

Abstract
Members of the muscarinic acetylcholine receptor family are thought to play key roles in the regulation of a large number of important functions of the CNS. However, the precise roles of the individual muscarinic receptor subtypes in modulating these processes are not well understood at present, primarily because of the lack of ligands with sufficient receptor subtype selectivity. To investigate the behavioral significance of the M1 muscarinic receptor (M1R), which is abundantly expressed in the forebrain, we subjected M1 receptor-deficient mice (M1R−/− mice) to a battery of behavioral tests. M1R−/− mice showed no significant impairments in neurological reflexes, motor coordination, pain sensitivity, and prepulse inhibition. Strikingly, however, M1R−/− mice consistently exhibited a pronounced increase in locomotor activity in various tests, including open field, elevated plus maze, and light/dark transition tests. Moreover,M1R−/− mice showed reduced immobilization in the Porsolt forced swim test and reduced levels of freezing after inescapable footshocks, suggesting thatM1R−/− mice are hyperactive under stressful conditions as well. An increased number of social contacts was observed in a social interaction test. Surprisingly,M1R−/− mice displayed no significant cognitive impairments in the Morris water maze and in contextual fear conditioning.M1R−/− mice showed slight performance deficits in auditory-cued fear conditioning and in an eight-arm radial maze, most likely because of the hyperactivity phenotype displayed by theM1R−/− mice. Our results indicate that M1 muscarinic receptors play an important role in the regulation of locomotor activity but appear to be less critical for cognitive processes, as generally assumed.
- acetylcholine
- muscarinic receptor
- M1receptor
- hyperactivity
- learning
- gene targeting
- knock-out
- behavioral phenotyping
Molecular cloning studies have revealed the existence of five distinct mammalian muscarinic acetylcholine receptor subtypes (M1–M5) (Caulfield, 1993;Wess, 1996). Muscarinic receptors are known to play key roles in many functions of the CNS, including the control of locomotor activity, emotional behavior, and higher cognitive processes such as learning and memory (Wess et al., 1990; Brown and Taylor, 1996). The identification of specific muscarinic receptor subtype(s) involved in these diverse functions is complicated by the fact that muscarinic ligands with high selectivity for the individual receptor subtypes are not available at present and most brain areas express multiple muscarinic receptor subtypes (Levey, 1993; Vilaro et al., 1993; Wess, 1996).
The M1 muscarinic receptor (M1R) is abundantly expressed in higher brains regions, including cerebral cortex, hippocampus, amygdala, and striatum (Buckley et al., 1988; Levey et al., 1991; Levey et al., 1995). Nonselective muscarinic antagonists, such as scopolamine and atropine, impair performance in various learning and memory tasks in rodents, such as the eight-arm radial maze test (Eckerman et al., 1980; Okaichi and Jarrard, 1982), contextual fear conditioning (Anagnostaras et al., 1995; Rudy, 1996), and the Morris water maze (Sutherland et al., 1982;Whishaw et al., 1985). Similar impairments have been observed after administration of muscarinic antagonists with limited M1R selectivity or of an M1R antisense oligodeoxynucleotide (Hagan et al., 1987; Roldan et al., 1997; Ghelardini et al., 1999; Fornari et al., 2000). On the basis of these observations and the finding that Alzheimer's disease is associated with a progressive loss of cholinergic input into higher brain regions, it has been proposed that M1R agonists might become useful in ameliorating the cognitive deficits associated with this disease (Coyle et al., 1983; Quirion et al., 1989; Fisher et al., 1996; Iversen, 1997).
M1Rs are abundantly expressed in the striatum (Weiner et al., 1990; Levey et al., 1991; Bernard et al., 1992; Hersch et al., 1994), and muscarinic receptor antagonists with high affinity for M1Rs are clinically useful in the treatment of Parkinson's disease (Fahn et al., 1990; Standaert and Young, 1996). It has therefore been proposed that the M1R subtype may play an important role in the regulation of extrapyramidal locomotor function (Levey et al., 1991). Consistent with this idea, pharmacological blockade of muscarinic receptors produces pronounced hyperactivity in rodents (Whishaw et al., 1985; Toide, 1989; Sipos et al., 1999).
Several lines of evidence indicate that muscarinic receptors including the M1R subtype may also play a role in certain aspects of schizophrenia (Bymaster et al., 1999), nociception (Bartolini et al., 1992; Ghelardini et al., 2000), anxiety-like behaviors (Smythe et al., 1996; File et al., 1998), and depressive disorders (Daws et al., 1991; Chau et al., 1999).
To gain more direct insight into the physiological roles of the M1R subtype, we have subjectedM1R−/− mice to a comprehensive battery of behavioral tests, covering sensory/motor functions, emotional behaviors, and learning abilities. Our data indicate that the lack of M1Rs is associated with a pronounced hyperactivity phenotype but does not lead to major learning deficits.
MATERIALS AND METHODS
M1 muscarinic receptor mutant mice. The M1R gene was disrupted in mouse TC1(129SvEv) embryonic stem cells, andM1R−/− mice were generated by standard techniques (A. Fisahn, M. Yamada, A. Duttaroy, C. Deng, C. McBain, and J. Wess, unpublished data). M1R function was abolished by replacing a genomic fragment that included the translation start site and the region coding for the first 54 amino acids of the M1R protein with a PGK-neomycin resistance cassette. All experiments were performed with male F2 littermates (C57BL/6J × 129SvEv hybrids).
Radioligand binding studies. Mouse striata were removed, dissected, frozen immediately on dry ice, and stored at −70°C until use. Tissues were homogenized by hand with 20 strokes of a Dounce tissue grinder in 0.32 m sucrose, 5 mm Tris-HCl, pH 7.5, and 1 mm phenylmethylsulfonyl fluoride. Membranes were prepared, and ligand binding experiments were performed using a saturating concentration (2 nm) of the nonselective muscarinic antagonist, [3H]quinuclidinyl benzilate ([3H]QNB), essentially as described (Dorje et al., 1991). Binding reactions were performed for 1 hr at room temperature (22°C). Nonspecific binding was determined in the presence of 10 μm atropine.
Immunoprecipitation assays. For immunoprecipitation studies, M1–M5 receptor-specific rabbit polyclonal antisera were raised against nonconserved regions of the third cytoplasmic loops of the mouse M1–M5 receptor proteins according to Levey et al. (1991). Membranes derived from mouse striata were prepared as described above, incubated with 2 nm [3H]QNB, washed thoroughly, and solubilized with 1% digitonin, followed by immunoprecipitation of solubilized [3H]QNB-labeled receptors (Gomeza et al., 1999a; Yamada et al., 2001).
Animals and experiment design. All behavioral tests were performed with male mice that were 11 weeks old at the start of the testing (M1R−/− mice,n = 25; wild-type littermates, n = 21). Mice were housed four to five per cage in a room with a 12 hr light/dark cycle (lights on at 6:00 A.M.) with ad libitumaccess to food and water (except for the period during which the radial maze test was conducted). Behavioral testing was performed between 9:00 A.M. and 3:00 P.M. The neurological screen, light/dark transition, open field, hot plate, rotarod, wire hang, elevated plus maze, eight-arm radial maze, prepulse inhibition, contextual and cued fear conditioning, social interaction, Morris water maze, and Porsolt swim tests were conducted in this sequence, with each test separated at least by 2 d. All behavioral testing procedures were approved by the Vanderbilt University Institutional Animal Care and Use Committee.
