

Abstract
Rats learned to lever-press for microinjections of the cholinergic agonist carbachol (30–500 pmol per infusion) or the acetylcholinesterase inhibitor neostigmine (7.5–75 pmol per infusion) into the posterior ventral tegmental area (VTA) of the brain. Intracranial carbachol self-administration was site-specific. Carbachol was not reliably self-administered into a site just dorsal to the VTA or into the adjacent substantia nigra and was self-administered only weakly into the adjacent anterior VTA or interpeduncular nucleus. Carbachol produced conditioned place preferences when injected into the posterior but not into the anterior VTA or sites dorsal to the posterior VTA. Rats self-administered carbachol less when it was co-infused with the muscarinic cholinergic receptor antagonist scopolamine or the nicotinic cholinergic receptor antagonist dihydro-β-erythroidine, and also when the rats were pretreated with the D1 dopamine antagonist SCH 23390. These findings implicate both nicotinic and muscarinic cholinergic neurotransmission in ventral tegmental reward function and suggest special involvement of the posterior portion of the VTA in cholinergic reward function.
- reinforcement
- self-administration
- acetylcholine
- nicotinic
- muscarinic receptors
- D1 receptors
- mesocorticolimbic dopamine neurons
The dopamine neurons projecting from the ventral tegmental area (VTA) to the nucleus accumbens have been implicated in instrumental behavior reinforced with lateral hypothalamic electrical stimulation, nicotine and a number of other drugs of abuse, and several natural rewards such as food, water, and sexual interaction (Wise and Rompre, 1989; Ikemoto and Panksepp, 1999). Studies of intracranial self-stimulation have also implicated acetylcholine in reward-related processes and suggested its interaction with ventral tegmental dopamine neurons (Redgrave and Horrell, 1976;Gratton and Wise, 1985; Yeomans et al., 1985). Recent evidence suggests that one of the excitatory inputs to the VTA dopamine neurons is acetylcholine, which is released by the axons from the laterodorsal and pedunculopontine tegmental nuclei (Oakman et al., 1995). Ventral tegmental dopamine neurons express both muscarinic and nicotinic cholinergic receptors (Clarke and Pert, 1985; Weiner et al., 1990), and administration of muscarinic or nicotinic agonists into the VTA can stimulate the dopamine neurons (Mereu et al., 1987; Calabresi et al., 1989; Lacey et al., 1990; Gronier and Rasmussen, 1998; Fiorillo and Williams, 2000) and cause dopamine release in the VTA (Gronier et al., 2000) and nucleus accumbens (Nisell et al., 1994; Blaha et al., 1996; Westerink et al., 1996; Gronier et al., 2000).
The rewarding effects of lateral hypothalamic brain stimulation are enhanced by ventral tegmental infusion of acetylcholine (Redgrave and Horrell, 1976) and attenuated by ventral tegmental infusions of muscarinic (Yeomans et al., 1985; Kofman and Yeomans, 1989; Yeomans and Baptista, 1997) or nicotinic (Yeomans and Baptista, 1997) antagonists. Ventral tegmental muscarinic antagonists also disrupt instrumental responses for food rewards (Ikemoto and Panksepp, 1996).
The direct rewarding effects of cholinergic agonists in the VTA have been demonstrated only with the conditioned place-preference paradigm. Both cytisine (Museo and Wise, 1994), a nicotinic agonist, and carbachol (Yeomans et al., 1985), a nonspecific cholinergic agonist, induce conditioned place preferences when injected into the VTA. In each case, however, anatomical localization of the site of rewarding action has not been determined by ineffective injections in surrounding regions. Moreover, the muscarinic specificity of the carbachol findings has not been confirmed. Finally, it has not been confirmed that animals will work for injections of either substance into this region. The purpose of the present investigation was to determine whether VTA carbachol or the acetylcholinesterase inhibitor neostigmine serves as an instrumental reinforcer and, if so, to characterize the rewarding effects of carbachol pharmacologically and anatomically.
MATERIALS AND METHODS
Subjects. The present study used 101 male albino rats [21 Sprague Dawley rats for experiment 1 from Charles River Laboratories (Raleigh, NC) and 80 Wistar rats for experiments 2–5 from Harlan Industries (Indianapolis, IN); 250–350 gm at the time of surgery]. Rats were housed initially in groups of two or three in a colony room with a constant temperature (21°C) and kept on a reversed 12 hr light/dark cycle (lights on at 9 P.M.). After surgery, they were housed individually. Food and water were available ad libitum except during testing. The animals were treated in accordance with the guidelines of the National Institutes of Health and the protocol was approved by the Animal Care and Use Committee of the Intramural Research Program.
