

Article Figures & Data
Figures
- Figure 1.
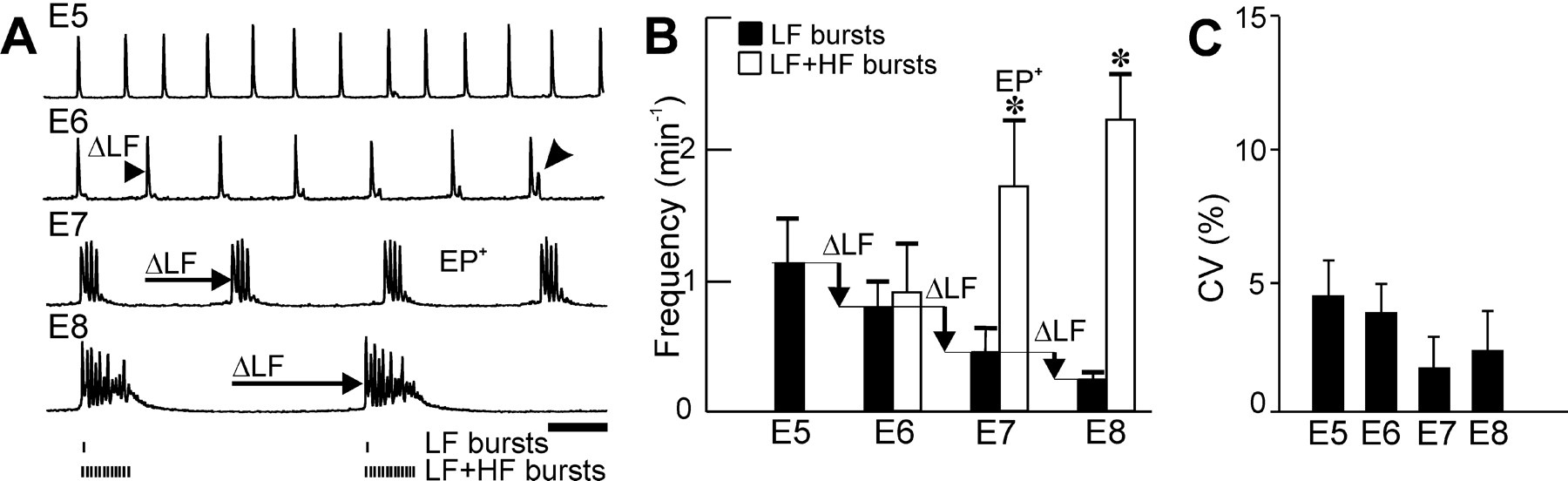
Appearance of episodic patterning of activities (EP+): transition from LF primordial activity. Central pattern generators for LF and episodes are identified by recording neuronal activities after isolation of the hindbrain in vitro. A, Integrated facial neurograms recorded at different developmental stages (E5-E8) show a progressive decrease (ΔLF; horizontal arrows) in the frequency of LF bursts initiated before E6 and maintained thereafter. B, The EP+ pattern is identified by the significant difference (*) between the frequency of LF bursts (black bars) and that of LF + HF bursts (empty bars). Note the exclusive presence of LF bursts at E5 and the emergence (A, arrowhead on the E6 trace) of HF bursts at E6. C, Regularity of rhythm generation, measured by the CV of inter-LF burst interval, is maintained during development. Scale bar, 1 min.
- Figure 2.
Anterior-odd rhombomeric code for the induction of EP+. Integrated glossopharyngeal (A, B) or trigeminal (C) neurograms obtained after heterotopic grafting and isolation. Transplantations of the odd r3 (in r5 position; A, arrowhead) and of the even r6 (in r4 position; B, arrowhead) or r2 (in r4 position; C, arrowhead) lead to r3r6 or r3r2 preparations showing an episodic patterning similar to that obtained from the normal configuration r3r4. D, E, Summary histograms of the frequencies of LF bursts and LF + HF bursts (D, filled and empty bars, respectively) and of CV (E) obtained after pooling preparations according to the heterotopic status of either odd (r3, r5) or even (r2, r4, r6) rhombomeres (detailed in Table 1) and the anterior or posterior status of the displacement along the neuroaxis (anterior shift and posterior shift) of rhombomeres, whatever their parity (detailed in Table 1). In all cases, an EP+ pattern comparable with that of the normal r3r4 pair resulted. Surgical isolation, shift, and grafting of rhombomeres do not interfere with the development of the EP+ in artificial rhombomere pairs in which the odd rhombomere is in contact with the anterior edge of the even rhombomere. Scale bar, 1 min.
