

Abstract
NMDA receptor activity is important for many physiological functions, including synapse formation and alterations in synaptic strength. NMDA receptors are composed most commonly of NR1 and NR2 subunits. There are four NR2 subunits (NR2A–NR2D). NR2 subunit expression varies across both brain regions and developmental stages. The identity of the NR2 subunit within a functional NMDA receptor helps to determine many pharmacological and biophysical receptor properties, including strength of block by external Mg2+ (Mgo2+). Mgo2+ block confers strong voltage dependence to NMDA receptor-mediated responses and is critically important for many of the functions that the NMDA receptor plays within the CNS. Here we describe the NR2 subunit dependence of the kinetics of Mgo2+ unblock after rapid depolarizations. We find that Mgo2+ unblocks from NR1/2A and NR1/2B receptors with a prominent slow component similar to that previously described in native hippocampal and cortical NMDA receptors. Strikingly, this slow component of Mgo2+ unblock is completely absent from NR1/2C and NR1/2D receptors. Thus currents from NR1/2C and NR1/2D receptors respond more rapidly to fast depolarizations than currents from NR1/2A and NR1/2B receptors. In addition, the slow component of Mgo2+ unblock from NR1/2B receptors is consistently slower than from NR1/2A receptors. This makes rapid depolarizations, such as action potential waveforms, more efficacious at stimulating Mgo2+ unblock from NR1/2A than from NR1/2B receptors. These NR2 subunit differences in the kinetics of Mgo2+ unblock are likely to help determine the contribution of each NMDA receptor subtype to current flow during synaptic activity.
Introduction
Postsynaptic NMDA receptor responses are characterized by strong voltage dependence, slow decay, and a large calcium (Ca2+) conductance. In part because of these unique characteristics, NMDA receptors are involved critically in synapse formation and modification during development (Bear et al., 1990; Cline et al., 1990; Iwasato et al., 2000; Ramoa et al., 2001; Erisir and Harris, 2003) as well as changes in synaptic strength in adulthood (Bliss and Collingridge, 1993; Heynen et al., 2000; Lisman and McIntyre, 2001). NMDA receptor dysfunction also has been implicated in many diseases such as epilepsy, schizophrenia, and several neurodegenerative disorders (Meldrum, 1992; Chapman, 2000; Cull-Candy et al., 2001; Tsai and Coyle, 2002; Zeron et al., 2002; Moghaddam, 2003).
Functional NMDA receptors are obligate heterotetramers, composed primarily of NR1 and NR2 subunits. The NR1 subunit has one gene product but many splice variants, which are expressed ubiquitously (Laurie et al., 1995). In contrast, expression of the four NR2 subunit gene products (NR2A–NR2D) shows tight temporal and spatial regulation. For example, NR2B and NR2D subunits are expressed prenatally, whereas NR2A and NR2C subunit expression rises sharply after birth (Monyer et al., 1994; Wenzel et al., 1996). Differential NR2 subunit expression also occurs across brain regions. In adulthood NR2A and NR2B subunits predominate within the cortex, whereas NR2C and NR2D subunits are found within midbrain and hindbrain structures (Monyer et al., 1994; Wenzel et al., 1996). NR2 subunits even show differential expression within individual neurons; NR2A subunits cluster around synapses, whereas NR2B subunits often occupy extrasynaptic sites (Tovar and Westbrook, 1999). NR2 subunit expression is controlled tightly because NR2 subunits help to determine many pharmacological and biophysical receptor properties, including external Mgo2+ (Mgo2+) affinity (Dingledine et al., 1999).
Because of voltage-dependent channel block by Mgo2+ (Mayer et al., 1984; Nowak et al., 1984; Ascher and Nowak, 1988), NMDA receptor-mediated currents are prominent only during periods of coincident glutamate release and postsynaptic depolarization. Recent reports have shown that, after rapid membrane depolarization, Mgo2+ unblock from native NMDA receptors contains fast (τ < 1 ms) and slow (τ = 3–20 ms) components (Spruston et al., 1995; Vargas-Caballero and Robinson, 2003; Kampa et al., 2004). These findings are surprising because the kinetics of Mgo2+ unblock at the single-channel level (Nowak et al., 1984; Ascher and Nowak, 1988) predict only rapid unblock. Instead, the slow component or components account for as much as 50% of the total current relaxation, which reduces NMDA receptor currents during the upstroke of action potentials (APs) (Vargas-Caballero and Robinson, 2003) and shortens the time window for spike timing-dependent plasticity (STDP) (Kampa et al., 2004).
