

Article Figures & Data
Figures
- Fig. 1.
In the presence of carbachol, anItail was observed under voltage clamp in the same cells that revealed a PP in current-clamp mode.A, Typical responses of a hippocampal CA1 pyramidal neuron under current-clamp conditions to hyperpolarizing and depolarizing current injection in control aCSF. Depolarizing current injection elicited robust Ca2+ spikes, and the membrane potential immediately returned to baseline levels after cessation of the current pulse. B, In the presence of 20 μm carbachol (CCH), Ca2+ spike firing evoked by identical stimuli resulted in a long-lasting PP. C, The PP was reversible after carbachol was washed from the slice. D, In the same cell, under voltage-clamp conditions in the absence of carbachol, an 800 msec depolarizing voltage pulse to 0 mV from a holding potential of −70 mV resulted in unclamped Ca2+ currents. At the offset of the pulse, Itail was not observed. E, In the presence of carbachol, a long-lasting inward Itail was induced at the offset of the depolarizing voltage pulse under voltage-clamp conditions. F, Itail was reversible after wash of carbachol. Recordings were made with a Cs+-based intracellular solution and 1.2 μm TTX in the external solution.
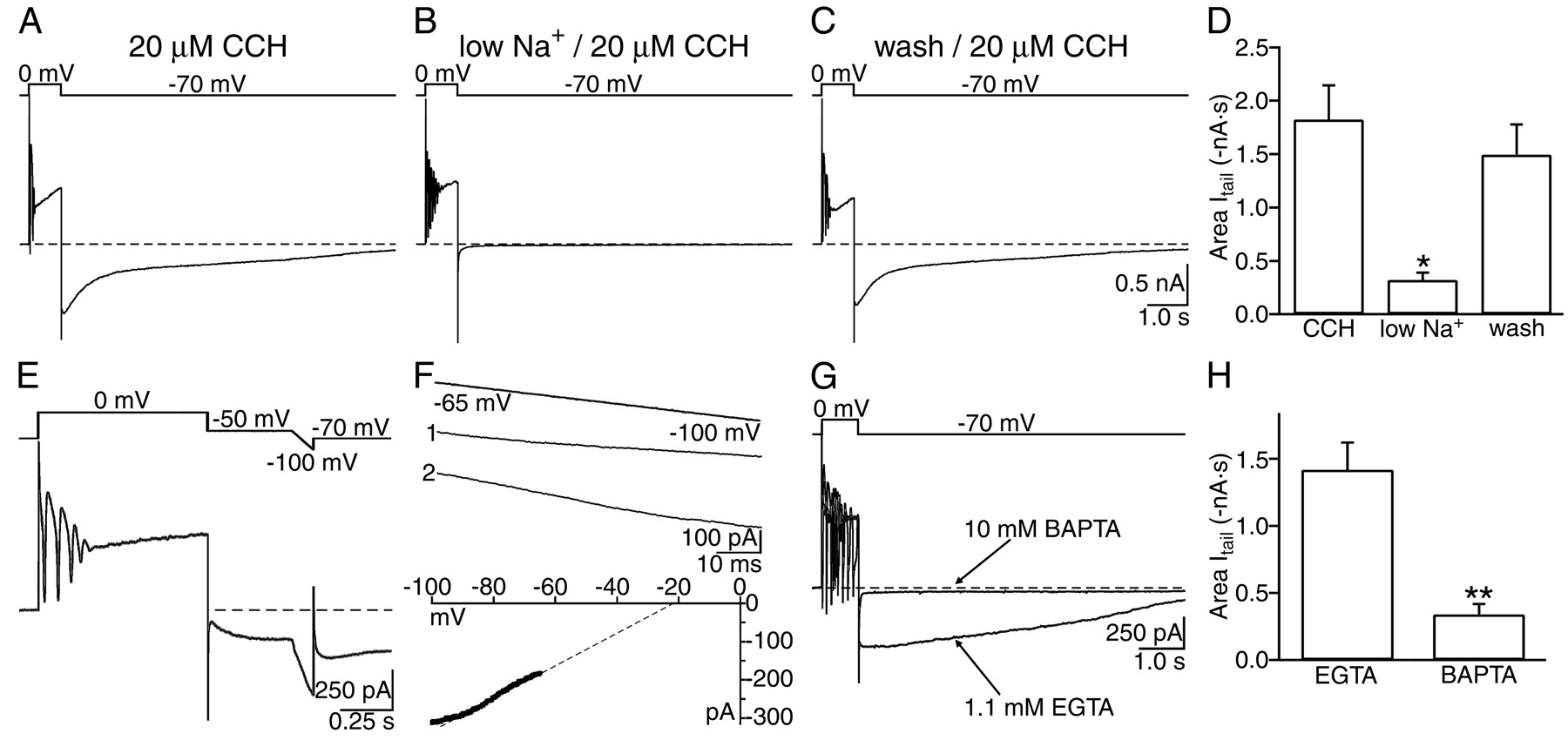
- Fig. 2.
