

Abstract
The members of the pentameric ligand-gated receptor channel family are involved in information transfer in synapses and the neuromuscular junction. They often contain several copies of the same subunit isoform. Here, we present a method to functionally dissect the role of individual subunits that occur in multiple copies in these receptors.
Opening of the inherent chloride channel in the GABAA receptor is achieved through the binding of two agonist molecules; however, it has been difficult to obtain information on the contribution of the two individual binding sites. The sites are both located at β(+)/α(-) subunit interfaces, suggesting similar properties. One pair of subunits is flanked by γ and β (site 1) and the other by α and γ (site 2), the different environment possibly affecting the binding sites. Here, we used concatenated subunits and two point mutations of amino acid residues, each in α and β subunits, both located in the agonist binding pocket, to investigate the properties of these two sites. The sites were individually mutated, and consequences of these mutations on GABA and muscimol-induced channel opening and its competitive inhibition by bicuculline were studied. A model predicts that opening also occurs for receptors occupied with a single agonist molecule but is promoted approximately 60-fold in those occupied by two agonists and that site 2 has an approximately threefold higher affinity for GABA than site 1, whereas muscimol and bicuculline show some preference for site 1.
Introduction
GABAA receptors are the major inhibitory neuronal ion channels in the mammalian brain. They belong to the family of ligand-gated ion channels that includes nicotinic acetylcholine, glycine, and serotonin type 3 receptors. Biochemical purification of a bovine GABAA receptor (Sigel et al., 1983) was followed by cloning, which led to the identification of 18 different subunit isoforms (Macdonald and Olsen, 1994; Rabow et al., 1995; Davies et al., 1997; Hedblom and Kirkness, 1997; Whiting et al., 1997, 1999; Barnard et al., 1998). Most GABAA receptors are thought to be pentameric assemblies containing α, β, and γ or α, β, and δ subunits (McKernan and Whiting, 1996). The five subunits are arranged pseudosymmetrically around a central Cl--selective channel (Macdonald and Olsen, 1994). The most likely stoichiometry is two α subunits, two β subunits, and one γ or δ subunit (Backus et al., 1993; Chang et al., 1996; Tretter et al., 1997; Farrar et al., 1999; Baumann et al., 2001). The major receptor isoform of the GABAA receptor in the brain probably consists of α1, β2, and γ2 subunits (Laurie et al., 1992; Benke et al., 1994; Macdonald and Olsen, 1994; Rabow et al., 1995; McKernan and Whiting, 1996). The inferred arrangement of subunits around the channel pore is γβαβα counterclockwise when viewed from the synaptic cleft (Baumann et al., 2002).
The functional GABA binding site in GABAA receptors is located at intersubunit contacts between α and β subunits (Sigel et al., 1992; Amin and Weiss, 1993; Smith and Olsen, 1994; Westh-Hansen et al., 1997; Boileau et al., 1999), and homologous amino acid residues of α and γ subunits form the benzodiazepine binding pocket (Wieland et al., 1992; Amin et al., 1997; Buhr and Sigel, 1997; Buhr et al., 1997a, b; Sigel and Buhr, 1997; Teissere and Czajkowski, 2001).
Here, we address the question of whether or not the two agonist sites on the receptor have identical properties. To our knowledge, this question has never been addressed for GABAA receptors. In the homologous nicotinic acetylcholine receptor, there is indirect evidence both for (Edelstein et al., 1997) and against (Neubig and Cohen, 1979) different properties of the agonist at the two sites. Some competitive antagonists (e.g., d-tubocurarine) differ widely in their affinity for the two sites (Martinez et al., 2000).
In the present study, we used two mutations, α1F65L located on the minus side and β2Y205S located on the plus side, to dissect the contribution of each of the two individual functional GABA binding sites. Concatenated subunits of the GABAA receptor were used for a forced assembly. The point mutations were introduced in the α1 and β2 subunits flanked by the γ subunit and a β subunit (site 1), by an α subunit and the γ subunit (site 2), or both. Mutation of the two sites leads to subtly different consequences.
Materials and Methods
Construction of tandem and triple subunit cDNAs. The tandem constructs, β2-α1 and γ2-β2, and triple constructs, γ2-β2-α1 and α1-β2-α1, were made as described previously (Baumann et al., 2001, 2002). Site-directed mutagenesis of α1F65 to L, β2Y62 to L, or β2Y205 to S were done in the tandem constructs β2-α1 or γ2-β2 and α1-β2 using the QuickChange mutagenesis kit (Stratagene, Amsterdam, The Netherlands). Subsequently, the triple constructs γ2-β2-α1F65L, γ2-β2Y62L-α1, and α1-β2Y205S-α1 were made similar as described by Baumann et al. (2001, 2002).
Expression of linked constructs in Xenopus oocytes. Capped cRNAs were synthesized (Ambion, Lugano, Switzerland) from linearized pCMV vectors containing different tandem or triple constructs, respectively, and from the vector pVA2580 (Kuhn and Greeff, 1999) encoding a neuronal voltage-gated sodium channel. A poly-A tail of ∼400 residues was added to each transcript using yeast poly-A polymerase (United States Biochemicals, Dūbendorf, Switzerland). The concentration of the cRNA was quantified on a formaldehyde gel using Radiant Red stain (Bio-Rad, Reinach, Switzerland) for visualization of the RNA and known concentrations of RNA ladder (Invitrogen, Basel, Switzerland) as standard on the same gel. cRNA combinations were precipitated in ethanol/isoamylalcohol, 19:1, and stored at -20°C. For injection, the alcohol was removed, and the cRNAs were dissolved in water. Isolation of oocytes from the frogs, culturing of the oocytes, injection of cRNA, and defolliculation were done as described previously (Sigel, 1987). Oocytes were injected with 50 nl of the cRNA solution. For cRNA combinations of the triple constructs with the tandem construct, ratios of 10:10 nm were investigated. The combination of wild-type α1, β2, and γ2 subunits was expressed at a ratio of 10:10:50 nm (Boileau et al., 2002). To allow standardization of expressed GABA currents, cRNA coding for the voltage-gated sodium channel was always added to a concentration of 40 nm. The injected oocytes were incubated in modified Barth's solution [10 mm HEPES, pH 7.5, 88 mm NaCl, 1 mm KCl, 2.4 mm NaHCO3, 0.82 mm MgSO4, 0.34 mm Ca(NO3)2, 0.41 mm CaCl2, 100 U/ml of penicillin, 100 μg/ml of streptomycin] at 18°C for 2 d before the measurements.