Surgery. Stainless steel guide cannulas (24 gauge) were implanted under sodium pentobarbital (31 mg/kg, i.p.) and chloral hydrate (142 mg/kg, i.p.) anesthesia. Each animal was implanted with a guide cannula that ended 1.0 mm above one of several target sites. The cannulas were inserted at a 6° angle toward the midline for sites in the anterior or posterior VTA, the region just dorsal to the posterior VTA, or the interpeduncular nucleus (IPN); guides were implanted vertically for injections in the substantia nigra. Stereotaxic coordinates were 5.0 mm posterior to bregma, 1.6 mm lateral to the midline, and 7.8 mm ventral to the skull surface (measured along the trajectory of the angled cannula) for anterior VTA placements; 6.2 or 5.8 posterior, 1.3 lateral, and 7.8 ventral for posterior VTA placements; 6.2 posterior, 1.3 lateral, and 6.8 ventral for the region dorsal to posterior VTA; 6.0 posterior, 1.8 lateral, and 7.5 ventral for the substantia nigra; and 6.2 or 6.7 posterior, 1.1 lateral, and 8.2 ventral for the interpeduncular nucleus (incisor bar set at 3.3 mm below the interaural line). We started testing animals 5–7 d after surgery, according to the procedure described below.
Drugs. The muscarinic–nicotinic agonist carbachol, the cholinesterase inhibitor neostigmine bromide, the nicotinic antagonist dihydro-β-erythroidine hydrobromide, the muscarinic antagonist (−)-scopolamine methyl bromide, and the dopamine D1 antagonist R(+)-SCH 23390 hydrochloride, obtained from Research Biochemical International (Natick, MA), were used. SCH 23390 was used for intraperitoneal treatment and dissolved in 0.9% saline; the other drugs were used for intracranial treatments and dissolved in an artificial CSF consisting of (in mm): 148 NaCl, 2.7 KCl, 1.2 CaCl2, and 0.85 MgCl2 (pH adjusted to 6.5–7.8).
General procedure and apparatus. For operant testing, each animal was placed in a 30 × 22 × 24 cm chamber with a grid floor. The chamber was equipped with one (experiments 2, 3, and 4) or two (experiment 1) levers (4.5 cm wide × 2 mm thick protruding 2 cm from the wall) with a cue light just above each lever. The chamber was enclosed in a sound-attenuating box equipped with a ventilating fan. Each rat's 31 gauge injection cannula was connected by polyethylene tubing to a micropump (Ikemoto and Sharpe, 2001) hanging a few millimeters above the rat's head. Each pump consisted of a miniature step motor and a small plastic reservoir. When activated, the motor advanced a shaft into the reservoir, displacing drug solution into the injection cannula. The maximum number of infusions available per session was limited to 50 (in experiment 1) or 60 (in experiments 3 and 4) for carbachol and 80 (in experiment 2) for neostigmine, to minimize the possibility of tissue damage with repeated days of testing.
Experiment 1: self-administration of the cholinergic agonist carbachol. Unilateral guide cannulas were aimed at the anterior VTA of nine rats, the posterior VTA of six rats, or the interpeduncular nucleus of six rats. Each rat was placed in an operant chamber equipped with a cue light and one lever on each of two opposing walls. A response on one lever (active lever) caused an intracranial injection (50 nl) over 3 sec and illuminated the cue light just above the lever for 5 sec. Additional lever-presses during this 5 sec period were counted but did not have any experimental consequence. Responses on the other lever (inactive lever) had no experimental consequence. These rats were trained for five sessions. Sessions lasted for 3 hr or until 50 infusions were earned. In the first session, each rat earned 10 mm carbachol; in subsequent sessions it earned vehicle, 0.1, 1, and 10 mm carbachol.
Experiment 2: self-administration of the cholinesterase inhibitor neostigmine. Six rats that had guide cannulas aimed at the posterior VTA were placed individually in the operant chambers, each of which was equipped with a single lever and a cue light just above the lever. A response on the lever caused a 75 nl injection over 5 sec and illuminated the cue light for 5 sec. Additional lever-presses during this 5 sec period did not have any experimental consequence. Sessions lasted for 90 min or until 80 infusions were earned. Each rat was initially trained with 1 mm carbachol and then with 1, 0.3, 0.1, and 1 mm neostigmine. The sessions with carbachol and neostigmine solutions were always preceded by vehicle sessions.
To determine whether neostigmine is self-administered by drug- and procedure-naive rats, another group of six rats with posterior VTA cannulas was tested with the same operant program as the first group. Each earned vehicle infusions during sessions 1 and 4 and 1 mm neostigmine during sessions 2 and 3.
Experiment 3: neurochemical specificity. We determined whether the blockade of muscarinic, nicotinic, or D1-type dopamine antagonists attenuated carbachol-induced reward. Seven rats that had guide cannulas aimed for the posterior VTA were placed in operant chambers as described in experiment 2. A response on the lever caused a 100 nl carbachol injection over 6 sec and illuminated the cue light for 7 sec. Additional lever-presses during this 7 sec period did not have any experimental consequence. Sessions lasted for 60 min or until 60 infusions were earned. Each rat was trained with 10 mm carbachol during session 1 and with 1 mm carbachol during subsequent sessions 2–8. For sessions 4 and 5, the effects of the D1-type antagonist SCH 23390 on carbachol self-administration were examined. Thirty minutes before carbachol self-administration in session 4, half of the rats received 0.9% saline (1 ml/kg, i.p.) and half received SCH 23390 (0.05 mg/kg, i.p.); in session 5 the conditions were reversed. For sessions 6–8, the effects of co-infusion of the muscarinic antagonist methyl-scopolamine or the nicotinic antagonist dihydro-β-erythroidine with carbachol were tested. In session 6, half of the rats earned 1 mm carbachol co-infused with 1 mm scopolamine; the other half earned 1 mm carbachol co-infused with 10 mm DHβE. In session 7, each rat earned carbachol without the antagonists. In session 8, each rat earned carbachol co-infused with the antagonist that it did not get in session 6. For sessions 9 and 10, the effects of SCH 23390 (0.05 mg/kg) on 0.3 mm carbachol self-administration were examined. The testing procedure was the same as the one described for sessions 4–5, except that the animals earned 0.3 mmcarbachol instead of the 1 mm solution.