- Figure 3.
Electroporation of Krox20 confers the EP+ pattern to the r4 and r1r4 EP- segments. Integrated neurograms (top) and corresponding compound distribution of interburst interval durations (bottom; n% ordinates; duration abscissae) recorded at E7 from the facial nerve root after development of the neuronal network deriving from EP- r4 (A) and r1r4 (C). An EP+ pattern of activity (B, D) results in 4 misexpressing Krox20 (4K20 in B) and in r1r4 segments when r1 misexpresses Krox20 in the pair (1K20 in D) resulting in a bimodal distribution of interburst interval. E, Summary histogram of the frequency of LF bursts and LF + HF bursts (E, filled and empty bars, respectively) showing that r2, r4, r6, r1r2 (normal configurations), and r1r4 are EP-. Electroporation of Krox20 induces EP+ in r4 and r1r4 (asterisks; compare with the normal configuration, r3r4). F, CV is not significantly affected in these experiments and is particularly large for r6. Scale bar, 1 min.
- Figure 4.
Whole-mount r1(K20)r4 islands 24 hr after electroporation grafting and isolation. A, B, Two examples of rhombomere islands (dorsal views) produced by heterotopic grafting in r3 position of r1 territories taken from donor embryos that had been submitted to unilateral electroporation of mixed GFP and Krox20 expression plasmids at the r1 level, before engraftment and isolation of the r1r4 island. Note the overt healing at boundaries (A and B, dotted lines) separating r1 anterior and r4 posterior and the ovoid aspect taken by the isolated rhombomere pairs. Forced Krox20 expression (red in A, dark blue in B) is visible on the electroporated side (ep.) on the left of the medial axis (A, vertical line) but not on the control side (cont.; right) of the r1 territory. Krox20 expression is confined within r1 (A, B), and a sharp AP limit of expression is visible on the ep. side in B, where as endogenous bilateral Hoxb1 expression (blue in A, not probed in B) is restricted to r4. The activity produced by this rhombomeric configuration is illustrated in Figure 3 D. A, Anterior; P, posterior. Scale bar, 50 μm.
- Figure 5.
Anteroposterior orientation of r4, but not r3, is required for the development of a stable rhythm generator. A-D, Two examples (superposed traces) of integrated motor activity (left) recorded from distinct rhombomere pair preparations (left schematic) and compound histograms of the corresponding distribution of interburst interval duration (right) recorded from the facial nerve. A, B, Episodic patterning is present in r3r4 (normal configuration) and r3r4 after AP reversal of r3 (B), absent (C, D) in r3r4 after AP reversal of r4 (C), and in r4r3 after heterotopic transplantation of r3 (in r5 position; D, arrowhead). E, F, Summary histogram of the frequency of LF bursts and LF+HF bursts (E, filled and empty bars, respectively) and of CV (F) of the different preparations indicated by their lettering in the figure. Reversal of r4 eliminates regularity of the rhythm as indicated by significantly higher mean CV in F (compare c, a). Note also in E that modifying the relative position of r3 with respect to anteroposterior polarity of r4 abolishes (in c and d) the development of episodes (a and b). Scale bar, 1 min.
- Figure 7.
Horizontal sections of preparations, including the anteroposterior reversal of r4, 24 hr after surgical manipulation. Insitu hybridization for Krox20 (blue) and Hoxb1 (red) in a control hindbrain (A) and in isolated rhombomere island preparations (B-F). Normal configurations, r2r3 (B) and r4r5 (D); modified configurations, AP inverted (f) pair of segment r3r4 (C) and r4r5 (F) and flipped r4 within the r4r5 pair (E). In all cases, Hoxb1 expression is preserved in r4 and Krox20 expression in r3 and r5, although episodic patterning is present (EP+ in A, C, E) or absent (EP- in B, D, F). Scale bar, 50 μm.
- Figure 6.