Neurological screen. A neurological screen was conducted as described previously (Miyakawa et al., 2001). The righting, whisker touch, eye blink, and ear twitch reflexes were evaluated. A number of physical features, including the presence of whiskers and bald patches, were also recorded.
Motor function tests. Motor coordination and balance were tested with the rotarod test, and neuromuscular strength was tested with the wire hang test, as described previously (Miyakawa et al., 2001). The rotarod test using an accelerating rotarod (UGO Basile Accelerating Rotarod) was performed by placing a mouse on a rotating drum (3 cm diameter) and measuring the time each animal was able to maintain its balance on the rod. The speed of the rotarod accelerated from 4 to 40 rpm over a 5 min period. In the wire hang test, the mouse was placed on a wire cage lid that was then inverted and gently waved in the air, so that the subject gripped the wire. Latency to fall onto the bedding was recorded, with a 60 sec cutoff time.
Open field test. Locomotor activity was measured using an open field test. Each subject was placed in the center of the open field apparatus (27.4 × 27.4 × 20 cm; MED-associates, Albans, VT). The maze was cleaned with water after each trial. Horizontal activity (in centimeters), vertical activity (rearing measured by counting the number of photobeam interruptions), time spent in the center, and number of fecal boli were recorded. Data were collected for 30 min.
Light/dark transition test. The apparatus used for the light/dark transition test consisted of a cage (27.4 × 27.4 × 20 cm) equally divided into two by a black partition containing a small opening (MED-associates). One chamber was open and brightly illuminated, whereas the other chamber was closed and dark. Mice were placed into the lit side and allowed to move freely between the two chambers for 10 min. The chambers were cleaned with water after each trial. The total number of transitions, time spent in the dark side, and horizontal activity (in centimeters) were recorded.
Elevated plus-maze. The elevated plus-maze consisted of two open arms (30 × 5 cm) and two enclosed arms of the same size, with 15-cm-high transparent walls. The arms and central square were made of white plastic plates and were elevated to a height of 50 cm above the floor. To minimize the likelihood of animals falling from the apparatus, 3-mm-high Plexiglas ledges were provided for the open arms. Arms of the same type were arranged at opposite sides to each other. Each mouse was placed in the central square of the maze (5 × 5 cm), facing one of the open arms. Mouse behavior was recorded during a 10 min test period. The maze was cleaned with water after each trial. The number of entries onto and the time spent on open and enclosed arms were recorded. For data analysis, we used the following four measures: the percentage of open arm entries, the percentage of time spent on the open arms, the total number of arm entries, and total distance traveled (centimeters). Data acquisition and analysis were performed automatically, using Image EP software (see Image analysis).
Pain test. The hot plate test was used to evaluate the sensitivity to a painful stimulus. Mice were placed on a 55.0 (±0.3)°C hot plate (Columbus Instruments, Columbus, OH), and latency to the first hindpaw response was recorded. The hindpaw response was either a foot shake or a paw lick.
Social interaction test. M1R+/+ orM1R−/− mice were introduced into a novel box (40 × 40 × 30 cm) with a male DBA/2J mouse and allowed to explore freely for 10 min. Social behavior was monitored by a color charge-coupled device camera (Sony DXC-151A) that was connected to a Macintosh computer. Analysis was performed automatically using Image SI software (see Image analysis). The software can discriminate between a mouse with a darker coat color and one with a brighter coat color. The number of active contacts initiated by the target animal (i.e., M1R+/+ or M1R−/− mice), mean duration per contact, total duration of contact, and total distance traveled were measured. The number of active contacts was defined as follows. Images were captured at one frame per second, and the distance traveled between two successive frames was calculated for each mouse. If the two mice contacted each other and the distance traveled by either mouse was longer than 5 cm, the behavior was considered to be “active contact.” The mouse that traveled a longer distance from the previous frame was considered to have approached the other subject actively.
Startle response/prepulse inhibition tests. A startle reflex measurement system was used (MED-associates). A test session began by placing a mouse in a Plexiglas cylinder where it was left undisturbed for 5 min. The duration of white noise that was used as the startle stimulus was 40 msec for all trial types. The startle response was recorded for 160 msec (measuring the response every 1 msec) starting with the onset of the prepulse stimulus. The background noise level in each chamber was 70 dB. The peak startle amplitude recorded during the 160 msec sampling window was used as the dependent variable. A test session consisted of six trial types (i.e., two types for startle stimulus-only trials and four types for prepulse inhibition trials). The intensity of startle stimulus was 110 or 120 dB. The prepulse sound was presented 100 msec before the startle stimulus, and its intensity was 74 or 78 dB. Four combinations of prepulse and startle stimuli were used (74–110, 78–110, 74–120, and 78–120). Six blocks of the six trial types were presented in pseudorandom order such that each trial type was presented once within a block. The average intertrial interval was 15 sec (range, 10–20 sec).
Eight-arm radial maze test. The eight-arm radial maze test was conducted in a manner similar to that described previously (Miyakawa et al., 1996). The floor of the maze was made of white Plexiglas, and the wall (16 cm high) consisted of transparent Plexiglas. Each arm (9 × 50 cm) radiated from an octagonal central starting platform (perimeter 12 × 8 cm) like the spokes of a wheel. Identical food wells (1.4 cm deep and 1.4 cm in diameter) with pellet sensors were placed at the distal end of each arm. The pellet sensors were able to automatically record pellet intake by the mice. The maze was elevated 35 cm above the floor and placed in a dimly lit room with several extra-maze cues. During the experiment, the maze was maintained in a constant orientation. One week before pretraining, animals were deprived of food until their body weight was reduced to 80–85% of the initial level. Pretraining started on the eighth day. Each mouse was placed in the central starting platform and allowed to explore and to consume food pellets scattered on the whole maze for a 5 min period (one session per mouse). After completion of the initial pretraining, mice received another pretraining to take a pellet from each food well after being placed at the distal end of each arm. A trial was finished after the subject consumed the pellet. This was repeated eight times, using eight different arms, for each mouse. After these pretraining trials, actual maze acquisition trials were performed. All eight arms were baited with food pellets. Mice were placed on the central platform and allowed to get all eight pellets within 15 min. A trial was terminated immediately after all eight pellets were consumed or 15 min had elapsed. An “arm visit” was defined as traveling for >5 cm from the central platform. The mice were confined in the center platform for 5 sec after each arm choice. The animals went through 1 trial per day (18 trials total). For each trial, choices of arms, latency to get all pellets, distance traveled, the number of different arms chosen within the first eight choices, and the number of revisiting and omission errors were automatically recorded.
During the 15th acquisition trial, a 30 sec delay was initiated after four pellets had been taken by confining the mice in the center platform. From the 16th to the 18th acquisition trial, the delay period was extended to 2 min. After each trial, the maze was cleaned with water. The locations of the maze arms were randomly relocated after each session to prevent animals from using intra-maze cues. Data acquisition, control of guillotine doors, and data analysis were performed by Image RM software (see Image analysis).
