

Abstract
Mouse Zic genes encode zinc finger proteins and are expressed in the developing and mature CNS. Reduced expression of Zic2 in mice results in spina bifida and holoprosencephaly. However, the disruption of Zic1, a strong homolog of Zic2 that has an overlapping expression pattern, results in cerebellar malformation with no apparent abnormalities in the forebrain or in posterior neuropore closure. Here we revealed that Zic2 and Zic1cooperatively control cerebellar development by regulating neuronal differentiation. Both Zic1 and Zic2 are expressed in the precursor cells of the granule neuron and the neurons in cerebellar nuclei. Mice carrying one mutatedZic1 allele together with one mutatedZic2 allele (Zic1+/−Zic2+/kd ) showed a marked cerebellar folial abnormality similar to, but distinct from that found in mice homozygous for the Zic1 mutation (Zic1−/− ). TheZic1+/−Zic2+/kd cerebellum is missing a lobule in the anterior vermis and has a truncation of the most posterior lobule. Expression of transverse zonal markers is shifted anteriorly in the developing cerebellum, indicating that the anterior part of the cerebellum is poorly developed. Abnormalities in the developingZic1+/−Zic2+/kd cerebellum share the following features with those of theZic1−/− cerebellum: a preceding reduction of cell proliferation in the anterior external germinal layer, a reduction in cyclin D1 expression, and enhanced expression of the mitosis inhibitors p27 andp16, and enhancement of Wnt7a expression. These results indicate that Zic1 and Zic2 may have very similar functions in the regulation of cerebellar development.
The mammalian cerebellum is composed of multiple folia and fissures that show a uniform and predictable pattern within a given species (Altman and Bayer, 1997; Voogd, 1998). This elaborate pattern of foliation involves the coordinated expression of genes within specific locations in the developing neural tube. Recent studies using genetically modified mice have revealed several genes controlling cerebellar development (Hatten et al., 1997; Herrup and Kuemerle, 1997; Wassef and Joyner, 1997; Goldowitz and Hamre, 1998).
Mouse Zic1 is one of the genes controlling cerebellar development (Aruga et al., 1994; 1998). Zic1 is expressed in the dorsal neural tube and its derivatives and is strongly expressed in the developing and mature cerebellum. In the Zic1-deficient (Zic1−/− ) mice, the cerebella are hypoplastic, and their folia are malformed. The abnormalities are closely related to the loss of granule cells, which is most dramatic in the anterior vermis.
Several other Zic-related genes have also been identified and implicated in neural development in mice and other species. Among the mouse Zic genes, Zic2 is most similar toZic1 (Aruga et al., 1996) and is expressed in overlapping regions (Nagai et al., 1997). However, the phenotypes of the mutant mice with reduced expression of Zic2(Zic2kd/kd ; kd,knock-down) are rather different from those ofZic1-defienct mice. TheZic2kd/kd mice show holoprosencephaly and spina bifida (Nagai et al., 2000), which derive from malformations in the anterior and posterior ends of the neural tube, respectively. The apparent differences in the mutant phenotypes raise the question of whether Zic1 and Zic2 are functionally related.
To address this question, we first examined the cerebellar abnormalities in the Zic2kd/kd mice. Because these mice die just after birth with severe brain deformities, we were not able to detect any abnormalities similar to those ofZic1-deficient mice. As many vertebrate genes can be functionally compensated by structurally related genes, we started generating mice with Zic1/Zic2 compound mutations. Mice carrying mutations in one allele of Zic1 and one allele of Zic2(Zic1+/−Zic2+/kd ) showed a behavioral abnormality with cerebellar malformation. This indicates that mouse Zic2 is involved in cerebellar patterning in cooperation with Zic1.
Furthermore, a number of genes controlling cell proliferation and differentiation are deregulated in the cerebella of bothZic1+/−Zic2+/kd and Zic1−/− mice, suggesting that both Zic1 and Zic2 function upstream of these genes to regulate cerebellar development.
MATERIALS AND METHODS
Mutant mice. Mice heterozygous for theZic1 “null” mutation (Aruga et al., 1998) were backcrossed onto C57BL/6 mice 7–10 times, and mice heterozygous for the Zic2 “knock-down” mutation (Nagai et al., 2000) were backcrossed four or five times. These backcrossed mice were then used in subsequent matings. Mice were genotyped and maintained as described (Aruga et al., 1998; Nagai et al., 2000). Noon of the day on which vaginal plugs were first observed in the morning was defined as embryonic day 0.5 (E0.5). The mice were maintained by the Advanced Technology Development Center, RIKEN Brain Science Institute and Division of Experimental Animal Research, Tsukuba Life Science Center, RIKEN. All animal experiments were done according to the guidelines for animal experimentation in RIKEN.