Two-electrode voltage-clamp measurements. All measurements were done in medium containing (in mm): 90 NaCl, 1 MgCl2, 1 KCl, 1 CaCl2, and 5 HEPES, pH 7.4, at a holding potential of -80 mV. For the determination of maximal current amplitudes, 10 mm GABA (Fluka, Buchs, Switzerland) was applied for 20 sec. The perfusion solution (6 ml/min) was applied through a glass capillary with an inner diameter of 1.35 mm, the mouth of which was placed ∼0.4 mm from the surface of the oocyte. The rate of solution change under our conditions has been estimated 70% within <0.5 sec (Sigel et al., 1990). Voltage-dependent sodium currents were determined by a potential jump from a holding potential of -100 to -15 mV. Functional GABAA receptor expression was determined by application of 10 mm GABA. The elicited current amplitude was normalized to the Na current measured. GABA-evoked currents (at 8-12% of the maximal current amplitude) were inhibited by varying concentrations of bicuculline methiodide (Sigma/RBI, Buchs, Switzerland). Concentration-response curves for GABA were fitted with the equation I(c) = Imax/[1 + (EC50/c)n], where c is the concentration of GABA, EC50 is the concentration of GABA eliciting half-maximal current amplitude, Imax is the maximal current amplitude, I is the current amplitude, and n is the Hill coefficient. Inhibition curves for bicuculline were fitted with the equation I(c) = I(0)/[1 + (IC50/c)n], where I(0) is the control current in the absence of bicuculline standardized to 100%, I(c) is the relative current amplitude, c is the concentration of bicuculline, IC50 is the concentration of bicuculline causing 50% inhibition of the current, and n is the Hill coefficient. Data are given as mean ± SEM (number of experiments, number of batches of oocytes). Relative current stimulation by diazepam (DZ) was determined at a GABA concentration evoking 2-5% of the maximal current amplitude in combination with varying concentrations of DZ (Roche Pharma, Reinach, Switzerland) and expressed as [(I(GABA + DZ)/I(GABA)) - 1] · 100.
Western blotting. Oocytes were homogenized in lysis buffer (10 mm HEPES, pH 8.0, 100 mm NaCl, 10 mm EDTA, 1% Triton X-100, Pepstatin, Leustatin, Antipain, and PMSF, each at 5 μg/ml) using a Teflon glass homogenizer. The homogenate was incubated on ice for 15 min and centrifuged at 15,000 × g for 15 min at 4°C. The supernatant was extracted with chloroform-methanol and subjected to SDS-PAGE (Laemmli, 1970). Proteins were transferred to nitrocellulose membranes (HybondECL; Amersham Pharmacia, Dūbendorf, Switzerland) according to Towbin et al. (1979) and decorated with the monoclonal antibodies bd24 and bd17 (Häring et al., 1985; Ewert et al., 1990), which recognize the N-terminal of the α1 and β2 subunits of the GABAA receptor, respectively. Bands were detected using the ECL system (Amersham Pharmacia).
Model. Because some of the parameters were in common for linked and mutated receptors, the averaged standardized data including SDs were fitted simultaneously with using a Levenberg-Marquardt algorithm (ProFit 5.1 software; ProFit, Zurich, Switzerland) in combination with the following equations: loose subunits, I(c) = [100 · (1 + L)]/(1/[1 + (L1 · K1)/(L2 · K2) + (L1 · c)/(L · K2)]) · [1 + (K1 · L1)/c + L1 + (L1 · K1)/K2 + (c · L1)/(K2) + (L1 · K1)/(L2 · K2) + (c · L1)/(K2 · L)]; receptors linked αβα/γβ, I(c) = [100 · (1 + fA · L)]/(1/[1 + (L1 · K1)/(L2 · K2) + (fa · L1 · c)/(fA · L · K2)]) · [1 + (K1 · fa · L1)/c + fa · L1 + (fa · L1 · K1)/K2 + (c · fa · L1)/(K2) + (L1 · K1)/(L2 · K2) + (c · fa · L1)/(K2 · fA · L)]; receptors linked γβα/βα, I(c) = [100 · (1 + fG · L)]/(1/[1 + (L1 · K1)/(L2 · K2) + (fg · L1 · c)/(fG · L · K2)]) · [1 + (K1 · fg · L1)/c + fg · L1 + (fg · L1 · K1)/K2 + (c · fg · L1)/(K2) + (L1 · K1)/(L2 · K2) + (c · fg · L1)/(K2 · fG · L)]; αβα/γβ receptors mutated in site 1 and an analogous equation for the mutation in residue α165 in site 1, I(c) = [100 · (1 + fA · L)]/(1/[1 + (L1 · K1205)/(L2 · ta205 · K2) + (fa · L1 · c)/(fA · L · ta205 · K2)]) · [1 + (K1205 · fa · L1)/c + fa · L1 + (fa · L1 · K1205)/(ta205 · K2) + (c · fa · L1)/(ta205 · K2) + (L1 · K1205)/(L2 · ta205 · K2) + (fa · L1 · c)/(fA · L · ta205 · K2)]; αβα/γβ receptors mutated in site 2 and an analogous equation for the mutation in residue α165 in site 2, I(c) = [100 · (1 + fA · L)]/(1/[1 + (L1 · ta205 · K1)/(L2 · K2205) + (fa · L1 · c)/(fA · L · K2205)]) · [1 + (ta205 · K1 · fa · L1)/c + fa · L1 + (fa · L1 · ta205 · K1)/K2205 + (c · fa · L1)/(K2205) + (L1 · ta205 · K1)/(L2 · K2205) + (fa · L1 · c)/(fA · L · K2205)]; and γβα/βα receptors mutated both sites, I(c) = [100 · (1 + fG · L)]/(1/[1 + (L1 · K165)/(L2 · K265) + (fg · L1 · c)/(fG · L · ta65 · K265)]) · [1 + (ta65 · K165 · fg · L1)/c + fg · L1 + (fg · L1 · K165)/K265 + (c · fg · L1)/(t65 · K265) + (L1 · K165)/(L2 · K265) + (c · fg · L1)/(ta65 · K265 · fG · L)]); I is the current, K1 and K2 are dissociation constants for agonists of the unmuted and K165, K265, K1205, and K2205 of the mutated receptors illustrated in Figure 5, L describes the isomerization between the closed (ARA) and open (ARA*) channel [L = (ARA)/(ARA*)], L1 and L2 describe the isomerization between closed receptors occupied by a single agonist molecule (AR) or (RA) and the corresponding open channels (AR*) and (RA*), respectively, c is the concentration of GABA or muscimol, b is the concentration of bicuculline or SR95531, fG and fA, fg, and fa are coefficients modulating L, L1, and L2, respectively (gating of the channel), as a consequence of subunit linkage, and ta65 and ta205 coefficients describe the allosteric effect of the mutation in one site on the dissociation constant for GABA of the second site.

A, Bicuculline inhibition curves of γβα/βα (•), γβα65/βα65 (○), γβα65/βα (▪), and γβα/βα65 (□) receptors. Bicuculline was applied in increasing concentrations together with a GABA concentration eliciting 9-11% of the maximal current amplitude. B, Bicuculline inhibition curves of γβα/βα (•), αβ205α/γβ (▴), and αβα/γβ205 (▾) receptors. Mean values with SEM from four to five oocytes from two batches for each subunit combination are shown. Individual curves were normalized to the current found in the absence of bicuculline and were subsequently averaged.
Inhibition was fitted with the equation I(c) = Imax/(1/[1 + (L1 · K1)/(L2 · K2) + (L1 · c)/(L · K2)]) · [1 + (K1 · L1)/c + L1 + (L1 · K1)/K2 + (c · L1)/(K2) + (L1 · K1)/(L2 · K2) + (c · L1)/(K2 · L)] + [(b · K1 · L1)/c] · [1/K1i + 1/K2i + b/(K1i · K2i)] for loose subunits, I(c) = Imax/[(1/[1 + (L1 · K1205)/(L2 · ta205 · K2) + (fa · L1 · c)/(fA · L · ta205 ·K2)]) · [1 + (K1205 · fa · L1)/c + fa · L1 + (fa · L1 · K1205)/(ta205 · K2) + (c · fa · L1)/(ta205 · K2) + (L1 · K1205)/(L2 · ta205 · K2) + (fa · L1 · c)/(fA · L · ta205 · K2)] + [(b · K1 · fa · L1)/c] · [1/(m · K1I) + 1/(tb205 · K2I) + b/(m · K1i · tb205 · K2i)])] for αβα/γβ receptors mutated in site 1, and analogously for the other receptors. K1i and K2i are dissociation constants for antagonists, m is the ratio between dissociation constants of mutated divided by that of unmutated receptors, and tb65 and tb205 coefficients describe the allosteric effect of the mutation in one site on the dissociation constant for bicuculline of the second site.
Results
Mutations in the agonist binding site
We made use of the two point mutations, α1F65L and β2Y205S, to alter agonist binding sites in recombinant α1β2γ2 GABAA receptors. The first mutation has been shown previously to decrease the apparent affinity of GABA for channel gating and the apparent affinity for the competitive antagonist bicuculline (Sigel et al., 1992). The mutation β2Y205S has been reported to compromise GABA binding without affecting channel opening induced by pentobarbital (Amin and Weiss, 1993). It is now known that a receptor pentamer contains two α and two β subunits. Therefore, in each case, both subunits and thereby both GABA sites were affected by the point mutations in the previous studies (Fig. 1, top). We have shown previously that it is feasible to covalently link α, β, and γ subunits of the GABAA receptor while retaining full receptor function (Baumann et al., 2001, 2002). This allows the introduction of a point mutation in one individual α1 or β2 subunit exclusively. Thus, the concatenated subunits γ2-β2-α1 (γβα), β2-α1 (βα), γ2-β2-α1F65L (γβα65), and β2-α1F65L (βα65) were prepared and functionally expressed in Xenopus oocytes as a channel containing no mutation γβα/βα, one mutation γβα65/βα and γβα/βα65, or two mutations γβα65/βα65 (Fig. 1). γ2-β2Y62L-α1 (γβ62α) and β2Y62L-α1 (β62α) were also prepared for control purposes. To study a second contact point of agonists with the receptor and a differently concatenated channel, we similarly prepared γ2-β2 (γβ), γ2-β2Y205S (γβ205), α1-β2-α1 (αβα), and α1-β2Y205S-α1 (αβ205α). αβα/γβ, αβ205α/γβ205, αβ205α/γβ, and αβα/γβ205 (Fig. 1) were expressed in Xenopus oocytes and functionally characterized.

