ reduced the spontaneous firing of the type B cells (Fig. 6B–D). Cs+ alone induced membrane hyperpolarization and reduced the rate of firing of B cells (Fig. 7). We further illustrate that membrane depolarization by current injection was insufficient to increase the firing rate, after block ofIh by Cs+. The effects of Cs+ were reversible after washout.

Pharmacological manipulation of the electrical activity of type B cells with Ni2+ and Cs+. A, The characteristic tonic activity of B cells in control solutions is transformed into alternating burst activity after bath perfusion with 1.5 mmNi2+-containing artificial seawater. Addition of 2 mm Cs+ to the Ni2+solution completely abolished spontaneous activity of the B cell in dark-adapted conditions. B, Top, Light-elicited generator potential and the corresponding spike rate in the predark-adapted stage were reduced in the presence of Ni2+ and Cs+.Bottom, Replacement of Ni2+- and Cs+-containing solution with normal artificial seawater restored the excitability of the B cells. C, In the cell shown in B, Ni2+ decreased the spike rate from ∼3 to 1 Hz; Cs+ reduced the spike rate further to 0 Hz. D, Data from five cells on the effects of Ni2+ and Cs+ on spike rate and membrane potential are shown.

Effects of Cs+ on the membrane properties of type B photoreceptors. A, Spontaneous activity of B photoreceptors in normal artificial seawater consists of uninterrupted burst spikes in type B photoreceptors in the dark. After application of 2 mm Cs+, the steady-state potential dropped to a more hyperpolarized potential, and the spike rate was reduced. Injection of positive current to depolarize the steady-state potential close to control conditions was insufficient to restore the spike rate. However, after 10 min of washout, the spike rate returned to control levels. B, In the B cell shown in A, Cs+ induced a spike rate of ∼0.5 Hz compared with a rate in controls of 2.5 Hz. C, Cumulative data from five type B cells are shown. Cs+ reduced the spike rate from 2.7 ± 0.5 to 0.7 ± 0.3 Hz. However, the spike rate returned to 2.5 ± 0.4 Hz that was not significantly different from control after washout.
Type A and B cells have similar spike rates in response to light (∼40 Hz), but the two cell types are dissimilar in their dark-response properties (Fig. 8). The B cells remain tonically active with a spike rate of ∼3 Hz in darkness, whereas the A cells remain quiet after termination of light stimuli (Fig. 8). Ba2+, a specific blocker ofIK1, significantly increased the spontaneous activity of A cells in darkness [spike frequency in the dark, 0.2 ± 0.2 Hz (mean ± SD); spike frequency in darkness in the presence of Ba2+, 2.2 ± 1.1 Hz (mean ± SD); n = 5; p < 0.005] but failed to produce substantial changes in the light-elicited spike activity in type A and B cells (see Fig. 8B). In contrast to the activity in type A cells, the tonic activity of type B cells in darkness was not significantly altered by Ba2+ (spike frequency of B cells in the dark, 2.5 ± 0.6 Hz; spike frequency in darkness in the presence of Ba2+, 2.6 ± 0.6 Hz; n = 5;p = 0.8, not significant).

Effects of Ba2+ on spike rates of type A and B cells. A, Profile of electrical activity of type A and B photoreceptors before and after stimulation with a 5 sec light stimulus. As seen in previous figures, the B cells retained spontaneous activity after the light was turned off. B, Histograms generated from five A and B photoreceptors. Ba2+ (1 mm) increased the spike rate of type A cells approximately 10-fold from 0.2 ± 0.2 to 2.2 ± 1.1 Hz. However, the light-elicited spike rate remained virtually the same in the presence or absence of Ba2+. The postlight electrical activity of the A cells was enhanced further in Ba2+-containing artificial seawater than in normal artificial seawater. In contrast, Ba2+ produced no significant change in spike rates in type B cells both in the pre- and postlight conditions. Prelight (pre) corresponds to 20 min of dark adaptation, and postlight (post) corresponds to 2 min after light stimulation.
