

Abstract
Rabphilin, a putative rab effector, interacts specifically with the GTP-bound form of the synaptic vesicle-associated protein rab3a. In this study, we define in vivo functions for rabphilin through the characterization of mutants that disrupt theCaenorhabditis elegans rabphilin homolog. The mutants do not display the general synaptic defects associated with rab3 lesions, as assayed at the pharmacological, physiological, and ultrastructural level. However, rabphilin mutants exhibit severe lethargy in the absence of mechanical stimulation. Furthermore, rabphilin mutations display strong synergistic interactions with hypomorphic lesions in the syntaxin, synaptosomal-associated protein of 25 kDa, and synaptobrevin soluble N-ethylmaleimide sensitive factor attachment protein receptor (SNARE) genes; double mutants were nonresponsive to mechanical stimulation. These synergistic interactions were independent of rab3 function and were not observed in rab3–SNARE double mutants. Our data reveal rab3-independent functions for rabphilin in the potentiation of SNARE function.
- synaptic vesicle
- GTPase effector
- vesicle fusion
- neurotransmitter release
- SNARE
- synaptobrevin
- SNAP-25
- syntaxin
Rabphilin interacts directly and specifically with the GTP-associated form of the small GTPase, rab3, and as such has been suggested as a candidate effector for the function of rab3 in synaptic transmission (Shirataki et al., 1993; Ostermeier and Brunger, 1999). Rab3 is expressed in neurons and neuroendocrine cells and appears to play a regulatory role in synaptic transmission (for review, see Geppert and Südhof, 1998). The rab family consists of a large number of GTPases, each of which cycles between a membrane-associated GTP-bound form and a soluble GDP-bound form, and each of which has been implicated in a distinct type of vesicle-mediated transport (for review, see Novick and Zerial, 1997;Schimmoller et al., 1998). Association of guanine nucleotide hydrolysis with membrane fusion suggests a regulatory role for rab proteins in vesicle transport or fusion (Bourne et al., 1991; Fischer von Mollard et al., 1991; Nuoffer and Blach, 1994). Several rab proteins have been shown to function independently of GTP hydrolysis, suggesting that the GTPase activity may not directly facilitate fusion, but may instead regulate the amount of rab-GTP available to downstream effectors (Rybin et al., 1996; Richardson et al., 1998). Thus, the action of candidate effectors, including rabphilin, may hold clues to the role of rab3 in synaptic transmission.
Two functional domains have been identified in rabphilin. Association with rab3 requires the N-terminal cysteine-rich Zn2+-binding region (McKiernan et al., 1996; Stahl et al., 1996; Ostermeier and Brunger, 1999). It remains unclear whether rab3 recruits rabphilin to synaptic vesicles (Li et al., 1994; McKiernan et al., 1996; Stahl et al., 1996) or whether rabphilin also interacts with vesicles independently of rab3 (Shirataki et al., 1994; Arribas et al., 1997). The rabphilin C terminus contains two protein kinase C-like C2 domains that bind phospholipid in a Ca2+-dependent manner and that may play a role in membrane association (Yamaguchi et al., 1993; McKiernan et al., 1996; Chung et al., 1998; Ubach et al., 1999). The C2 domains resemble those of synaptotagmin, a protein with a critical role in synaptic transmission (for review, see Geppert and Südhof, 1998;Südhof and Rizo, 1996).
A number of studies suggest a role for rabphilin in synaptic function. Experiments that alter rabphilin levels in isolated synapses and cultured cells lead to changes in regulated exocytosis (Arribas et al., 1997; Chung et al., 1995, 1998; Komuro et al., 1996; Burns et al., 1998), and in some cases, endocytosis (Ohya et al., 1998). However, these experiments offer no consensus for a mechanism of action, because the nature and direction of the observed effects vary with experimental approach. Other studies have suggested that rabphilin plays a role in a rab3a-dependent mossy fiber pathway of long-term potentiation (Lonart and Südhof, 1998). Mouse knock-out studies failed to clarify a role for rabphilin, because a detailed physiological analysis revealed no abnormalities in either synaptic transmission or synaptic plasticity (Schlüter et al., 1999). Hence, the relationship between in vitro studies and the in vivo role of rabphilin remains unclear. We have investigated the role of rabphilin in the nematode using a molecular genetic approach to clarify its role in regulating synaptic function and its relationship to rab3.
MATERIALS AND METHODS
Growth and culture of Caenorhabditis elegans. Caenorhabditis elegans was grown at 20°C on solid medium as described by Sulston and Hodgkin (1988). 2-methyl-2-[methylthio]proprionaldehydeO-[methylcarbamoyl]oxime (aldicarb) was obtained from Chem Services, Inc. (West Chester, PA). Pharyngeal pumping and aldicarb sensitivity were tested as described previously (Nonet et al., 1997).
rab-3(js49), unc-10(md1117), andsnt-1(md290) represent null alleles based on molecular, genetic, and immunohistochemical criteria (Nonet et al., 1993, 1997;Schaefer et al., 2000). aex-3(y255) behaves as a strong loss-of-function allele (Iwasaki et al., 1997). However, the null phenotype has not been strictly defined. snb-1(md247),snb-1(js17), snb-1(js44), unc-64(js21) unc-64(e246), ric-4(js20), and ric-4(md1088)are hypomorphic alleles based on molecular, genetic, and immunohistochemical criteria (Nonet et al., 1998; Saifee et al., 1998) (Y. Lee, J. Rand, M. L. Nonet, B. J. Meyer, and J. Lee, unpublished observations). Double and triple mutants were constructed using standard methods. The genotypes of the strains were confirmed by molecular genotyping or noncomplementation assays. The genotype of strains contains the js232, md1117, and md290 deletions, and the md247 duplication was confirmed using PCR. The js49, e246,js21, and js20 lesions each result in a restriction-site polymorphism, and the presence of these alleles was confirmed using PCR. The presence of other lesions was confirmed using noncomplementation assays.
Electrophysiology. Electropharyngeograms (EPGs) were performed as described by Saifee et al. (1998). Bath solution consisted of Dent's saline with 2–5 mm serotonin to stimulate pumping. The presence of MC and M3 transients was scored qualitatively as distinct spikes greater than the background noise.
Electron microscopy. Worms were cut in 0.8% glutaraldehyde and 0.7% osmium tetroxide in 0.1 m cacodylate, pH 7.4, on ice. After 2 hr, they were moved to 2% osmium tetroxide in 0.1 m cacodylate, pH 7.4, and left at 4°C overnight. Processing and sectioning were as described previously (McIntire et al., 1992).