Anteroposterior reversal of r4 produces the development of an episodic generator in r4r5. A-D, Two examples (superposed traces) of integrated motor activity (left) recorded from distinct rhombomere pair preparations (left schematic) and compound histograms of the corresponding distribution of interburst interval duration (right) recorded from the facial nerve. A, B, Episodic patterning is absent in r4r5 (A, normal configuration) and after reversal of the entire r4r5 pair (B) and is produced (C, D) in r4r5 after AP reversal of r4 alone (C) and in r5r4 after heterotopic transplantation of r5 (in r3 position; D, arrowhead). E, F, Summary histogram of the frequency of LF bursts and LF+HF bursts (E, filled and empty bars, respectively) and of CV (F) of the different preparations. The EP+ pattern is produced (E, asterisks) by modifying the relative position of r5 with respect to the anteroposterior orientation of r4 (in c and d but not in a and b). Reversal of r4 (B, C) also eliminates regularity of the rhythm as indicated by significantly higher mean CVs (F, asterisks). Scale bar, 1 min.







Tables
Preparations n LF bursts LF+HF bursts EP− E5 hindbrain 7 1.15 ± 0.34 id. E6 hindbrain 5 0.80 ± 0.20 0.92 ± 0.21, NS r2 3 2.66 ± 1.12 id. r4 4 2.38 ± 0.28 id. r6 5 2.15 ± 0.66 id. r2r3 3 1.11 ± 0.27 id. r4r5 (at E6) 5 2.25 ± 1.01 id. r4r5 9 2.20 ± 0.67 id. r1r2 2 1.42 ± 0.49 id. r1r4 7 1.26 ± 0.57 id. r1(GFP)r4 3 1.10 ± 0.23 id. f(r4r5) 9 1.21 ± 0.35 1.96 ± 1.03, NS r3f(r4) 4 1.28 ± 0.20 1.64 ± 0.36, NS r4r3* 2 2.02 ± 0.12 2.21 ± 0.27, NS EP+ E7 hindbrain 21 0.45 ± 0.21 1.75 ± 0.51 E8 hindbrain 8 0.24 ± 0.06 2.27 ± 0.35 r3r4 6 0.72 ± 0.08 2.78 ± 0.21 r5r6 4 0.79 ± 0.10 2.55 ± 0.24 r5*r4 3 1.10 ± 0.51 3.46 ± 0.33 Heterotopic odd 5 0.72 ± 0.14 2.96 ± 0.54 Heterotopic even 4 0.67 ± 0.12 2.84 ± 0.39 Anterior shift 5 0.96 ± 0.41 3.02 ± 0.69 Posterior shift 4 0.66 ± 0.12 2.75 ± 0.49 r4(K20) 4 0.84 ± 0.15 3.01 ± 0.38 r1(K20)r4 5 1.05 ± 0.32 2.85 ± 0.81 f(r3)r4 7 0.75 ± 0.13 2.95 ± 0.62 f(r3r4) 7 0.72 ± 0.14 3.11 ± 0.94 f(r4)r5 5 0.96 ± 0.62 3.13 ± 0.82 -
Summary table of the values of LF burst and LF+HF burst frequencies (minute−1) measured on the different preparations at E7. Anteroposterior position of rhombomeres in rhombomere pairs is indicated from left to right (e.g., r4r5-r4 anterior to r5). An asterisk to the right of a rhombomere indicates that the rhombomere has been heterotopically grafted (e.g., r4r3*-r3 grafted in heterotopic r5 position). The preceding rhombomeres in brackets indicates their flipped anteroposterior orientation. GFP or K20 in a bracket to the right of a rhombomere indicates that this rhombomere was targeted by electroporation of the sole GFP or the GFP and Krox20 expression vectors, respectively. Heterotopic odd and even refer to compound measurements pooling, respectively, r3*r6 (n = 1), r5*r4 (n = 3), and r5*r2 (n = 1) and r3r6* (n = 1), r3r2* (n = 2), and r5r4* (n = 1) preparations. Anterior and posterior shift refer to compound measurements obtained pooling, respectively, r3r6* (n = 1), r5*r4 (n = 3), and r5*r2 (n = 1) and r3*r6 (n = 1), r3r2* (n = 2), and r5r4* (n = 1) preparations. id., Idem; NS, no significant difference between frequency values of LF bursts and LF+HF bursts.
-





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}