Experiment 4: additional injection-site analysis. Thirty-seven rats received unilateral guide cannulas aimed for the VTA and surrounding regions. Each was trained with the same testing procedure as described in experiment 3 except that it received 1 mm carbachol in three consecutive sessions. The rates of self-administration in the third session were compared across injection sites to identify the most probable site of effective action.
Experiment 5: conditioned place preference. To resolve concerns about the high rates of responding on the inactive lever in experiment 1, a conditioned place preference study was done. High rates of responding on the inactive lever might be thought to reflect accidental lever activations caused by drug-induced general activation [but see Bozarth and Wise (1981)] and not simply generalization between the otherwise identical active and inactive levers. The conditioned place-preference study was done to assess carbachol reward with a paradigm in which drug-induced general activation is not a possible confound (Bozarth, 1987; van der Kooy, 1987).
The place-conditioning chamber consisted of two compartments and an area connecting the compartments; a guillotine door separated each compartment from the connecting area. One compartment differed from the other by wall color (black vs white), floor type (net vs grid), and lighting; the amount of light was modulated in each compartment such that rats do not prefer one compartment to the other before place conditioning. In session 1, each rat was placed in the place-conditioning chamber for 15 min without any treatment; the rat had access to both compartments, and the time spent in each compartment was recorded. In session 2, each rat was confined to one of the compartments for 15 min, and in session 3, it was confined to the other compartment for 15 min during and after an injection. One half of the rats received 1 mm carbachol in session 2 and vehicle in session 3; the other half received these treatments in reversed order over the two sessions. Pairings of carbachol to the two compartments were counterbalanced among the rats. In session 4, each rat was placed in the chamber without any treatment with access to both compartments; the time spent in each compartment was recorded for 15 min. Sessions were separated by 24 hr.
Microinjections were given in the test apparatus. Each rat was connected to a microinjection cannula and placed in the assigned compartment. Each of the injections (500 nl) was delivered over 60 sec, with an additional 30 sec before the injection cannula was removed. Immediately after the injection, each rat was picked up, disconnected from the microinjection cannula, and placed back in the compartment. The rat stayed in the compartment for 15 min from the beginning of the injection.
A two-way mixed ANOVA was performed on place-preference time (the time spent in the carbachol-paired compartment minus the time spent in the vehicle-paired compartment) with injection site (anterior VTA, posterior VTA, and dorsal site) and conditioning (before and after injections). When a significant site × conditioning interaction was found, an additional paired t test was performed separately in each site on place-preference time between before and after injections.
Histology. When each rat completed its experimental procedure, it was anesthetized with a mixture of sodium pentobarbital (31 mg/kg) and chloral hydrate (142 mg/kg) and was perfused transcardially with 50 ml of 0.9% saline with 0.2% sodium nitrite followed by 100 ml of 4% paraformaldehyde in phosphate buffer solution (PBS). Its brain was removed and placed in 4% paraformaldehyde PBS for a day and stored in 18% sucrose PBS for 1–5 d until sectioning. Coronal sections (30 μm) at the microinjection site were cut with a cryostat. Sections were stained with cresyl violet. The placement of the injection cannula was confirmed by microscopic examination.
RESULTS
Experiment 1: self-administration of carbachol
Rats with cannulas in each of the three injection sites learned to self-administer carbachol (Fig.1A). This was reflected in a significant main effect of concentration on rate in the one-way ANOVA (anterior VTA, F(3,24) = 5.08,p < 0.01; posterior VTA,F(3,24) = 8.38, p < 0.005; and IPN, F(3,15) = 4.16,p < 0.05). With anterior VTA injections, rats self-administered the medial and high concentrations (1 and 10 mm) of carbachol more reliably than they self-administered the lower concentration or vehicle. With posterior VTA injections, rats self-administered more of the medial concentration (1 mm) than of the high (10 mm) or low concentration. With IPN injections, rats self-administered the highest concentration (10 mm) of carbachol more than vehicle. In the case of VTA injections, responding on the “active” lever was significantly greater than responding on the “inactive” lever (anterior VTA, F(1,8) = 21.57,p < 0.005; posterior VTA,F(1,5) = 13.71, p < 0.05) (Fig. 1B). In the case of IPN injections, there was a trend in the same direction, but it was not statistically significant (F(1,5) = 5.18,p = 0.07). When the animals were tested for the second time (fifth day) with the 10 mm concentration, self-administration was the same as it had been on the first day (F(2,18) = 1.44, p = 0.26, and F(2,18) = 0.12,p = 0.89). Representative injection sites are shown in Figure 2.
