

Abstract
Neurofilament assembly requires at minimum the polymerization of neurofilament light chain (NF-L) with either neurofilament medium chain (NF-M) or neurofilament heavy chain (NF-H) subunits, but requirements for their axonal transport have long been controversial. Using a gene deletion approach, we generated mice containing only NF-L or NF-M. In vivo pulse radiolabeling analyses in retinal ganglion cell neurons revealed that NF-L alone is incapable of efficient transport, whereas nearly one-half of the normal level of NF-M is transported along optic axons in the absence of the other triplet subunits. Under these conditions, however, NF-M transport is completely abolished by deleting α-internexin. Our results strongly suggest that efficient neurofilament protein transport in vivo minimally requires hetero-oligomer formation. They also show that NF-M can partner with intermediate filament proteins other than the NF-H and NF-L subunits in neurons to support slow transport and possibly other functions of neuronal intermediate filaments.
- neurofilaments
- neurofilament medium subunit
- neurofilament light subunit
- α-internexin
- hetero-oligomer
- slow axonal transport
- optic axons
- knock-out mice
- intermediate filaments
- cytoskeletal protein
Introduction
Neurofilaments are the most abundant structural components of large myelinated axons. They are obligate heteropolymers in vivo composed of neurofilament light chain (NF-L), neurofilament medium chain (NF-M), and neurofilament heavy chain (NF-H) (Ching and Liem, 1993; Lee et al., 1993; Nakagawa et al., 1995). Neurofilament formation is believed to begin with the association of two subunits into coiled-coil dimers, which then form tetramers and eventually higher-order assemblies of 10 nm filaments (Fuchs and Weber, 1994). The NF-M and NF-H by themselves cannot form filaments in vivo (Ching and Liem, 1993; Lee et al., 1993; Nakagawa et al., 1995). Many CNS neurons also contain varying levels of α-internexin, a type IV intermediate filament subunit (Fliegner and Liem, 1991) expressed abundantly in embryonic neurons but at lower levels after birth (Kaplan et al., 1990; Fliegner et al., 1994). Although α-internexin colocalizes with neurofilaments and can form filaments with each of the triplet proteins in vitro (Ching and Liem, 1993), the composition of the filaments it forms is not known.
The prominence of neurofilament pathology in neurodegenerative diseases, including the pathogenic role of NF-L mutations in Charcot-Marie-Tooth disease (Mersiyanova et al., 2000; De Jonghe et al., 2001; Brownlees et al., 2002; Perez-Olle et al., 2002; Jordanova et al., 2003), has focused increased attention on the mechanisms of neurofilament assembly and transport in the adult nervous system in vivo. Specifically, the molecular requirements for neurofilament protein movement, including the motors and the subunit(s) essential for translocation, are unknown. Moreover, a long-standing debate has centered on the form of the transported neurofilament proteins (Baas and Brown, 1997; Bray, 1997; Hirokawa et al., 1997; Galbraith et al., 1999; Yabe et al., 1999; Prahlad et al., 2000; Terada and Hirokawa, 2000; Yuan et al., 2000; Shah and Cleveland, 2002). By one view, neurofilament proteins are assembled in the cell body and transported into axons as fully assembled neurofilaments (Baas and Brown, 1997). Supporting this view is the direct observation of a subpopulation of green fluorescent protein-tagged neurofilaments moving bidirectionally in growing axons of cultured embryonic neurons (Roy et al., 2000; Wang et al., 2000; Ackerley et al., 2003). Alternatively, neurofilament proteins have been proposed to move in subunit form (monomer or oligomer) and exchange with those in the neurofilaments that form a stable network in the axon (Hirokawa et al., 1997). In support of this model, virally expressed NF-M were transported at slow rates without NF-L or NF-H into axons in a transgenic mouse model that expresses a mutant NF-H linked to β-galactosidase, which aggregates the neurofilaments within the perikaryon and substantially reduces the numbers of neurofilaments in the axons (Terada et al., 1996). The differential dynamics of NF-L and NF-H in axons in vivo have also been interpreted as support for subunit movement (Takeda et al., 1994).
