

Abstract
Behavioral sensitization caused by repeated and intermittent administration of psychostimulants, such as cocaine andd-amphetamine, is accompanied by enhanced function in limbic-motor circuitry that is involved in the generation of motivated behavior. The present microdialysis study investigated the effect ofd-amphetamine-induced sensitization on dopamine (DA) efflux in the nucleus accumbens (NAC) of male rats during sexual behavior. Male rats were given one injection of d-amphetamine (1.5 mg/kg, i.p.) or saline every other day for a total of 10 injections. Three weeks after discontinuation of drug treatment, rats were tested for sexual behavior during a test in which microdialysis was performed. There was an augmented efflux of DA in the NAC ofd-amphetamine-sensitized rats compared with nonsensitized control rats when a receptive female was present behind a screen (35 vs 17%). Sensitized rats exhibited facilitated sexual behavior when the screen was removed, as indicated by a significantly shorter latency to mount and an overall increase in the amount of copulatory behavior. Although there was a significant increase in NAC DA concentrations from baseline in both sensitized and nonsensitized rats during copulation, there was a greater increase in DA efflux in the NAC of sensitized rats during the first 10 min copulatory sample (60 vs 37%). These results demonstrate that behavioral sensitization caused by repeated psychostimulant administration can “cross-sensitize” to a natural behavior, such as sex, and that increased NAC DA release may contribute to the facilitation of appetitive and consummatory aspects of this behavior.
- sensitization
- d-amphetamine
- sexual behavior
- motivation
- appetitive
- consummatory
- mesolimbic
- nucleus accumbens
- dopamine
- microdialysis
Previous exposure to psychostimulants, such as d-amphetamine, can result in an enhanced behavioral response to the drug, a phenomenon known as behavioral sensitization. The development and expression of behavioral sensitization is coincident with functional changes in limbic-motor circuitry (Robinson and Berridge, 1993; Self and Nestler, 1995; Pierce and Kalivas, 1997). This circuitry plays an integral role in the expression of motivated or reward-driven behaviors (Phillips et al., 1991; Kalivas et al., 1993; Robbins and Everitt, 1996). It has been hypothesized for some time that the same neural mechanisms that mediate the response to addictive drugs, sexual behavior, and feeding behavior, are functionally related (Robinson and Berridge, 1993; Pierce and Kalivas, 1997).
Although the expression of behavioral sensitization to psychostimulants appears to be the result of complex changes that involve numerous neurotransmitters and nuclei, many studies have reported an enhancement of function within the mesolimbic dopamine (DA) system (Robinson and Berridge, 1993; Self and Nestler, 1995; Pierce and Kalivas, 1997). Indeed, augmented release of DA in the nucleus accumbens (NAC), a terminal area of mesolimbic dopaminergic neurons, is one of the most consistent findings associated with the long-term expression of behavioral sensitization (Robinson and Becker, 1986; Paulson and Robinson, 1995; Pierce and Kalivas, 1997; but see Kuczenski et al., 1997).
Mesolimbic DA plays an important role as a modulator of complex reward processes that organize motivated behaviors, such as drinking, feeding, and sex, by the evaluation of salient environmental stimuli (Blackburn et al., 1992; Kalivas et al., 1993; Kiyatkin, 1995; Salamone, 1996). The incentive sensitization theory of drug addiction proposes that repeated administration of drugs of abuse renders this DA pathway hypersensitive and, in so doing, augments its function of attributing incentive salience to cues associated with reward (Robinson and Berridge, 1993). Recently, we demonstrated that behavioral sensitization to psychostimulants can also enhance the incentive properties of a natural reward (Fiorino and Phillips, 1995).
It is well established that the incentive properties of a sexually receptive female are critical for sexual behavior in inexperienced male rats (Beach, 1941; Madlafousek and Hlinak, 1983). Mesolimbic dopamine is known to play an important facilitatory role in copulation (Everitt, 1990; Pfaus and Phillips, 1991; Mas, 1995; Melis and Argiolas, 1995), and a number of microdialysis studies have reported increased NAC DA efflux associated with both appetitive/motivational and consummatory components of sexual behavior in male rats (Damsma et al., 1992;Wenkstern et al., 1993; Mas, 1995; Fiorino et al., 1997a). In our initial study, previous exposure to d-amphetamine facilitated sexual behavior in sexually inexperienced male rats (Fiorino and Phillips, 1995). Given the hypothesis that this facilitation of sexual behavior is mediated by enhanced release of NAC DA, the present study used microdialysis to determine whether the increase in extracellular NAC DA concentration is augmented during sexual behavior in sexually inexperienced male rats sensitized tod-amphetamine. Subsequently, a d-amphetamine challenge was given to provide independent confirmation that augmented NAC DA efflux is evident in d-amphetamine-sensitized rats.
