

Abstract
Wakefulness depends on the activity of hypocretin–orexin neurons because their lesion results in narcolepsy. How these neurons maintain their activity to promote wakefulness is not known. Here, by recording for the first time from hypocretin–orexin neurons and comparing their properties with those of neurons expressing melanin-concentrating hormone, we show that hypocretin–orexin neurons are in an intrinsic state of membrane depolarization that promotes their spontaneous activity. We propose that wakefulness and associated energy expenditure thus depend on that property, which allows the hypocretin–orexin neurons to maintain a tonic excitatory influence on the central arousal and peripheral sympathetic systems.
Introduction
Wakefulness has been shown to depend on the newly identified hypocretin–orexin (Hcrt/Orx) neuropeptides (de Lecea et al., 1998; Sakurai et al., 1998) by findings that alterations in their precursor protein, their receptors, or the neurons that produce them lead to the sleep disorder narcolepsy in both animals and humans (Chemelli et al., 1999; Lin et al., 1999; Peyron et al., 2000;Thannickal et al., 2000). In normal individuals, Hcrt/Orx neurons, by their activity, presumably provide a waking drive to the multiple activating systems to which they project (Peyron et al., 1998), including the central arousal and peripheral sympathetic systems. A persistent drive on these targeted systems would underlie the dual role of the Hcrt/Orx neurons in simultaneously promoting waking and energy expenditure, functions that are compromised in narcolepsy and the moderate obesity that follows their lesion (Hara et al., 2001). Without recording from identified Hcrt/Orx neurons, the properties that allow them to fulfill this unique role remain unknown (for review, seeSiegel, 1999; Kilduff and Peyron, 2000; Hungs and Mignot, 2001; Willie et al., 2001). Indeed, whether these cells depend on inputs from other systems or could be endowed with particular membrane properties for their activity remains to be determined.
Materials and Methods
Electrophysiology. Coronal brain slices containing numerous Hcrt/Orx neurons in the perifornical area of the hypothalamus were chosen according to the atlas of Peyron et al. (1998). In the same area, but in smaller number, neurons containing melanin-concentrating hormone (MCH) are intermingled with Hcrt/Orx neurons. Before use, slices (300-μm-thick) obtained from young rats (15–28 d) were incubated at room temperature in artificial CSF (ACSF) containing the following (in mm): 130 NaCl, 5 KCl, 1.25 KH2PO4, 1.3 MgSO4, 20 NaHCO3, 10 glucose, and 2.4 CaCl2 (bubbled with 95% O2 and 5% CO2). For sodium substitutions, we used the following (in mm): 145N-methyl-d-glucamine (NMDG) (or choline), 5 KCl, 10 HEPES, 10 glucose, 1.3 MgSO4, 2.4 CaCl2, pH 7.4. Whole-cell recordings were obtained (Bayer et al., 2002) with patch electrodes (8–12 MΩ) containing the following (in mm): 126 KMeSO4, 4 KCl, 5 MgCl2, 10 HEPES, 8 phosphocreatine, 3 Na2ATP, 0.1 NaGTP, and 0.1 BAPTA, pH 7.3. Access resistance was between 15 and 25 MΩ. Values for membrane potentials are uncompensated for junction potentials [estimated junction potential, −9.6 mV; based on the JpCalc software provided with the data acquisition system by Axon Instruments (Foster City, CA)]. In some experiments, BAPTA was raised to 20 mm(KMeSO4 and lowered to 86 mm). To identify cells, 0.2% neurobiotin was added to the intrapipette solution.
Immunohistochemistry. After recordings, slices were immersed successively in ice-cold 3% paraformaldehyde for 2–12 hr and in 30% sucrose for 24–48 hr. They were then stored at −80°C to be later cut with a cryostat in 45-μm-thick sections. After two rinses with Tris NaCl, pH 7.4, sections were submitted to staining for Hcrt/Orx and MCH. First, sections were incubated 24 hr at room temperature with a goat polyclonal IgG against Hcrt/Orx A (dilution at 1:200; Santa Cruz Biotechnology, Santa Cruz, CA) and with a rabbit polyclonal IgG against MCH (dilution at 1:200; gift from Prof. D. Fellmann, University of Besançon, Besançon, France). Second, they were rinsed (two times) with Tris NaCl and incubated for 3 hr at room temperature in the presence of an anti-goat IgG-Cy3, an anti-rabbit IgG-7-amino-4-methylcoumarin-3-acetic acid, and Cy2-conjugated streptavidin (dilutions at 1:200 for all antibodies; Jackson ImmunoResearch, West Grove, PA). Neurobiotin-filled neurons appeared in green, Hcrt/Orx-positive cells in red, and MCH-positive cells in blue. In a few occasions only Hcrt/Orx-injected neurons were sought, in which case a similar protocol but without MCH antiserum was used. Specificity of MCH staining has been checked in previous studies (references in Brischoux et al., 2002). For Hcrt/Orx staining, specificity was demonstrated by preabsorption of the antiserum (1:100) with orexin A (20 μg/ml).