In the present study, we directly addressed in mice whether or not neurofilament subunits are capable of independent translocation and whether subunit assembly is essential for movement. By analyzing slow transport in mutant mice containing only NF-L or NF-M, we show that the minimal requirement for efficient transport is a hetero-oligomer, that either NF-L or α-internexin can partner with NF-M to support slow transport of the complex along axons, and that NF-L alone is not efficiently transported despite its ability to form filaments in vitro. These findings define important structural requirements for the interactions of neurofilament subunits with the putative motors that mediate slow transport.
Materials and Methods
Generation of knock-out animals. Adult male or female mice of the C57BL/6J strain, aged 3-4 months at the time of injection or death, were used in all experiments. Mice were housed at 23°C on a 12 hr light/dark cycle and were maintained on Lab Chow (Purina Mills, Gray Summit, MO) supplied ad libitum. Production of NF-L knock-out (LKO) (Zhu et al., 1997) and α-internexin knock-out (α-IKO) mice (Levavasseur et al., 1999) has been described previously. NF-H and NF-M double knock-out (HM-DKO) mice were generated by crossbreeding HKO (Rao et al., 1998) with MKO mice (Elder et al., 1998; Jacomy et al., 1999). NF-H and NF-L double knock-out (HL-DKO) were generated by crossbreeding HKO (Rao et al., 1998) with LKO mice. α-IL-DKO was generated by crossbreeding α-IKO with LKO mice.
Isotope injections. Adult mice were anesthetized with 13-20 μl/gm body weight of Avertin (0.5 gm of tribromoethanol and 0.25 gm of 2-methyl-2-butanol in 39.5 ml of distilled water) and received 0.20 μl of PBS, pH 7.4, which contained 50 μCi of l-[35S]methionine (specific activity, 1175 Ci/mmol) purchased from PerkinElmer Life Sciences (Boston, MA). Injections were made into the vitreous of each eye with a glass micropipette (70-100 μm) apparatus (Nixon and Logvinenko, 1986).
Axonal transport studies. The retinal ganglion cells of adult LKO, α-IKO, HM-DKO, HL-DKO, α-IL-DKO mice, and their corresponding wild-type controls were radiolabeled by intravitreal injection. The primary optic pathway consists of the optic nerve, the optic chiasm, and part of the optic tract extending to, but not including, terminals in the lateral geniculate nucleus. Five hours or 3, 7, and 14 d after injection, optic pathways from groups of three animals were dissected and cut into eight consecutive 1 mm segments on a micrometer slide on dry ice. Triton X-100-soluble and Triton X-100-insoluble preparations from each segment were subjected to SDS-PAGE, electrotransfer of proteins, phosphorimaging, and autoradiography. These experimental protocols were approved by the New York University and Nathan Kline Institute Institutional Animal Care and Use Committees under the guidelines of the Institutional Animal Care and Use Committee of the United States.
Tissue preparation, SDS-PAGE, and immunoblot analysis. Retina or each optic pathway segment pooled from three mice was homogenized in 125 μl of cytoskeleton extraction buffer (50 mm Tris-HCl, pH 6.8, 1% Triton X-100, 10% glycerol, 0.15 m NaCl, 2 mm EDTA, 2 mm PMSF, 50 μg/ml leupeptin, and 2.5 μg/ml aprotinin) (Chiu and Norton, 1982). The homogenate was centrifuged at 15,000 × g for 30 min at 4°C to be separated into Triton X-100-soluble and Triton X-100-insoluble particulate fractions. One-dimensional SDS-PAGE was performed (Laemmli, 1970) using 320 mm slab gels containing 5-15% linear polyacrylamide gradients. Two-dimensional gel electrophoresis was performed as described previously (Yuan et al., 2000). PAGE-separated proteins were transferred to nitrocellulose membranes. The transferred proteins were immunostained by means of an alkaline-phosphatase-conjugated indirect antibody procedure. The distribution of [35S]methionine-labeled proteins was visualized by autoradiography or phosphorimaging. The radioactivity associated with each of the NF subunits was determined by phosporimaging or computer-assisted densitometry. Antibodies used were monoclonal antibodies (mAbs) to NF-L (NR4), NF-M (NN18), NF-H (N52) (Sigma, St. Loiis, MO), and α-internexin (MAB5224; Chemicon, Temecula, CA), respectively.