Histology and morphometric analysis. For histological examination, tissue was fixed in Bouin's fixative and processed for paraffin sections and stained with hematoxylin and eosin. The morphometric analysis was performed as described (Aruga et al., 1998). Serial sagittal sections (6 μm) were prepared, and the most comparable sets of sections was selected for the analysis. All the pictures were first digitized by Fuji HC-2500 3CCD camera. The areas were measured using density slice function of NIH Image program (developed at the National Institutes of Health, Bethesda, MD; available on the Internet at http://rsb.info.nih.gov/nih-image/). EGL length was calculated by the same software, measuring manually fitted curves on the EGL. The subjects for the morphometric analysis in this study were two sets of (Zic1+/+, Zic2+/+ ), (Zic1+/−, Zic2+/+ ), (Zic1+/+, Zic2+/kd ), and (Zic1+/−, Zic2+/kd ) at postnatal day 16 (P16), three pairs of Zic2+/+ andZic2+/kd , and two pairs ofZic2+/+ andZic2+/kd at 1 year old. Each set or pair was derived from the same litter.
In situ hybridization and immunohistochemistry. In situhybridization was performed as described (Nagai et al., 1997) withZic1, Zic2, and En2 probes (Aruga et al., 1994, 1998; Nagai et al., 1997). Cryosections were prepared at 10 μm. Adjacent sections were used in hybridizations to compare the distribution of two different transcripts.
For immunohistochemical staining, cryosections were incubated for 12 hr at 4°C with antibody binding solution consisting of primary antibody [anti-calbindin (Chemicon, Temecula, CA), anti-Eph3 (Santa Cruz Biotechnology, Santa Cruz, CA), anti-βIII tubulin (Promega, Madison, WI), anti-p16 (Santa Cruz Biotechnology), and anti-phospho-histone H3 antibody (Upstate Biotechnology, Lake Placid, NY)], 2% normal goat serum, and 0.1% Triton X-100 in PBS. The bound primary antibody was detected by Cy3-conjugated anti-rabbit IgG, TRITC-conjugated anti-mouse IgG (Jackson ImmunoResearch, West Grove, PA), or a vector stain ABC kit (Vector Laboratories, Burlingame, CA). Immunohistochemical staining with anti-phospho-histone H3 antibody was performed on four pairs of (Zic1+/+, Zic2+/+ ) and (Zic1+/−, Zic2+/kd ) embryos. Five comparable sections were prepared from each embryo, and the mean number of the labeled cells were calculated. Statistical significance was determined using at test. P values < 0.05 were considered significant.
RT-PCR analysis. E17.5 cerebella from each genotype were excised and frozen in liquid nitrogen. Cerebellar peduncles were trimmed, and pia maters were peeled off before freezing. Total RNA was isolated using Trizol reagent (Life Technologies, Gaithersburg, MD). After DNaseI treatment, reverse transcription was performed with Superscript II reverse transcriptase (Life Technologies). Glyceraldehyde-3-phosphate dehydrogenase (G3PDH) expression was measured to monitor RNA recovery. RT-PCR was performed after they were in the log-linear phase of the amplification curve at the indicated cycles. The PCR cycles, annealing temperature, and primer sequences are: G3PDH, 23 cycles, 68°C, 5′ CCGGTGCTGAGTATGTCGTGGAGTCTAC 3′ and 5′ CTTTCCAGAGGGGCCATCCACAGTCTTC 3′;cyclin D1, 30 cycles, 55°C, 5′ AACTACCTGGACCGCTTCCT 3′ and 5′ CCACTTGAGCTTGTTCACCA 3′; cyclin D2, 26 cycles, 55°C, 5′ AGACCCATCTTCAGCTCCTG 3′ and 5′ TGCTCAATGAAGTCGTGAGG 3′; p27, 30 cycles, 55°C, 5′ CTGTGTGCAGTCGCAGAACT 3′ and 5′ CCAGGGGCTTATGATTCTGA 3′; and Wnt7a, 32 cycles, 55°C, 5′ CACAGTTCCGAGAGCTAGGC 3′ and 5′ CCTGTCACTGGGTCCTCTTC 3′.