Contextual and cued fear conditioning. Each mouse was placed in a test chamber (20 × 16 × 36 cm) inside a sound-attenuated chamber (MED-associates) and allowed to explore freely for 2 min. A 4.5 kHz pure tone, which served as the conditioned stimulus (CS), was presented for 30 sec, followed by a mild (2 sec, 0.5 mA) footshock, which served as the unconditioned stimulus (US). Two more CS–US pairings were presented with 2 min interstimulus intervals. Context testing was conducted 24 hr and 4 weeks after conditioning in the same chamber. Cued testing with altered context was conducted 48 hr after conditioning using a triangular box (25 × 25 × 30 cm) made of white opaque Plexiglas that was located in a different room. Banana extract odor was added to further change the context. Data acquisition, control of stimuli (i.e., tones and shocks), and data analysis were performed automatically, using Image FZ software (see Image analysis). Images were captured at one frame per second. For each pair of successive frames, the amount of area (pixels) within which the mouse moved was measured. When this area was below a certain threshold (i.e., 10 pixels), the behavior was judged as “freezing.” When the amount of area equaled or exceeded the threshold, the behavior was considered to be “nonfreezing.” The optimal threshold (amount of pixels) by which to judge freezing was determined by adjusting it to the amount of freezing measured by human observation. A frame capturing rate of one frame per second yielded results that were in good agreement with those obtained by human observation (correlation between human observation and computer generated data; r > 0.95). Freezing that lasted less than the defined time threshold (i.e., 2 sec) was not included in the analysis. To assess mouse reactivity to footshocks, distance traveled during shock presentation was assessed by capturing images at 2.5 frames per second for 6 sec from 2 sec before shock presentation to 2 sec after shock termination.
Morris water maze test. A “hidden platform” version of the Morris water maze test was conducted to assess spatial learning ability, as described previously (Miyakawa et al., 2001). The apparatus consisted of a circular tank (40 cm high × 95 cm diameter) filled with water (up to 30 cm deep) maintained at room temperature (23°C) and made opaque with nontoxic white paint. The surface of the platform (8 × 8 cm) was 1 cm below the water surface. Four trials per day were conducted for 6 successive days with the original platform location and for 5 successive days with a new platform location. There were four possible locations for the platform. One of these platform positions was assigned to each mouse as correct location during the original training (blocks 1–6). During reversal training (blocks 7–11), the platform location was rotated by 180° from the original platform location. Latency to reach the platform, distance traveled to reach the platform, average swim speed, and percentage of time spent at the perimeter of the pool were recorded. When the distance between mice and the wall of the pool was <8 cm, mice were considered to be at the perimeter. On the 6th and 11th days of training, the platform was removed, and a 60 sec probe trial was conducted. Time spent in each quadrant, number of crossings above the former target site, average swim speed, and percentage of time spent at the perimeter of the pool were recorded during the probe trials. Data acquisition and analysis were performed using Image WM software (see Image analysis).
Porsolt forced swim test. The apparatus consisted of two Plexiglas cylinders (20 cm height × 10 cm diameter). The cylinders were separated by a nontransparent panel to prevent mice from seeing each other. The cylinders were filled with water (23°C), up to a height of 7.5 cm. Mice were placed into the cylinders, and their behavior was recorded over a 10 min test period. Data acquisition and analysis were performed automatically, using Image FZ software (see Image analysis). Images were captured at one frame per second. For each pair of successive frames, the amount of area (pixels) within which the mouse moved was measured. When the amount of area was below a certain threshold (i.e., 40 pixels), mouse behavior was judged as “immobile.” When the amount of area equaled or exceeded the threshold, the mouse was considered to be “moving.” The optimal threshold (amount of pixels) by which to judge immobility was determined by adjusting it to the amount of immobility measured by human observation. A capturing rate of one frame per second yielded results that were in good agreement with those obtained by human observation. Immobility lasting for less than a defined time threshold (i.e., 2 sec) was not included in the analysis. Distance traveled was measured by Image OF software (see Image analysis) using stored image files.
Image analysis. All applications used for the behavioral studies (Image EP, Image RM, Image FZ, Image SI, Image WM, and Image FZ) were run on a Macintosh computer. Applications were based on the public domain NIH Image program (developed by Wayne Rasband at the U.S. National Institute of Mental Health) and were modified for each test by Tsuyoshi Miyakawa (available through O'Hara & Co., Tokyo, Japan).
Statistical analysis. Statistical analysis was conducted using StatView (SAS Institute, Cary, NC) or SAS (SAS Institute). Data were analyzed by two-tailed t test, χ2 test, two-way ANOVA, or two-way repeated measures ANOVA, unless noted otherwise. Values in Tables and graphs are expressed as mean ± SEM.
RESULTS
Hyperactivity under normal, social, or stressful conditions inM1R−/− mice
All experiments were performed with male mice (littermates) that were 11 weeks old at the beginning of the behavioral studies. As indicated in Table 1,M1R−/− mice weighed ∼10% less than their wild-type littermates (p < 0.01). The lack of M1Rs had no significant effect on physical characteristics (whiskers and fur), sensory-motor reflexes (eye blink, ear twitch, whisker twitch, righting reflex, and acoustic startle response), sensory-motor gating (prepulse inhibition), pain sensitivity (hot plate test), or motor coordination (wire hang and accelerating rotarod tests) (Table 1) (p > 0.05).
General physical characteristics and sensory/motor functions of M1R−/− mice and their wild-type littermates
Strikingly, M1R−/− mice showed a pronounced increase in locomotor activity in several different tests. This hyperactivity phenotype was consistently found in all locomotor activity-related indices measured.M1R−/− mice traveled significantly longer distances in the light/dark transition test (Fig.1A) (genotype effect,F(1,44) = 24.1; p <0.0001), open field test (Fig.2A) (genotype effect,F(1,44) = 25.7; p <0.0001), elevated plus maze test (Fig.3A) (t = 2.80; df = 43; p = 0.008), and social interaction test (Fig.4B) (t= 3.17; df = 40; p = 0.003). The number of transitions in the light/dark transition test (Fig. 1B) (t = 2.15; df = 44; p = 0.037), the number of vertical activities in the open field test (Fig.2B) (genotype effect,F(1,44) = 7.17; p = 0.010), and the number of total arm entries in the elevated plus maze test (Fig. 3B) (t = 3.53; df = 43;p = 0.001) were also significantly increased inM1R−/− mice.

Increased locomotor activity ofM1R−/− mice in the light/dark transition test. Data are given as means (±SEM) for horizontal activity (centimeters) (A), number of transitions between the light and dark sides (B), and time spent in light (C). Horizontal activity was significantly increased in M1R−/− mice (genotype effect, p < 0.0001) (M1R−/− mice,n = 25; wild-type mice, n = 21). *Significantly different from wild-type mice (p < 0.05).

Increased locomotor activity ofM1R−/− mice in an open field test. Data are given as means (±SEM) for horizontal activity (centimeters) (A), vertical activity (B), time spent in the center area (C), and the number of fecal boli (D). Horizontal activity and vertical activity were significantly increased inM1R−/− mice (genotype effect, p < 0.0001 for horizontal activity andp = 0.010 for vertical activity) (M1R−/− mice,n = 25; wild-type mice, n = 21). Time spent in the center of the open field was significantly longer in M1R−/− mice (genotype effect, p = 0.012).