MATERIALS AND METHODS
The following experiments were conducted in accordance with the standards of the Canadian Council on Animal Care.
Subjects. Male Sprague Dawley rats (225–250 gm) were obtained from the University of British Columbia Animal Care Center and housed individually in plastic cages. The colony room was maintained on a reverse light/dark cycle (lights off 7 A.M. to 7 P.M.). Female Long–Evans rats (Charles River Canada, St. Constant, Quebec, Canada) were housed in a separate colony room on the same reverse light/dark cycle. Ambient temperature was ∼20°C, and rats had ad libitum access to food and water. Testing occurred during the middle third of the dark cycle.
Surgery. Female rats were ovariectomized bilaterally under halothane gas anesthesia (Fluothane; Ayerst Laboratories) at least 4 weeks before testing. Sexual receptivity in the stimulus females was induced by subcutaneous injections of estradiol benzoate (10 μg) and progesterone (500 μg), 48 and 4 hr, respectively, before each test session. All females were sexually experienced before tests of sexual behavior were initiated.
Male rats for each experiment were anesthetized with ketamine hydrochloride (100 mg/kg, i.p.) and xylazine (10 mg/kg, i.p.) before stereotaxic surgery. Microdialysis probe guide cannulae (19 gauge) were implanted over the NAC (coordinates from bregma: anterior, +1.7 mm; medial, −1.1 mm; and ventral, −1.0 mm from dura; flat skull), and secured to the skull with dental acrylic and jeweler’s screws. A 19 gauge wire “training post” was cemented on top of the skull behind the guide cannulae.
Apparatus. Testing was conducted in chambers (48 × 24 × 35 cm) bisected by a removable transparent Plexiglas partition (32 cm), creating two 12 cm wide alleys along the length. Rats could move freely between each side on both ends of the chamber. An antechamber (24 × 8 × 16 cm), which was used to hold an estrous female, was located at one end of the apparatus and separated from the main body of the chamber by a wire mesh screen. Infrared photobeam emitters/detectors allowed the number of activity counts (movements from one side of the partition to the other) to be monitored automatically by a computer (2 Hz scan rate). Chambers were cleaned with a dilute Windex solution between behavioral tests to minimize the influence of residual rat odors.
Induction of behavioral sensitization. Injections were administered in the same chamber that was used for testing of sexual behavior (see below) in each experiment. In each experiment, male rats were assigned randomly to either the repeated d-amphetamine (AMPH) or saline control (CONT) groups. At the beginning of each activity test, a steel coil, which was attached to a liquid swivel (Instech, 375 sec) mounted on top of the testing chamber, was secured to the training post using a microdialysis probe guide collar (see Fiorino et al., 1993 for details). In this manner, rats became accustomed to the apparatus necessary for microdialysis. Rats were placed into testing chambers and, after a 30 min habituation period, given intraperitoneal injections of either d-amphetamine sulfate (1.5 mg/kg; Smith-Kline Beecham, Oakville, ON) or saline vehicle (1 ml/kg; Baxter Corporation, Toronto, Ontario, Canada). Two hours after the injection, rats were returned to their home cages. Injections were given once every 2 d for a total of 10 injections. Activity counts were collected in 10 min bins, and a significant increase from the first to the tenth test was taken as evidence for behavioral sensitization.