RESULTS
Expression of mouse Zic2 in the developing cerebellum
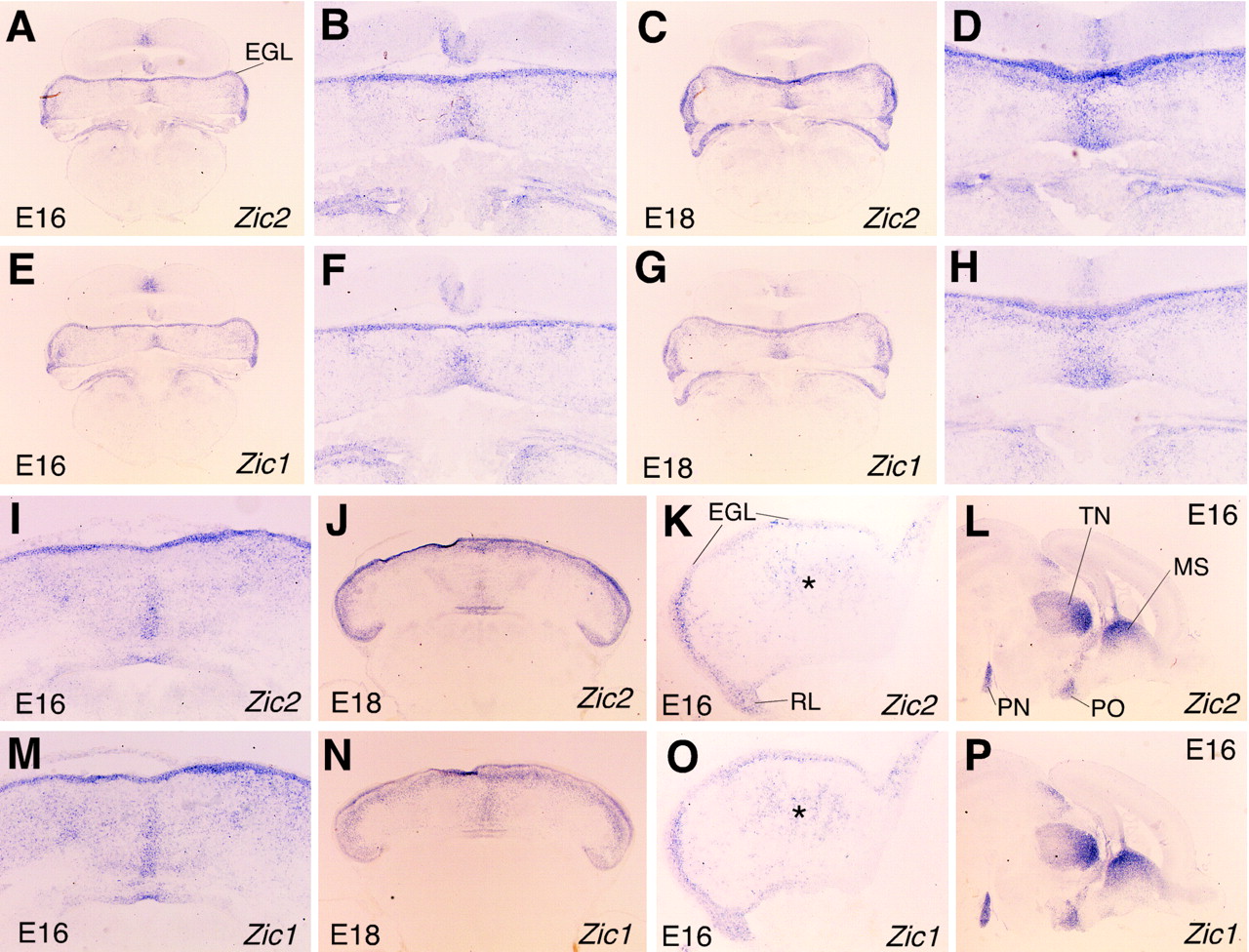
We have previously shown the expression pattern of Zic2at times earlier than E14 (Nagai et al., 1997). To clarify the role ofZic2 in cerebellar development, we first examined the expression of Zic2 at later stages (Fig.1), compared with that ofZic1. Sagittal sections of E16 and E18 brains showed very similar expression patterns for the two genes (Fig.1K,L,O,P), both being strongly expressed in the external germinal layer (EGL) and rhombic lip of the cerebellum and pons in the hindbrain structure. In the telencephalon and mesencephalon, they are expressed in the medial septal nucleus, the thalamic nuclei, and preoptic nucleus.