DISCUSSION
The properties of a transient Ca2+ and two distinct inward rectifier currents in the photoreceptors ofHermissenda were studied to understand the underlying mechanisms for the electrophysiological differences between type A and B photoreceptors. Here, we report the following. (1) A transient Ca2+ current is expressed in both cells at similar densities, and this current contributes to the generation of action potentials in both cell types. (2) Both type A and B photoreceptors express two inward rectifier currents, a time-dependent (Ih ) current and a time-independent (IK1) current. However,IK1 is predominantly expressed by the type A cells. (3) In contrast, type B photoreceptors express mainlyIh that has kinetic and pharmacological properties that are distinct from IK1. (4)IK1 contributes to membrane hyperpolarization and controls the resting membrane potential of cells during dark adaptation, whereas Ih enhances membrane depolarization and promotes the spontaneous firing properties of the cells in the dark. The additive properties of the Ca2+ current and Ih induce spontaneous activity in the B cells. However, the predominant expression of IK1 in type A cells results in membrane hyperpolarization and virtual quiescence in the dark.
Transient Ca2+ current
Previous reports attest that the photoreceptors express two distinct Ca2+ currents, transient (ICat) and sustained (ICas) currents, and that no significant differences were observed in the level of expression of the two currents in the two cell types (Yamoah and Crow, 1996). The transient current is classified as a T-like current (Nowycky et al., 1985) because it is activated at low voltages, has a rapid onset, is inactivated at a relatively faster rate than is the sustained Ca2+ current, and is selectively blocked by Ni2+. ICat contributes to rhythmic membrane spike activity in the type B photoreceptors, based on the evidence that Ni2+ alters the frequency and phasic properties of spontaneous activity of the cells. The voltage dependence of both the activation and inactivation ofICat revealed the existence of a window current at −50 to −30 mV, suggesting that at rest and during the repolarization phase of a generator potential,ICat may contribute to the total depolarization-induced currents.
Inward rectifiers
Evidence of the distinction betweenIh and IK1 is as follows. (1) Ih was activated with time, whereas activation of IK1 was time-independent. (2) Ih was permeable to both Na+ and K+, whereasIK1 was relatively K+-selective. (3) Ba2+selectively blocked IK1 without having any blocking action on Ih . (4) The kinetics of block of the two currents by monovalent cations was different. From these data, we conclude that two distinct inward-rectifying currents exist in Hermissenda photoreceptors. The existence of K+-selective and Na+/K+-selective inward rectifier currents in seemingly homogeneous cells has been reported in frog hair cells (Holt and Eatock, 1995) and cardiac myocytes (Wu et al., 1991). It is worth noting that in the right atrium of cats, ∼90% of myocytes that exhibit spontaneous pacemaker activity also expressIh -like currents calledIf , whereas approximately the same population of quiescent cells was found to exhibitIK1-like currents (Wu et al., 1991). Presumably, the predominance of Ih -like current is one of the requirements for spontaneous activity, whereas quiescence cells may require IK1-like background current.
The properties of the inward-rectifying currents in the photoreceptors account for some of the characteristic properties of the photoreceptors. Specifically, Ih shares features that are consistent with spontaneous pacemaker activity in the B cells. (1) The activation threshold ofIh (−40 to −50 mV) is compatible with the induction of afterdepolarization during the repolarization phase of action and generator potentials. Also, the permeation properties of the channels (Na+ and K+) and the corresponding Erev ofIh (approximately −40 mV) indicate that at rest (approximately −50 mV) Ih promotes membrane depolarization. (2) The time-dependent activation kinetics of Ih is consistent with the pacing of type B cells after dark adaptation (∼3 Hz); however, as depicted in Figure 6, it is clear that spontaneous activity of the B cells results from the transient Ca2+ current as well. (3) The fact that Ih blocked by Cs+ reduces the frequency of spontaneous activity is in agreement with the hypothesis that Ih contributes to the initiation of spontaneous activity in the B cells. The present hypothesis predicts that in vivo Ih is activated by an ensemble of synaptic inhibitory potentials from adjacent photoreceptors and vestibular neurons (Rogers et al., 1994). Activation ofIh is expected to follow membrane repolarization of action and generator potentials. If inhibitory input from the cells adjacent to type B cells is stronger than that from those adjacent to type A cells, as suggested by Alkon and Fuortes (1972), then Ih in the B cells will be poised to induce an enhanced membrane depolarization to produce tonic activity. Because Ih exhibits no inactivation at potentials close to the resting potential of the B cells (approximately −50 mV), activation ofIh can be cumulative, thus resulting in phasic depolarization of the membrane potential. As seen in Figure6A, after suppressing the transient Ca2+ currents with Ni2+, nonphasic spike activity in the B cells was transformed into a phasic firing pattern that was abolished after application of Cs+. Thus, the properties ofIh can account, at least in part, for the reported properties of the B cells.