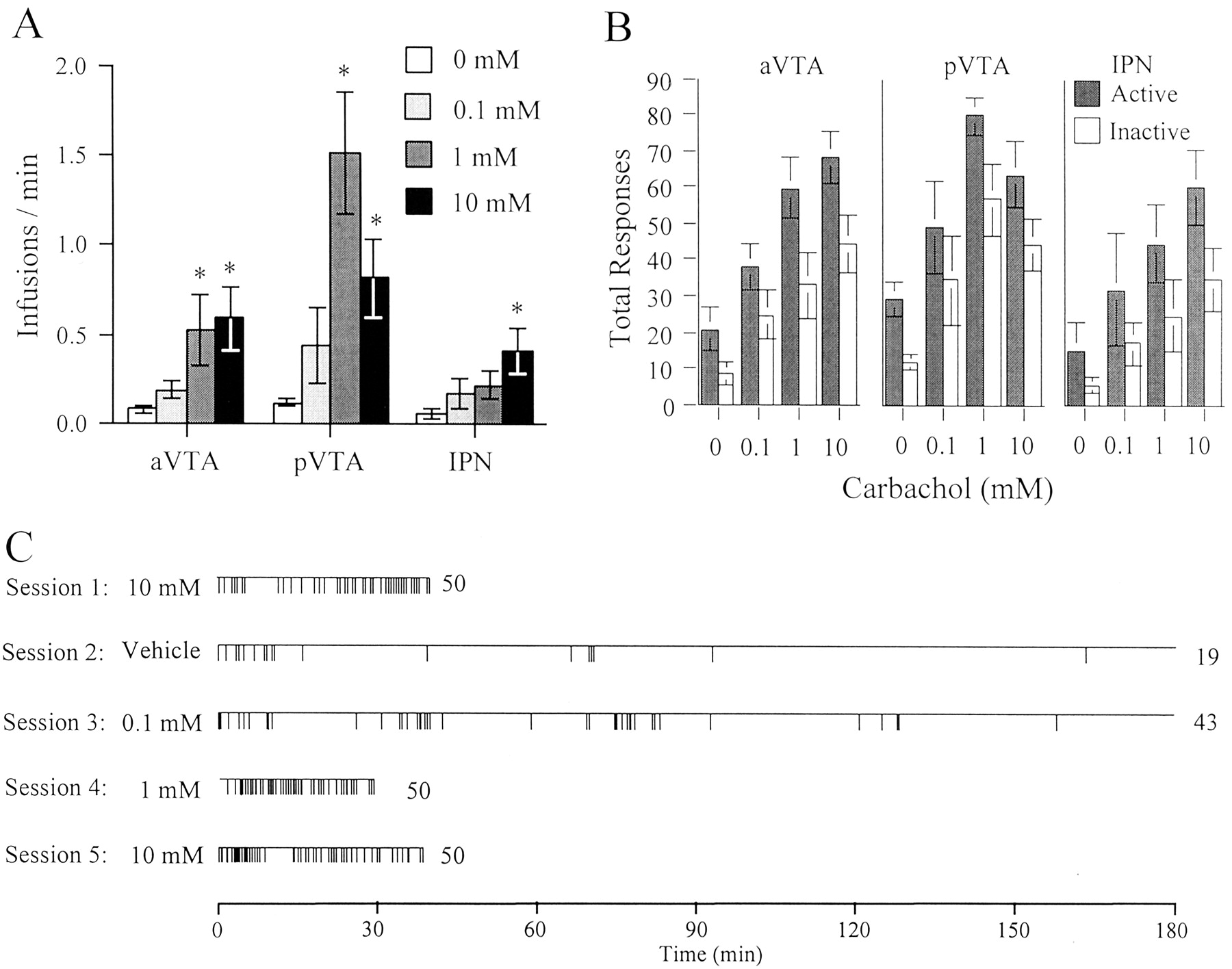
Carbachol self-administration into the ventral tegmental area. A, B, Mean (±SEM) infusions per minute and lever responses for 0, 0.1, 0.3, and 10 mm carbachol in sessions 2, 3, 4, and 5, respectively. Rats self-administered carbachol into the posterior VTA (pVTA) at faster rates than at the other sites (p < 0.05). In particular, 1 mmcarbachol solution elicited faster self-administration in the pVTA than the same concentration in the other sites (p< 0.01), as well as other concentrations (0 and 0.1 mm) in the same sites (p < 0.01). *p < 0.05, compared with respective vehicle. Rats administering carbachol solutions into the anterior VTA (aVTA) or pVTA, but not the interpeduncular nucleus (IPN), exhibited preference for the active lever over the inactive lever (B) (p < 0.05). Event records of a representative rat with pVTA injections are shown in C. Each vertical line on the horizontal lineindicates the time point of an infusion. The number justright of the horizontal line indicates total infusions in that session.