The expression of Zic1 andZic2 in the developing cerebellum. In situ hybridization was performed on coronal (A–H), horizontal (I, J, M, N), and sagittal (K, L, O, P) sections of E16 (A, B, E, F, I, K, L, M, O, P) and E18 (C, D, G, H, J, N) wild-type cerebella with Zic2(A–D, I–L) and Zic1 (E–H, M–P) probes. B, D,F, and H are higher magnifications ofA, C, E, andG, respectively. Zic1 andZic2 are expressed in a similar pattern. A significant difference between Zic1 and Zic2expression was found in the vermal (medial) intermediate region (I, J, M, N). Asterisks inK and O indicate midsagittal expression in I and M, respectively.EGL, External germinal cell layer; MS, medial septum; PN, pontine nuclei; PO, preoptic nuclei, TN, thalamic nuclei.
In coronal sections of the same stage, there was a region expressingZic1 and Zic2 in the midline of the cerebellum that was widest at the ventral part of the cerebellar anlagen (Fig.1A–H). In horizontal sections at E18,Zic2 expression is composed of three lines, whereasZic1 was expressed in a single region that was strongest in the midline (Fig. 1J,N). They are similarly expressed at E16 (Fig. 1I,M). The midline expression of Zic1 and Zic2 corresponds to a weak but significant expression in the center of the cerebellar primordia in the midsagittal sections (Fig. 1K,O, asterisk). Along the anterior–posterior (AP) axis, Zic1 is more specifically expressed in the midline of the anterior region. In the mature brain,Zic2 and Zic1 are expressed in almost identical patterns (Aruga et al., 1998; data not shown).
The cerebellar abnormalities in the Zic1+/− Zic2+/kd mice
We next examined cerebellar phenotypes of the Zic2knock-down mice (Zic2kd/kd ). In the wild-type E16 cerebellum, the left and right cerebellar plates are fusing at the midlines, and one can normally observe a shallow groove at the ventral side of the cerebellar primordia in coronal section (Fig. 2A). There was a narrow groove at the midline of theZic2kd/kd cerebellum (Fig. 2C), and the ventricular spaces and the subarachnoid space were markedly reduced. In contrast to these gross morphological abnormalities, there was no obvious histological abnormality (Fig. 2B,D).Zic2kd/kd mice had shrinkage or dilatation of the ventricles at the later embryonic stages, which may have been caused by disturbed CSF circulation because of major neural tube anomalies present (spina bifida, holoprosencephaly, and exencephaly) (data not shown).

Cerebellar phenotype in Zic2knock-down mice. Coronal (A, C) and sagittal (B, D) hematoxylin and eosin-stained sections were prepared from E16 wild-type (Zic2+/+) (A, B) and Zic2kd/kd (C, D) embryos. The distorted shape suggests oppression, which is probably caused by disturbed CSF circulation. It is difficult to judge whether Zic2 has a primary role in the cerebellar development, from Zic2kd/kd phenotypes. CN, Cerebellar nuclei forming cells;EGL, external germinal cell layer.
Because the Zic2kd/kd phenotype was less informative about the role of Zic2 in cerebellar development, we next asked whether Zic2 may act in cooperation with Zic1. To address this question, we started generating Zic1/Zic2 compound mutant mice. This was possible because Zic1 and Zic2 are located on different chromosomes (chromosome 9 and 14, respectively). Initially, we had planned matings betweenZic1+/−Zic2+/kd mice to obtain double homozygous mutant mice. However, theZic1+/−Zic2+/kd mice had a poor survival rate. The frequency of the genotypes obtained by the mating between Zic1+/− andZic2+/kd mice were:Zic1+/−Zic2+/kd , 8%;Zic1+/−Zic2+/+ , 29%;Zic1+/+Zic2+/kd , 29% andZic1+/+Zic2+/+ , 34% (n = 115) at 4 weeks after birth. In addition, theZic1+/−Zic2+/kd mice frequently showed hydrocephalus (four of nine at 4 weeks) and always showed a gait and postural abnormalities. We therefore gave up mating betweenZic1+/−Zic2+/kd mice and investigated the abnormalities appearing in theZic1+/−Zic2+/kd mice.
First, the mature brain was examined macroscopically. There was a marked change in the fissure pattern in the cerebellum. The mutant cerebellum apparently lacks the primary fissure in the vermis and posterior superior fissure in the hemisphere (Fig.3). Comparison of parasagittal sections through the median vermis and the hemisphere indicated that a lobule in the anterior lobe was missing (Fig. 4). Although folial identities are not conclusive at this point, we assigned presumable numbers to the lobules, as in Figures 2,B and C, and 3D. The elongated lobule VI covers the anterior lobe, whereas lobule IX is stubbed in comparison with the wild type. In Zic1+/− mice, lobule V was slightly smaller than the wild type, consistent with a previous study (Aruga et al., 1998), whereas theZic2+/kd did not show any significant alterations (Fig. 4C,E). Foliation patterning in the cerebellar hemisphere was also altered in theZic1+/−Zic2+/kd mice (Fig. 4D, right). In this case, the position of simple lobules shifted anteriorly, and the CrusI of the ansiform lobule extended anteriorly in a similar manner to the change in lobule VI of the vermis.