Results
Identification of Hcrt/Orx neurons
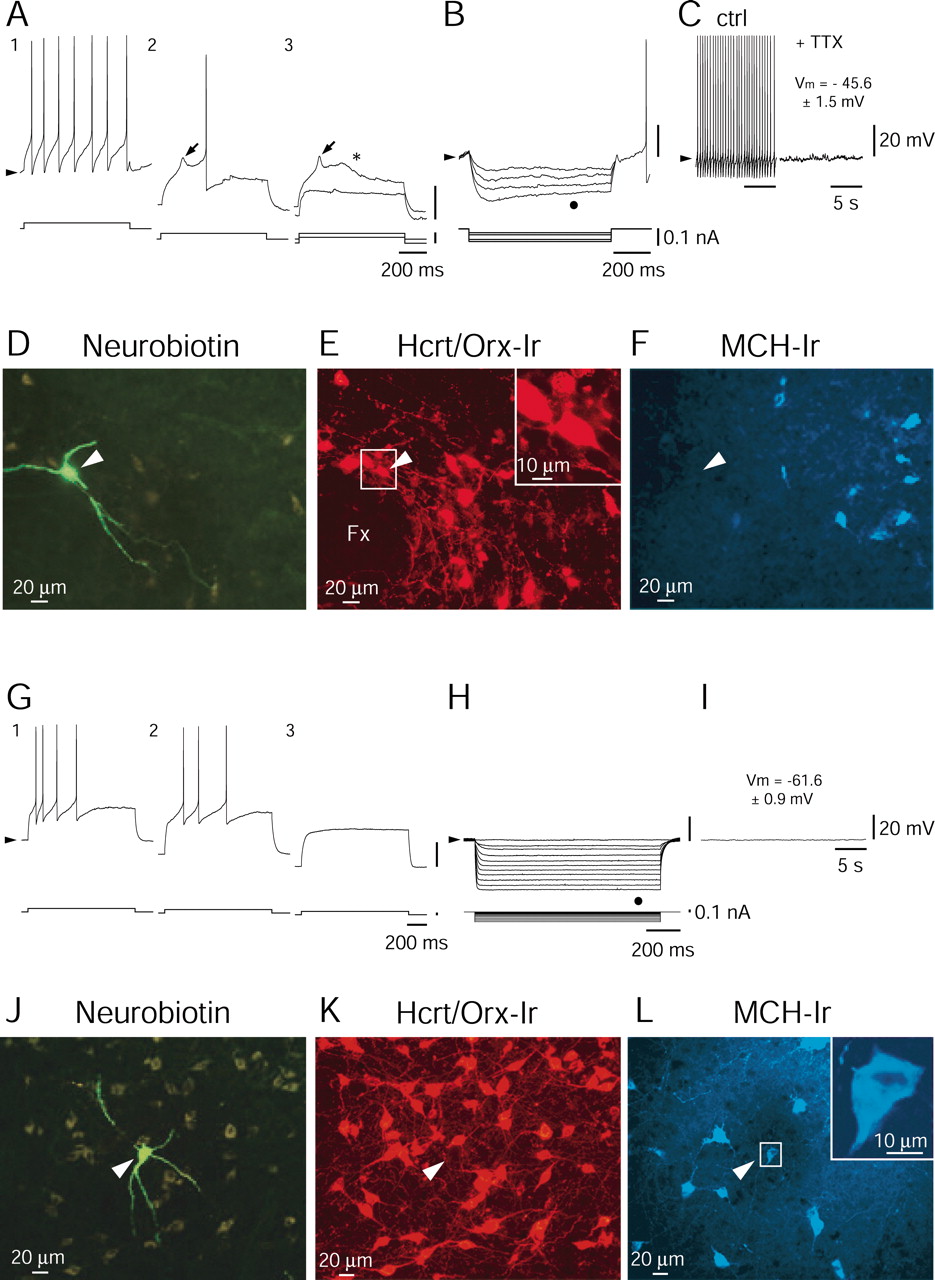
Hypothalamic neurons were recorded in rat brain slices using the whole-cell technique. Under infrared video microscopy, large-sized (≥20 μm) neurons were selected in the perifornical area, which is known to contain a large number of Hcrt/Orx neurons intermingled with a smaller population of cells containing the MCH (Broberger et al., 1998), a peptide thought to exert an action opposite to Hcrt/Orx on energy metabolism (Qu et al., 1996; Shimada et al., 1998). One class of cells immediately became of interest as potentially expressing Hcrt/Orx on the basis of its clear preponderance, representing 49.7% of cells recorded in rats aged 15–20 d (n = 82 of 165) and up to 71.7% in those aged 25–28 d (n = 33 of 46). The main characteristic of cells belonging to this class are illustrated in Figure 1A–C. First, when they were depolarized from the resting level, they responded by tonic firing with little adaptation (Fig.1A1 ). As they were challenged by depolarizing current pulses delivered from a hyperpolarized level, a low-threshold spike (LTS) was revealed (Fig.1A2 ,A3 , arrows), followed by a slow afterdepolarization (ADP) (Fig.1A3 , asterisk). Together, the LTS and ADP, shown separately in Figure1A3 , represent the key features that distinguish these cells from all neighboring cells. It is also noteworthy that the LTS in these neurons was never crowned (at any membrane potential) by a high-frequency burst of action potentials. Second, in the presence of hyperpolarizing current pulses, all of these cells showed a membrane rectification characterized by a sag (Fig.1B, dot), which in other neurons has been identified as a time- and voltage-dependent rectification resulting from the presence of an Ih current. Finally, most neurons (n = 59 of 82, or 72%) with the above characteristics were spontaneously active (Fig. 1C), with a mean firing frequency of 3.17 ± 1.53 Hz (for those active,n = 59). As evidenced in the presence of tetrodotoxin (TTX) (1.0 μm) (Fig. 1C), neurons of this type had a rather depolarized resting membrane potential (mean ± SEM, −45.6 ± 1.53 mV; n = 