Coronal sections showing the locations of injection sites (indicated by arrows) of representative rats. Each rat had a unilateral guide cannula aiming for one of the following sites. A depicts the anterior ventral tegmental area (aVTA), which is defined as the region just dorsal to the medial mammillary nucleus (MM) and just medial to the substantia nigra (SN).B depicts the posterior VTA (pVTA), which is defined as the region just dorsal to the interpeduncular nucleus (IPN), just medial to the SN, and just ventral to the red nucleus (RN). C depicts the IPN, which is found just ventral to the pVTA. FR, Fasciculus retroflexus; ML, medial lemniscus.
In the case of the posterior VTA injections but not the anterior VTA or IPN injections, the animals worked less for the highest concentration than for the intermediate concentration (significant interaction between concentration and site,F(6,54) = 3.92, p < 0.005). This was reminiscent of the inverted “U”-shaped dose–response curves seen with intravenous amphetamine and heroin self-administration. The animals worked more (that is, they earned their maximum number of injections more quickly) for posterior VTA injections than for injections in either of the other two sites (p < 0.01, Student-Newman–Keuls post hoc tests after a significant main effect for site in the one-way ANOVA, F(2,18) = 7.93,p < 0.005). The patterns of responding for the 5 d of testing are shown for a representative animal in Figure1C. In the first session, this animal quickly learned to self-administer carbachol and self-administered the 10 mm concentration regularly throughout the session. In the second session, the rat was switched to vehicle reinforcement. Here this rat lever-pressed quickly in the first few minutes of the session but did not sustain regular responding after the first 15 min. In the third session this animal was switched to the lowest concentration of carbachol (0.1 mm). Here the rat again self-administered the drug, but in intermittent binges reminiscent of the patterns seen with saline reinforcement. In this case, however, the binges were repeated and sustained for longer periods, and the total number of infusions per session was greater than in the case of the saline condition. In the fourth session, this animal was switched to the intermediate concentration of carbachol (1 mm); this concentration maintained the highest injection rate in the posterior VTA group, with sustained responding and regular spacing throughout the session. Finally, in the fifth session, this animal was switched back to the highest concentration (10 mm); here again the rat responded steadily throughout the session, at a rate comparable with the earlier test with the same dose.
Experiment 2: self-administration of neostigmine
In the second experiment, a new group of rats learned to self-administer either 1 mm neostigmine or, replicating experiment 1, 1 mm carbachol into the posterior VTA. Self-administration of 1 mm neostigmine was similar to the self-administration of 1 mm carbachol, both in overall rate of infusions (Fig. 3A) and in temporal pattern of self-administration (Fig. 3B). Lower concentrations of neostigmine (0.1 and 0.3 mm) were also self-administered more than vehicle (T5 = 3.28, p < 0.05 and T5 = 3.29, p < 0.05, respectively). Self-administration of the 1 mm concentration of neostigmine in the fourth session was as strong and reliable as responding for the 1 mm concentration in the last session (Fig.3A), indicating that there was no deterioration of reinforcement with repeated injections in repeated tests. When another group of drug-naive rats was tested with 1 mmneostigmine for the first time, similar results were found (a significant difference between the first and second sessions,T5 = 4.81, p < 0.005) (Fig. 3C).

Self-administration of the cholinesterase inhibitor neostigmine into the posterior VTA. Each session lasted 90 min or until 80 infusions were obtained. A depicts mean infusion rates with SEM (n = 6) of 1 mmcarbachol (CARB) and 1, 0.3, 0.1, and 1 mmneostigmine (NEO) examined in this sequence. Effects of these chemical treatments were compared with preceding vehicle treatments, using paired t tests. *p< 0.05 compared with vehicle. B depicts event records showing self-administration of a representative rat. Eachvertical line on the horizontal lineindicates the time point of an infusion. The number justright of the horizontal line indicates total infusions in that session. The rat exhibited well spaced self-administration when receiving 1 mm carbachol (T5 = 3.89; p < 0.05) or 1 mm neostigmine (T5 = 3.77; p < 0.05). When receiving vehicle, the rat obtained infusions mostly during the first 15 min of the session. C depicts the acquisition of neostigmine self-administration. Drug-naive and procedure-naive rats (n = 6) received vehicle (sessions 1 and 4) and 1 mm neostigmine (sessions 2 and 3) into the posterior VTA. The rats learned to self-administer neostigmine quickly when receiving it for the first time during session 2 and significantly reduced self-administration rates when receiving vehicle during session 4 (*p < 0.05). The data are mean infusion rates with SEM.
Experiment 3: neurochemical specificity
As found in the first experiment and replicated in the second, rats quickly learned in the third experiment to self-administer carbachol into the posterior VTA. Responding for 1 mmcarbachol was evenly paced. Co-infusion of the nicotinic antagonist DHβE or the muscarinic antagonist scopolamine attenuated responding (Fig. 4A) (Student's–Newman–Keuls tests: a significant difference,p < 0.05, between carbachol alone and carbachol with DHβE and a significant difference, p < 0.05, between carbachol alone and carbachol with scopolamine, after a significant main effect for treatment in the one-way ANOVA;F(2,12) = 11.73; p < 0.005).