Macroscopic observation of theZic1+/−Zic2+/kd cerebella. Cerebella from wild-type (Zic1+/+Zic2+/+ ) (A) andZic1+/−Zic2+/kd (B, C) at P17 are seen from dorsoposterior (A, B) or posterior (C) direction. Folial patterns are markedly altered in theZic1+/−Zic2+/kd , different from that ofZic1−/− . Vermis lobules are indicated by V–IX. a, Anterior colliculus; p, posterior colliculus; CI, CrusI lobule; CII, CrusII lobule; S, simplex lobule; P, paramedian lobule. Scale bar, 5 mm.

Comparison of the cerebellar folial patterns in sections. Sagittal sections through cerebellar vermis (left) and hemisphere (right) fromZic1+/+Zic2+/+ (A),Zic1+/−Zic2+/+ (B),Zic1+/+Zic2+/kd (C),Zic1+/−Zic2+/kd (D), andZic1−/−Zic2+/+ (E), at P16 (A–D) or 3 weeks (E) mice. Vermis lobules are indicated byIII–X. A, Anterior lobe;CI, CrusI lobule; CII, CrusII lobule;S, simplex lobule; PM, paramedian lobule;P, pyramidis. Asterisks indicate dorsal (*) and ventral (**) paraflocculus lobules. Circlesindicate abnormally located lobules, which are not assigned at this point.
These features are essentially similar to the folial abnormality in theZic1−/− cerebellum (Aruga et al., 1998) in that a lobule in the anterior lobe is missing. However, there are some differences. First, the cerebellum in theZic1+/−Zic2+/kd mouse is not as small as that in theZic1−/− mouse, which shows a greater reduction in the medial to lateral aspect. Second, the extension of lobule VI was greater than that seen in theZic1−/− cerebellum. Third, the truncation of lobule IX is observed only inZic1+/−Zic2+/kd cerebella. This feature produces a steep posterior face of the cerebellar mass (Fig. 3B,C).
Morphometric analysis was used to quantitatively evaluate differences in the mutants. In young adults, there was a reduction in the area of the granule cell layer in Zic1+/− (Fig. 5), consistent with a previous study (Aruga et al., 1998). In theZic2+/kd , there was a slight reduction of the granule cell layer area of the hemisphere. A slight reduction in the hemisphere was also observed in 1-year-oldZic2+/kd cerebellum (data not shown). In contrast to the cerebella of single mutants, theZic1+/−Zic2+/kd cerebella showed a marked reduction in the area of both granule cell layer and molecular layer. The reduction was more than that of single mutants. The area (mass) reduction reflected a decrease in the number of granule cells because the cell density of the granule cell layer was not changed (Zic1+/+Zic2+/+ , 4.4 × 105 ± 2.5 × 104(cells/mm2);Zic1+/−Zic2+/+ , 4.3 × 105 ± 4.9 × 104;Zic1+/+Zic2+/kd , 4.3 × 105 ± 5.0 × 104;Zic1+/−Zic2+/kd , 4.2 × 105 ± 5.0 × 104).

Morphometric analyses of the mutant cerebella. The areas of the granular cell layer (GCL) and the molecular layer (ML) in midsagittal (vermis) and parasagittal sections through hemisphere (hemisphere) were measured. The cerebella were derived from P16 single litter. The areas are indicated in square millimeters. The error bars indicate SD.White bar,Zic1+/+Zic2+/+ (+/+); black bar,Zic1+/−Zic2+/kd (Z1/Z2); hatched bar,Zic1+/−Zic2+/+ (Z1/+); stippled bar,Zic1+/+Zic2+/kd (+/Z2); *p < 0.05; **p < 0.01.
In contrast to the folial pattern abnormalities, laminar organization of the cerebellar cortex is not disturbed, and the Purkinje and granule cell markers were expressed properly (data not shown), indicating that lineage-specific differentiation occurs normally inZic1+/−Zic2+/kd mice.