Electron microscopy. Mice were anesthetized with halothane gas, and the tissue was fixed through intracardial perfusion with 4% paraformaldehyde, and 5% glutaraldehyde in 0.1 m PBS, pH 7.4, at room temperature. The optic nerve was dissected and processed as described previously (Nixon et al., 1994), segmented in 1.2 mm pieces, cleared in propylene oxide, and embedded in Medcast (Ted Pella, Redding, CA). Ultrathin sections were collected, and the section containing the initial portion of the retinal excavation was then used as a standard reference point. Ultrathin sections were stained with uranyl acetate and lead citrate and examined in an EX electron microscope (model JEM1200; Jeol, Peabody, MA) at 80 kV.
Results
NF-L alone is not efficiently transported in vivo
To determine whether NF-L alone is efficiently transported without NF-H and NF-M, we generated a line of NF-H and NF-M double knock-out (HM-DKO) mice containing only NF-L by crossbreeding NF-H null mice (Rao et al., 1998) with NF-M knock-out mice (Elder et al., 1998; Jacomy et al., 1999). Analysis of slow axonal transport in optic axons at 3 d after injection of [35S]methionine demonstrated not only the absence of NF-H and NF-M transport but also the disappearance of NF-L transport in the HM-DKO animals (Fig. 1). The complete loss of NF-L in HM-DKO mice was confirmed by two-dimensional gel electrophoresis (Fig. 1d,f). Immunoblotting studies with mAb to NF-L (NR4) revealed no detectable NF-L in the optic nerves of HM-DKO mice (Fig. 1c,e,g). The observations that NF-L mRNA levels were not affected by the disruption of NF-H and NF-M genes (Jacomy et al., 1999) but NF-L protein by Western blot analysis was at low level (<10%) in the retinal ganglion cells compared with wild-type controls imply additional posttranscriptional effects on NF-L (e.g., reduced translation of NF-L mRNA or faster degradation of the protein) (Jacomy et al., 1999). At present, we cannot differentiate among these possibilities.

No detectable transport of NF-L in optic axons in the absence of NF-M and NF-H. Slow axonal transport was determined by intravitreal injection of radiolabeled [35S]methionine into 3- to 4-month-old control (a) and HM-DKO (b) mice. At 3 d after injection, the optic pathways were cut into eight 1 mm segments at consecutive levels extending from the eye to the lateral geniculate body and were fractionated into Triton X-100-soluble and Triton X-100-insoluble fractions. The cytoskeleton proteins were separated on 5-15% SDS-polyacrylamide gels, transferred to nitrocellulose, and visualized by autoradiography and phosphorimaging. The positions of each cytoskeletal proteins are indicated. Labeling of NF-M and NF-H proteins are seen in control but not in HM-DKO optic axons. The absence of a labeled NF-L in HM-DKO mice traveling at an SCa rate is partially obscured by the labeled Hsc70 (heat shock cognate 70) protein, which is transported at a faster SCb rate of transport (Yuan et al., 2000). NF-L subunit, however, was not detected in optic axons of HM-DKO mice by immunoblotting using mAb to NF-L (NR4) (c). Moreover, further resolution of Hsc70 (small arrow) and NF-L (large arrow) using two-dimensional gel electrophoresis confirmed that NF-L is radiolabeled (d) and immunostained with anti-NF-L in control optic axons (e), whereas, in HM-DKO optic axons, NF-L is neither detectably labeled (f) nor immunostained (g). WT, Wild-type mice; MAP1A, microtubule-associated protein 1A.
NF-M is capable of transport without NF-L
We compared subunit protein levels in optic axons of LKO and wild-type mice after total protein extracts were subjected to SDS-PAGE, transferred to nitrocellulose, and immunoblotted with mAbs to NF-L, NF-M, NF-H, or α-internexin. Densitometric quantification of immunostained proteins showed that optic axons from LKO mice have no NF-L (Fig. 2a) but contain a substantial proportion of the NF-M (52%) measured in wild-type mouse optic nerves (Fig. 2b) and smaller proportions of the NF-H proteins (13%) (Fig. 2c). The level of α-internexin was essentially unchanged by NF-L deletion (92.5% of wild type) (Fig. 2d).

A major proportion of the normal level of NF-M protein is present in optic nerve axons of LKO mice. a-d, Immunoblots of 20 μg of total optic nerve protein extract from 3- to 4-month-old mice and equal amount of protein was loaded. Neurofilament triplet subunits and α-internexin were identified with mAbs to NF-L, NF-M, NF-H, and α-internexin (a, b, c, d), respectively. Relative levels of each protein were determined by NIH Image software. Measurements are mean ± SD from seven to eight animals for each group. WT, Wild-type mice.