Tests of sexual behavior. Before conducting the microdialysis study, a behavioral experiment (experiment 1) was performed on separate groups of rats (n = 10, both groups) to confirm the facilitation of sexual behavior afterd-amphetamine sensitization, within the constraints of the microdialysis procedure. As noted above, all rats were habituated to the coil that connected the head assembly to the liquid swivel, during 10 tests of locomotor activity. Twenty-one days after the last injection of d-amphetamine or saline, sexually naive rats in the AMPH and CONT groups were tested for sexual behavior. The experiment consisted of a 60 min baseline period in the unilevel chamber, after which an estrous female rat was placed in the antechamber that was separated from the main test chamber by a wire screen. After 10 min, the screen was removed, allowing copulation to proceed for 30 min. The female was subsequently removed, and the male rat remained in the chamber for an additional 60 min postcopulation period. Sexual behavior was videotaped and, subsequently, a computer and appropriate software (courtesy of Sonoko Ogawa, Rockefeller University) were used to record standard measures of sexual behavior: (1) mount frequency (MF), (2) intromission frequency (IF), (3) ejaculation frequency (EF), (4) mount and (5) intromission latencies [ML and IL; the time (sec) from the presentation of the female to the first mount or intromission], (6) ejaculation latency [EL; time (sec) from the first intromission to the first ejaculation], (7) number of intromissions to the first ejaculation (IE1), (8) postejaculatory interval [PEI; the time (sec) from the first ejaculation to the next intromission], (9) interintromission interval (III; EL/IE1), and (10) the intromission ratio (IR; IF/(MF + IF)). Criteria for mounts and intromissions are described inSachs and Barfield (1976).
Microdialysis. Rats (n = 10, both groups) were implanted with microdialysis probes 12–18 hr before experiment 2 and placed in the testing chamber with ad libitumaccess to food and water. On the morning of the experiment, microdialysis samples were collected every 10 min. The experiment consisted of five phases: (1) baseline (at least 60 min); (2) estrous female behind the screen (10 min); (3) copulation with estrous female (30 min); (4) postcopulation interval (60 min) after which rats were injected with d-amphetamine (1.5 mg/kg, i.p.); and (5) postinjection period (120 min). Sexual behavior was videotaped and analyzed as described in experiment 1.
Microdialysis procedures and probe characteristics have been reported previously (Fiorino et al., 1997a). The concentrations of microdialysate analytes, which included DA and its metabolites dihyroxyphenylacetic acid (DOPAC) and homovanillic acid (HVA), were assessed by HPLC electrochemical detection methods (Fiorino et al., 1997a). Typical probe recoveries, conducted in vitro and at room temperature, were: for DA, 20.0 ± 0.9%; DOPAC, 15.2 ± 0.9%; and HVA, 14.2 ± 0.6%.
After the microdialysis experiment, animals were given an overdose of chloral hydrate and perfused intracardially with saline and formalin (4%). Frozen brains were sliced, and coronal sections were stained with cresyl violet to determine placement of microdialysis probes. Rats with probe placements within the NAC in experiment 2 were used for behavioral and neurochemical analyses.
Statistics.Activity counts and measures of sexual behavior were assessed using ANOVAs. When a significant main effect was obtained, between-groups comparisons were made using simple main effects analyses. Within-groups post hoc comparisons of total activity counts used the Newman–Keuls test. Neurochemical data were analyzed in the same manner except that Dunnett’s test was used to make within-group comparisons from control means.
A separate statistical analysis of sexual behavior data used Kaplan–Meier plots of both ML and IL, which were assessed by survival analyses (Bloch et al., 1993; Liu et al., 1997). The latencies to mount and intromit and the percentage of subjects showing these behaviors are measures of motivational components of sexual behavior. Both of these measures are used in Kaplan–Meier plots, generated by plotting latencies against the percentage of rats that showed the behavior within the test. This procedure has the added advantage that data from subjects who fail to mount or intromit are not omitted or given an arbitrary value.
Analyses were performed using SPSS and Statistica statistical software packages.