Abnormalities in the developing cerebella
To understand how these malformations occurred, we examined the developing cerebella. At E18, the folial pattern of theZic1+/−Zic2+/kd was distinct from the wild type (Fig.6C,D). The primary fissure was located anteriorly in the cerebellum. Correspondingly, the immature Purkinje cells, which were aligned in a defined pattern along the AP axis, were shifted anteriorly, resulting in the shorter anterior segment (Fig. 6E,F). These features are similar to those of Zic1−/− mice (Fig. 6A,B). The folial pattern abnormality could be traced back to E17.5 when the position of the initial primary fissure was located more anteriorly in theZic1+/−Zic2+/kd (data not shown). Ephrin receptors are differentially expressed along the AP axis in the cerebellar primordium (Rogers et al., 1999). The EphA3 receptor, which is usually expressed in the lobule VI region, was detected more anteriorly at E16.5 (Fig. 6G,H). We therefore concluded that the cerebellar patterning along AP axis had been differentially established at this point.

Anterior to posterior patterning abnormality found in theZic1+/−Zic2+/kd cerebella. Sagittal sections from E18 (A–F) and E16 (G, H) cerebella ofZic1+/+Zic2+/+ (A, C, E, G),Zic1−/−Zic2+/+ (B), andZic1+/−Zic2+/kd (D, F, H). A, C,E, and G are the littermates ofB, D, F, andH, respectively. A–D, Hematoxylin and eosin staining; E, F, immunohistochemical staining using anti-calbindin antibody (immature Purkinje cells).Arrowheads indicate the corresponding posterior and anterior boundaries of the clusters of Purkinje cells.G, H, Anti-EphA3 antibody staining, which stains a transversal zone corresponding to prospective lobule VII.Arrowheads indicate the strongly stained zone in each panel. I, J, Coronal sections of E18Zic1+/+Zic2+/+ (I) andZic1+/−Zic2+/kd (J) embryos. K, L, En2expression in the E17 horizontal section ofZic1+/+Zic2+/+ (K) andZic1+/−Zic2+/kd (L) are shown by in situhybridization. Note that the anterior region is reduced, as indicated by the folial and the marker expression patterns, whereas there are no apparent abnormalities in the medial to lateral patterning.
We also examined the patterning abnormality along the medial to lateral axis. At E18, a comparison of coronal sections revealed poorly formed foliations in the hemisphere of theZic1+/−Zic2+/kd cerebella without obvious alteration in medial to lateral extension of the cerebellar mass or the location of the cerebellar nuclei (Fig.6I,J). En2 is expressed in three parasagittal stripes in horizontal sections of the cerebellar primordium (Millen et al., 1995). This three-striped pattern was also evident in theZic1+/−Zic2+/kd cerebellum (Fig. 6K,L), suggesting that the medial to lateral pattern was not grossly affected in the cerebellar primordium. Taken together, these findings indicate a definite patterning abnormality along the AP axis in theZic1+/−Zic2+/kd cerebellum.
The reduced proliferation in the EGL underlies theZic1+/− Zic2+/kd cerebellar abnormality
Our previous study onZic1−/− mice showed that proliferating granule cell precursor levels were decreased in the EGL (Aruga et al., 1998). To examine whether a similar change occurs inZic1+/−Zic2+/kd , numbers of mitotic cells were compared. At E16, mitotic cells were decreased in the EGL, but not in the intermediate zone or the ventricular zone (Fig. 7A). At E17.5, the greatest difference was seen in the EGL anterior to the primary fissure (Fig. 7B). In contrast to the anterior EGL segment, the reductions in the posterior segment were slight. Mitotic cell number per a unit length (1 mm) was significantly decreased only in the E16 EGL (Fig. 7C), suggesting that the reduction of the total mitotic cell numbers at E17 reflects an impaired cell proliferation at the preceding stage. In contrast to the abnormality in cell proliferation, there was no change in cell death frequency (data not shown). These results indicate that a reduction in proliferating granule cell precursors is a notable feature in theZic1+/−Zic2+/kd cerebellum.