11; see Materials and Methods). Triple immunohistochemical staining for neurobiotin (Fig. 1D), Hcrt/Orx (Fig.1E), and MCH (Fig. 1F) revealed that all neurons (n = 11 of 11) of this type were Hcrt/Orx positive. In contrast, none of them were MCH positive. Similar results were obtained with dual immunohistochemical staining for neurobiotin and Hcrt/Orx alone (n = 7 of 7; data not shown). In those injected cells in which morphology could be properly evaluated (n = 12), we found that two Hcrt/Orx neurons appeared bipolar, whereas the remaining (10 of 12) were multipolar (three main dendrites).

Characterization of neurons expressing Hcrt/Orx or MCH. A1, Tonic firing in response to a depolarizing current pulse delivered from the level of resting potential (arrowhead).A2 ,A3 , LTS (arrow) and ADP (asterisk) triggered by a depolarizing current pulse delivered from an hyperpolarized level. Additional hyperpolarization eliminates the LTS and the ADP (bottom trace in A3 ).B, Superimposed responses to hyperpolarizing pulses suggesting the presence of an Ih current (dot). Note that only the trace with the deepest hyperpolarization is shown in full. C, Tonic firing at rest and its elimination by TTX (1.0 μm) to determine resting potentials. D, F, Immunohistochemical identification of an Hcrt/Orx neuron injected with neurobiotin (arrowhead in D) and expressing immunoreactivity for Hcrt/Orx (E) but not for the MCH (F). G1 , Firing with accommodation triggered by a depolarizing current pulse delivered from the resting potential level.G2 ,G3 , Absence of either LTS or ADP in response to depolarizing pulses applied from more hyperpolarized levels. H, Responses to hyperpolarizing current pulses demonstrating the absence (dot) of any sag that could have indicated the presence of an Ihcurrent. I, Absence of spontaneous firing in such neurons and their mean resting potential. J,L, Immunohistochemical identification of an MCH neuron injected with neurobiotin (arrowhead inJ) and expressing immunoreactivity for MCH (L) but not for Hcrt/Orx (K). Membrane potentials are as follows (arrowheads): −47 mV (A), −44 mV (B), −48 mV (C), −61 mV (G), and −61 mV (H,I). Fx, Fornix.