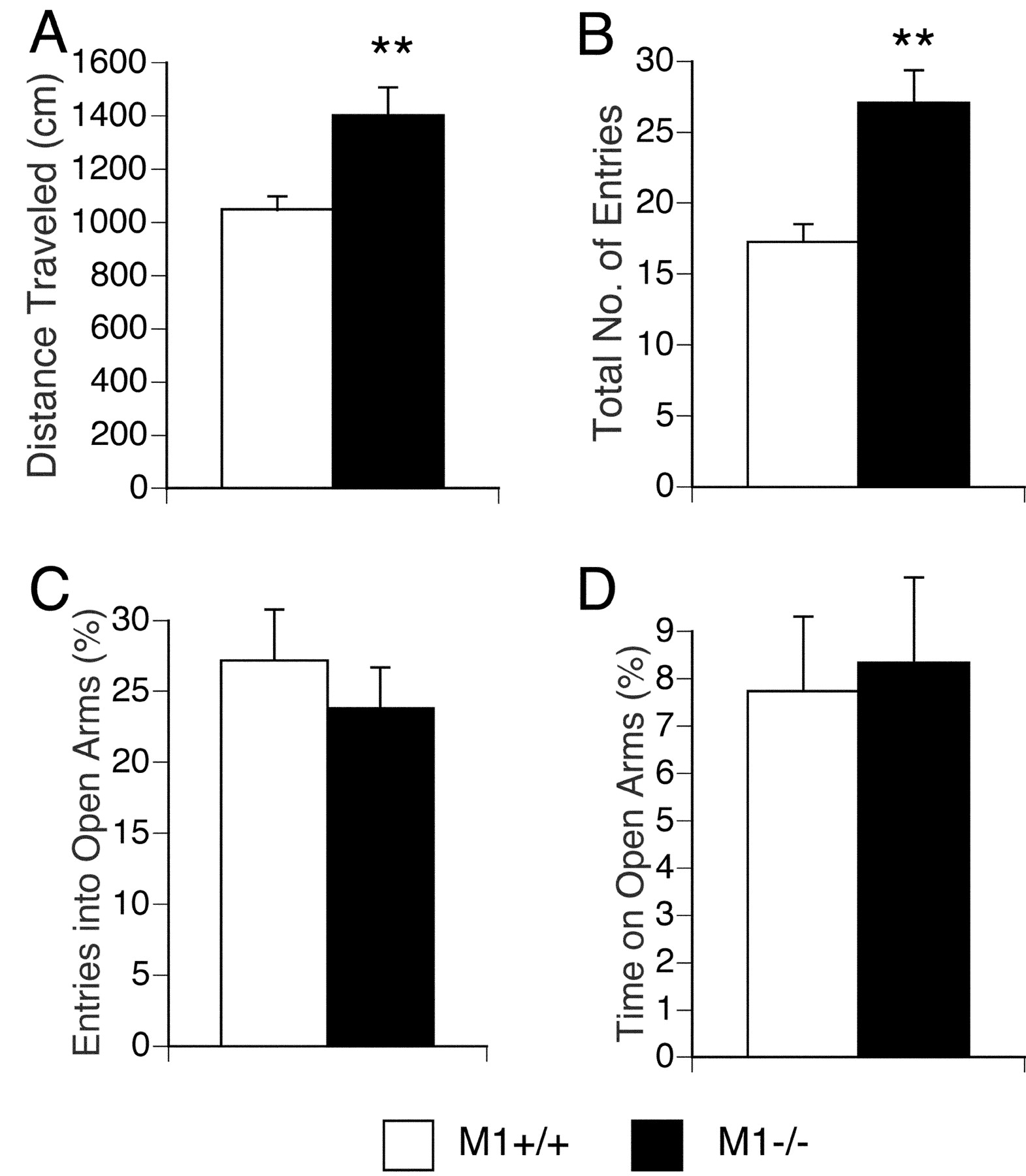
Increased locomotor activity ofM1R−/− mice in the elevated plus maze test. Data are given as means (±SEM) for distance traveled (A), total number of arm entries (B), percentage entries into open arms (C), and percentage time spent on open arms (D). Distance traveled and total number of entries were significantly increased inM1R−/− mice (**p < 0.01) (M1R−/− mice,n = 25; wild-type mice, n = 20).
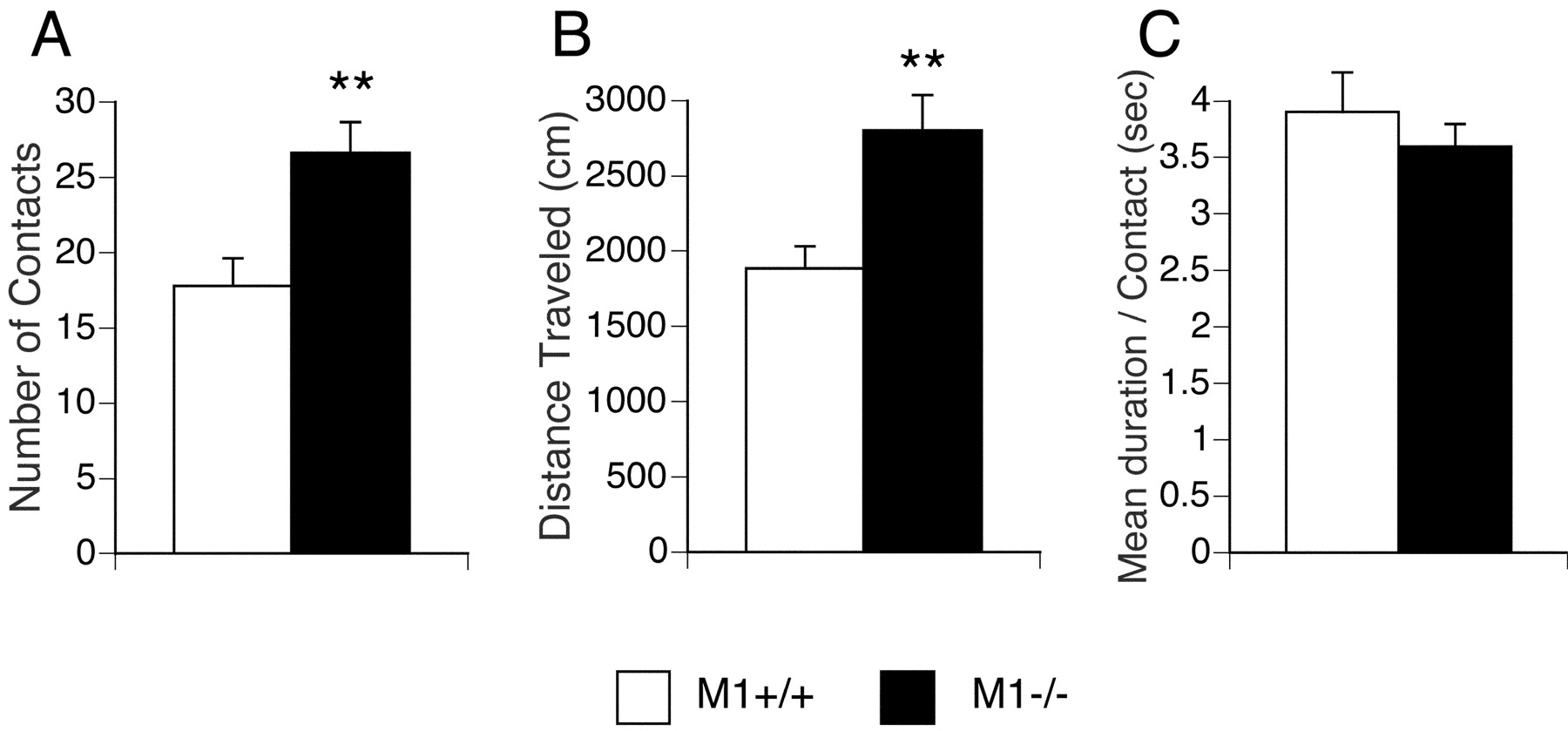
Increased number of social contacts ofM1R−/− mice in a social interaction test. Data are given as means (±SEM) for number of contacts (A), distance traveled (B), and mean duration per contact (C). The number of contacts and distance traveled were significantly increased inM1R−/− mice (**p < 0.01) (M1R−/− mice,n = 23; wild-type mice, n = 19).