Effects of the muscarinic antagonist scopolamine (SCOP) or the nicotinic antagonist dihydro-β-erythroidine (DHβE) on carbachol self-administration into the posterior VTA. These antagonists were administered in mixtures with 1 mm carbachol (CARB). Each session lasted 60 min or until 60 infusions were obtained. The presence of SCOP or DHβE significantly attenuated infusion rates of intra-VTA carbachol (*p < 0.05).A depicts mean infusion rates with SEM (n = 7). B depicts event records of a representative animal. Each vertical line on thehorizontal line indicates the time point of an infusion. The number just right of thehorizontal line indicates total infusions per session.
Pretreatment with the D1 antagonist SCH 23390 caused cessation of self-administration of 0.3 mm carbachol and dramatically decreased, but did not completely cease, self-administration of 1 mm carbachol. These observations were confirmed by analyses in which the first six interinfusion intervals were compared with the last six interinfusion intervals (Fig.5C). Latency to the first infusion was not affected by SCH 23390 pretreatment (Table1); the latencies to self-administer the first infusion when animals were treated with saline did not differ from those when the animals were treated with the D1 antagonist for either low or high carbachol reward (T6 = 0.44, p = 0.68 or T6 = 0.63, p = 0.55, respectively). Thus the treatment did not seriously compromise the response capacity of the animals.

Effects of the D1 antagonist SCH 23390 (SCH) on carbachol (CARB) self-administration into the posterior VTA. The D1antagonist (0.05 mg/kg, i.p.) or saline (1 ml/kg) was administered 30 min before the self-administration session with 0.3 or 1 mmcarbachol. Each session lasted 60 min or until 60 infusions were obtained. The SCH 23390 treatment significantly reduced the rate of self-infusion of intra-VTA 0.3 or 1 mm carbachol (**p < 0.005 or *p < 0.05, respectively). A depicts mean infusion rates with SEM (n = 7). B depicts event records of a representative animal. A vertical line on thehorizontal line indicates the time of an infusion. Thenumber just right of thehorizontal line indicates total infusions per session.C depicts the first six and last six infusion intervals when animals earned 0.3 and 1 mm carbachol. The first six infusion intervals when animals were treated with the D1antagonist did not differ from those when treated with saline for both low (0.3 mm) and high (1 mm) carbachol rewards (F(1,6) = 2.28, p = 0.18 and F(1,6) = 2.32,p = 0.18, respectively). Moreover, animals delivered infusions at similar intervals between the first and last six intervals when treated with saline for both low and high carbachol rewards (F(1,6) = 0.26,p = 0.63 and F(1,6)= 3.55, p = 0.11, respectively). However, when rats were treated with SCH 23390, the last intervals increased more significantly than the first intervals for both low and high carbachol rewards (F(1,6) = 28.53,p < 0.005 andF(1,6) = 12.67, p< 0.05, respectively) and the last intervals of saline treatment (F(1,6) = 40.13, p< 0.001 and F(1,6) = 14.42,p < 0.01, respectively).
Mean latency in seconds (SEM) to self-administer the first infusion as a function of pretreatments (saline vs the dopamine antagonist SCH 23390) (n = 7)
Experiment 4: additional injection-site analysis
When additional rats were tested with 1 mm carbachol, it was confirmed that the strongest self-administration (highest rates of injection) occurred when carbachol was injected into an area just dorsal and lateral to the IPN at the level of the posterior VTA (Fig. 6). Rats self-administered carbachol into the posterior VTA more than they did into the anterior VTA, interpeduncular nucleus, or the regions just lateral or dorsal to the VTA (Student's–Newman–Keuls tests: significant differences,p < 0.05, after a significant main site effect in the one-way ANOVA, F(4,60) = 13.80,p < 0.0001). Mean infusion rates were 1.13, 0.52, 0.17, 0.30, and 0.16, respectively. Minimal responding was seen with injections directly into the IPN or with injections dorsal or lateral to the region of VTA dopamine neurons. Infusions in the anterior VTA, just rostral to the best posterior VTA sites, were more effective than IPN injections or more dorsal or lateral injections.

Effectiveness of carbachol self-administration. The injection sites of the rats from experiments 1 (n = 21), 3 (n = 7), and 4 (n = 37) are depicted on coronal drawings [adapted and modified from Paxinos and Watson (1997)]. Self-administration rates with 1 mm carbachol were classified into six levels. The numbers in the drawings indicate distances from bregma. aVTA, Anterior ventral tegmental area;IPN, interpeduncular nucleus; pVTA, posterior ventral tegmental area; RN, red nucleus;SN, substantia nigra; SUM, supramammillary nucleus.
Experiment 5: conditioned place preference
Injections of carbachol into the posterior VTA induced conditioned place preference (Fig. 7) (a significant difference in place-preference times between before and after injections; T7 = 3.52;p < 0.01). On the other hand, injections of carbachol into the anterior VTA or the regions just dorsal to the posterior VTA did not induce conditioned place preference (no reliable differences in place-preference times between before and after injections;T7 = 1.48 , p= 0.89 and T7 = 1.19,p = 0.28, respectively). Differential conditioning effects of carbachol among the three sties are confirmed by a significant site × conditioning interaction (F(2,21) = 4.31; p < 0.05 with a two-way ANOVA).