Mutations in α or β subunits of the GABAA receptor. If a point mutation is introduced in an α or β subunit, two mutations are always present in an altered receptor (top row). The availability of concatenated subunits allows construction of linked receptors containing only one mutation (bottom row). This single mutated α1 subunit can be placed between either two β subunits or a β and γ subunit. This way, the point mutation α1F65L has been investigated. Similarly, the mutation β2Y205S subunit has been investigated in a differently concatenated channel.
Nonlinked receptors and linked receptors
Linked subunits might be proteolysed during expression. If the linker region(s) specifically would be affected, nonlinked subunits could be produced from dual constructs and nonlinked subunits and dual subunit constructs could be produced from triple constructs, all products carrying part of the corresponding linkers. Such an event would complicate interpretation of the results in the event that the newly liberated subunits retain the ability to reassemble to form functional channels. Because the assembly of multi-subunit proteins is taking place in an early phase after translation in the endoplasmic reticulum, this hypothetical process would have to take place there. Figure 2A shows a Western blot analysis of noninjected oocytes (lane 1), oocytes expressing loose subunits α/β/γ (lane 3), or the combination of concatenated subunits αβα/γβ (lane 2). All lanes were decorated with the α1-specific antibody bd24. As expected for the case of absent proteolysis, we detected no band in lane 1, a single specific band at ∼50 kDa in lane 3, and a single specific band at ∼170 kDa in lane 2. Different exposures allowed an estimate of the detection limit, which was ∼2% for a single proteolyzed fragment. Thus, our results obtained with αβα/γβ and its mutational variants (see below) cannot be explained by proteolysis and subsequent reassembly. Figure 2B shows a similar analysis for oocytes injected with γβα/βα and decorated with bd17, which recognizes free N-termini of β subunits. In this case, a stronger nonspecific reaction occurred. However, the band corresponding to βα is clearly seen at ∼110 kDa. A similar analysis using antibodies directed against loops of α and β and the N-terminal of γ subunits failed because of small amounts of antigens formed in the Xenopus oocyte that were not detected with the corresponding antibodies (data not shown).

Western blot analysis. A, Resistance to proteolysis of the fusion protein αβα. Lanes 1-3 were decorated with bd24, reacting with free termini of α subunits. Lane 1, Noninjected oocytes (n.i.). No signal was detected in noninjected oocytes. Lane 2, Oocytes injected with αβα/γβ. The αβα triple construct migrates at 170 kDa. No specific signal was detected at ∼50 or 110 kDa, the size of a monomeric α1 subunit or dimeric subunit carrying α1 at the N-terminus, as expected as a consequence of proteolysis in one of the linker regions. The absence of specific signals in other areas indicates that no N-terminal breakdown product of this triple subunit construct larger than 21 kDa is formed. Lane 3, Oocytes injected with α/β/γ. The two loose α1 subunits from the wild-type α1β2γ2 receptor migrate at ∼50 kDa. B, Lanes 1-3 were decorated with bd17 and reacted with free termini of β subunits. Lane 1, Noninjected oocytes. Lane 2, Oocytes injected with γβα/βα. The βα dual construct migrates at ∼110 kDa. No specific signal was detected at 53 kDa, the size of a monomeric β2 subunit, as expected during proteolysis in the linker region. Lane 3, Oocytes injected with α/β/γ. The two loose β2 subunits from the wild-type α1β2γ2 receptor migrate at ∼53 kDa.
Apart from these observations, we believe it for several reasons unlikely that proteolysis of subunits is followed by assembly of the fragments to form functional channels. First, we did not obtain functional channels when we chose the linker too short. Second, we tried to functionally express a huge variety of combinations of dual and triple constructs without succeeding unless a channel γβαβα could be formed directly (Baumann et al., 2001, 2002). If proteolysis would lead to functional channels, many of the non-γβαβα channels should have resulted in functional expression. Specifically, we expressed βα alone (Baumann et al., 2001) and γβα in combination with α or β subunits (Baumann et al., 2002) and observed very little, if any, current. Third, the hypothetical proteolysis leading to different functional channels than envisaged would be predicted to lead to a channel mixture. We never observed evidence for a two or more phasic dose-response curve for nonmutated channels.
At a functional level, the effect of subunit concatenation was first studied. Thus, the functional properties of nonlinked receptors, α/β/γ, and linked receptors, αβα/γβ and γβα/βα, were compared. Concentration-response curves for GABA and muscimol, as agonists, and inhibition of currents elicited by GABA by bicuculline were measured for all three types of receptors. Results are summarized in Table 1. Subunit concatenation resulted in a 2- to 3.5-fold loss in agonist sensitivity. In contrast, subunit concatenation did not significantly affect the IC50 for bicuculline in αβα/γβ receptors and only to a very small extent in γβα/βα receptors.
Agonist and antagonist properties of wild-type, concatenated, and mutated receptors
Effect of the mutation α1F65L on agonist concentration-response properties for GABA and muscimol
Figure 3A summarizes average concentration-reponse curves for wild-type γβα/βα and mutant γβα65/βα65 channels and also shows curves derived from channels harboring a single mutation γβα65/βα and γβα/βα65. The curves obtained from the channels containing one mutation displayed properties more similar to channels with two mutated sites. Data are summarized in Table 1.

A, GABA concentration-response curves of γβα/βα (•), γβα65/βα65 (○), γβα65/βα (▪), and γβα/βα65 (□) receptors. B, Muscimol concentration-response curves of γβα/βα (•) and γβα65/βα65 (○) receptors. Mean values with SEM from four to five oocytes from two batches for each subunit combination are shown. Individual curves were first normalized to the observed maximal current amplitude and subsequently averaged.