As shown in Figure 2C,M1R−/− mice spent significantly more time in the center of the open field apparatus (genotype effect, F(1,44) = 6.80;p = 0.012), which is usually considered to reflect reduced anxiety (Crawley, 2000). However, the total time spent in the lit compartment in the light/dark transition test (Fig. 1C) (t = 1.30; df = 44; p = 0.20), the percentage of entries into the open arms (Fig. 3C) (t = 0.24; df = 43; p = 0.810), and the percentage of time spent on the open arms in the elevated plus maze test (Fig. 3D) (t = 0.77; df = 43;p = 0.449) were not significantly affected by the lack of M1Rs. These data indicate that the lack of M1Rs was not associated with consistent changes in anxiety-related behaviors.
In a social interaction test,M1R−/− mice showed a significant increase in the number of social contacts as compared with their wild-type littermates (Fig. 4A) (t = 3.14; df = 40; p = 0.003). The mean duration of contacts did not differ significantly among the two genotypes (Fig. 4C) (t = 0.81; df = 40;p = 0.423). Although an increase in the number of social interactions is usually considered a measure of reduced anxiety (File, 1980), it is likely that the increased number of social contacts displayed by the M1R−/− mice is primarily caused by their hyperactivity phenotype (see previous paragraph).
The behavior of M1R−/− mice and their wild-type littermates was also analyzed in the Porsolt swim test, which is widely used as a screening test for antidepressants (Porsolt et al., 1977; Crawley, 2000). In this test,M1R−/− mice spent significantly less time in immobility (usually indicative of decreased “behavioral despair”) thanM1R+/+ mice (Fig.5B) (genotype effect,F(1,39) = 10.04, p = 0.003; genotype × time interaction,F(9,351) = 1.58, p = 0.121). Moreover,M1R−/− mice traveled significantly longer distances than their wild-type littermates in this test (Fig. 5A) (genotype effect,F(1,39) = 8.56, p = 0.006; genotype × time interaction,F(9,351) = 1.47,p = 0.158), indicating thatM1R−/− mice displayed a hyperactivity phenotype also under extremely stressful conditions.

Decreased behavioral despair ofM1R−/− mice in the Porsolt forced swim test. M1R−/− mice traveled significantly longer distances (A) (p = 0.006) and spent significantly less time in immobility (B) (p = 0.003) than their wild-type littermates. Data are given as means (±SEM) (M1R−/− mice,n = 22; wild-type mice, n = 19).
Performance deficits ofM1R−/− mice in the eight-arm radial maze test
To examine whether the loss of M1Rs was associated with cognitive deficits,M1R−/− mice and their wild-type littermates were analyzed in several different memory and learning tasks. Initially, animals were tested in an eight-arm radial maze (Fig. 6), a test of spatial working memory. The angles between two successive arm choices were distributed equally from 45 to 180°, indicating that mice from either group were not using an “adjacent arm” search strategy (data not shown). The number of revisiting errors, in which subjects returned to the arms that had been visited previously to retrieve a food pellet, were significantly higher inM1R−/− mice during trials without delay (1st to 14th trials) (Fig. 6A) (genotype effect, F(1,42) = 6.94,p = 0.012; genotype × trial interaction,F(13,546) = 0.64, p = 0.823). There was no significant genotype effect or genotype × trial interaction in the number of omission errors (Fig.6E) (genotype effect,F(1,42) = 0.103, p = 0.750; genotype × trial interaction,F(13,546) = 0.984, p = 0.466), suggesting that reduced motivation to take the food pellets is unlikely to be responsible for the increase in revisiting errors displayed by the M1R−/− mice in the trials without delay.

Performance deficit ofM1R−/− mice in the eight-arm radial maze test. Mice were confined for 5 sec in the center platform after each arm choice. Delay (30 sec for the 15th trial and 120 sec for the 16th to 18th trial) was initiated after intake of the fourth pellet by confining the mice in the center platform. Data are given as means (±SEM) (M1R−/− ,n = 25; wild-type, n = 21).A, Number of revisiting errors. A small but significant genotype effect (p = 0.012) was observed during training without delay (trials 1–14). However, no significant genotype effect was found in training with delay (p = 0.185). B, Number of different arms chosen within the first eight choices. There were no significant differences between genotypes in training without or with delay (p = 0.901 and p = 0.376, respectively). C, Distance traveled during trials. M1R−/− mice traveled significantly longer distances during trials (p = 0.001 for training without delay andp = 0.042 for training with delay).D, Latency to take all eight pellets. There were no significant differences between genotypes in training without or with delay (p = 0.367 and p = 0.060, respectively). E, Number of omission errors (p > 0.05).
On the other hand, the number of revisiting errors during trials with delay (30 sec delay in the 15th trial, and 2 min delay in the 16th to 18th trials) was not significantly different between genotypes (Fig.6A) (genotype effect,F(1,42) = 1.82, p = 0.185; genotype × trial interaction,F(13,546) = 0.62, p = 0.602). Moreover, the number of different arms chosen during the first eight choices, which is considered a measure of working memory that is relatively independent of locomotor activity levels and the total number of choices, was not significantly affected by the lack of M1Rs, either during trials without delay (Fig.6B) (genotype effect,F(1,42) = 0.13, p = 0.901; genotype × trial interaction,F(13,546) = 1.08, p = 0.376) or during trials with delay (Fig. 6B) (genotype effect, F(1,42) = 0.54,p = 0.466; genotype × trial interaction,F(13,546) = 1.42, p = 0.241).