Isolation of rbf-1 cDNAs. Manipulations of DNA and RNA including electrophoresis, blotting, and probing of blots were performed using standard procedures (Sambrook et al., 1989) except where noted. cDNA was made by reverse transcribing RNA using random hexanucleotide primers as described previously (Sambrook et al., 1989).rbf-1 cDNAs were isolated by PCR and obtained from theC. elegans expression project (Kohara, 1996). Oligonucleotides used to amplify rbf-1 were chosen based onC. elegans spliced leader sequences and the genomic sequence of the rbf-1 locus (Wilson et al., 1994; Blumenthal, 1995) and included the following: SL-1, 5′-TGTAGAATTCCGCGGTTTAATTACCCAAGTTTG-3′; SL-2, 5′-TCTAGAATTCCGCGGTTTTAACCCAGTTACTC-3′; Rbf-A5, 5′-GGGATCCTAGAAAAAATGAGGAGGCG-3′; Rbf-C5, 5′-GCCTGCAGGCAATGGACTCAAATGG-3′; Rbf-A3, 5′-CGGGTACCTTTCTCCTCATCATCCAC-3′; Rbf-C3, 5′-CGGAATTCATGCTTGCCCAAATCGTG-3′; and Rbf-G3, 5′-TACCCATCTTGGTTGAATCCTGGGTC-3′.
Amplified cDNAs were sequenced directly or cloned into pBluescript and subsequently sequenced using Sequenase (United States Biochemicals, Cleveland, OH). Four amplified cDNAs (RBF-5-1f, RBF-5-1g, RBF-5-2h, and RBF-5-2i) that spanned most of the coding sequence were sequenced, as well as one from the C. elegans expression project (Kohara, 1996). A reference RBF-1 sequence was submitted to GenBank (accession number AF399852). Alternately spliced cDNA clones encoded products that differed from the reference as follows: exon 6 (amino acid 269–286) was found in cDNA yk39h5, but not in RBF-5-1f, RBF-5-1g, RBF-5-2h, or RBF-5-2i; exon 12 (amino acids 572–655) was present in RBF-5-1f and RBF-5-1g but absent from RBF-5-2h and RBF-5-2i; in RBF-5-1g, an alternate splice donor was used extending exon 8, replacing V435 with GNFSTI. The 5′ region of the gene was determined by amplifying and sequencing additional cDNAs that were spliced to the SL-1 spliced leader sequence and were identified on the basis of this 5′ sequence (Blumenthal, 1995). An additional partial 5′ cDNA was identified that contains an SL-2 leader sequence spliced to an upstream exon in addition to several of the first rbf-1 exons, but this transcript reaches a stop codon only six amino acids (aa) beyond the presumptive start ATG. Thebriggsae rbf-1 gene structure was deduced by comparison of the genomic sequences of C. elegans andCaenorhabditis briggsae. Intronic sequences were identified as regions with divergent sequences flanked by consensus splice acceptor and donor sequences. Coding regions were identified by homology and strong bias for third-codon silent substitutions.
Isolation of rbf-1 deletion strain. An expanded library of Tc1 insertions was screened as described previously (Zwaal et al., 1993). An insertion into the rbf-1 locus was identified using two rounds of PCR. Tc1-specific oligonucleotides were used as described previously (Zwaal et al., 1993). Therbf-1-specific oligonucleotides used were as follows: Rbf-H5, 5′-GAGGTACCTGATGAATGATTGGGAAATCG-3′; Rbf-H3, 5′-TAGCGGCCGCGAGAATCGTTCCATATTAG-3′; Rbf-K5, 5′-AATGTTTAGGCTGGCAGATGCTG-3′; and Rbf-K3, 5′-GAAGGATTCACCATCCACGAAC-3′.
A deletion event was detected with two rounds of PCR using the following oligonucleotides: Rbf-D5, 5′-TACTGCAGCATACCCTCATACAGTAATC-3′; Rbf-D3, 5′-GTGGATCCACATCAGTTGTCTCGACTC-3′; Rbf-E5, 5′-AACTGCAGTTTTCTTTGCTCCTCCCAC- 3′; and Rbf-E3, 5′-GCGGATCCACACACATTGCTGCATAGC-3′.
The js232 deletion deletes 1500 bp, resulting in the deletion junction cagaatttctgcaaaaat/ttccccccaagag. The resulting lesion was backcrossed 10 times to the wild-type strain N2 to eliminate background effects.
Behavioral assays. Defecation, pharyngeal pumping, and mating were assayed as described previously (Nonet et al., 1997). To quantify locomotor defects, 25–30 L4 larval stage animals were placed on a Petri dish recently seeded withEscherichia coli OP50. After overnight growth, the plate was immobilized on a Leica (Deerfield, IL) dissecting scope with tape and imaged using a Cohu (San Diego, CA) CCD camera controlled by Scion (Frederick, MD) Image software. An image series usually consisted of 300 frames captured at 5 sec intervals. To provide a consistent mechanical stimulus, a steel bar was dropped five times in rapid succession from a height of 5.1 cm at frame number 121. Mean velocities were calculated by summing the distances traveled by all worms in a pair of images using a custom macro. Mean basal velocities in Figure 6 were calculated by combining the data for the following time points: 750, 800, and 850 sec after stimulation (a group of 50–75 worms). Time points after stimulation were used instead of time points before stimulation because the stimulation history is better controlled using this protocol. Mean stimulated velocities were calculated by averaging the following time points: 20, 25, and 30 sec after stimulation. This time period was chosen for two reasons, First, tracking individual animals is difficult immediately after stimulation for wild-type and other strains with negligible locomotor defects. Second, certain soluble N-ethylmaleimide sensitive factor attachment protein receptor (SNARE) mutants have a delayed response to the stimulus and do not initiate movements immediately after stimulation. Color images in Figures 5 and 7 were created from grayscale images by inverting the images, placing one image in each of the green and red channels, and increasing the contrast.