RESULTS
Experiment 1: effect of d-amphetamine sensitization on male rat sexual behavior
Locomotor activity
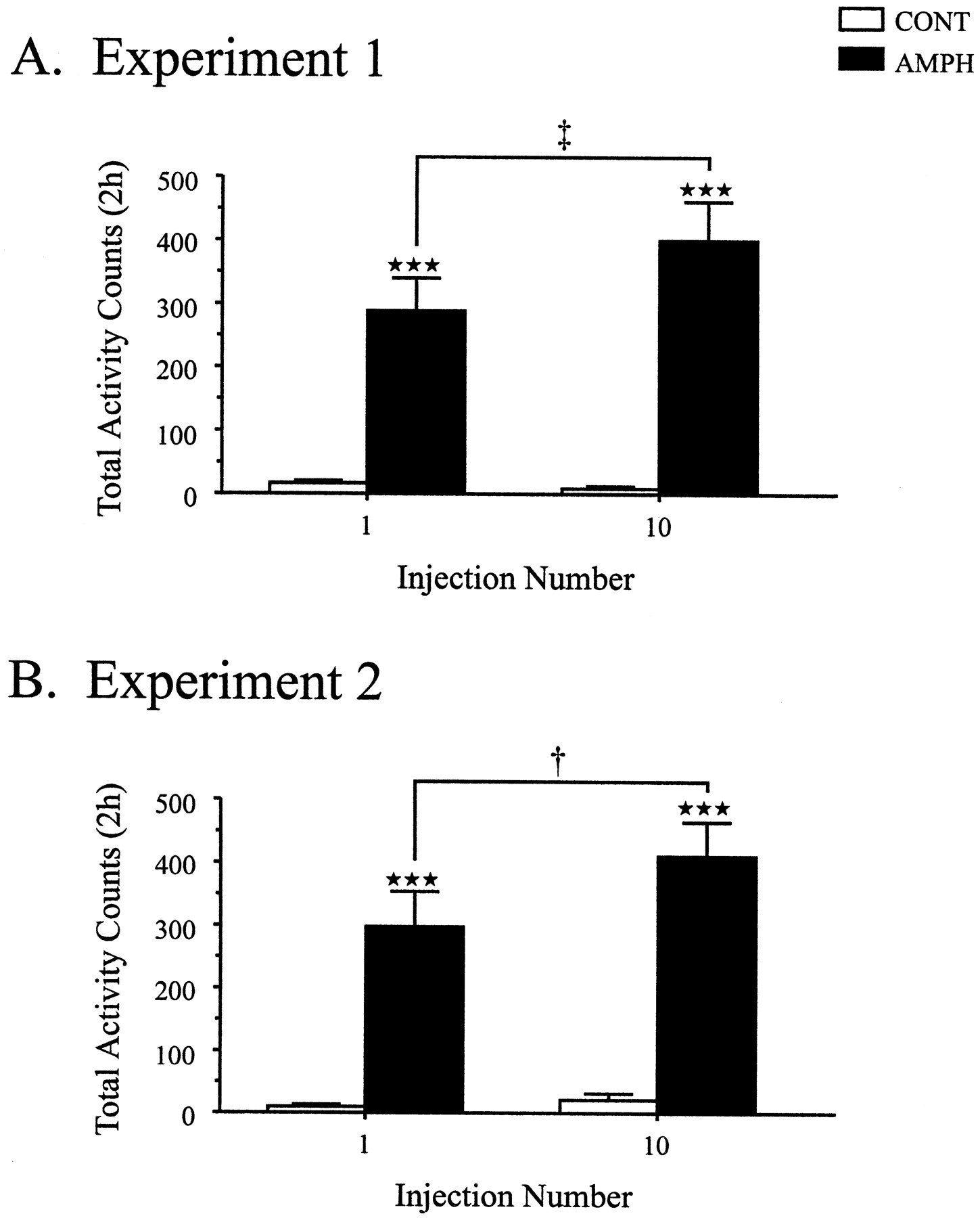
Rats in the AMPH group showed a progressive enhancement ind-amphetamine-induced activity over the course of 10 injections (Fig. 1A). There was a significant increase in the total number of activity counts across injections in the AMPH group (Newman–Keuls; p< 0.01), indicating behavioral sensitization tod-amphetamine.

A, B, Effect of repeated d-amphetamine or saline injections on activity counts accumulated over 2 hr after injection in experiment 1 (A) and experiment 2 (B). The data are represented as mean (± SEM) activity counts. There was a significant interaction between group and injection number in both experiments: experiment 1, F(1,16) = 5.60,p < 0.05; experiment 2,F(1,18) = 12.44, p < 0.01. Between-group comparisons, ***p < 0.001, using simple main effects analysis. Within-group comparisons, †p < 0.01, ‡p < 0.001, using Newman–Keuls post hoc test.
Sexual behavior
Table 1 summarizes measures of sexual behavior in experiment 1. There was facilitation of sexual behavior in the AMPH (n = 10) group relative to the CONT (n = 10) group, as indicated by significantly shorter latencies to mount (F(1,14) = 4.80;p < 0.05) and intromit (F(1,14)= 5.92; p < 0.05). Survival analyses of Kaplan–Meier curves for mount latency (Fig.2A, left panel) and intromission latency (Fig.2A, right panel) confirmed a facilitation of behavior in AMPH-treated rats (Log rank statistic = 9.43, p = 0.0021; Log rank statistic = 10.48,p = 0.0012, respectively).