In the eight-arm radial maze test,M1R−/− mice traveled significantly longer distances during trials without delay (Fig.6C) (genotype effect,F(1,42) = 12.58, p = 0.001; genotype × trial interaction,F(13,546) = 1.03, p = 0. 427) and during trials with delay (Fig. 6C) (genotype effect, F(1,42) = 4.38,p = 0.042; genotype × trial interaction,F(13,546) = 0.05, p = 0.985). However, no significant differences were observed between the two mouse lines in latency to take all pellets during trials without delay (Fig. 6D) (genotype effect,F(1,42) = 0.83, p = 0.367; genotype × trial interaction,F(13,546) = 1.57, p = 0. 088) or during trials with delay (Fig. 6D) (genotype effect, F(1,42) = 3.73,p = 0.060; genotype × trial interaction,F(13,546) = 0.96, p = 0.412). On the basis of the observed increase in locomotor activity displayed by the M1R−/− mice, we reanalyzed the number of revisiting errors by analysis of covariance (ANCOVA), using distance traveled in the open field test as covariance. This analysis revealed that there was no genotype effect concerning the number of revisiting errors (genotype effect,F(1,40) = 0.01; p = 0.926). Consistent with this observation, there was a highly significant correlation between the mean number of revisiting errors (trials 1–14) and several activity indices measured in the open field, light/dark transition, and elevated plus maze tests (Table2). Taken together, these findings suggest that the increased number in revisiting errors displayed by theM1R−/− mice during trials without delay may be caused, at least to a major extent, by the general hyperactivity associated with the loss of M1Rs.
Correlation between mean number of revisiting errors during training without delay and locomotor activity indices measured in the open field, light/dark transition, and elevated plus maze tests
Reduced freezing ofM1R−/− mice in a fear conditioning test
The cognitive functions ofM1R−/− mice and their wild-type littermates were also analyzed in a contextual and cued fear conditioning test. During the conditioning period,M1R−/− mice showed lower levels of freezing after footshocks (Fig.7A, Conditioning) (genotype effect, F(1,41) = 13.34,p = 0.0007; genotype × time interaction,F(15,615) = 3.49, p < 0.0001). Sensitivity to footshocks was evaluated by measuring distance traveled during and immediately after footshocks were applied (Fig.7B). The distance traveled during the 4 sec interval after the initiation of footshocks was slightly increased inM1R−/− mice (Fig.7B) (genotype effect,F(1,40) = 4.43, p = 0.041; genotype × shock number interaction,F(2,80) = 0.22, p = 0.801), excluding the possibility that reduced sensitivity to footshocks is responsible for the reduced freezing levels observed after shock with the M1R−/− mice. There was a highly significant correlation between the distance traveled during the period before shock and freezing levels during the period after shock (r = −0.503; p = 0.0005), suggesting that the hyperactivity phenotype caused by the lack of M1Rs may be the primary cause of the reduced freezing levels after shock displayed by theM1R−/− mice.

Reduced levels of freezing inM1R−/− mice during conditioning, cued testing with altered context, and context testing.A, M1R−/− showed less freezing during the conditioning phase (p = 0.0007) and cued testing with altered context (p < 0.0001). There was no significant difference in freezing between genotypes during context testing conducted 24 hr after conditioning (p = 0.161). A small but significant difference (p = 0.024) was observed during context testing conducted 4 weeks after conditioning. B, Distance traveled during and immediately after application of footshocks during conditioning. Footshocks (bars) were presented three times for 2 sec. The distance traveled during 4 sec after the initiation of the footshocks was slightly increased (p = 0.041) inM1R−/− mice. Data are given as means (±SEM) (M1R−/− mice, n = 23; wild-type mice, n= 19).
When the conditioned stimulus (tone) was presented in an altered context 48 hr after conditioning (cued testing),M1R−/− mice showed significantly reduced levels of freezing (Fig. 7A,Cued Testing with Altered Context) (genotype effect,F(1,40) = 25.73; p < 0.0001). We noted that there was a highly significant correlation (1) between freezing levels while the tone was presented in the altered context and the distance traveled before footshocks during the conditioning phase (r = −0.493; p = 0.0008) and (2) between freezing levels while the tone was presented in the altered context and freezing levels after footshocks applied during the conditioning phase (r = 0.535; p = 0.0002). Moreover, M1R−/− mice traveled significantly longer distances during the period before tone in cued testing (p = 0.0035), and ANCOVA revealed (using the distance traveled during the period before tone in cued testing as a covariance) no significant genotype effect in freezing levels during the period after tone in cued testing (F(1,248) = 0.003; p = 0.955). These observations suggest that the reduced freezing levels displayed by the M1R−/− mice during cued testing may be caused primarily by the general hyperactivity associated with the lack of M1Rs.
Strikingly, M1R−/− mice and their wild-type littermates showed similar levels of freezing during context testing conducted 24 hr after conditioning [Fig. 7A,Context Testing (24 hr)] (genotype effect,F(1,41) = 2.04; p = 0.161). In contrast, when context testing was conducted 4 weeks after conditioning, M1R−/− mice displayed a significant reduction in freezing levels [Fig.7A, Context Testing (4w)] (genotype effect,F(1,40) = 5.52; p = 0.024). However, freezing levels did not differ significantly between genotypes during the first 2 min of testing (t = 1.60;p = 0.118). Again, the overall freezing levels observed during context testing (4 weeks after conditioning) correlated well with freezing levels after footshocks applied during the conditioning phase (r = 0.407; p = 0.007).
Intact performance ofM1R−/− mice in the Morris water maze test
M1R−/− mice and their wild-type littermates were also tested in the Morris water maze (hidden platform version), which is frequently used to assess spatial reference memory in rodents. Generally,M1R−/− mice showed no significant cognitive deficits in this test. Escape latencies (time required to reach the platform) were not significantly different between genotypes either during the original training (genotype effect,F(1,39) = 0.32, p = 0.572; genotype × trial interaction,F(5,195) = 1.17, p = 0.337) or during the reversal training (genotype effect,F(1,39) = 0.72, p = 0.401; genotype × trial interaction,F(4,156) = 0.50, p = 0.735) (Fig. 8A). Similarly, there was no genotype effect or genotype × trial interaction either in swimming speed (Fig. 8B) (p > 0.05) or in time spent in the perimeter of the pool (Fig. 8C) (p > 0.05).

Intact performance ofM1R−/− mice in the Morris water maze task. Escape latency (A), swimming speed (B), and time spent in the perimeter of the pool (C) did not differ significantly (p > 0.05) between the two genotypes either during original or during reversal learning. In probe trials, bothM1R+/+ andM1R−/− mice selectively searched the location where the platform had been located. Both genotypes spent significantly more time in the training quadrant (black bars) compared with the other quadrants (opposite quadrant, white bars; right quadrant, dark gray bars; left quadrant, light gray bars) in the probe trials conducted after original training (D) (p < 0.0001) and after reversal training (F) (p < 0.0001). Also, both genotypes crossed the training site significantly more often than the equivalent sites in the other three quadrants in the probe trials conducted after original training (E) (p < 0.0001) and after reversal training (G) (p = 0.003 forM1R+/+ mice andp < 0.003 forM1R−/− mice). Averaged swimming traces of swim pattern for wild-type mice (I, original learning; K, reversal learning) andM1R−/− mice (J, original learning; L, reversal learning) show that both genotypes selectively searched the trained quadrant. The configuration of the four quadrants is shown inH (numbers 0, 1,2, and 3 indicate trained quadrant, opposite quadrant, right quadrant, and left quadrant, respectively). Data are given as means (±SEM) (M1R−/− mice,n = 22; wild-type mice, n = 19).