To examine the transport behavior of NF-M and NF-H in the absence of NF-L, we injected [35S]methionine intravitreally into LKO and wild-type mice and analyzed the distribution of labeled Triton X-100-soluble and Triton X-100-insoluble proteins along segments of the optic pathway at 3 and 7 d after injection (Fig. 3). NF-M and NF-H proteins were exclusively Triton X-100 insoluble. Densitometry of autoradiographs revealed that the NF-M protein in optic axons of LKO animals was transported at the same rate (0.25 mm/d) as that in wild-type mice (Fig. 3c,f), which corresponded to the SCa (slow component a) rate of slow transport. The radioactivity of transported NF-H in LKO optic axons was too low to be visible on these autoradiographs (Fig. 3b,e). Because NF-M and NF-H subunits accumulate abnormally in motor neuron cell bodies of LKO mice (Williamson et al., 1998) and of neurofilament-deficient Japanese quail (Toyoshima et al., 2000) caused by a nonsense mutation in the NF-L gene (Ohara et al., 1993), we also compared in LKO and wild-type mice the levels of radiolabeled neurofilament proteins in retinas, which contain the ganglion cell perikarya that give rise to optic axons. NF-M and NF-H proteins at 3 d after intravitreal injection of [35S]methionine were exclusively Triton X-100 insoluble and, in LKO mice, were at the same levels (NF-M) or lower levels (NF-H) compared with those in wild-type mice, indicating that these two subunits did not accumulate in the retina. Western blot analysis using mAbs to NF-M and NF-H confirmed this conclusion (data not shown). It is possible that either NF-M proteins are degraded faster in the absence of NF-L in axons or that RNA translation is reduced because NF-M protein levels in the retina were the same as wild type and 52% relative to wild type in optic axons, but the levels of NF-M mRNA are unaffected by the disruption of the NF-L gene (Levavasseur et al., 1999).

NF-M protein is transported at a normal SCa rate along optic axons in the absence of NF-L. Slow axonal transport was measured as noted in the legend to Figure 1, except using wild-type (WT; a, d) and LKO (b, e) mice. c and f show transport of NF-M protein quantified by densitometry scanning of autoradiograph profiles and plotted against optic pathway segment numbers. The vertical axis indicates relative radioactivity of labeled NF-M protein. On the horizontal axis, the nerve segments are numbered consecutively from the level of the eye. Insoluble NF-M protein was transported at the same SCa rate (0.25 mm/d) in LKO mice as in wild-type mice.
NF-M is transported without NF-L and NF-H
To eliminate the possibility of a role for NF-H in the movement of NF-M in LKO mice, we generated NF-H and NF-L double knock-out (HL-DKO) mice by crossbreeding NF-H knock-out mutants (Rao et al., 1998) with LKO animals (Zhu et al., 1997). Deleting NF-H completely did not influence NF-M levels in the optic nerves of HL-DKO mice, which remained comparable with those in LKO mice and substantial (43.3 ± 3.2%) compared with those in wild-type mice (Fig. 4a). A larger decrease of NF-M levels in brain (Fig. 4b) than in spinal cord of HL-DKO mice (Fig. 4c) is likely to be attributable in part to much lower α-internexin levels in brain and higher proportions of axons in spinal cord than in brain. As in LKO mice, levels of α-internexin in HL-DKO optic nerves were unchanged (97.1 ± 9.9%) (Fig. 4d). Similarly, the translocation rate of newly synthesized Triton X-100-insoluble NF-M along optic axons in the absence of NF-L and NF-H was the same as that in wild-type mice (Fig. 5) and LKO mice (Fig. 3), with the peak rate calculated at 0.25 mm/d between 3 (Fig. 5c) and 7 d (Fig. 5f) and at 0.14 mm/d between 7 and 14 d (Fig. 5i).

NF-M protein remains at high levels in optic nerve axons of HL-DKO mice. Immunoblots of 20 μg of protein extract from 3- to 4-month-old mice and equal amounts of protein were loaded. NF-M protein and α-internexin were identified with mAbs to NF-M (a-c) and α-internexin (d), respectively. Relative levels of each protein were determined by NIH Image software. Measurements are mean ± SD from four animals for each group. NF-M levels in optic axons of HL-DKO and LKO mice (Fig. 2) do not significantly differ.