Identification of MCH neurons
In striking contrast to the Hcrt/Orx neurons, another group of cells (17.58% in rats aged 15–20 d, n = 29 of 165; and 13.04% in those aged 25–28 d, n = 6 of 46) was identified that had completely different characteristics. The first defining property of this group was the complete absence of a sag during hyperpolarizing pulses delivered from rest (Fig.1H), thus indicating the absence of anIh current. Second, neurons having the above-described property never displayed an LTS or an ADP (Fig.1G1–G3 ). Finally, these cells also differed from Hcrt/Orx neurons by the level of their resting membrane potential, which was much more hyperpolarized (mean ± SEM, −61.6 ± 0.86 mV; n = 29). None of these neurons discharged spontaneously (Fig.1I). Indeed, their membrane characteristics appeared to render them silent, at least in the absence of synaptic activity. As illustrated in Figure1J–L, triple labeling revealed that all (n = 8 of 8) of the neurobiotin-stained cells (Fig. 1J) of this group were Hcrt/Orx negative (Fig. 1K) but MCH positive (Fig. 1L). In those injected cells in which morphology could be properly evaluated (n = 6), we found that MCH neurons were all multipolar (three to four main dendrites).
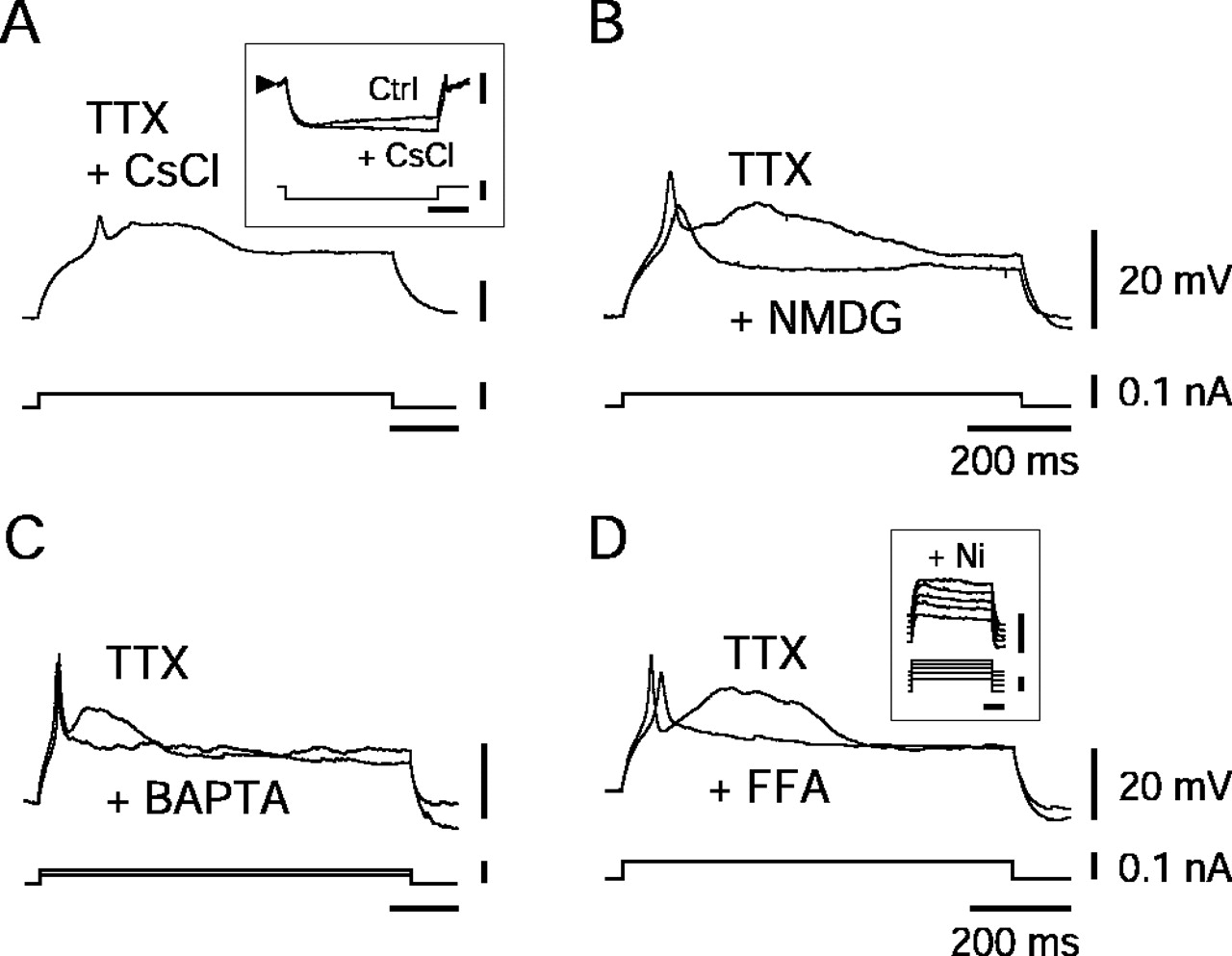
Properties of hypocretin–orexin neurons.A, Persistence of the LTS and ADP in the presence of TTX (1.0 μm) and cesium chloride (2 mm).Inset illustrates elimination of the time- and voltage-dependent sag by cesium chloride. B,D, Elimination of the ADP but persistence of the LTS in presence of NMDG (B), BAPTA (20 mm;C), or FFA (100 μm; D).Inset in D shows that further addition of Ni (200 μm) eliminates the LTS.
Properties of Hcrt/Orx neurons
Given the evident importance of activity in Hcrt/Orx neurons for maintaining wakefulness and promoting energy expenditure (for review, see Kilduff and Peyron, 2000; Hungs and Mignot, 2001; Siegel et al., 2001; Willie et al., 2001), we proceeded to further investigate their intrinsic properties. We began by examining the nature of the ADP by using TTX, a specific blocker of voltage-dependent sodium channels. Although TTX (1.0 μm) eliminated the action potentials, it failed (n = 11 of 11) to