Production of antisera. His6-tagged RBF fusion proteins were generated from pRSET vectors (Invitrogen, San Diego, CA), expressed in E. coli BL21, and purified on NTI-Sepharose (Qiagen, Hilden, Germany). The N-terminal fusion protein contains 172 aa of RBF-1 and includes most of the zinc-finger domain. The N-terminal construct, pRFN, included a 700 bp cDNA insert amplified with Rbf-H5 and Rbf-G3 oligonucleotides (sequences described above). Rbf-C5 and Rbf-C3 oligonucleotides were used to amplify the C-terminal region, and this PCR product was cloned into pRSETC to make the C-terminal construct pRfC2. Fusion proteins were purified and used to immunize rabbits as described previously (Nonet et al., 1993). For purifying antisera, Rbf-N–glutathioneS-transferase (GST) and Rbf-C2–GST were expressed using the pGEX vectors (Amersham Pharmacia Biotech, Arlington Heights, IL). GST fusion proteins bound to glutathione agarose beads were electrophoresed, transferred to polyvinylidene difluoride membranes, and used as a matrix for affinity purification of antisera (Smith and Fisher, 1984). Anti-RAB-3 and anti-synaptotagmin (SNT-1) antisera were used as described previously (Nonet et al., 1993, 1997). Antibody staining was visualized with anti-rabbit and anti-mouse antibodies coupled to cy5 and alexa-568 (Molecular Probes, Sunnyvale, CA) and with a Bio-Rad (Richmond, CA) confocal microscope.
RBF-1–green fluorescent protein fusion proteins. Vectors for producing green fluorescent protein (GFP) fusions were generously supplied by Andy Fire (Carnegie Institution of Washington, Baltimore, MD). For RBF-1–GFP, pRf9 was generated by cloning two genomic fragments (PstI–PstI andPstI–EaeI) from cosmid F34A7 (kindly provided by Alan Coulson, Sanger Centre, Hinxton, UK) into pPD95.75. The resulting fusion contains the entire RBF-1 open reading frame minus the last 25 codons fused to GFP and 7 kb of genomic sequence upstream of the ATG start codon.
RESULTS
rbf-1 encodes a C. elegans rabphilin homolog
A single rabphilin homolog was identified in C. elegansby a BLAST search of the C. elegans genome data (TheC. elegans Sequencing Consortium, 1998). No other rabphilin homologs were detected in the genome, with the most similar sequence being the synaptotagmin gene snt-1 (Nonet et al., 1993). We have designated this gene rbf-1, in keeping with C. elegans nomenclature conventions (Horvitz et al., 1979). The structure of the rbf-1 gene was deduced from sequence analysis of the partial expressed sequence tag cDNAs yk39h5 and yk129d3 (Kohara, 1996) and cDNAs representing the 5′ end of the mRNA isolated by PCRs (Fig. 1). The conservation between C. elegans and vertebrate rabphilin is strongest in the N-terminal cysteine-rich rab3 binding domain (45% identity; Fig. 1A) (Ostermeier and Brunger, 1999) and in the two C-terminal C2 domains implicated in Ca2+–phospholipid binding (48% identity; Fig. 1B) (Yamaguchi et al., 1993). However, in the “linker” domain between these two conserved regions, C. elegans rabphilin diverges from other rabphilins. In C. elegans this domain is 275 aa larger than of rat rabphilin and its sequence offers no clues to its function. Analysis of cDNAs also revealed several regions of alternate splicing. mRNA species of 2.8 and 3.1 kb were detected by Northern blot analysis (Fig. 1C), consistent with the structure of cDNAs we isolated. The largest alternately spliced exon codes for amino acids 572–655, a region within the large unique linker sequence (Fig. 1); see Materials and Methods for a complete description of alternative splicing. The sequence of the rbf-1 homolog from the closely related species C. briggsae was also analyzed. Rabphilin coding was highly conserved between these two species (Fig.1B,D; 95% identity in the zinc finger, 78% in the linker, and 94% in C2 domains). The noncoding sequence was completely divergent except for several small presumptive regulatory elements (Fig. 1D). Furthermore, all of the alternatively spliced exons were also present in C. briggsae, although 5 of the 16 introns found in the C. elegans gene were absent (Fig. 1D). The sequence similarity among theCaenorhabditis species indicates that the function of rabphilin domains, including the linker region, has been selected for during the evolution of the nematode lineage.

C. elegans rabphilin gene structure. A, A schematic diagram of the C. elegans and rat rabphilin proteins illustrating the conservation in domain structure. The percentage of amino acid identities among the two proteins in the zinc-finger (Zn2+) and calcium–phospholipid binding (C2) domains is indicated. B, Protein sequence alignments of the C. elegans, Drosophila, bovine, and rat rabphilin zinc-finger and C2 domains. Amino acids conserved among three of the four proteins in the alignment are boxed. The standard single-letter code was used. Introduced gaps are represented as dashes. Alignments were made using ClustalW (Thompson et al., 1994). The Drosophilasequence presented is a conceptual translation with no experimental confirmation. C, Northern blot of wild-type mixed-staged poly(A)-selected RNA probed with a rabphilin genomic clone. RNA size markers are labeled on the right (in kilobases).Arrows depict the two major rabphilin transcripts,C. briggsae and C. elegans rabphilin genes. Exons are depicted as patterned boxes. Exons encoding the zinc-finger domain (▨), the C2 domains (▩), and the alternatively spliced domains (▤) are uniquely labeled. Putative regulatory elements are depicted assmall black filled boxes. The position of thejs232 lesion and Tc1 transposon insertion (
 ) from which thejs232 lesion was derived are depicted below the diagram. The genomic regions included in two plasmid clones used in the study are also diagrammed.
) from which thejs232 lesion was derived are depicted below the diagram. The genomic regions included in two plasmid clones used in the study are also diagrammed.
RBF-1 colocalizes with synaptic vesicle markers independently of RAB-3
C. elegans rbf-1, like its mammalian homologs, is expressed in neurons. Antibodies against both the N-terminal and C-terminal domain of RBF-1 fusion proteins label the nerve ring, ventral nerve cord, and dorsal nerve cords in a punctate pattern (Figs.2 and 3). This staining pattern is characteristic of and colocalized with the staining pattern of antibodies directed against other C. elegans synaptic vesicle proteins, including RAB-3, SNT-1, and SNB-1 [synaptobrevin/vesicle associated membrane protein (VAMP)] (Fig. 2 and data not shown) (Nonet et al., 1993, 1997,1998). A rabphilin–GFP fusion expressed in transgenic animals also localized to the synaptic-rich neuropil (data not shown). Localization of synaptic vesicle-associated proteins in C. elegans is dependent on the UNC-104 kinesin (Hall and Hedgecock, 1991). In unc-104 hypomorphs, synaptic vesicles and vesicle-associated proteins accumulate in cell bodies (Hall and Hedgecock, 1991; Nonet et al., 1993). RBF-1 protein detected with antibodies and rabphilin–GFP was also mislocalized and accumulated in cell bodies rather than in processes in unc-104 worms (Fig.2 and data not shown). Our data demonstrate that RBF-1 is localized in the synaptic-rich neuropil of C. elegans in concert with other synaptic vesicle proteins and suggest that C. elegansrabphilin is associated with synaptic vesicles, as has been shown for vertebrate rabphilin (Li et al., 1994; Mizoguchi et al., 1994;Shirataki et al., 1994).