NF-M protein is transported at a normal SCa rate along optic axons independently of NF-L and NF-H. Slow axonal transport was measured as noted in the legend to Figure 3, except using wild-type (WT; a, d, g) and HL-DKO (b, e, h) mice. Insoluble NF-M protein from HL-DKO and wild-type mice was transported at the same SCa rate (c, f, i).
An attempt to generate NF-H alone mutant mice by crossbreeding NF-L knock-out with NF-M null animals was unsuccessful because mating of heterozygous NF-L+/-/NF-M+/- parents gave rise to only three genotypes: NF-L+/+/NF-M+/+ wild-type, NF-L -/-/NF-M +/+ single NF-L knock-out, and NF-L +/+/NF-M -/- single NF-M null animals. This is likely attributable to the fact that the NF-L and NF-M genes map too close to each other (∼30 kb apart) to allow segregation by breeding procedure. Thus, generation of mice expressing only NF-H will require an entirely different knock-out strategy.
NF-M transport without other triplet proteins requires α-internexin
The known ability of α-internexin to form filaments with NF-M raised the possibility that NF-M transport might be modulated by interactions with this intermediate filament protein. Indeed, we detected small numbers of intermediate-sized filaments in the optic axons of HL-DKO mice by electron microscopy. These filaments were visible in electron micrographs of axons in cross sections and in longitudinal sections (Fig. 6). Although axon caliber was significantly reduced in HL-DKO mice (data not shown), axoplasmic architecture appeared otherwise normal with the exception of a small increase in microtubule number. To determine the influence of α-internexin on NF-M transport in the absence of other triplet proteins, we generated α-internexin and NF-L double knock-out (α-IL-DKO) mice by crossbreeding α-internexin null (α-IKO) mutant (Levavasseur et al., 1999) with LKO mice (Zhu et al., 1997). As shown in Figure 7, a and b, deletion of α-internexin abolished the slow axonal transport of NF-M in the optic nerve axons. Labeled NF-M was found in the fast axonal transport phase at 5 hr after isotope injection in wild-type mice (g) but undetectable in α-IL-DKO animals (h). Western blot analyses of optic pathways from α-IL-DKO mice confirmed the absence of α-internexin and NF-L and only small amounts (<2%) of NF-M and NF-H, which may reflect adventitious attachment to other organelles or possible minute levels of protein synthesis in axons (Alvarez et al., 2000). Additional analyses of single α-IKO mice revealed that lack of α-internexin itself does not affect the transport of NF-M protein in optic axons when NF-L and NF-H are present (Fig. 7i,j). These results demonstrate that NF-M transport does not require other neurofilament triplet proteins in axons that contain another type IV neuronal intermediate filament protein.