suppress either the ADP or the LTS, as illustrated in Figure 2A. As also shown in this figure, cesium chloride (2–3 mm), which completely eliminated the time- and voltage-dependent sag attributable to the Ih(n = 7 of 7) (Fig. 2A,inset), affected neither the LTS nor the ADP. We then hypothesized that the ADP might reflect the presence of a nonselective cation current (Bal and McCormick, 1993). To test this hypothesis, we first substituted sodium chloride with NMDG in the perfusion solution and indeed found that it completely eliminated (n = 2 of 2) the ADP (Fig. 2B) without affecting the LTS. This result was confirmed (n = 3 of 3) by substituting sodium chloride with choline chloride that again eliminated the ADP but left the LTS unaltered (data not shown). We subsequently tested the calcium dependence of this ADP by internally perfusing BAPTA (20 mm) into the neurons (Fig. 2C) and again observed that the ADP was eliminated (n = 7 of 7) but the LTS persisted. These results suggest that the ADP uncovered in Hcrt/Orx neurons must be attributable to the presence of a strong calcium-activated nonselective cation current (ICAN) (Bal and McCormick, 1993;Partridge et al., 1994). To further test this possibility, we finally applied flufenamate (FFA), an anti-inflammatory drug known to block such currents (Partridge and Valenzuela, 2000). As illustrated in Figure 2D, when added to TTX, FFA (100–200 μm) completely eliminated the ADP (n = 6 of 6) but left the LTS unaffected. As evidenced in the inset of Figure2D, adding small doses of nickel (Ni) (100–200 μm) to FFA (in the presence of TTX) completely eliminated the LTS (n = 2 of 2). The latter result suggests that the LTS must depend on the presence of low-voltage-activated (LVA) calcium channels, which are known to be sensitive to small doses of nickel. It is probable that, in the experimental condition used to reveal the ADP, it is actually the calcium entering through these LVA channels that activates theICAN. As evidence for this point, it is noteworthy that small doses of nickel (100–200 μm), when applied in the presence of TTX but in the absence of FFA, indeed completely eliminated both the LTS and the ADP (n = 3 of 3; data not shown).