Rabphilin is localized to synaptic-rich regions of the C. elegans nervous system. Lateral views of the head region of whole adult animals fixed, permeabilized, coincubated with mouse and rabbit antibodies, and visualized with fluorescent tagged secondary antibodies are shown. RAB-3 (A) and rabphilin (B) colocalize in wild-type animals double-labeled with α-RAB-3 and α-RBF-1 antibodies. Synaptotagmin (C) and rabphilin (D) colocalize in wild-type animals double-labeled with α-SNT-1 and α-RBF-1 antibodies. RAB-3 (E) and rabphilin (F) both accumulate in neuronal cell bodies inunc-104(e1265) animals double-labeled with α-RAB-3 and α-RBF-1 antibodies. Synaptotagmin (G) and rabphilin (H) both remain localized to synapse-rich regions in rab-3(js49) animals double-labeled with α-SNT-1 and α-RBF-1 antibodies. Scale bar, 20 μm.

Rabphilin protein is absent inrbf-1(js232) mutant animals. Lateral views of the head region of whole adult animals fixed, permeabilized, incubated with mouse and rabbit antibodies, and visualized with fluorescent tagged secondary antibodies are shown. RAB-3 (A) and rabphilin (B, C) are localized in wild-type animals labeled with α-RAB-3 antibodies (A) as well as antibodies directed against the RBF-1 N terminus (B) and RBF-1 C terminus (C). However, rabphilin but notRAB-3 is absent in rabphilin mutants labeled with α-RAB-3 (D), α-RBF-1 N terminus (E), and α-RBF-1 C terminus (F) antibodies. Scale bar, 20 μm.
Previous studies in vertebrates have found that vertebrate rabphilin is recruited to vesicles through its association with rab3 (Stahl et al., 1996), whereas other studies have found localization of rabphilin to be independent of an interaction with rab3 (Shirataki et al., 1994). We examined whether RBF-1 localization is mediated through rab3. InC. elegans, RBF-1 localization was unaffected by the loss of rab3. RBF-1 localized to synaptic processes in a pattern that resembles that of the vesicle protein SNT-1 in both wild-type andrab-3(js49) null worms (Fig. 2G,H). Furthermore, rabphilin–GFP remained sequestered in the soma inunc-104 mutants even in absence of rab3, suggesting that rabphilin associates with vesicle membranes even in the absence of RAB-3 (data not shown). Thus, localization of rabphilin to synaptic vesicles and regulation of the transport of rabphilin were both independent of rab3.
rbf-1 mutants display no behavioral defects associated withrab-3 mutants
A deletion mutation in rbf-1 was isolated using Tc1-transposon-mediated mutagenesis (Rushforth et al., 1993; Zwaal et al., 1993) (see Materials and Methods). The js232deletion mutation removes 1500 bp including proximal promoter regions and the first three exons of the rbf-1 locus (Fig.1C). The deleted region includes the start codon and the first two CXXC motifs that have been implicated previously in the binding of Zn2+ and rab3 (McKiernan et al., 1996; Stahl et al., 1996). Although a transcript derived from the mutant locus can be detected using reverse transcriptase-PCR, the proximal ATG of this transcript is out of frame with the remaining RBF-1 coding sequences. Furthermore, antisera directed against both the N-terminal domain and the C-terminal domain of RBF-1 show no detectable staining in js232 animals (Fig. 3E,F). Thus, js232 completely disrupts the rab-3 binding domain of rabphilin and very likely represents a complete loss-of-function allele of rbf-1.
Homozygous rbf-1 mutant animals are viable, develop at comparable rates, and have brood sizes that are similar to those seen for the wild type (data not shown). Because rabphilin is thought to act as an effector of rab3, an rbf-1 mutation might be expected to disrupt the same functions that are disrupted in rab-3mutants. C. elegans rab-3 mutants were originally identified based on their resistance to the acetylcholinesterase inhibitor aldicarb (Nonet et al., 1997). Defects in synaptic function lead to aldicarb resistance, presumably because they lower the accumulation of acetylcholine in the synaptic cleft (Rand and Nonet, 1997). In contrast to rab-3 and many other synaptic transmission mutants, rbf-1 worms exhibit a sensitivity to aldicarb exposure that is similar to that seen for wild-type animals (Fig.4A). Therab-3 phenotype is also characterized by aberrant synaptic physiology that can be assessed using the EPG, a simple extracellular recording technique developed by Raizen and Avery (1994)and Avery et al. (1995). Mutations in several genes implicated in synaptic transmission significantly alter the EPG trace (Iwasaki et al., 1997; Nonet et al., 1997, 1998; Saifee et al., 1998). In contrast to rab-3, which exhibits EPG defects (Nonet et al., 1997), the EPGs of most rbf-1 worms were not distinguishable from those of wild-type animals (Fig. 4B). Recordings from a minority of rbf-1 animals did reveal slight abnormalities that were distinguishable from the wild type (Fig. 4C). However, the defects were much less pronounced than those ofrab-3 mutants (Fig. 4D). In these animals, pump length was prolonged and addition M3 transients were observed. These could reflect a slight decrease in the efficacy of M3 transmission, for which the inhibitory input onto muscle hastens repolarization. rab-3 mutations have also been shown to alter the distribution of synaptic vesicles surrounding neuromuscular junction presynaptic densities (Nonet et al., 1997). However, inrbf-1 mutants the general morphology of neuromuscular synapses appears normal (Fig. 4E–G). In particular, the distribution of synaptic vesicles in sections containing a presynaptic density was similar to the wild type (wild type, 33.0 ± 2.5, n = 34; rbf-1, 33.5 ± 2.4,n = 30). In summary, we found no defects in synaptic function or structure in the rabphilin mutant that were comparable with those of rab-3 mutants.