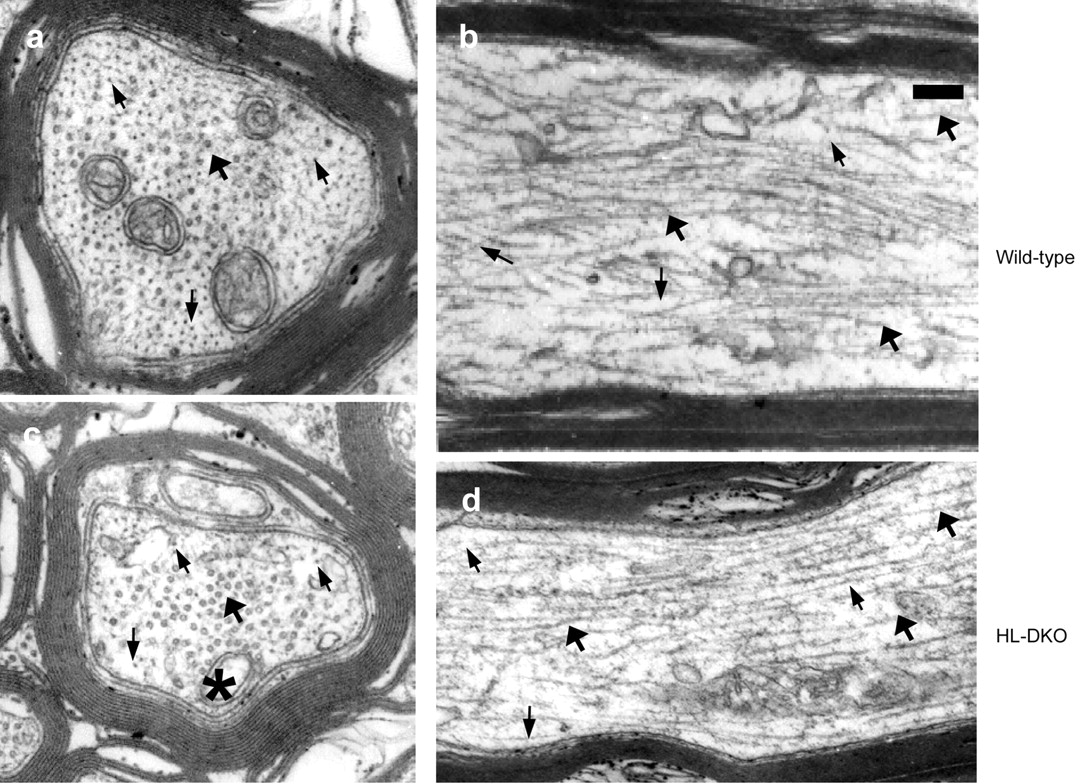
Intermediate filaments are present in small numbers in optic axons of HL-DKO mice. Electron microscopy of transverse (a, c) and longitudinal (b, d) sections of optic nerve at 2 mm from the eye in 3-month-old wild-type (a, b) and HL-DKO (c, d) mice. Intermediate filaments and microtubules are indicated by small and large arrows, respectively. Note that the number of intermediate filaments in the HL-DKO axon depicted is higher than that for the whole axon population. The asterisk in C is a mitochondrion. Scale bar, 0.25 μm.

Slow transport of NF-M protein is abolished in optic axons of α-IL-DKO mice. Autoradiographs of slow transport profiles at 3 or 7 d were generated as noted in the legend to Figure 1, except using wild-type (WT; a, i),α-IL-DKO (b), and α-IKO (j) mice. Enlargement of regions from lane 1 of these gels are shown in c, e, and k. The absence of NF-L (c, d) and α-internexin (e, f) in α-IL-DKO mice was confirmed by autoradiography and immunoblotting. Note that the absence of labeled NF-L is partially obscured by labeled Hsc70 (c). Autoradiographs of fast transport profiles at 5 hr were also generated as noted in the legend to Figure 1, except using wild-type (g) and α-IL-DKO (h) mice, and only the first two lanes of NF-M regions were shown. A small portion of NF-M was fast transported in wild-type mice but undetectable in α-IL-DKO animals. Deletion of α-internexin alone (α-IKO) did not affect the slow transport of NF-M and other slowly transported proteins (i, j). The α-internexin migrates on gels just above a 57 kDa SCb protein (e) (indicated as X). The absence of α-internexin (k, l) in α-IKO mice was confirmed by autoradiography and immunoblotting. MAP1A, Microtubule-associated protein 1A.
Discussion
Despite experimental efforts over two decades, the form for neurofilament protein transport has remained a controversial issue. In this report, we showed that NF-L alone is incapable of efficient transport in optic axons, confirming a previous finding in peripheral axons (Jacomy et al., 1999). In addition, however, we demonstrated for the first time that a major proportion of the normal level of endogenous NF-M can be transported into axons without other triplet proteins. Thus, NF-M transport in vivo does not necessarily require the other neurofilament triplet subunits or assembly into neurofilaments as they are currently defined.