Persistence of the membrane depolarization and spontaneous activity of hypocretin–orexin neurons in conditions of synaptic blockade and their inhibition by GABA. A, Persistence of the membrane depolarization in TTX (1.0 μm), ionotropic blockers (MK801 at 20 μm, NBQX at 10 μm, and bicuculline at 10 μm), and an ACSF with 0.1 mm Ca2+ and 10 mm Mg2+. B, Persistence of the spontaneous activity in the presence of synaptic blockade (right is an enlargement of the area identified by anasterisk in the left). C, Inhibition by a brief application of muscimol (5 sec at 100 μm). Membrane potentials are as follows (arrowheads): −42 mV (A), −43 mV (B), and −44 mV (C).
The membrane depolarization of Hcrt/Orx neurons persists when either synaptic transmission or Ih are suppressed
Although, as demonstrated above, Hcrt/Orx neurons are equipped with a set of intrinsic properties, all potentially conducive of a high level of electrical activity, it remains possible that the major determinant for their depolarized and active state is of synaptic origin. Given the presence of spontaneous synaptic potentials in these cells, we tackled that question first by blocking ionotropic receptors to glutamate [(+)-5-methyl-10,11-dihydro-5H-dibenzo [a,d] cyclohepten-5,10-imine maleate (MK801) (20 μm) and 2,3-dihydroxy-6-nitro-7-sulfonyl-benzo[f]quinoxaline (NBQX) (10 μm) to block NMDA and AMPA receptors, respectively] and GABA (bicuculline at 10 μm to block GABAAreceptors) and show that such a condition had no effect on either their resting membrane potential (Fig.3A) (n = 3 of 3 in the presence of TTX) or their spontaneous activity (n = 6 of 6; data not shown). As a second step, we tested the effect of a more general condition of synaptic blockade (0.1 mm Ca2+ and 10 mm Mg2+) and found again that it affected neither the resting potential (Fig.3A) (n = 5 of 5 in the presence of TTX) nor the spontaneous activity (n = 3 of 3) (Fig.3B) of these cells.
In addition to showing that the persistent depolarization and activity of Hcrt/Orx neurons are not of synaptic origin, the above results indicate that voltage-dependent calcium currents are not implicated either. Along this line, it is noteworthy that theIh current, which could have been involved in the depolarization and activity of Hcrt/Orx neurons, also plays no role. Indeed, cesium (3 mm), which was shown to block the Ih, affected neither the resting potential (n = 5 of 5; data not shown) nor the activity (n = 3 of 3; data not shown) of these neurons.
Inhibition of Hcrt/Orx neurons by GABA
Of final notice, Hcrt/Orx neurons were strongly inhibited (Fig.3C) by the GABAA agonist muscimol, briefly applied at 100 μm (n = 4 of 4). This effect persisted in the presence of either TTX or a high Mg2+–low Ca2+ solution (altogethern = 3 of 3; data not shown).
Discussion
This study presents the first recordings of identified Hcrt/Orx and MCH neurons. It shows that Hcrt/Orx neurons, in contrast to MCH neurons, are in a depolarized state that promotes their activity. This property of Hcrt/Orx neurons could contribute importantly to their suggested role in maintaining wakefulness.
Our data show first that the depolarized and active state of Hcrt/Orx neurons is not of synaptic origin because it persists in a condition in which synaptic transmission is blocked. As a result, one is led to propose that the depolarized and active state of Hcrt/Orx neurons is intrinsic in nature. Among the intrinsic properties that might play a role in directly promoting the spontaneous firing of Hcrt/Orx neurons are the presumed Ih and LVA currents, which in other neurons have been implicated in rhythmic activity. Such is not the case here, however, because blocking either of these currents had no influence on the spontaneous firing of Hcrt/Orx neurons. The conditions in which these currents would become active is not known, but one might speculate that they could support activity in the face of hyperpolarizing synaptic inputs. Hcrt/Orx cells are also endowed with a presumed ICAN (Bal and McCormick, 1993; Partridge et al., 1994), revealed as an ADP after activation of an LVA current when the cells are depolarized from an hyperpolarized level. An ICAN that would only depend on the previous activation of an LVA current cannot, however, explain the spontaneous firing of Hcrt/Orx cells as recorded here, because the activity of these cells persists when voltage-dependent calcium currents are blocked.