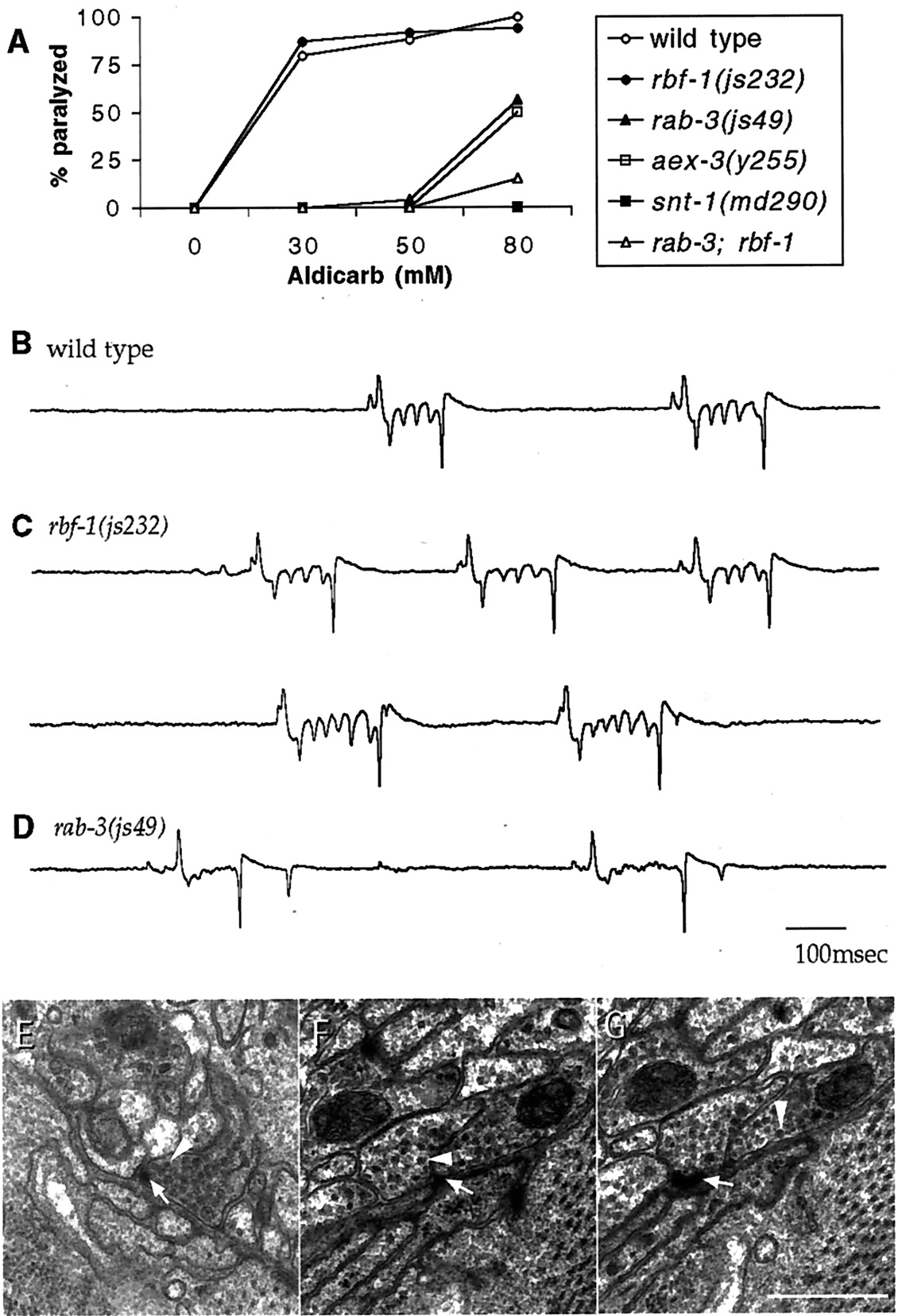
Rabphilin mutants appear normal in many assays of synaptic function. A, Rabphilin animals show wild-type responses to the acetylcholinesterase inhibitor aldicarb. Animals were placed on plates containing various concentrations of aldicarb, and their behavior was assayed 4.5 hr later. The percentage of animals paralyzed is plotted as a function of aldicarb concentration for the indicated genotypes in this representative experiment.B–D, Pharyngeal recordings from wild-type animals and mutants. Characteristic electropharyngeogram traces from the wild-type strain N2 (B), the rabphilin mutant (C), and the rab-3(js49) mutant (D) are shown. Recordings from rabphilin mutants were more variable than those from wild-type animals. The lower trace in C illustrates a typical abnormal electropharyngeogram observed in the rabphilin mutant. Alltraces are plotted as millivolts versus time.E–G, Ultrastructure of neuromuscular junctions from a wild-type animal (E) and two neighboring sections of the rabphilin mutant (F, G). Electron micrographs of ventral cord of adult animals are shown. A thick electron-dense specialization is visible at the opposition nerve and muscle (arrow) surrounded by a cluster of vesicles. The arrowhead identifies a representative vesicle. Scale bar, 500 nm.
Rabphilin mutants are lethargic
We examined the behavior of rbf-1 mutants using several different assays. The rate of pharyngeal pumping on food (wild type, 240 ± 17 pumps/min, n = 10; rbf-1, 227 ± 23 pumps/min, n = 10), the defecation cycle [wild type, 44.0 ± 7.9 sec cycle time and 95% enteric muscle contraction (EMC), n = 122; rbf-1, 47.0 ± 4.1 sec cycle time and 96% EMC, n = 90], egg laying (data not shown), and response to gentle touch (data not shown) all were similar to that of wild-type animals. Nevertheless, with careful observation, rbf-1 mutants could be distinguished from the wild type. Specifically, rbf-1mutants appeared lethargic when observed in the absence of exogenous stimulation. Manipulation of the Petri dish housing the mutants was sufficient to alter the behavior of rbf-1 mutants. Hence, we designed a time-lapse assay to observe the locomotor behavior ofC. elegans. In this assay, 25–30 young adult animals are placed on an agar-containing Petri dish freshly seeded with E. coli. The animals are imaged every 5 sec for 25 min. Ten minutes after initiating the recording, the plate is tapped five times in rapid succession with a standardized stimulus. This evokes a strong and reproducible increase in locomotion in the wild type. To quantify locomotion, the change in position of all animals was averaged over various 5 sec intervals during the recording (Fig.5). Wild-type animals exhibited a mean locomotor rate of 38.7 μm/sec in absence of stimulation, a condition we define as the basal rate of locomotion. Immediately after stimulation, locomotor rates increased to >200 μm/sec and slowly receded to the basal level over the intervening 10 min.