During probe trials (platform removed), bothM1R+/+ andM1R−/− mice selectively searched the correct area where the platform had been located. Both mouse strains spent significantly more time in the training quadrant compared with the other quadrants in the probe trials conducted after the original training (Fig. 8D) [M1R+/+ mice, F(3,72) = 20.14, p < 0.0001; M1R−/− mice,F(3,84) = 25.42, p < 0.0001; Newman–Keuls post hoc comparison (trained quadrant more than all the other quadrants); p <0.01 for both genotypes] and after the reversal training (Fig.8F) [M1R+/+mice, F(3,72) = 8.31,p < 0.0001;M1R−/− mice,F(3,84) = 15.68, p < 0.0001; Newman–Keuls post hoc comparison (trained quadrant more than all the other quadrants); p < 0.01 for both genotypes]. In addition, bothM1R−/− mice and their wild-type littermates crossed the training site significantly more often than equivalent sites in the other three quadrants in the probe trials conducted after the original training (Fig.8E) [M1R+/+mice, F(3,72) = 15.26,p < 0.0001;M1R−/− mice,F(3,84) = 11.94, p < 0.0001; Newman–Keuls post hoc comparison (trained quadrant more than all the other quadrants); p < 0.01 for both genotypes] and after the reversal training (Fig. 8G) [M1R+/+ mice,F(3,72) = 5.09, p = 0.003; M1R−/− mice,F(3,84) = 5.00, p = 0.002; Newman–Keuls post hoc comparison (trained quadrant more than all the other quadrants); p < 0.05 for both genotypes].
Muscarinic receptor expression in the striatum ofM1R−/− and wild-type mice
As outlined above,M1R−/− mice showed a pronounced hyperactivity phenotype that was observed consistently in a number of different behavioral tests. Because striatal muscarinic receptors are thought to play an important role in the regulation of locomotor activity (Hornykiewicz, 1981; Fahn et al., 1990; Di Chiara et al., 1994), we wanted to determine at which levels the M1R is expressed in the mouse striatum and whether inactivation of the M1R gene led to compensatory changes in the levels of the remaining four muscarinic receptor subtypes (M2–M5).
To address this issue, muscarinic receptors (M1–M5) present in mouse striatal membrane preparations were labeled with a saturating concentration (2 nm) of the nonselective muscarinic antagonist, [3H]QNB, solubilized with 1% digitonin, and then immunoprecipitated by using receptor subtype-selective antisera (Gomeza et al., 1999a; Yamada et al., 2001). The specificity of the antisera (which were raised in rabbits against nonconserved regions of the third cytoplasmic loops of the mouse M1–M5 receptor proteins) was verified by using Chinese hamster ovary cell lines transfected with the M1–M5 receptor subtypes (data not shown). Use of the M1R antiserum showed that M1Rs are abundantly expressed in the striatum of wild-type mice (Fig.9A). As expected, the M1R antiserum was unable to immunoprecipitate significant amounts of radioactivity ([3H]QNB-labeled receptors) from striata derived from M1R−/− mice, confirming the lack of functional M1Rs in these animals. These studies also showed that the lack of M1Rs did not lead to compensatory changes in the expression levels of the remaining four muscarinic receptor subtypes (M2–M5) (Fig.9A). Essentially similar results were obtained when we performed analogous studies with cortical and hippocampal preparations (Fisahn, Yamada, Duttaroy, Deng, McBain, and Wess, unpublished observations).

Muscarinic receptor expression in mouse striatum studied via radioligand binding and immunoprecipitation assays.A, Immunoprecipitation studies. Striatal membranes prepared from wild-type orM1R−/− mice were incubated with 2 nm of the nonselective muscarinic antagonist [3H]QNB. [3H]QNB-labeled muscarinic receptors were solubilized and immunoprecipitated with M1–M5 receptor-selective antisera as described in Materials and Methods. B, Radioligand binding studies. Membranes prepared from mouse striata of the indicated M1R genotypes were incubated with a saturating concentration (2 nm) of the nonselective muscarinic antagonist [3H]QNB. Radioligand binding studies were performed as described in Materials and Methods. Data are given as means ± SD (n = 4). *p < 0.05 (Student's t test).
Consistent with the immunoprecipitation studies, [3H]QNB binding studies showed that muscarinic receptors are abundantly expressed in the striatum of wild-type mice (Fig. 9B). Strikingly, the total number of muscarinic binding sites was found to be strongly reduced, by ∼50%, in the striatal preparations fromM1R−/− mice (Fig.9B), suggesting that approximately half of the muscarinic receptors in the mouse striatum consist of M1Rs.
DISCUSSION
Hyperactivity ofM1R−/− mice
The lack of M1Rs did not lead to any obvious abnormalities in sensory-motor gating, nociception, motor coordination, and anxiety-related behavior. However,M1R−/− mice displayed a pronounced increase in locomotor activity that was consistently observed in all tests that included locomotor activity measurements. Consistent with this observation, it is well known that centrally active muscarinic agonists or antagonists can cause pronounced changes in locomotor activity levels (Molinengo et al., 1989; Shannon and Peters, 1990; Ukai et al., 1994). A hyperactivity phenotype was not seen with mutant mouse strains lacking M2 (Gomeza et al., 1999a) or M3 muscarinic receptors (Yamada et al., 2001). M4 muscarinic receptor knock-out mice showed increased locomotor activity in an open field test (Gomeza et al., 1999b). However, this phenotype was relatively mild compared with the robust hyperactivity phenotype displayed by theM1R−/− mice.
Muscarinic neurotransmission in the striatum is considered to play a major role in the regulation of locomotor control (Hornykiewicz, 1981;Di Chiara et al., 1994). Centrally acting muscarinic antagonists and dopamine receptor agonists are clinically useful in the treatment of Parkinson's disease (Fahn et al., 1990; Standaert and Young, 1996), supporting the concept that muscarinic and dopamine receptors interact in the striatum to promote normal locomotion. Radioligand binding and immunoprecipitation studies confirmed that M1Rs are abundantly expressed in the mouse striatum. Moreover, in agreement with a previous study by Hamilton et al. (1997), inactivation of theM1R gene did not trigger secondary changes in the expression levels of other muscarinic receptor subtypes in the striatum or other forebrain regions, excluding the possibility that upregulation or downregulation of non-M1muscarinic receptors is responsible for the observed hyperactivity phenotype.
Immunohistochemical studies indicate that M1Rs are expressed by most striatal projection neurons. Although M1Rs are colocalized with M4 muscarinic receptors in projection neurons that give rise to the striatonigral pathway, M1Rs represent the predominant muscarinic receptor subtype expressed in projection neurons giving rise to the striatopallidal pathway (Weiner et al., 1990; Bernard et al., 1992; Hersch et al., 1994). Activation of the striatopallidal pathway is thought to inhibit locomotor activity, whereas its inhibition is predicted to facilitate locomotion (Di Chiara et al., 1994; Standaert and Young, 1996). Unilateral ablation of striatal cholinergic interneurons induces contralateral rotation behavior, consistent with the concept that striatal muscarinic neurotransmission exerts a suppressive function on locomotor activity (Kaneko et al., 2000). One possibility therefore is that the loss of stimulatory M1Rs on striatopallidal projection neurons (M1 receptors are coupled to stimulatory G-proteins of the Gq family) (Wess, 1996) suppresses the activity of this inhibitory striatal output pathway, thus causing hyperactivity inM1R−/− mice. However, other mechanisms involving cortical or hippocampal M1Rs may contribute to or even be the primary cause of the locomotor phenotype observed with theM1R−/− mice. Clearly, more detailed studies will be needed to identify the molecular mechanisms by which M1Rs regulate locomotor activity.