Measures of sexual behavior from experiments 1 and 2

Effect of d-amphetamine or saline pretreatment on latencies to mount and intromit. A, Kaplan–Meier curve for mount latency (left panel) and intromission latency (right panel) in experiment 1. Independent survival analyses showed a significant difference between sensitized (AMPH) and nonsensitized (CONT) rats with respect to mount latency (Log rank statistic = 9.43; p = 0.0021) and intromission latency (Log rank statistic = 10.48; p = 0.0012). B, Kaplan–Meier curve for mount latency (left panel) and intromission latency (right panel) in experiment 2. Independent survival analyses showed a significant difference between sensitized (AMPH) and nonsensitized (CONT) rats with respect to mount latency (Log rank statistic = 6.12; p = 0.0134) and intromission latency (Log rank statistic = 4.38;p = 0.0364).
There was an overall increase in the amount of sexual behavior caused by previous d-amphetamine exposure, as indicated by a significantly greater number of intromissions (F(1,16) = 5.89; p < 0.05) and ejaculations (F(1,16) = 6.37; p< 0.05) over the 30 min copulation period as compared with the CONT group. However, other measures from the first ejaculatory series once copulation was initiated, such as EL, PEI, IE1, III, and IR, were not different between groups.
Experiment 2: dopamine efflux in the NAC and sexual behavior afterd-amphetamine-induced sensitization
Locomotor activity
Repeated d-amphetamine administration again induced behavioral sensitization in AMPH rats (Fig.1B). Rats in the AMPH group displayed a sensitized behavioral response to d-amphetamine, as indicated by a significant increase in the total number of activity counts from injection 1 to injection 10 (Newman–Keuls; p < 0.05).
Sexual behavior
Measures of sexual behavior in experiment 2 (n = 8, both groups) are shown in Table 1. Although the AMPH group displayed shorter latencies to mount and intromit, these measures did not differ significantly from those of the CONT group. However, survival analyses of Kaplan–Meier curves for ML and IL (see Fig. 4A) did reveal a significant enhancement of sexual behavior in AMPH rats (Log rank statistic = 6.12,p = 0.0134; Log Rank statistic = 4.38,p = 0.0364, respectively). The AMPH group also achieved more ejaculations within the 30 min copulatory period than the CONT group (F(1,14) = 5.56; p < 0.03). Generally, the rats in experiment 2 displayed a more proficient level of sexual behavior than rats of experiment 1, with respect to latencies to mount, intromit, and ejaculate, as well as the overall amount of copulation.
Neurochemistry of sexual behavior
Previous exposure to d-amphetamine did not alter basal concentrations of DA, DOPAC, or HVA (Table2). There was an overall change in NAC DA efflux associated with sexual behavior (Fig.3). Specifically, concentrations of DA in AMPH rats were significantly elevated relative to baseline throughout all phases of the test session, including the period when the female was present behind a screen, during copulation, and for the 20 min period after her removal. In contrast, significant increases in DA concentrations in CONT rats were restricted to samples associated with copulation. With respect to the different phases of the test, presentation of a receptive female behind the screen (Scr; time = 20 min) resulted in a significant increase in NAC DA concentrations from baseline values in the AMPH group (+35%; p < 0.01) but not in the CONT group (+17%). There was a further increase in DA efflux during copulation in both groups. In AMPH rats, DA reached maximum concentrations during the first copulatory sample (time = 30 min; +60%), and NAC DA remained elevated for 20 min after removal of the female. Maximum DA concentrations in the CONT group occurred in the second 10 min copulatory sample (time = 40 min; +47%).
Mean basal concentrations of analytes corresponding to 100% baseline

Changes in nucleus accumbens dopamine efflux (line graph) during baseline (Bas), while a receptive female was present behind the screen (Scr), during copulation, and after copulation for CONT and AMPH rats. Bar graphs show the number of mounts plus intromissions (top bar graph) and ejaculations (bottom bar graph) displayed for each group during three 10 min samples. *p < 0.05; **p < 0.01 using simple main effects analysis. Within-group Newman–Keuls post hoc tests revealed significant (p < 0.05) increases in nucleus accumbens DA concentrations from baseline in AMPH (time = 20–50 min) and CONT (time = 30–50 min) rats.