Proliferating cell numbers in theZic1/Zic2 cerebellum. Cells stained with anti-phospho-histone H3 antibodies (mitotic cells) were counted in the indicated regions of the E16 (A) and E17 (B) cerebellar sagittal sections. The ordinate indicates the mean number of the labeled cells per section. InC, mean numbers of the labeled cells in 1 mm of the EGL are indicated. The error bars indicate SD. ant EGL, Anterior EGL; post EGL, posterior EGL;VL, ventricular layer; int(intermed), intermediate zone that includes the area in the cerebellar primorium except EGL and VL; +/+,Zic1+/+Zic2+/+ ; Z1/Z2,Zic1+/−Zic2+/kd ; **p < 0.01
Progression of neuronal cell differentiation is de-regulated in the Zic1−/− and Zic1+/−Zic2+/kd cerebella
Recent studies revealed several genes controlling cerebellar development. We tested their expression in theZic1−/− andZic1+/−Zic2+/kd mice to evaluate their relationship with Zic proteins. Transcript levels of cyclin D1, cyclin D2, p27,and Wnt7a were quantitated in the cerebella of E17.5Zic1+/−Zic2+/kd ,Zic1−/− and their littermate wild-type controls (Fig. 8).d-cyclins are thought to drive cell cycle progression by regulating the phosphorylation of cellular substrates by the cyclin-dependent kinases Cdk4 and Cdk6 (Sherr and Roberts, 1999).Cyclin D1 levels were clearly decreased both in theZic1+/−Zic2+/kd and Zic1−/− cerebella, whereas the amount of cyclin D2 was not significantly changed. On the other hand, the Cdk inhibitor p27, which stops proliferation of granule cell precursors (Miyazawa et al., 2000), was increased in Zic1−/− and slightly increased inZic1+/−Zic2+/kd . In addition, expression of Wnt7a, which is usually expressed in the granule neurons postnatally (Lucas and Salinas, 1997), was prematurely enhanced both inZic1+/−Zic2+/kd and Zic1−/− .

Genes controlling cerebellar development are deregulated in theZic1−/− andZic1+/−Zic2+/kd cerebella. RT-PCR analysis was performed on cDNAs prepared from cerebella of E17.5Zic1−/− andZic1+/−Zic2+/kd and their littermate wild-type controls. G3PDH is a ubiquitously expressed gene. In the absence of reverse transcriptase [G3PDH (-RT)], no bands are detected, ensuring the absence of DNA. Amounts of cyclin D1, cyclin D2, p27, and Wnt7a transcripts were compared.
To further examine whether neuronal differentiation is affected, we performed immunohistochemical staining on the E15 cerebella. We detected a significant change in the distribution of the βIII tubulin protein, which is a marker of the differentiated neuron. In theZic1−/− cerebellum, the region of clustered cerebellar nuclei forming cells where both the Zic1 andZic2 transcripts were detected (Fig. 1K,O) was more strongly stained with the anti-βIII tubulin antibody than the wild-type control (Fig.9A,B). Corresponding areas contained more strongly p16-immunopositive cells (Fig.9C,D). TheZic1+/−Zic2+/kd cerebella showed a slightly enhanced distribution of βIII tubulin protein in the corresponding region (data not shown). These results indicate that premature neuronal differentiation occurs in bothZic1+/−Zic2+/kd and Zic1−/− cerebella at this stage.

Premature neuronal differentiation occurs in theZic1−/− cerebella. Immunohistochemical staining showing localization of βIII tubulin (A, B) and p16 (C, D) proteins were examined in the in E16 cerebella ofZic1−/− . Wild-type littermates (A, C) are shown as controls forB and D, respectively. The area surrounded by the arrowheads indicates the corresponding region between the mutants and wild-type controls, which showed significant differences.
DISCUSSION
Functional cooperation between Zic1 andZic2 in cerebellar development
This study investigated the involvement of Zic2 in cerebellar development by generatingZic1+/−Zic2+/kd mutant mice. The Zic1+/− andZic2+/kd cerebella were almost indistinguishable from wild type, although a subtle reduction of the size was observed in both theZic1+/− andZic2+/kd . In contrast, theZic1+/−Zic2+/kd cerebellum was abnormal and distinct from wild type. This is the first evidence for the cooperative role of the Zic family of genes in vertebrate development. Besides the cerebellar abnormality,Zic1+/−Zic2+/kd mice also showed changes in the septum, which was not affected either in Zic1+/− orZic2+/kd (J. Aruga, unpublished observation). These findings are consistent with the idea that Zic1 and Zic2 proteins have essentially similar functions. This idea is also supported by a recent study revealing that both Zic1 and Zic2 have similar DNA-binding properties and that both activate reporter genes in cultured cells (Mizugishi et al., 2001).
How can we explain the phenotypic difference betweenZic1−/− andZic2kd/kd mice? The difference may be attributed to the difference in the spatiotemporal expression patterns. At a primitive streak stage, Zic2 expression is strong in the forebrain region while Zic1 expression occurs later in the neuroectoderm and is enhanced in the hindbrain and the spinal cord (Nagai et al., 1997). This may explain the absence of apparent forebrain abnormality in the Zic1 mutant. It remains inconclusive as to whether the cerebellar phenotype of theZic2kd/kd mice reflects the lack of intrinsic Zic2 function in the cerebellum. Because the malformation of the posterior neuropore and forebrain is obvious as early as E9.5 and E10.5, respectively, cerebellar development may be secondarily affected by the preceding malformation.