GABA concentration-response curves were also obtained for mutant γβ62α/β62α channels (data are given in Table 1). β62 is the homologous residue in the β2 subunit to α65 in the α1 subunit and has been proposed to contribute to high-affinity binding of agonists (Newell et al., 2000). The mutation introduced in both β2 subunits had only a very small effect on EC50. Each of the channels harboring a single mutated subunit was studied only twice. As expected, an intermediate EC50 resulted (data not shown).
The concentration-response curves for the agonist muscimol was also performed with wild-type γβα/βα and mutant γβα65/βα65 channels (Fig. 3B). Quantitative data are given in Table 1. Although double mutation caused a 12-fold shift in the EC50 for GABA, it amounted to only fourfold for muscimol.
Effect of the mutation β2Y205S on agonist concentration-response properties for GABA and muscimol
Figure 4A illustrates concentration-response curves obtained from wild-type αβα/γβ channels with GABA and muscimol, respectively (values are given in Table 1). The channel containing two mutations did not result in any detectable current for GABA concentrations up to 10 mm. At 100 mm, the elicited current amounted to 0.1% of the maximal current amplitude of wild-type αβα/γβ channels. Expression of all channels was verified by determining the amplitude of the coexpessed voltage-gated Na channel and using direct activation of the GABA channel by 1 mm pentobarbital. The observed current amplitudes elicited by pentobarbital were in the range of 200-1200 nA for the wild-type channel (10 oocytes), 300-2800 nA for the αβα/γβ205 channel (13 oocytes), 70-400 nA for αβ205α/γβ channels (14 oocytes), and 2-60 nA for αβ205α/γβ205 channels (15 oocytes). The reasons for the small currents in the latter case is not known; however, because the corresponding oocytes expressed microampere-sized Na currents, it cannot be explained by either failure of injection or presence of RNase activity in the injection solution.
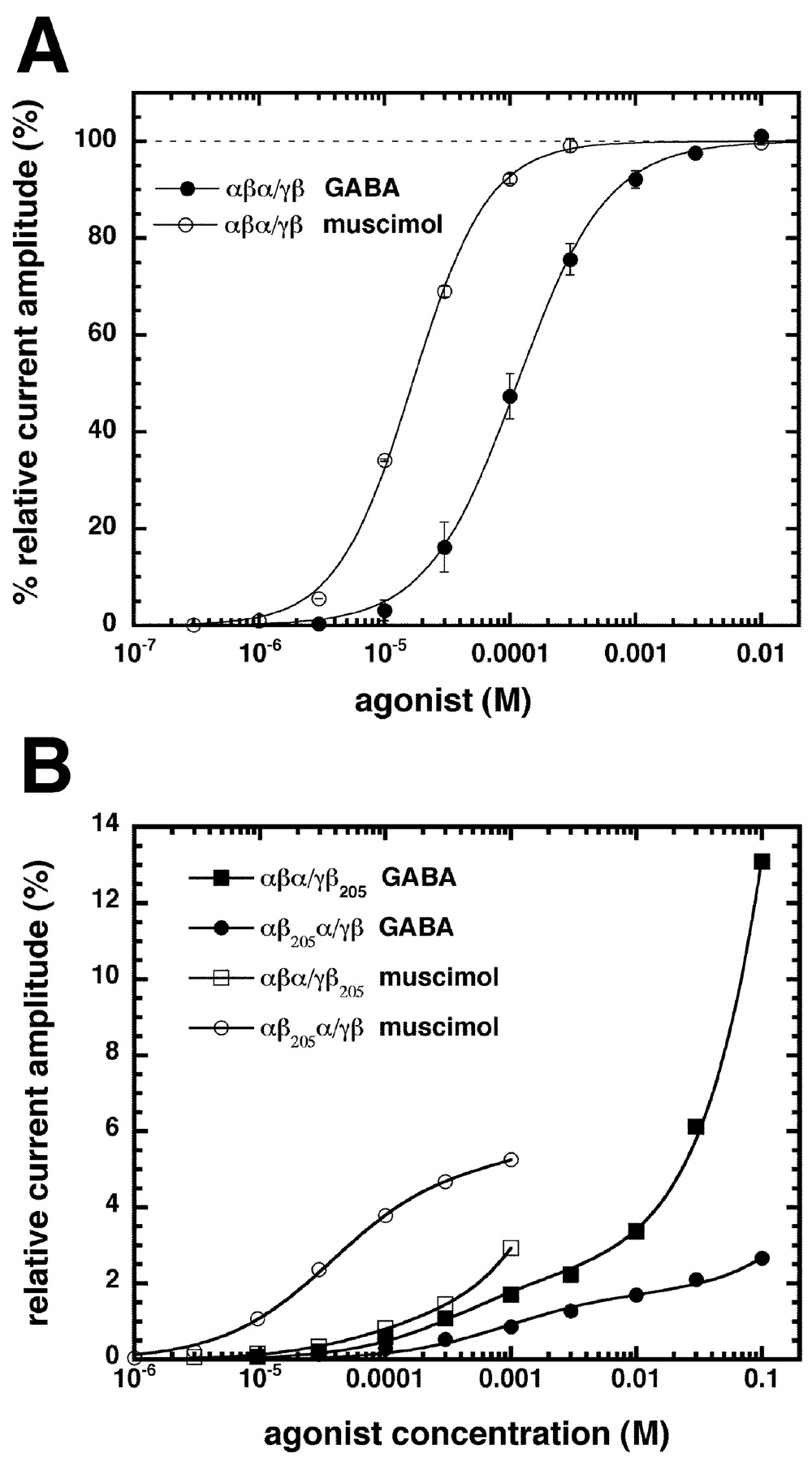
A, GABA (•) and muscimol (○) concentration-response curves of αβα/γβ receptors. B, GABA (closed symbols) and muscimol (open symbols) concentration-response curves of αβα/γβ205 (square symbols) and αβ205α/γβ (round symbols) receptors. Mean values with SEM from four to five oocytes from two batches for each subunit combination are shown. For receptors containing no mutation, individual curves were first normalized to the observed maximal current amplitude and subsequently averaged. For mutated receptors, amplitudes were normalized to the average maximal response elicited by GABA in αβα/γβ receptors.