The increase in NAC DA in the AMPH group was significantly greater than the increase obtained from the CONT group, both when the female was behind the screen (time = 20 min) (F(1,182)= 4.76; p < 0.05) and during the first 10 min of copulation (time = 30 min) (F(1,182) = 6.32; p < 0.05). The significant augmentation of NAC DA efflux in the AMPH group (time = 30 min) coincided with an increased number of mounts plus intromissions (F(1,28) = 18.56; p < 0.01) and ejaculations (F(1,18) = 5.09; p< 0.05) relative to the CONT group.
Extracellular concentrations of DOPAC and HVA also increased after copulation (Fig. 4). A significant increase in DOPAC concentrations relative to baseline occurred during the first copulatory period in both groups and remained significantly elevated for 70 min after injection in CONT rats (maximum, time = 40 min; +40%) and 60 min after injection in AMPH rats (maximum, time = 40 min; +55%). Increases in the concentration of HVA from baseline achieved statistical significance in the first 10 min period of copulation for the AMPH group (maximum, time = 60 min; +50%) and remained significantly elevated for 30 min after copulation. However, HVA concentrations in AMPH rats were still elevated just before the d-amphetamine challenge (+28%). Concentrations of HVA in CONT rats were significantly elevated from the last 10 min copulatory period (time = 50 min) until thed-amphetamine injection (time = 110 min), and reached a maximum increase of +49% (time = 70 min). There was no statistical difference in metabolite concentrations between groups at any point in the experiment.

Changes in DOPAC (top panel) and HVA (bottom panel) concentrations in the nucleus accumbens during baseline (Bas), while a receptive female was present behind the screen (Scr), during copulation, and after copulation. Within-group Newman–Keulspost hoc tests revealed significant (p < 0.05) increases in nucleus accumbens metabolite concentrations from baseline in AMPH (DOPAC, time = 30–80 min; HVA, time = 30–90 min) and CONT (DOPAC, time = 30–90 min; HVA, time = 50–100 min) rats.
Behavior and neurochemistry after ad-amphetamine challenge
Systemic administration of d-amphetamine (1.5 mg/kg) resulted in a sensitized behavioral and neurochemical response by the AMPH group relative to the CONT group (Fig.5). A repeated measures ANOVA on activity counts revealed a significant effect of group (F(14,196) = 28.50; p < 0.01). The increase in activity counts after d-amphetamine administration persisted for 2 hr after the injection in both groups relative to baseline, as assessed by Newman–Keuls post hoctests. The AMPH group displayed a significantly greater number of activity counts than the CONT group in the first 30 min after the drug injection and with respect to total activity counts after injection (413.5 ± 37.7 vs 303.6 ± 37.8) (F(1,14) = 4.84; p < 0.05).

Changes in nucleus accumbens dopamine efflux in response to a d-amphetamine challenge (1.5 mg/kg, i.p.). *p < 0.05; **p < 0.01 using simple main effects analysis. Dopamine concentrations and activity counts remained elevated throughout the 2 hr postinjection period relative to baseline (Bas) in both groups.
There was a concomitant increase in extracellular NAC DA after injection in the CONT and AMPH groups (F(14,196)= 39.16; p < 0.01), and the increase observed in the AMPH group was significantly greater than the CONT group in the first 40 min after injection (Fig. 5, top panel). Both metabolites of DA decreased after injection in the CONT and AMPH groups. Systemic administration of d-amphetamine resulted in a significant decrease in DOPAC efflux 10 min after injection, reaching a minimum concentration of 48% in the CONT group and 44% in the AMPH group (time = 50 min, both groups). The maximum decrease in HVA concentrations was delayed further, reaching significantly depressed values 40 and 50 min after injection in CONT (minimum, time = 80 min; 82%) and AMPH (minimum, time = 100 min; 83%) rats, respectively. There were no differences between groups in DA metabolite concentrations at any point after d-amphetamine administration.
The disparate changes in extracellular NAC DA metabolite concentrations in response to sexual incentives andd-amphetamine administration (i.e., increase and decrease, respectively) during a period when DA efflux invariably increased, once again emphasizes the inherent problems associated with inferring DA transmission by changes in DA metabolite efflux (Fiorino et al., 1997a;O’Neill et al., 1998).
Histology
Microdialysis probes tracts were found in both the shell and core regions of the NAC in a range extending from +1.60 to +2.20 mm from bregma (Fig. 6).

Location of microdialysis probes within the nucleus accumbens of rats used in experiment 2. Vertical black lines correspond to the location of the active fiber area of the microdialysis probes. For reasons of clarity, probe placements of CONT rats are displayed on the left, and those of AMPH rats are shown on the right. Coronal brain sections redrawn from Paxinos and Watson (1997).