In view of the above, it is reasonable to assume that the key factor in promoting the spontaneous activity of Hcrt/Orx cells is an intrinsic state of membrane depolarization that keeps them constantly near their firing threshold. Our data suggest, however, that this state does not depend on the presence of either sodium or calcium voltage-dependent currents and that the Ih current is not implicated. Given the importance of the presumedICAN in Hcrt/Orx neurons, one is led to speculate that this current could play an important role in the persistent membrane depolarization. In that case, the calcium needed for activation of ICAN could originate from channels other than the voltage-dependent calcium channels or from intracellular stores. A clear-cut demonstration that anICAN contributes to the state of membrane depolarization of Hcrt/Orx neurons cannot, however, be achieved at present given the absence of selective antagonists of calcium-activated cation currents.
The enduring activity that results from the properties of Hcrt/Orx neurons would allow them to excite in a persistent manner their multiple targets (Peyron et al., 1998) that include the major central activating systems (Jones, 2000). Indeed, the Hcrt/Orx peptides have been shown recently to exert a depolarizing and excitatory effect on noradrenergic (Hagan et al., 1999; Horvath et al., 1999; Bourgin et al., 2000; Brown et al., 2001), histaminergic (Bayer et al., 2001;Eriksson et al., 2001), cholinergic (Methippara et al., 2000; Eggermann et al., 2001; Xi et al., 2001; Burlet et al., 2002), and thalamic intralaminar (Bayer et al., 2002) neurons. The autochthonous drive to these activating systems could in fact derive from the Hcrt/Orx neurons, as evidenced by the dramatic consequences during wakefulness of their destruction seen in transgenic mice and human narcoleptics (Nishino et al., 2000; Peyron et al., 2000; Thannickal et al., 2000;Hara et al., 2001).
The intrinsic properties of the Hcrt/Orx neurons suggest that their natural state is depolarized and active, by which they would promote wakefulness, and that their inhibition would be necessary for allowing sleep when, from c-Fos studies, they indeed appear to be less active (Estabrooke et al., 2001). Here, not surprisingly, we were able to verify that Hcrt/Orx neurons are indeed inhibited by GABA. GABAergic neurons, located in the preoptic and basal forebrain areas, which project to the posterior hypothalamus and become active during sleep, could provide this inhibition (Sherin et al., 1998; Szymusiak et al., 1998; Gallopin et al., 2000) (for review, see Jones, 2000; Kilduff and Peyron, 2000; Hungs and Mignot, 2001; Saper et al., 2001).
With respect to the influence of Hcrt/Orx neurons on metabolism, the findings that narcoleptic patients have an increased body-mass index (Schuld et al., 2000) and that mice with selective destruction of Hcrt/Orx neurons become moderately obese although hypophagic (Hara et al., 2001) suggest that Hcrt/Orx neurons also stimulate energy expenditure. One possible mechanism for this action could be the stimulatory effect of the Hcrt/Orx peptides on the sympathetic nervous system (Antunes et al., 2001) to which they project (Peyron et al., 1998). Here again, the intrinsic properties of Hcrt/Orx neurons, by favoring their depolarized and active state, could enable them to exert a tonic influence on the preganglionic sympathetic neurons. It is notable that MCH neurons, which also have widespread projections throughout the CNS (Bittencourt et al., 1992), but have in contrast been linked to energy conservation and decreased metabolic rate (Qu et al., 1996; Shimada et al., 1998), are hyperpolarized and inactive in the resting state. As suggested by a number of studies, MCH neurons must depend for their activation, as occurring particularly in conditions of food deprivation, on various hunger–satiety-related signals arising from the periphery and/or hypothalamic centers involved in the control of feeding behavior (for review, see Spiegelman and Flier, 2001).
In conclusion, our results lead us to propose that wakefulness and associated energy expenditure could depend on the intrinsic characteristics of Hcrt/Orx neurons, which, by maintaining them active, allow them to maintain an excitatory influence on their targeted central arousal and peripheral sympathetic systems.
Footnotes
↵* E.E., L.B., and M.S. contributed equally to this work.
This study was supported by the following: grants from the Swiss Fonds National, Novartis, OTT, and de Reuter and Schmidheiny Foundations (M.M. and M.S.); the Canadian Medical Research Council (B.E.J.); and a Roche fellowship (L.B.).
Correspondence should be addressed to Dr. M. Mühlethaler, Centre Médical Universitaire, Département de Physiologie, 1 Rue Michel-Servet, 1211 Genève 4, Suisse. E-mail:michel.muhlethaler{at}medecine.unige.ch.





{kind=link}
{kind=link}
{kind=link}