Itail was dependent on Na+ influx independent of TTX-sensitive channels, varied linearly with voltage, and was blocked by intracellular BAPTA.A, In 20 μm carbachol,Itail was observed after a depolarizing voltage step (800 msec) to 0 mV from a holding potential of −70 mV.B, Reducing [Na+]o from 152 to 26 mm depressed Itail. External NaCl was substituted with equimolarN-methyl-d-glucamine. C, Restoring external NaCl to 152 mm completely reversed the depression of Itail induced by low [Na+]o. D, Summary of the effects of reducing [Na+]o and after washout on the area of Itail. Mean ± SE areas of Itail are plotted (n = 6). *p < 0.002 compared with control data. E, The reversal potential ofItail was assessed by ramping the voltage from −50 to −100 mV in 100 msec in the presence of carbachol. Ramps obtained during Itail were compared with ramps obtained from identical voltages without an evokedItail. F, Subtraction of the ramp currents without Itail(ramp 1) from the ramp currents duringItail (ramp 2) from −65 to −100 mV revealed that Itail varied linearly with membrane potential in the voltage range tested. The reversal potential in this cell determined by linear extrapolation (dotted line) was −20.1 mV. G, Superimposed recordings in carbachol from different CA1 pyramidal neurons from the same slice loaded with intracellular pipette solution containing either 1.1 mm EGTA or 10 mm BAPTA (indicated by arrows). Chelating [Ca2+]i with BAPTA depressedItail. H, Summary of the mean ± SE Itail areas from pyramidal neurons recorded with either (in the micropipette) 1.1 mmEGTA (n = 10) or 10 mm BAPTA (n = 9). **p < 0.001, EGTA compared with BAPTA.
- Fig. 3.
Itail was refractory when evoked at short intervals but was stable for up to 1 hr when evoked at >3 min intervals. A,Itail was evoked in the presence of 20 μm carbachol with a depolarizing voltage step to 0 mV from a holding potential of −70 mV (left). Another depolarizing voltage step to 0 mV 15 sec after the initialItail was evoked resulted in anItail with a reduced area (middle). A depolarizing voltage step to 0 mV 180 sec after an initial Itail was evoked resulted in a Itail with a similar area (right). B, Plot of the normalized mean ± SE Itail areas at intervals of 7.5, 15, 30, 45, 60, 90, and 180 sec with respect to the initial evokedItail, showing that activation ofItail had a refractory period (n = 9). A single exponential curve was fit (solid line), and the time constant of recovery was calculated to be 43.5 sec (indicated by dotted lines).C, The normalized mean ± SEItail areas evoked every 3 min were plotted against time for 51 min (n = 6).
- Fig. 4.
Itail required sGC activity but was independent of PKG activity. A, In the presence of 20 μm carbachol (CCH),Itail could be evoked with a depolarizing voltage step to 0 mV from a holding potential of −70 mV.B, After bath application of 20 μmLY83583, Itail was depressed.C, Summary of the mean ± SEItail areas in the presence of the sGC inhibitors LY83583 or ODQ or a PKG inhibitor compared withItail controls. Bath application of either 20 μm LY83583 (n = 5) or 20 μm ODQ (n = 5) significantly depressed Itail area. *p< 0.02, compared with control Itail areas. Intracellular perfusion of the PKG inhibitor KT5823 (10 μm) for >30 min did not alterItail area (n = 5, KT5823; n = 5, control).Itail areas from cells perfused with KT5823 were compared with control areas of Itailobtained from pyramidal neurons in the same slices. D,Itails of reduced areas were evoked with a depolarizing voltage step to 0 mV from a holding potential of −70 mV with perfusion of a submaximal dose of carbachol (5 μm).E, Bath application of zaprinast enhanced the evokedItail. F, The increase inItail area by zaprinast was reversible after wash.
- Fig. 5.
Antagonists of cyclic nucleotide-gated channels depressed Itail. A, In the presence of 20 μm carbachol (CCH),Itail was evoked with depolarizing voltage steps to 0 mV from a holding potential of −70 mV. B, Bath application of 100 μm 2′,4′-dichlorobenzamil (DCB) depressed generation ofItail. C, The inhibition ofItail was reversible after wash of 2′,4′-dichlorobenzamil. D, Summary plot of the mean ± SE areas of Itail before, after, and wash of 2′,4′-dichlorobenzamil. Application of 2′,4′-dichlorobenzamil reversibly depressed Itail(n = 7; *p < 0.001) compared with Itail control. E, Summary of the effects of bath application ofl-cis-diltiazem on mean ± SEItail area. Itailwas depressed by l-cis-diltiazem (n = 7; **p < 0.01 compared with control). In three neurons that were stable in the wash for >30 min, Itail partially recovered from the depression induced by l-cis-diltiazem.
- Fig. 6.
Itail was independent of nitric oxide production. A, The effects of the nitric oxide inhibitors l-NAME and l-NNA are summarized in the histogram. Itail area was not depressed by either bath application for >1 hr (1 mm) (n = 5) or intracellular perfusion (1 mm) (n = 10) of l-NAME. Inhibition of nitric oxide synthase by bath application (n = 5) or intracellular perfusion (n = 10) of l-NNA did not affectItail area.
- Fig. 7.
Proposed model for the activation ofItail and generation of plateau potentials. We propose that stimulation of muscarinic receptors (m1/m3) coupled to G-proteins in combination with Ca2+ influx through high-voltage-activated Ca2+ channels (HVA) can activate sGC, leading to an increase in intracellular cGMP and opening of CNG channels. The mechanism by which Ca2+ activates sCG does not apparently require nitric oxide. Influx of Na+ and Ca2+ through CNG channel openings mediates Itail and the prolonged depolarization during the plateau potential. Activation of protein phosphatase (PP) is required for plateau potential generation (Fraser et al., 2001), which may serve to increase the sensitivity of CNG channels to cyclic nucleotides. As part ofItail termination, cGMP-specific phosphodiesterase (PDE) can metabolize cGMP to 5′-GMP.












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}