DISCUSSION
Repeated and intermittent exposure to d-amphetamine, sufficient to induce behavioral sensitization, facilitated sexual behavior in sexually inexperienced male rats, thus confirming our previous observations (Fiorino and Phillips, 1995). This effect was replicated again in the microdialysis experiment, which showed clearly that enhanced sexual behavior was correlated with augmented NAC DA efflux. The results of the present experiments not only provide support for the role of mesolimbic DA in motivated behavior, but buttress the hypothesis that changes in limbic-motor circuitry, specifically to mesolimbic dopaminergic pathways, contribute to sensitized behavior in response to both psychostimulant administration and natural incentives.
An earlier study found that sexual behavior was facilitated in male rats when tested in an environment that had been paired repeatedly with systemic morphine injections (Mitchell and Stewart, 1990). Specifically, there was a preferential enhancement in motivational measures, such as the amount of anogenital exploration, the percentage of animals copulating, and latency to mount, rather than indices of copulation itself. Although the aim of that study was to investigate whether sexual behavior could be enhanced by a conditioned association between environmental cues and opiate reward, the injection regimen used has been reported to induce behavioral sensitization to the locomotor-activating effects of morphine (Kalivas and Stewart, 1991). The present experiments directly examined the influence of behavioral sensitization on sexual behavior in male rats and found a similar facilitation of motivational components of sexual behavior, including mount and intromission latencies. Although the overall amount of copulation was increased in sensitized rats, as indicated by a greater EF and IF in the 30 min copulation period (experiment 1) and the number intromissions plus mounts and ejaculations during the first 10 min of copulation (experiment 2), consummatory measures within bouts of copulation, such as IE1, III, IR, and EL were not significantly altered. The preferential effect of sensitization on appetitive aspects of sexual behavior confirms previous observations (Fiorino and Phillips, 1995). In contrast to the study by Mitchell and Stewart (1990), we have found this effect to be independent of the context in which rats received drug injections (our unpublished observations). It should be noted that, despite the fact that all rats in the AMPH group copulated in the present experiments,d-amphetamine sensitization, induced by this injection regimen, does not guarantee that sexually naive rats will copulate (Fiorino and Phillips, 1995). Nevertheless, we have observed that a very high percentage of rats preexposed to d-amphetamine will copulate during their first test of sexual behavior (i.e., >85%).
Previous reports have noted that increases in medial preoptic area (mPOA) or NAC DA efflux in response to a receptive female behind a screen were only observed in male rats that also copulated once the screen was removed (Hull et al., 1995; Wang et al., 1995; Fiorino et al., 1997a). The same relationship was observed in the present study; in the case of an individual rat, a precopulatory increase in NAC DA efflux was predictive of subsequent copulatory behavior. There was no significant appetitive increase in NAC DA efflux in the CONT group. However, two CONT rats (25%) did not copulate, and the corresponding change in mean NAC DA concentrations in response to the receptive female in the CONT group was attenuated. Our observation that increased mesolimbic DA transmission was associated with copulation in naive rats is in agreement with the results of an earlier microdialysis experiment (Wenkstern et al., 1993). The present results demonstrate that increases in NAC DA concentrations in response to sexual incentives occur in sexually naive male rats.
There was a significant augmentation of NAC DA efflux in the AMPH group relative to the CONT group during sexual behavior, and this was evident during both the appetitive phase (i.e., female behind the screen) and the first copulatory sample. As mentioned above, noncopulating CONT rats did not contribute to the appetitive or copulatory rise in NAC DA concentrations. Therefore, the increased amount of copulation observed during the first copulatory sample in AMPH rats relative to CONT rats, and in particular the greater number of ejaculations, may explain the augmented efflux of NAC DA in the AMPH group. Chronoamperometry experiments conducted in our laboratory have shown that peak oxidation currents associated with DA are correlated with ejaculations (Phillips et al., 1991; Fiorino et al., 1997b), although it is difficult to determine whether ejaculation or the vigorous pursuit activity leading to ejaculation is correlated with maximal NAC DA efflux. In this regard, it is important to note that the “consummatory” phase of male rat sexual behavior (i.e., copulation) contains many appetitive components (Fiorino et al., 1997a), and it is impossible, in the present study, to correlate NAC DA efflux preferentially with one component or the other. Nevertheless, the intense behavior of sensitized rats during the first copulatory sample, relative to CONT rats, may account for differences in neurochemical profiles between groups.