The mutation β2Y205S destroys the agonist binding site and theoretically should uncover the properties of the second site. Thus, in αβ205α/γβ mutant receptors, only site 1 should be visible and in αβα/γβ205 mutant receptors, only site 2 should be visible. The maximal current amplitudes of αβ205α/γβ and αβα/γβ205 mutant receptors could not be determined because the affinity of the second site is in the molar range. For evaluation of data, it was assumed that this parameter was the same as for αβα/γβ channels. Figure 4B summarizes the dose-response curves in response to the agonists GABA and muscimol on αβ205α/γβ and αβα/γβ205 mutant receptors after normalization to the maximal current amplitude of unmutated αβα/γβ receptors. All average curves were fitted with a two component logistic equation (Fig. 4B, solid lines). Data are summarized in Table 1. The absolute amplitudes should be judged with care because they depend on the expression rate. Our observations are in line with a preference of site 2 for GABA and a preference of site 1 for muscimol. It was interesting to see whether the assumption that αβ205α/γβ and αβα/γβ205 mutant receptors display the same maximal current amplitudes as αβα/γβ receptors affected our basic findings. Therefore, three different alternative possibilities were tested. First, it was assumed that the maximal currents by both mutant receptors were only one-fifth of the standardized currents. Second, it was assumed that they were both five times the standardized currents and, third, that αβα/γβ205 and αβ205α/γβ mutant receptors had a twofold, respectively one-third, maximal current amplitude of wild-type receptors, as suggested by the currents elicited by 1 mm pentobarbital. Our basic conclusions were not affected in any of the three cases.
Inhibition by bicuculline
The concentration-dependent inhibition of GABA-induced currents by the competitive antagonist bicuculline was also studied. Opening of the GABA receptor channel was standardized by performing experiments approximately at the respective EC10 (EC8.9-EC11.2), except for the receptors containing a β2Y205S mutation. For the mutant γβα65/βα65 channels, as compared with wild-type γβα/βα channels, IC50 for the competitive antagonist bicuculline is shifted to 56-fold higher concentrations. Figure 5A summarizes these experiments and, in addition, shows average curves obtained with channels harboring a single mutation, γβα65/βα and γβα/βα65, which were shifted eightfold and fourfold, respectively. Quantitative data are summarized in Table 1.
Bicuculline concentration-inhibition curves were also obtained from mutant γβ62α/β62α channels (Table 1). The mutation introduced in both β2 subunits had no significant effect. Therefore, channels harboring a single mutated subunit were not studied.
Figure 5B summarizes bicuculline inhibition curves for wild-type αβα/γβ and single mutant αβ205α/γβ and αβα/γβ205 channels, respectively. Both curves of mutant receptors would be expected to superimpose with those of concatenated wild-type receptors, unless the effects of mutations were allosterically communicated to the other site. Because the shift amounts to approximately sevenfold in both cases, this seems to be the case. Quantitative data are summarized in Table 1. Inhibition for αβα/γβ205 and αβ205α/γβ receptors was measured at a GABA concentration of 100 and 300 μm, respectively.
Inhibition by SR95531
Inhibition by the competitive inhibitor SR95531 was determined for γβα/βα, γβα65/βα, γβα/βα65, γβα65/βα65, αβα/γβ, αβ205α/γβ, and αβα/γβ205 under conditions described for bicuculline. In all cases, a qualitatively similar behavior was found. The respective IC50 were ∼10-fold lower than with bicuculline.
Diazepam responsiveness
All receptors, γβα/βα, γβα65/βα, γβα/βα65, γβα65/βα65, γβ62α/βα, γβα/β62α, and γβ62α/β62α, were tested for stimulation of the current amplitude by 1 μm diazepam at an EC5 for GABA. In all cases, the current was stimulated to 240-330% of the control amplitude (data not shown).
Model
Different models were used to analyze the experimental data. Figure 6 shows the model that was in best agreement with our findings and then used for the fit of the data. The model incorporates two binding sites for agonists and competitive antagonists, site 1 and site 2. At least a single site has to be occupied by an agonist to produce an open state AR*, RA*, or ARA*. Concentration-response curves, including those with nonlinked subunits, were averaged and standardized. We initially tried to fit individual curves with the equations given in Materials and Methods containing variables describing binding (K1, K2) as well as gating (L, L1, L2) phenomena. However, many acceptable fits were obtained. Because different curves share at least some variables, a combined Levenberg-Marquardt fit was applied to the activation curve data collected with GABA as an agonist in receptors composed of nonlinked subunits, containing linked subunits (two receptor types), mutated in site 1 (two receptor types), mutated in site 2 (two receptor types), and containing the α1F65L mutation in both sites. Combined fitting led to a single solution. Another combined fit was applied to the data on bicuculline inhibition. Here, the fitted parameters of the activation curves were used. Results of the fits are given in the legend for Figure 6. Fitting suggested that concatenation of subunits modified gating of the channel, favoring closed channels by a factor of 4-31 (fa, fA, fg, fG). For unmutated channels, transition into the open state is ∼60 times less efficient when occupied by a single agonist molecule as compared with those occupied by two agonist molecules. Fitting also predicted that site 2 has an approximately threefold higher affinity for GABA than site 1. Data presented in Figure 4B indicate a preference of site 1 for muscimol. Occupation of a single site by bicuculline keeps the channel in a closed state. Site 1 has a slight preference for bicuculline. Simulated data were obtained with the above fit results for the gating of the channel by GABA and its inhibition by bicuculline for the mutation in the α1 subunit α1F65L (Fig. 7A,B) and the mutation in the β2 subunit β2Y205S (Fig. 7C,D). The model predicts the behavior of the channels quite precisely in both cases. The coefficients ta65 and tb65, and ta205 and tb205, describing an allosteric effect on agonist and antagonist binding elicited by the mutations, was indicative of little effect in both cases of the α1F65L mutation and an approximately twofold to fivefold effect in the case of the β2Y205S mutation.