Ih recorded from the photoreceptors exhibits many of the properties reported for the current in leech heart interneurons (Angstadt and Calabrese, 1989), neonatal spinal motor neurons (Takahashi, 1990), thalamic relay neurons (Huguenard and McCormick, 1992), and vertebrate photoreceptors (Maricq and Korenbrot, 1990). In addition, there are remarkable similarities between the properties of Ih and those of the currents termed If in cardiac pacemaker cells (Yanagihara and Irisawa, 1980; DiFrancesco, 1985) andIq in hippocampal neurons (Halliwell and Adams, 1982; Maccaferri et al., 1993). Permeation of the channels by Na+ and K+ and sensitivity to Cs+ block are a few examples of the common features shared by these hyperpolarization-activated currents. Furthermore, cells that express these hyperpolarization-activated currents invariably exhibit some form of membrane potential oscillations. Thus, the present findings on the contribution ofIh to type B photoreceptor spontaneous activity are consistent with previous reports on the functions ofIh . Although many of the characteristics of Ih are similar to those reported for hyperpolarizing-activated currents in other systems, the time course of activation was significantly faster than that forIh and/or If in other systems; e.g., at −60 mV, the time constant of activation ofIh in Hermissendaphotoreceptors was ∼300 msec, whereas values closer to 1 sec have been reported in leech interneurons (Angstadt and Calabrese, 1989), and values from 1 to 3 sec have been documented in sinoatrial node cells (Hagiwara and Irisawa, 1989; Zhou and Lipsius, 1992). The time course of activation is expected to affect the beat frequency of spontaneously active cells. However, a direct correlation between the activation of Ih and the frequency of membrane activity will be unlikely in the photoreceptors, where multiple conductances dictate baseline activity. Interestingly, the time course of activation of Ih in photoreceptors is similar to that of Ih in saccular hair cells (Holt and Eatock, 1995). Similar to the photoreceptors, saccular hair cells express two inward-rectifying currents, one Cs+- and the other Ba2+-sensitive (Table1).
Comparison of properties of inward rectifier currents
Inward rectification promotes membrane depolarization if theErev of the current is positive relative to the resting potential of the cell. The time-independent currentIK1 has permeation properties, which are K+ selective, making the Erevmore negative (−70 to −75 mV) than the recorded resting potential of the photoreceptors (approximately −50 mV). In cells that predominantly express IK1, the current acts as a background current that tends to restore the resting potential of the photoreceptors. Relatively high expression ofIK1 in A cells is consistent with the relatively hyperpolarized state of the A cells at rest and the lack of spontaneous activity in the A cells during dark adaptation.
Combined techniques of molecular cloning and electrophysiological studies of cloned channels in heterologous expression systems have identified several types of inward rectifier channels (Bond et al., 1994). These cloned channels exhibit distinct kinetics and pharmacology. It is of interest to correlate the properties of these expressed channels to the native currents recorded in a more physiological system, keeping in mind the possible differences attributable to heteromultimerization and subunit coassembly in the native systems. Emerging data on the inward rectifier currents in both vertebrate and invertebrate systems clearly indicate expression of these currents, namely Ih - andIK1-like currents, in maintaining similar functions in seemingly diverse systems, spanning from invertebrates to vertebrates.
Footnotes
This work was supported by a Grass Foundation fellowship to E.N.Y. and by National Institutes of Health grants to T.C. (MH40860) and L.M. We thank Dr. J. Byrne for the use of his osmometer during the initial phase of the experiments. We also appreciate constructive comments provided by Dr. N. Chiamvimonvat.
Correspondence should be addressed to Dr. Ebenezer N. Yamoah, Department of Cell Biology, Neurobiology, and Anatomy, University of Cincinnati School of Medicine, 231 Bethesda Avenue, Cincinnati, OH 45267.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}