The presence of augmented dopaminergic transmission in the NAC induced by repeated d-amphetamine administration lends support to the observation that enhanced dopaminergic activity can facilitate the initiation of sexual behavior in sexually naive male rats (Agmo and Picker, 1990). Dopamine in the mPOA is involved in both appetitive and consummatory aspects of male rat sexual behavior (Pfaus and Phillips, 1991; Hull et al., 1993, 1995; Shimura et al., 1994; Mas et al., 1995;Sato et al., 1995), and enhanced mPOA DA transmission may have contributed to facilitation of sexual behavior observed in the present study.
A systemic d-amphetamine challenge also resulted in augmented locomotor behavior and NAC DA efflux in the AMPH group relative to the CONT group. This finding contributes to the growing literature demonstrating that increased striatal DA transmission accompanies behavioral sensitization to psychostimulants, when assessed after extended periods (>14 d) after discontinuation of drug treatment (for review, see Pierce and Kalivas, 1997; but see Kuczenski et al., 1997). Caution must be exercised, however, in comparing these results with previous reports because the present drug challenge was administered after a period of sexual activity, and residual sex-related odors may have contributed, perhaps differentially, to neurochemical responses in both groups. Bedding from the cages of estrous females has been reported to increase extracellular NAC DA in male rats (Mitchell and Gratton, 1992).
Our results are consistent with the incentive-sensitization theory of drug addiction (Robinson and Berridge, 1993), which proposes that the incentive value of cues associated with reward is enhanced because of repeated administration of psychostimulants via an augmentation of mesotelencephalic DA function; enhanced mesotelencephalic DA transmission ultimately contributes to compulsive drug seeking and drug taking, which are defining features of drug addiction. Previous experiments with drug reward found that preexposure of animals to psychostimulants facilitated various measures of self-administration of drugs of abuse (Woolverton et al., 1984; Horger et al., 1990, 1992;Piazza et al., 1990; Mendrek et al., 1998; but see Li et al., 1994). In a recent study, Mendrek et al. (1998) demonstrated thatd-amphetamine-sensitized rats exhibited increased motivation to self-administer d-amphetamine, as indicated by a significantly higher break point under a progressive ratio schedule of reinforcement. The present study extends the enhancement of motivated behaviors caused by repeated psychostimulant administration to those elicited by natural incentives. In this case, previousd-amphetamine treatment may have amplified the importance of unconditioned incentive cues of the estrous female, which included pheromones, ultrasonic vocalizations, ear wiggling, and darting, and led to a facilitation of sexual behavior. The presence of augmented NAC DA transmission in response to an estrous female located behind a screen strengthens the argument that the mesolimbic DA system contributes to this effect.
Cross-sensitization between DA uptake blockers and sexual behavior is also supported by clinical observations. Bupropion, an antidepressant drug that can block DA uptake (Cooper et al., 1980; Nomikos et al., 1989), enhanced sexual function in male and female patients treated for sexual dysfunction (Crenshaw and Goldberg, 1995) and psychiatric disorders (Modell et al., 1997). It is interesting that preclinical studies have shown that long-term (10 mg/kg, b.i.d. × 21 d), but not acute (10 mg/kg, b.i.d. × 2 d), administration of bupropion results in augmented NAC DA efflux in response to a bupropion challenge (Nomikos et al., 1989, 1992). In contrast, there was no significant augmentation of DA transmission observed in the striatum after chronic bupropion treatment (Nomikos et al., 1992). The development of the prosexual effects of bupropion may be a consequence of neural changes similar to those involved in the initiation and expression of behavioral sensitization to psychostimulants. Therefore, the ability of a compound to induce long-term functional enhancement in the mesolimbic DA system may provide valuable information about its potential to treat sexual dysfunction.
Footnotes
This work was supported by Group Grant PG-12808 from the Medical Research Council of Canada. We thank David Mutch for his help in conducting these experiments and Fred LePiane and Keith Waldron for their assistance in construction of testing chambers. Also, many thanks to Ariane Coury and Jim Pfaus for their helpful comments and Liz McCririck for her secretarial services.
Correspondence should be addressed to Dr. A. G. Phillips, University of British Columbia, Department of Psychology, 2136 West Mall, Vancouver, British Columbia, Canada, V6T 1Z4.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}