Model of the receptor with two agonist binding sites 1 and 2. The receptor (R) can first bind GABA (A) to either site 1 (AR) or site 2 (RA). Analogously, the receptor can first bind bicuculline (I) to either site 1 (IR) or site 2 (RI). The receptor occupied by two agonist molecules (ARA) can isomerize to the open-state ARA*, and the receptors occupied by a single agonist molecule can isomerize to the open-states AR* and RA*. The model at the top left describes a receptor composed of loose subunits. Concentration modifies L and L1 with fG and fg, respectively. Effects of mutation in site 1 are allosterically transferred to site 2 and vice versa (ta, agonists; tb, antagonists). Constants are taken as dissociation constants and gating constants as closed state divided by open state. Combined Levenberg-Marquardt fit gave the following estimates for the parameters: L, 0.23; L1, 15.3; L2, 13.6; K1, 128 μm; K2, 42 μm; fA, 4.5; fa, 8.6; fG, 20.4; fg, 31.2; K165, 1440 μm; K265, 1320 μm; K1205, 1.3 m; K2205, 14.9 m; ta65, 0.84; tb65, 1.00; ta205, 4.6; tb205, 2.2; K1I, 2.2 μm; K2I, 3.0 μm; m, 97.

A-D, Simulation of the GABA dose-response curves (A, C) and bicuculline inhibition curves (B, D). A, C, γβα/βα, γβα65/βα65 (sites 1 and 2 mutated), γβα65/βα (site 1 mutated), and γβα/βα65 (site 2 mutated) receptors. B, D, αβα/γβ, αβα/γβ205 (site 1 mutated), and αβ205α/γβ (site 2 mutated) receptors. The parameters obtained in a fit are given in Figure 6.
Discussion
The GABAA receptor has been proposed to harbor two functional agonist binding sites. Occupancy of both sites greatly enhances the probability of opening the intrinsic channel. Both sites have been suggested to be located at βα subunit interfaces, but the flanking subunits of α are both β in one case and β and γ in the other. We wanted to alter individual agonist sites by point mutation. In nonlinked receptors, only investigation of wild-type receptors, receptors containing two mutations, or mixtures of receptors containing wild-type and mutated subunits is possible. To circumvent this problem, we used concatenated subunits to dissect the functional roles of the individual binding sites.
In these concatenated subunits, we introduced mutations that affect the properties of the binding sites. We altered αβα/βγ by introducing Y205S into one of the two β subunits or both to create αβ205α/βγ, αβα/β205γ, and αβ205α/β205γ channels. This point mutation has been suggested previously to abolish GABA agonist properties of the affected site (Amin and Weiss, 1993). Similarly, we introduced the point mutation F65L into one of the two α subunits or both in a differently concatenated channel γβα/βα. The homologous residue F64 in rat α1 has been shown to increase the IC50 for the competitive antagonists bicuculline and SR95531 and the EC50 for the agonist GABA for channel opening (Sigel et al., 1992). The same residue was labeled in photoaffinity experiments using the agonist [3H] muscimol (Smith and Olsen, 1994). Amino acid residues including F64 probably adopt a β-sheet structure involved in agonist interaction (Boileau et al., 1999). Thus, the channels γβα/βα, γβα65/βα, γβα/βα65, and γβα65/βα65 were investigated.
Introduction of the mutation β2Y205S into one subunit resulted in very small currents as compared with the corresponding channel containing no mutation. Mutation of site 2 leads to a channel with weaker GABA sensitivity than mutation of site 1. Introduction of the α1F65L mutation in both subunits shifted the GABA concentration-response curve to ∼12-fold higher concentrations. Introduction of the mutation in only one subunit, either the one flanked by two β subunits or the one flanked by a β and γ subunit, caused, in either case, a very similar shift of approximately fivefold to sixfold. Our observations intuitively indicate that both sites must be occupied for the channel to open efficiently. Furthermore, we provide direct evidence for the existence of two agonist sites involving individual α and β subunits.
The interaction with the agonist muscimol was also characterized. Introduction of the mutation β2Y205S indicated a higher apparent affinity of site 1 than site 2, because mutation of site 2 led to a lower EC50 than mutation of site 1. Introduction of the mutation α1F65L in both subunits shifted the muscimol concentration-response curve to only fourfold higher concentrations. This apparently smaller shift than that observed in the case of GABA as agonist could simply reflect a higher open tendency of the receptor with muscimol as agonist. A combined Levenberg-Marquardt fit indeed suggested a higher open tendency combined with a higher affinity of site 1 for muscimol than for site 2.
The interaction with the competitive inhibitor bicuculline was also investigated. Except for the channels carrying the mutations in position β2205, the experiments were always performed at an agonist concentration eliciting ∼10% of the maximal current amplitude in the corresponding mutant. Introduction of the mutation β2Y205S led to occupancy by the agonist of the unmutated site exclusively. Mutation of either site resulted in a similar IC50. Because agonist occupancy was chosen similarly in both cases, this already indicates a similar binding affinity of bicuculline for both sites. Introduction of the mutation α1F65L in both subunits shifted the bicuculline inhibition curve to higher concentrations; the shift was ∼57-fold. Introduction of the mutation in site 2 caused a fourfold shift, and introduction in site 1 caused an eightfold shift. The relative shift between the two curves containing a single mutation is approximately twofold.