Rabphilin mutants are lethargic.A, Superimposed images of animals of different genotypes foraging on an E. coli bacterial lawn before (left) and after (right) a stimulus; see Materials and Methods for details. Two images separated by 120 sec (left) or 5 sec (right) were pseudocolored in green and red and superimposed. Animals that have moved appear as two images, onegreen and one red, whereas animals that have remained stationary or moved a minimal distance appear as a singleyellow or partially yellow image. The genotypes are rbf-1(js232), unc-64(e246), and jsEx510[pRF6; pMR1]. B, Quantification of locomotor rates of animals of different genotypes foraging on an E. coli bacterial lawn before and after a stimulus; see Materials and Methods for details. The mean locomotor velocity of 20–25 animals is plotted at various time points before and after a stimulus applied at t = 0. Genotypes: as described above and rab-3(js49).
In this assay, rabphilin mutants exhibited a pronounced decrease in spontaneous locomotion, averaging only 6.7 μm/sec (Figs.5B and 6A). Their locomotion rates were comparable with that of the severe hypomorphic unc-64(e246) syntaxin mutant (Figs. 5 and 6) (Saifee et al., 1998). In contrast, after stimulation, locomotion rates were comparable with the wild type (Fig. 5) but returned to the low basal rate in the intervening 10 min period. A close examination of the animals indicated that stationary rbf-1 animals are actively feeding and performing typical foraging nose movements. Hours of extended video observation of rbf-1 animals revealed that individual animals occasionally initiate bouts of active movement and then settle back down into a stationary feeding mode. This phenotype is directly associated with the rabphilin lesion, because wild-type locomotion was restored in most animals when a wild-type rabphilin construct was introduced into the rbf-1 mutant (Fig.5A,B). The incomplete nature of the rescue is probably a consequence of the mosaic nature of transgene expression in this system, although we have not excluded the possibility that genetic modifiers remained in our strain after ten-fold backcrossing.

Rabphilin lesion effects on basal and stimulated locomotion. Mean basal velocities (open bars) and mean stimulated velocities (filled bars) of rabphilin and rab-3 single, double, and triple mutant combinations are shown. Error is expressed as SEM. A, Expression of the rabphilin lethargic phenotype requires rab-3 activity. The genotypes are rbf-1(js232) andrab-3(js49). B, Synaptic regulators show negligible interactions with rabphilin. The genotypes areunc-10(md1117), snt-1(md290),aex-3 (y255), rbf-1(js232), andrab-3(js49). rab-3(y251) was used for the double mutant with snt-1(md290).rab-3(y251) behaves like a null mutant (Nonet et al., 1997). C, Neuronal SNARE mutations show strong interactions with rabphilin. The genotypes arerab-3(js49), rbf-1(js232), and SNAREs as indicated on the figure.
Rabphilin lethargic phenotype in synaptic mutants
We examined the locomotor behavior of many synaptic mutants to determine whether lethargic behavior was present in other synaptic mutants, and constructed double mutants to determine whether rabphilin phenotypes were expressed in the presence of other synaptic lesions. Lethargic behavior in the absence of stimulation was not observed in synaptotagmin, Rim, rab-3, or aex-3 guanine nucleotide exchange factor (GEF) mutants or in SNARE hypomorphic mutants that remain capable of some locomotion (Fig. 6). Surprisingly, most double mutants between rabphilin and the same group of mutants also failed to display the lethargic phenotype (Fig. 6). The most notable exception was a double mutant between rabphilin and the mild ric-4(js20) lesion. Thus, expression of the rabphilin lethargy phenotype appears to be dependent on the presence of an otherwise functional release apparatus.
Rabphilin is unlikely to encode the missing calcium sensor for fusion
Rabphilin and synaptotagmin both contain two C2 domains that bind calcium in vitro (Südhof and Rizo, 1996; Chung et al., 1998). These molecules are present on synaptic vesicles, and they have been proposed to act as calcium sensors that control the calcium sensitivity of the fusion step the nerve terminal (Geppert and Südhof, 1998). Previously, synaptotagmin mutants in several systems have been demonstrated to dramatically reduce but not completely eliminate synaptic transmission (DiAntonio et al., 1993;Littleton et al., 1993; Nonet et al., 1993; Geppert et al., 1994b; Fernandez-Chacon et al., 2001). For example,snt-1 synaptotagmin deletion mutants in C. elegans remain viable and are capable of coordinated movements (Nonet et al., 1993). Although synaptotagmin mutants in the mouse,Drosophila, and C. elegans all support the role of synaptotagmin as a calcium sensor for fusion, they also suggest the presence of other calcium sensors. We created double mutants between synaptotagmin and rabphilin and assessed their behavior.snt-1–rbf-1 double mutants behaved very similarly tosnt-1 single mutants, suggesting that rbf-1 does not mediate a significant portion of the remaining regulated release in this mutant. Double mutants were viable and exhibited locomotion (Fig.6B), pharyngeal pumping (snt-1, 27 ± 19 pumps/min; snt-1–rbf-1, 41 ± 17 pumps/min;snt-1–rab-3, 32 ± 17 pumps/min; n = 10 for each genotype), and defecation behaviors (data not shown) that were similar to that seen for synaptotagmin single mutants. Thus, our genetic analysis is inconsistent with rabphilin functioning as a predominant additional calcium sensor regulating the calcium dependence of the fusion event at the synapse.