Our additional finding that deletion of α-internexin abolishes the slow transport of NF-M in optic axons of LKO mice indicates that NF-M monomer is not efficiently transported on its own. Thus, we show, for the first time, that efficient neurofilament protein transport in vivo requires the formation of at least hetero-oligomeric complexes and that NF-M may partner with NF-L, α-internexin, or both to achieve slow transport in vivo. The additional observations that a small number of intermediate filaments are present in HL-DKO optic axons (Fig. 6) but completely disappear in α-IL-DKO mice (Levavasseur et al., 1999) along with the disappearance of NF-M transport suggest that α-internexin and NF-M subunits can form intermediate filaments. Therefore, our results reveal a novel ability of neurofilament subunits to form intermediate filaments in vivo with a composition distinct from that of conventional neurofilaments. These findings and the additional observation that α-internexin and NF-M can colocalize on the same filament in axons of wild-type mice (our unpublished observations) also imply that neurofilaments are heterogeneous in composition and may contain α-internexin. This conclusion is consistent with reports that α-internexin coassembles with each of the neurofilament triplet proteins in vitro (Ching and Liem, 1993, 1998), colocalizes with triplet proteins in CNS axons (Kaplan et al., 1990), and accumulates abnormally with the triplet proteins when α-internexin is overexpressed in transgenic mice (Ching et al., 1999). The number of intermediate filaments in optic axons of HL-DKO mice is <10% of the number of neurofilaments in wild-type mouse axons (A. Yuan and R. A. Nixon, unpublished observations) and unlikely to account for the high proportion of NF-M that is still transported in HL-DKO mice. In these axons, more likely the transported NF-M in LKO or HL-DKO mice is in the form of both hetero-oligomers and heterpolymers. However, we cannot exclude the possibility that all of the NF-M transported in HL-DKO mice is represented by the remaining filaments. Because the gene deletion paradigm might promote a compensatory response that is not normally present in wild-type animals, additional studies of these filaments are needed to establish what proportion of NF-M is associated with α-internexin under normal conditions.
Terada et al. (1996) provided initial evidence for movement of a modified rat NF-M protein introduced by viral vector delivery of the gene into neurons of transgenic mice that lack the majority of their axonal neurofilaments (Eyer and Peterson, 1994). Because NF-M protein cannot assemble into homopolymers, detection of the transfected protein at substantial distance down the axon was interpreted as support for subunit transport. This interpretation, however, was challenged for not distinguishing between active transport and diffusion (Baas and Brown, 1997), which can occur over considerable distances along axons within hours of introducing a fluorescent molecule into the cell body (Popov and Poo, 1992). Our findings are consistent with the observations of Terada et al. that NF-M can move without the other triplet proteins but demonstrate unequivocally that the efficient slow transport of NF-M in the absence of NF-L and NF-H requires α-internexin.
NF-M subunit exists in all vertebrates, and all other neurofilament proteins are believed to have originated by gene duplication from an ancestral NF-M-like protein. The most primitive living vertebrate lamprey has only a single NF-M-like subunit (NF-180) (Hall et al., 2000), but more recent studies suggest that this subunit may need another element for filament assembly (Zhang et al., 2002). In tiger salamander olfactory axons, NF-M is abundant, but NF-L and NF-H subunits are present at trace levels, resulting in rare neurofilament formation (Burton and Wentz, 1992). In developing spinal cord neurons of Xenopus laevis and mouse myenteric neurons, NF-M and α-internexin (or α-internexin-like Xenopus protein) emerge simultaneously before the appearance of NF-L and NF-H subunits, indicating a close relationship between NF-M and α-internexin (Faussone-Pellegrini et al., 1999; Undamatla and Szaro, 2001). The NF-M subunit has also been shown to interact with the D1 dopamine receptor to regulate cell surface expression and desensitization (Kim et al., 2002). It is possible, therefore, that the unique transport behavior of NF-M, among the triplet proteins, may reflect a more complex role for NF-M in neuronal function, consistent with its extensive regulation by phosphorylation (Sihag and Nixon, 1990; Sihag et al., 1999).
In conclusion, we showed that neurofilament proteins require at least hetero-oligomer formation for efficient axonal transport. Previously, in vitro immunoprecipitation studies demonstrated binding of kinesin to NF-M and NF-H but not NF-L (Yabe et al., 2000), suggesting a role for the high molecular weight subunits in neurofilament transport. Our findings define minimal molecular requirements for efficient slow transport of neurofilament proteins and suggest that, among the triplet proteins, the NF-M protein is likely to be the critical subunit to interact with the putative transport motor.
Footnotes
This work was supported by National Institutes on Aging Grants AG05604 and P30-DK19525 (R.A.N.). We express deep appreciation to Hayley McAuliff for her help with manuscript preparation.
Correspondence should be addressed to Dr. Aidong Yuan, Center for Dementia Research, Nathan Kline Institute, New York University School of Medicine, 140 Old Orangeburg Road, Orangeburg, NY 10962. E-mail: yuan{at}nki.rfmh.org.
Copyright © 2003 Society for Neuroscience 0270-6474/03/239452-07$15.00/0





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}