Y62 is the homologous residue on the β2 subunit to F65 on the α1 subunit. Introduction of the mutation to L into Y62 on the β subunit to form γβ62α/β62α receptors had little or no effect on the GABA concentration-response curve and on the concentration-dependent inhibition by bicuculline of currents elicited by GABA. This indicates that the homologous site to α1F65 at the homologous subunit interfaces to βα, αβ, and γβ is not involved in GABA-mediated channel opening, and that the effect of the mutation in the β subunit is not allosterically transmitted to the functional binding site at the βα subunit interface. The lack of an effect on the stimulation by diazepam of the current elicited by GABA indicates that the effects of both mutations are not allosterically transmitted to the binding pocket for benzodiazepines at the αγ subunit interface.
Different models were evaluated. The first envisaged channel opening receptors occupied by zero, one, or two agonist molecules (data not shown), the second only of those occupied by one or two agonist molecules (Fig. 6), and the third exclusively of those occupied by two agonist molecules (data not shown). If a significant contribution to channel opening was assumed from nonoccupied receptors or those occupied by a single agonist molecule, the GABA concentration-response curves of channels harboring a single mutation were shifted to the left. Therefore, these reactions were assumed to be relatively minor.
A combined fit for the activation curves by GABA indicated that opening of receptors occupied by two agonist molecules was characterized by an L value of 0.23. This is in good agreement with previous patch-clamp data that indicated 0.24 for this parameter (Amin and Weiss, 1993). The opening of receptors occupied by a single agonist molecule is predicted to be governed by equlibirium constants L1 and L2, of 15 and 14, for sites 1 and 2, respectively. Linkage of receptor subunits is predicted to affect these parameters approximately fivefold for αβα/γβ and ∼20-fold for γβα/βα. The affinity of site 2 is predicted by this model to be approximately threefold higher for GABA than that of site 1. Note that our conclusions may be affected by the fact that we only studied slow phenomena, and we would miss fast desensitization events occurring within the first 500 msec after exposure to agonist in the oocyte expression system (Sigel et al., 1990). Data presented in Figure 4B indicate a preference of site 1 for muscimol. The combined fit for the inhibition curves by bicuculline indicated a slightly higher apparent affinity of site 1 for bicuculline than site 2. Note that for simplicity, our model assumes that bicuculline acts as a pure antagonist despite the fact that bicuculline action has an allosteric component (Ueno et al., 1997).
Our model only includes equilibrium constants. It is evident that the single-channel behavior of every subunit combination will have to be characterized to include rate constants and to see how the linked constructs perform in the fast time range. Natural nonconcatenated receptors have been investigated with the patch-clamp technique in cultured neurons (Macdonald et al., 1989; Weiss and Magleby, 1989; Twyman et al., 1990). Kinetic models explaining these observations have been presented. Results of experiments performed at low GABA concentrations strongly suggested that a single bound molecule of GABA inefficiently opens the receptor with brief open times and noncomplex bursts. The majority of channel openings was interpreted as reflecting the doubly occupied receptor. In the above cases, analysis was complicated by the presence of multiple GABAA receptor types in a single cell. However, a less comprehensive analysis of recombinant α1β1γ2S GABAA receptors indicated a similar kinetic behavior (Angelotti and Macdonald, 1993). In summary, the similarity of our experiments and model supports the intuitive suggestion that both sites must be occupied for the channel to open efficiently, a fact predicted by single-channel kinetic analysis, and that occupancy of a single site by bicuculline is sufficient to prevent channel opening.
How can it be explained that the two sites display different properties for agonist and similar properties for competitive antagonist interaction? Each agonist or antagonist molecule contacts several atoms in the receptor. The fact that both sites are located at the βα subunit interface but display subtly different properties may be explained by a subtly different conformation of the two sites, depending on the nature of the flanking subunits (Fig. 8). Note that the published structure of the acetylcholine binding protein has five identical subunits (Brejc et al., 2001), and the atom coordinates of the recently proposed model by Cromer et al. (2002) are not available, such that our predicted structural differences of the two binding sites cannot be visualized.

Schematic representation of the GABAA receptor showing the two functional agonist sites (G) and the benzodiazepine binding site (B). Agonist site 2 has a preference for the agonist GABA, and site 1 has a preference for muscimol and a slight preference for the antagonist bicuculline.
Analogously to the methods presented here, the role of any individual site located on an α or β subunit may be studied, which was impossible thus far because there were always two sites on the two subunits affected by a mutation in 2α2β1γ GABAA receptors. The method enables targeted introduction of a mutation in only one defined subunit. Because the GABAA receptor belongs to a superfamily of ligand-gated ion channels, procedures described here are applicable to the entire family of ligand-gated channels comprising nicotinic acetylcholine, glycine, and 5HT3 receptors.
Footnotes
This work was supported by Swiss National Science Foundation Grant 3100-064789.01/1. We thank Dr. C. Wagner for helpful discussions on modeling, Heleen van Hees for help in preparing the linked subunit constructs, and Dr. V. Niggli for carefully reading this manuscript.
Correspondence should be addressed to Erwin Sigel, Department of Pharmacology, Friedbuehlstrasse 49, CH-3010 Bern, Switzerland. E-mail: erwin.sigel{at}pki.unibe.ch.
Copyright © 2003 Society for Neuroscience 0270-6474/03/2311158-09$15.00/0





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}