Time spent in carbachol-paired compartment minus time spent in vehicle-paired compartment (place-preference score). Conditioned place preference induced by injections of the cholinergic agonist carbachol (1 mm in 500 nl) into the posterior VTA (n = 8) but not the anterior VTA (n = 8) or the region just dorsal to the posterior VTA (dorsal region; n = 8) (significant difference compared with place-preference time before conditioning; *p < 0.05). There was no differential preference for the compartment before pairing with carbachol among the three groups of rats.
DISCUSSION
Behavioral considerations
Our rats typically learned to lever-press for infusions of carbachol or neostigmine within the first session. Evidence that the animals had mastered the task and reached stable levels of responding so quickly includes the following: (1) responding was sustained throughout the first session with relatively regular inter-response times; (2) typical extinction patterns (responding in the early minutes of the session but response slowing and cessation thereafter) were seen when saline was substituted for carbachol; and (3) intake on the fifth day was similar to that on the first day, when animals were trained with the same dose. Responding was faster at the 1 mmconcentration than at the 10 mm concentration, suggesting saturation of the system or satiation of the animal for tens of seconds after each injection.
Responding for the lower concentrations of carbachol was strongest when the drug was injected into the posterior VTA. In this case responding decreased when the highest concentration was given, suggesting a biphasic dose–effect function similar to that seen with intravenous drug self-administration. Weaker responding and monotonic dose–effect curves for the adjacent anterior VTA and slightly more ventral interpeduncular nucleus suggest that satiating concentrations were reached only in the case of the posterior VTA, a finding that strongly implicates the posterior VTA as the site of rewarding action seen in the present study. This seems confirmed by the conditioned place-preference experiment, in which place preferences were established by carbachol only when it was injected into the posterior VTA.
Although high rates of responding were seen on each of the two levers, the animals showed reliable preferences for the active lever. High rates of responding on the inactive lever were not surprising, because the levers were identical and placed in similar positions on opposite walls of the cage. Generalization from the active to the inactive lever is normally seen in the early stages of instrumental conditioning. The more limited number of sessions in the intracranial drug self-administration paradigm precludes the stronger discrimination that can be established in intravenous drug self-administration studies. Although responding more on the inactive lever in the drug reward condition than in the saline condition can result from the sum of two factors—learned generalization between the two levers and the nonselective activating effect of the drug—it is unlikely that drug-induced activation contributed strongly to the inactive lever counts. In a similar study with intracranial morphine reinforcement, no inactive lever counts were seen in yoked animals that had only an inactive lever but received the same injections as those earned by their yoked partners (Bozarth and Wise, 1981). This was true despite the fact that the drug injections caused robust circling locomotion in the yoked animals. This suggests that the inactive lever responding in the two-lever task results not just from nonspecific activating effects of the rewarding drug, but because of the association of lever-pressing with that drug. In any case, that the posterior VTA carbachol injections were rewarding and not just activating is confirmed by fact that posterior but not anterior VTA carbachol established learned preferences for the drug-associated place that were expressed in the drug-free condition on the day after the drug treatment.
Neuronal mechanisms of reward induced by ventral tegmental carbachol and neostigmine
Possible inter-regional circuitry
Ventral tegmental dopamine neurons project to various forebrain regions. Of these regions, the nucleus accumbens has clearly been implicated in reward (Wise and Rompre, 1989; Ikemoto and Panksepp, 1999). Thus, the VTA–accumbens dopamine pathway seems the most likely mediator of the rewarding effects of ventral tegmental carbachol and neostigmine. Indeed, it has been shown that intra-VTA application of carbachol, neostigmine, the muscarinic agonist oxotremorine M, or nicotine increases extracellular levels of dopamine or dopamine signals in the nucleus accumbens (Nisell et al., 1994; Westerink et al., 1996; Gronier et al., 2000).
Additional evidence of the importance of the dopamine system is that the D1 receptor antagonist SCH 23390 (0.05 mg/kg) disrupted markedly the self-administration of carbachol (Fig. 5). It is unlikely that the reduction in carbachol self-administration was caused by sedation or motor impairment induced by SCH 23390, because rats initiated self-administration normally (Table 1) and delivered carbachol infusions at normal intervals at the beginning of those sessions (Fig. 5C). Moreover, the same dose of SCH 23390 tended to cause termination of self-administration of the low carbachol reward (0.3 mm), whereas it simply increased inter-infusion intervals in the case of the high reward (1 mm). These results are consistent with the hypothesis that the blockade of D1 receptors disrupts the rewarding effect of ventral tegmental carbachol. However, because D1 receptors in both the nucleus accumbens (Ikemoto et al., 1997a) and the VTA (Ranaldi and Wise, 2001) are each implicated in reward, the localization of the D1 receptors involved in intra-VTA carbachol reward remains to be determined.
The source of acetylcholine in the VTA is cholinergic neurons projecting from the laterodorsal and pedunculopontine tegmental nuclei (Oakman et al., 1995). It is not clear which of these cholinergic nuclei is more important; limited evidence, if anything, favors the laterodorsal tegmental nucleus. The lesions of the laterodorsal tegmental nucleus, but not the pedunculopontine tegmental nucleus, attenuate accumbal dopamine efflux induced by intra-VTA neostigmine (Blaha et al., 1996). Intra-VTA injections of scopolamine or the nicotinic antagonist mecamylamine attenuate accumbal dopamine efflux evoked by electrical stimulation of the laterodorsal tegmental nucleus (Forster and Blaha, 2000). These findings suggest that the pathway from the laterodorsal tegmental nucleus projecting to the VTA and the nucleus accumbens is involved in the induction of reward and related processes.