Although Zic1 and Zic2 appear to have largely redundant functions, there remains the possibility that specificZic1/Zic2 interactions are required for cerebellar development. This could explain why the folial pattern in the trans-heterozygote is not identical to that found inZic1−/− . A “swapping” experiment (Hanks et al., 1995) is required to further investigate the functional equivalency of both proteins.
The role of Zic1 and Zic2 in cerebellar development
We suggest that Zic1 and Zic2 proteins may have following roles in cerebellar development. One is the regulation of granule cell expansion by Zic proteins. Our previous study showed that decrease in granule cell number is the most significant change in theZic1−/− cerebella. This was partly explained by the reduction in cell proliferation of the granule cell precursors. In theZic1+/−Zic2+/kd cerebella, the level of proliferating cells was significantly reduced at the beginning of phenotypic expression in the EGL. Because bothZic1 and Zic2 are expressed in the granule cell precursors in a similar manner, it is likely that Zic2controls granule cell proliferation in a similar manner toZic1.
The stimulation of granule cell proliferation by Zic1 andZic2 proteins may be regulated via the expression of cell cycle regulatory genes. cyclin D1, which was decreased in the mutant mouse cerebella, is known to drive cell proliferation in multiple tissues (Fantl et al., 1995; Sicinski et al., 1995). In addition, another member of the d-cyclin family,cyclin D2, which is predominantly expressed in the postnatal EGL (Ross et al., 1996), was shown to control cerebellar histogenesis (Huard et al., 1999). Although cerebellar abnormalities have not been reported in cyclin D1-deficient mice (Fantl et al., 1995; Sicinski et al., 1995), it is possible that cyclin D1 normally contributes to the control of granule cell proliferation and that its function is compensated for by other related molecules. The enhanced expression of p27 may be acting synergistically in the reduction of cell proliferation in the granule cell precursors, because deletion of the p27 gene restores normal development in cyclin D1-deficient mice (Geng et al., 2001).
In addition to their expression in granule cells and their precursors, both Zic1 and Zic2 are also expressed in the midline and anterior part of the vermal region of the cerebellar primordia at E16 and E18. This region contains clusters of cerebellar nuclei forming cells. The progression of neuronal differentiation in this region was affected in theZic1+/−Zic2+/kd and Zic1−/− cerebella. In addition, there was a difference in the expression pattern betweenZic1 and Zic2, which also may participate in generating the difference in the folial pattern between these two mutants.
We recently observed an inhibitory effect of mis-expressedZic1 in the neuronal differentiation in chick spinal cord (J. Aruga, T. Tohmonda, S. Homma, and K. Mikoshiba, unpublished observations). In addition, Brewster et al. (1998) showed that neuronal differentiation, as indicated by N-tubulin expression, is inhibited by Xenopus Zic2 expression. These facts suggest that the Zic family of proteins may play a general role in the inhibition of neuronal differentiation, via their effect on the expression of regulatory cell cycle genes. It is interesting that the Zic proteins are able to promote the differentiation of neuroectoderm from ectoderm in the Xenopus embryo (Nakata et al., 1997,1998), while at the same time inhibit neuronal differentiation. Zic proteins are also expected to have another role in the adult granule neuron, whereas all Zic genes are abundantly expressed.
Although this study showed that cyclin D1, p27,and Wnt7a are possible targets of the Zic proteins as transcriptional regulators, it is not clear how the Zic proteins act in the regulatory process. A comprehensive analysis of the downstream target genes and the assessment of possible relationships with other regulatory molecules are necessary for further understanding. Recent studies have indicated a functional interaction between Zic proteins and Gli proteins, which share a highly conserved zinc finger domain (Aruga et al., 1996, 1999; Brewster et al., 1998; Koyabu et al., 2001;Mizugishi et al., 2001). Gli genes are also expressed in developing cerebella and mediate Shh signaling which enhances proliferation of granule cell precursors (Wechsler-Reya and Scott, 1999). The role of the Gli proteins in cerebellar development has not yet been shown, and the intriguing question of their relevance to Zic proteins is still to be answered.
Footnotes
This work was supported by Special Coordination Funds for Promoting Science and Technology and grants from the Japanese Ministry of Education, Science, and Culture (to J.A. and K.M.) We thank Dr. Takaki Miyata for critical reading of this manuscript.
Correspondence should be addressed to Jun Aruga, Laboratory for Developmental Neurobiology, RIKEN Brain Science Institute, 2-1 Hirosawa, Wako-shi, Saitama 351-0198, Japan. E-mail:jaruga{at}brain.riken.go.jp.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}