SNARE function modulated by rabphilin activity
To further address the role of rabphilin at the synapse, therbf-1 mutation was combined with other mutations that alter synaptic release to determine whether a role for rabphilin could be uncovered in other compromised backgrounds. We paid particular attention to mutations in genes with homologs that had been implicated in docking and fusion of synaptic vesicles, including the membrane-associated SNARE proteins (Sollner et al., 1993).snb-1, ric-4, and unc-64 encode synaptobrevin/VAMP, synaptosomal-associated protein of 25 kDa (SNAP-25), and syntaxin, respectively (Nonet et al., 1998; Saifee et al., 1998) (Lee, Rand, Nonet, Meyer, and Lee, unpublished observations). We built and characterized double mutant combinations with various C. elegans SNARE homologs, as well as rab-3 and its putative regulatorsaex-3 and Rim. The rbf-1(js232) mutation enhances the locomotor defects of mutations in three neuronal SNARE genes. Hypomorphic mutants of each of these SNARE genes exhibit uncoordinated locomotion. Nevertheless, most of the milder alleles respond well to exogenous mechanical stimuli and drastically increase locomotion velocity after stimulation (Fig. 6C). When therbf-1(js232) mutation is combined with snb- 1(md247), ric-4(md1088), or unc-64(js21), the resultant double mutants are much less active than the single mutants (Fig. 6C). Locomotion velocities were extremely low and unaffected by exogenous stimulation (Fig. 6C). For example, ric-4(md1088) SNAP-25 mutant animals respond well to stimuli, whereas ric-4(md1088) rbf-1 mutants do not (Fig.7). A similar synthetic effect was observed for some, but not all, other alleles of snb-1 andric-4 (Fig. 6C and data not shown). Introduction of an rbf-1(+) construct into rbf-1–snb-1(md247)double mutants restored both basal and stimulated locomotion rates to typical snb-1(md247) behavior, verifying that the synergistic effects were a direct consequence of the rbf-1lesion (snb-1(md247) rbf-1(js232) + rbf-1(+) array (basal locomotion, 32 ± 8 μm/sec; stimulated locomotion, 71 ± 10 μm/sec). In contrast, this synergistic locomotion defect is not observed when rbf-1 is combined with other mutants harboring lesions in several other synapse-associated molecules implicated in rab-3 function (unc-10 Rim, aex-3rab-3 GEF; Fig. 6B) or in a molecule implicated in regulation of the calcium sensitivity of the fusion process (snt-1; as discussed above). Thus, rabphilin function influences locomotor behavior in genetic backgrounds for which SNARE activity is compromised, suggesting that a normal role of rabphilin is to potentiate SNARE function.

Synergistic locomotor defects in rabphilin–SNARE double mutants. Superimposed images of animals of different genotypes foraging on an E. coli bacterial lawn after a stimulus are shown; see Materials and Methods for details. Two images separated by 5 sec were pseudocolored ingreen and red and superimposed. Animals that have moved appear as two images, one green and onered, whereas animals that have remained stationary or moved a minimal distance appear as a single yellow orpartially yellow image. The genotypes areric-4(md1088), rab-3(js49), and rbf-1(js232).
Rabphilin SNARE genetic interactions independent of rab3 activity
We analyzed the behaviors of rab3–SNARE double mutants and compared their behavioral deficits with those for rabphilin–SNARE double mutants. rab3–SNARE double mutants exhibited none of the synergistic locomotion defects observed with rabphilin mutants (Fig.6). For example, although a rabphilin–SNAP-25 double mutant does not respond to stimuli, the rab3–SNAP-25 double mutant behaves similarly to the SNAP-25 single mutant (Figs. 6C and 7). Hence, the synaptic defects associated with rab-3 mutations did not appear to be additive with SNARE mutant defects, whereas the phenotypically much milder rabphilin mutations produced strong synergistic defects. To assess whether the rabphilin-mediated synergy was dependent on rab3 function, we constructed rab3–rabphilin–SNAP-25 and rab3–rabphilin–synaptobrevin triple mutants. In both cases, these mutants were completely nonresponsive to mechanical stimuli (Fig.6C). This indicates that the synergistic effects of rabphilin are independent of rab3 activity. However, basal locomotion rates increased in both triple mutants (Fig. 6C) consistent with our previous finding that the rabphilin lethargy phenotype is influenced by the functionality of the synaptic apparatus. Thus, our analysis of rab3–rabphilin–SNARE double mutants and triple mutants revealed rab3-independent functions of rabphilin in the nematode.
DISCUSSION
We have characterized the unique rabphilin gene of the worm.C. elegans rabphilin displays strong similarity to vertebrate rabphilin in both the rab3-binding zinc-finger domain and two calcium–phospholipid binding C2 domains (Fig. 1). Our work provides several insights into rabphilin function. First, our analysis provides in vivo data supporting a role for rabphilin in regulating nerve function. Second, our work suggests that C. elegans rabphilin associates with synaptic vesicles independently of rab3. Third, the phenotypic characterization of rabphilin defects indicates that certain rabphilin functions are independent of rab3. Fourth, rabphilin mutants do not enhance the behavioral defects of synaptotagmin mutants. Thus, our genetic analysis is inconsistent with the calcium–phospholipid binding activities of the rabphilin C2 domains representing a major constituent of the calcium sensor that triggers vesicle fusion. Fifth, lesions in rabphilin exhibit strong synergy with lesions in the neuronal SNAREs, implicating rabphilin in modulating SNARE function.
Rabphilin is largely dispensable for neuronal function
The extensive conservation of the zinc-finger and C2 domains fromC. elegans to vertebrates strongly implies that rabphilin plays a critical role in regulating nervous system function. Nevertheless, our genetic analysis of rabphilin mutants found that these animals show none of the behavioral, pharmacological, and physiological defects typically associated with mutants with deficiencies in synaptic function. Defects in cholinergic transmission in many synaptic mutants of C. elegans are detectable as resistant to the acetylcholinesterase aldicarb (Miller et al., 1996). Glutamatergic transmission defects in the M3 neuron detected using EPG are also associated with most synaptic transmission mutants (Nonet, 1999). However, neither of these assays revealed defects in rabphilin mutants. More generally, mutants with lesions in synaptic components exhibit a broad range of behavioral deficits including abnormalities in defecation, pharyngeal pumping, and mating assays (Rand and Nonet, 1997). Rabphilin mutants exhibited normal defecation, pharyngeal pumping, and mating behavior. Previous studies of a mouse rabphilin knock-out mutant revealed no defects in the physiological, behavioral, or morphological attributes of these animals (Schlüter et al., 1999). Although redundancy could account for the lack of a phenotype in the mouse, the findings are largely consistent with ours. Both studies suggest that rabphilin is dispensable for the function of many neurons.
Our studies did reveal a unique locomotor behavior defect in rabphilin mutants. In the absence of exogenous mechanical stimuli, rabphilin mutants rarely move. Rather, the animals remain stationary, feeding vigorously while continuing head foraging movements. Indeed, C. elegans rabphilin mutants moved at rates that were comparable with those of severe syntaxin hypomorphs under these controlled conditions. What defects could account for this unusual behavioral change? Many behaviors including pharyngeal pumping, defecation, mating, and stimulated locomotion are normal, suggesting that the cellular functions of the neurons controlling these behaviors are not significantly perturbed. One possibility is that a small general reduction in neuronal function in all neurons causes this change. However, we favor the hypothesis that a class (or classes) of interneurons in the “higher order” centers of the simple C. elegans nervous system that regulate locomotion are critically dependent on rabphilin for proper function. First, similar behavioral defects have not been observed in the large number of hypomorphic mutant alleles of various strengths in many synaptic regulatory genes that have been characterized in C. elegans. Second, combining rabphilin with more severe synaptic defects blocks expression of the phenotype, suggesting that displaying the behavior associated with the rabphilin mutant requires significant neuronal functionality. Mosaic analysis using neuron-specific promoters should permit us to determine whether rabphilin functioning to regulate this behavior is limited or broadly distributed in the nervous system.