It should be noted that rats self-administered the cholinesterase inhibitor neostigmine without direct stimulation of acetylcholine release in the VTA. Therefore, ventral tegmental acetylcholine appears to be tonically released. Indeed, cholinergic neurons of the laterodorsal and pedunculopontine tegmental nuclei appear to fire tonically, in particular, during the awake state (el Mansari et al., 1989; Steriade et al., 1990; Kayama et al., 1992). These electrophysiologial findings, in turn, suggest that acetylcholine neurotransmission in the VTA is involved in more general processes necessary during the awake state, and the present findings add that phasic increase of acetylcholine neurotransmission in the VTA appears to trigger a rewarding state.
Differential self-administration as a function of injection site
The finding that cholinergic agents have differential rewarding effects in the anterior and posterior VTA should not be surprising given the heterogeneity of this region that has been seen with GABAergic agents. Rats self-administer GABAAagonists into the posterior but not the anterior VTA (Ikemoto et al., 1998); conversely, rats self-administer GABAAantagonists into the anterior but not the posterior VTA (Ikemoto et al., 1997b). Moreover, we found recently that rats self-administer the μ-opioid agonist endomorphin-1 more avidly into the posterior VTA (Zangen et al., 2002). Thus, the present study adds to the evidence that the anterior and posterior VTA play differential roles in reward function. The drugs that are most effective in the posterior VTA do not appear to share the identical “hot spot.” The most sensitive site for carbachol reward is as shown in Figure 6. The most sensitive site for endomorphin-1 reward appears to be slightly more posterior than that (Zangen et al., 2002), seemingly corresponding to the heaviest site of μ-receptors within the VTA (German et al., 1993). The most sensitive site of the GABAA agonist muscimol appears to be even more posterior, around the central linear nucleus of the VTA (Ikemoto et al., 1998).
Possible cytoarchitectonic mechanisms
Coadministration of carbachol with the muscarinic antagonist scopolamine or the nicotinic antagonist dihydro-β-erythroidine attenuated the self-administration of intra-VTA carbachol (Fig. 4). These results are consistent with previous studies on instrumental responding maintained by brain stimulation reward. Yeomans and Baptista (1997) reported that the intra-VTA microinjection of either muscarinic or nicotinic antagonists attenuates lever-pressing maintained by lateral hypothalamic stimulation reward. Moreover, such self-stimulation is also attenuated by intra-VTA injections of antisense oligonucleotide for M5 receptor mRNA, which specifically reduces functional muscarinic M5 receptors (Yeomans et al., 2000). Intra-VTA injections of dihydro-β-erythroidine or 6-OHDA lesions of the mesoaccumbens dopamine system also attenuate intravenous self-administration of nicotine in rats (Corrigall et al., 1992, 1994). These findings as well as the present ones implicate both muscarinic and nicotinic neurotransmission in ventral tegmental reward process. However, the relative contribution of each to the rewarding effect of carbachol remains to be determined.
We cannot exclude the possibility that cholinergic receptors localized on presynaptic terminals in the VTA may play some role in reward induced by ventral tegmental cholinergic transmission. Ventral tegmental dopamine neurons receive glutamatergic as well as GABAergic inputs. The stimulation of presynaptic muscarinic M3 receptors located on GABAergic terminals appears to attenuate GABA release onto ventral tegmental dopamine neurons (Grillner et al., 2000). Moreover, the stimulation of presynaptic nicotine receptors in the VTA appears to enhance glutamatergic inputs to dopamine neurons for a prolonged period (Mansvelder and McGehee, 2000) and inhibit GABAergic inputs (Mansvelder et al., 2002). This mechanism may potentially be more important in maintaining instrumental responding than the nicotinic receptors located on dopamine neurons that appear to desensitize in seconds (Pidoplichko et al., 1997). Thus, carbachol in the VTA may have indirect as well as direct effects on dopaminergic neurons that contribute to mesolimbic reward circuitry.
In summary, the present study demonstrates that the cholinergic agonist carbachol and the acetylcholinesterase inhibitor neostigmine are rewarding when injected directly into the posterior ventral tegmental area but not surrounding brain regions. The drugs are self-administered intracranially, and carbachol induces conditioned place preference when injected into this region. These data fit with several other lines of evidence suggesting a cholinergic input to the dopaminergic neurons of the ventral tegmental area that have been implicated broadly in reward function.
Footnotes
This work was supported by The Intramural Research Program of the National Institute on Drug Abuse/National Institutes of Health. We thank Emily Roach for conducting a pilot experiment and Brian Witkin for helping with neostigmine and place-conditioning experiments.
Correspondence should be addressed to Satoshi Ikemoto, National Institute on Drug Abuse, Behavioral Neuroscience Branch, 5500 Nathan Shock Drive, Baltimore, MD 21224. E-mail:sikemoto{at}intra.nida.nih.gov.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}