It has been hypothesized that muscarinic hypersensitivity is involved in the etiology of depression (Janowsky et al., 1994). The Flinders Sensitive Line (FSL) of rats is known to be hypersensitive to muscarinic agonists and considered an animal model of depressive disorder (Overstreet et al., 1996). The FSL rats have increased muscarinic receptor densities in the striatum and hippocampus (Overstreet et al., 1996) and show increased immobility (behavioral despair) in the forced swim test, increased locomotor depression induced by footshocks, and reduced locomotor activity in an open field test (Overstreet and Russell, 1982; Overstreet, 1986). Pharmacological studies also suggest that M1Rs located in the nucleus accumbens may mediate increased behavioral despair (Chau et al., 1999). We found that M1R−/− mice displayed phenotypical features that were essentially opposite to those observed with the FSL rats. Most notably,M1R−/− mice showed reduced behavioral despair in a forced swim test and reduced locomotor depression after inescapable footshocks. Although these behaviors were likely to be affected by the general hyperactivity displayed by theM1R−/− mice, these observations are consistent with a role of M1Rs in the manifestation of depression-like behavior.
Hyperactivity is a key symptom in many neurological disorders, including attention deficit-hyperactivity disorder (ADHD), Tourette's syndrome, and manic disorders (Paule et al., 2000). In the past, the lack of genetically defined animal models of hyperactivity has made it difficult to identify the biological basis underlying these disorders. Our results suggest thatM1R−/− mice may serve as a novel animal model of hyperactivity. It is possible that more detailed analysis of the M1R−/− mice may shed new light on the pathophysiology of ADHD and other hyperactivity disorders.
Analysis of M1R−/− mice in learning and memory tests
M1R−/− mice showed largely normal working memory performance in the eight-arm radial maze test. We noted, however, thatM1R−/− mice displayed an increased number of revisiting errors during trials without delay. It is likely that this deficit is attributable largely to the hyperactivity phenotype displayed by theM1R−/− mice, because there was an excellent correlation between the number of revisiting errors and different locomotor indices. The mild performance deficit in this test and the hyperactivity phenotype of theM1R−/− mice are reminiscent of human disorders such as ADHD in which hyperactivity is often accompanied by cognitive deficits (Paule et al., 2000).
Fear conditioning studies showed thatM1R−/− mice displayed normal freezing levels during context testing performed 24 hr after conditioning, a behavior that is sensitive to blockade by muscarinic antagonists (Anagnostaras et al., 1995; Rudy, 1996; Fornari et al., 2000). On the other hand,M1R−/− mice exhibited reduced freezing in auditory-cued testing and a small but significant reduction of freezing when context testing was performed 4 weeks after conditioning. However, there was a highly significant correlation between spontaneous activity-related parameters and freezing levels in the M1R−/− mice, and the freezing levels of M1R−/− mice were similar to those of their wild-type littermates during the first minute in context testing after 4 weeks. These observations indicate that the reduced levels of freezing displayed by theM1R−/− mice may be caused primarily by their hyperactivity phenotype.
Our observation that M1R−/− mice showed reduced freezing after shock is consistent with a study byAnagnostaras et al. (1999) showing that a scopolamine dose that affected contextual fear conditioning also reduced freezing levels after shock. These authors suggested that scopolamine-induced impairment of working memory caused hyperactivity by disrupting habituation. However, it is unlikely that the hyperactivity phenotype displayed by the M1R−/− mice is caused by deficits in habituation or working memory, because (1) the activity levels ofM1R−/− mice were already increased during the first minute of testing in the open field test, (2) M1R−/− mice showed normal habituation in the open field test, and (3)M1R−/− mice displayed no major working memory impairment in the eight-arm radial maze test.
M1R−/− mice showed normal behavior in the Morris water maze test, which is a frequently used test to assess spatial reference memory in rodents. They performed equally as well as their wild-type littermates during both the original and the reversal learning trials, without showing differences in swimming speed. Previous work has shown that muscarinic antagonists can impair performance in a nonspatial discrimination task (Hagan et al., 1986) and in a cued version of the Morris water maze (Paylor and Rudy, 1990), augment swimming in the perimeter of the pool (Whishaw and Tomie, 1987), and frequently cause aberrant behaviors such as “jumping off,” “deflecting,” and “swimming over” after reaching the platform (Saucier et al., 1996). These aberrant behaviors, by themselves, could severely interfere with the performance of the test. In support of this view, nonspatial pretraining before drug treatment (Saucier et al., 1996) or a procedure designed to facilitate use of a spatial search strategy (Day and Schallert, 1996) eliminates spatial learning impairment induced by muscarinic antagonists. However,M1R−/− mice were completely devoid of such behavioral abnormalities. In the absence of such confounding factors, our data convincingly demonstrate that M1Rs are not essential for spatial learning as assessed by the Morris water maze test.
Taken together, these data indicate that the lack of M1Rs does not lead to major cognitive deficits in three different hippocampus-dependent learning tasks. It is therefore possible that the memory deficits observed after administration of centrally active muscarinic antagonists are caused by blockade of muscarinic receptor subtypes other than or in addition to the M1R. Although our data and those of Hamilton et al. (1997) suggest that inactivation of the M1R gene does not lead to significant changes in the levels of the M2–M5 receptor subtypes, other compensatory mechanisms may exist to maintain proper cognitive functions in the M1R−/− mice. For example, simultaneous blockade of muscarinic cholinergic and serotonergic neurotransmission eliminates normal electrographic activity in neocortex and hippocampus and leads to highly maladaptive behaviors in learning tasks (Vanderwolf, 1987). These electrographic and behavioral effects are not observed after inhibition of either one of the two neurotransmitter systems alone (Vanderwolf, 1987). Thus, the possibility exists that M1Rs play a role in cognitive functions that may become observable only when other compensatory receptor systems are disrupted simultaneously.
Footnotes
This research was supported by a Cooperative Research and Development Agreement between the National Institute of Diabetes and Digestive and Kidney Diseases (J.W.) and the Eli Lilly Research Laboratories. We thank J. N. Crawley, A. Holmes, and D. L. McKinzie for thoughtful discussions and critical reading of this manuscript, J. Gan for expert technical assistance, and A. M. Spiegel and I. W. Levin for generous support of this work.
Correspondence should be addressed to Dr. Tsuyoshi Miyakawa, Center for Learning and Memory, Massachusetts Institute of Technology, Building E17-342, 77 Massachusetts Avenue, Cambridge, MA 02139-4307. E-mail:miyakawa{at}mit.edu.
M. Yamada's present address: Laboratory for Cell Culture Development, Brain Science Institute, RIKEN, Saitama 351-0198, Japan.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}