C. elegans RBF-1 localizes with synaptic vesicles independently of rab3
Models for rabphilin function have postulated that rab3 recruits rabphilin to synaptic vesicles (Li et al., 1994; Stahl et al., 1996;Ostermeier and Brunger, 1999). However, in C. elegans, we have shown that this is not the case: rab-3 null mutants properly localize rabphilin (Fig. 2). Consistent with our observations, previous biochemical studies also reported rab3-independent localization of rabphilin (Shirataki et al., 1994). Thus, rabphilin likely associates with vesicles via interaction with either phospholipids or another vesicle-associated protein. Stahl et al. (1996) demonstrated that incubation of vesicles in vitrowith guanine nucleotide dissociation inhibitor and GDP removes both rab3 and rabphilin from vesicles, suggesting that both dissociate from membranes during GTP hydrolysis by rab3. Although rabphilin localizes properly in the absence of rab3 in C. elegans, it is possible that in the presence of rab3 it could dissociate from the membrane during GTP hydrolysis. However, others have shown that the absence of rab3a in mice alone causes a reduction and mislocalization of rabphilin staining, a result that is more difficult to correlate with our own (Geppert et al., 1994a; Li et al., 1994). One major distinction between C. elegans and vertebrate neurons is the distances that components need to be transported to reach synaptic terminals. Perhaps a reduced affinity for vesicles in the absence of rab3 leads to greater degradation of rabphilin during the extended transport in vertebrate systems. Regardless of the localization mechanism in the mouse, our genetic data also indicate that rabphilin functions independently of rab3, and thus support the existence of a rab3-independent localization mechanism.
rabphilin functions independent of rab3
Although rabphilin mutants have subtle behavioral defects, when placed in combination with hypomorphic lesions in the neuronal SNARE genes, the resultant double mutants exhibited severe defects in stimulated locomotion. Several lines of evidence suggest that these genetic interactions are specific. First, rab3–SNARE double mutants do not exhibit the same synergistic locomotor defects. Because rab3 mutants exhibit much stronger general behavioral defects than rabphilin mutants, these synergistic behavioral phenotypes are not the simple consequence of placing any two synaptic mutants together. Second, rabphilin mutants do not enhance the stimulated locomotion defects of several other synaptic mutants; notably, no interactions are observed with rab3, rab3 GEF (aex-3), Rim (unc-10), orsnt-1 mutants. Qualitatively, synaptotagmin mutants and Rim mutants show equal or more severe behavioral defects than those of the most severe hypomorphic SNARE lesions we examined. Third, the rabphilin SNARE paralysis was not dependent on rab3 activity. rab3–rabphilin–SNARE triple mutants were also nonresponsive to stimuli. Because the paralysis phenotype observed in the rabphilin–SNARE double mutant is specific to the neuronal SNAREs, independent of rab3 activity, and not observed with rab3 mutants, we conclude that the rabphilin SNARE interactions are not consistent with a rab3 effector function.
rab3-independent effects of rabphilin on regulated release have also been observed in cell culture (Arribas et al., 1997; Joberty et al., 1999). Rabphilin constructs containing either a deletion of the zinc-finger domain or point mutants in the zinc-finger domain that render the molecule unable to interact with rab3 still stimulated calcium-regulated exocytosis in two distinct systems. Our data are consistent with these findings and suggest that this rab3-independent rabphilin activity may involve regulation of SNARE function.
A critical role for rabphilin to potentiate SNARE functions: an accessory docking protein
Several rabs, notably rab5 and ypt7, have been demonstrated to play roles in docking through interactions with effectors (Christoforidis et al., 1999; Price et al., 2000). Furthermore, these effectors interact directly with SNAREs linking the vesicle to the target membrane (McBride et al., 1999; Simonsen et al., 1999). Indeed, rab proteins in yeast are essential components of the secretory pathways they regulate (Novick and Zerial, 1997). In contrast, previous studies of rab-3 mutants in C. elegans and rab-3a mutants in mice came to the unexpected conclusion that rab-3 was not required for synaptic vesicle release (Geppert et al., 1994a;Nonet et al., 1997). A major difference between the synapse and most other secretory pathways is that vesicles are found in abundance at most synapses. The strict requirements for reliable, efficient, and timely release at the synapse may necessitate that docking not be the rate-limiting step in the synaptic vesicle cycle. To accomplish this, one possibility is that some docking functions have been parceled out to other molecules.
Our analysis of rabphilin is consistent with rabphilin potentiating SNARE function. Furthermore, SNAP-25 interacts directly with rabphilin in a manner that can be detected both by immunoprecipitation and SNAP-25 affinity chromatography of brain extracts (X. Zhang, P. Hanson, J. Ubach, G. Lonart, R. Jahn, and T. C. Sudhof, personal communication). In light of these biochemical data, we propose that rabphilin acts directly to potentiate SNARE functions, perhaps by regulating SNARE complex assembly. For example, rabphilin–SNAP-25 interactions could help orient SNAREs for efficient pairing. This interaction, although effectively redundant in a wild-type genetic background, becomes critical when SNARE function is compromised.
Footnotes
This work was supported by Grants to B.G. and M.L.N. from the United States Public Health Service. J.S. was supported by a National Research Service Award. We thank Michael Crowder, Maya Kunkel, Tom Sudhof, Aguan Wei, and the Nonet laboratory for critical comments; Liping Wei and Gayla Hadwiger for technical assistance; Tom Sudhof and Phyllis Hanson for sharing unpublished data; and Jim Rand, Junho Lee, and Erik Jorgensen and the Caenorhabditis Genetic Center for providing genetic strains.
Correspondence should be addressed to Michael L. Nonet, Department of Anatomy and Neurobiology, Washington University School of Medicine, 660 South Euclid Avenue, St. Louis, MO 63110. E-mail:nonetm{at}thalamus.wustl.edu.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}