Previous studies describing slow Mgo2+ unblock were performed on native NMDA receptors of an undefined subunit composition. Here, we set out to determine whether the properties of Mgo2+ unblock are NR2 subunit-dependent. We find that the relative speed of Mgo2+ unblock from NMDA receptor subtypes is represented by the following: NR1/2C, NR1/2D ≫> NR1/2A > NR1/2B. These NR2 subunit differences in Mgo2+ unblock render brief depolarizations, such as APs, more effective at stimulating Mgo2+ unblock from NR1/2A than from NR1/2B receptors.
Materials and Methods
Cell culture and transfection.
Human embryonic kidney (HEK) 293T cells were maintained as previously described (Qian et al., 2005). HEK 293T cells were plated onto untreated glass coverslips or onto glass coverslips pretreated with poly-d-lysine (0.1 mg/ml) and rat-tail collagen (0.1 mg/ml; BD Biosciences, San Jose, CA) in 35 mm culture dishes at 1–2 × 105 cells per dish. At 18–24 h after plating the cells were transiently transfected with cDNAs encoding the NR1-1a and one of the four NR2 subunits (NR2A–NR2D), using a modified Ca2+ precipitation procedure (Qian et al., 2005). The cDNA for enhanced green fluorescent protein (eGFP) was cotransfected as a marker of successful transfection. A total of 0.7 μg of eGFP, 1.3 μg of NR1-1a, and 2–8 μg of NR2A–NR2D cDNA were used per dish. After incubation of the cells with the transfection solution for 6–8 h, the precipitates were washed off with fresh culture medium that contained 200–1000 μm APV and 2 mm Mg2+. Experiments were performed 20–72 h after transfection.
Solutions
Solutions were prepared daily from frozen stocks. Currents were activated by the indicated concentration of NMDA or glutamate either in the absence of Mgo2+ or with 1 mm added Mgo2+. Glycine (10 μm) was added to all solutions. We did not adjust for changes in osmolality that resulted from the addition of Mgo2+. The external solution contained the following (in mm): 140 NaCl, 1 CaCl2, 2.8 KCl, and 10 HEPES, pH 7.2, adjusted with NaOH (osmolality, 290 ± 10 mmol/kg). The internal solution contained the following (in mm): 125 CsCl, 10 EGTA, and 10 HEPES, pH 7.2, adjusted with CsOH (osmolality, 275 ± 10 mmol/kg). Sucrose was used as needed to adjust the osmolality of the external solution. The junction potential between the pipette and bath solution was 5 mV, and all holding potentials were corrected for junction potentials. Ultrapure salts were used when available. All chemicals were from Sigma (St. Louis, MO).
Whole-cell recording.
Whole-cell recordings from transfected HEK 293T cells were performed as described previously (Qian et al., 2005). Briefly, pipettes were pulled from borosilicate standard walled glass with filaments (1.5 mm outer diameter; 0.86 mm inner diameter; Warner Instruments, Hamden, CT) and fire-polished to a resistance of 2–5 MΩ. Solutions were delivered with the use of an in-house fabricated fast perfusion system (Qian et al., 2002) connected to an eight chamber gravity-fed solution reservoir (AutoMate Scientific, San Francisco, CA). Solution exchanges were 90% complete within 20 ms for standard whole-cell experiments or 90% complete within 1 ms for the lifted cell experiments. Solution exchange measurements were made by recording whole-cell current from a transfected HEK 293T cell while moving between two barrels. Both barrels contained the same concentration of NMDA and glycine, but one barrel contained normal extracellular solution and the second barrel contained an extracellular solution with the impermeant ion N-methyl-d-glucamine (NMDG) in place of NaCl. The time course of current decrease after movement into the NMDG extracellular solution was used to estimate solution exchange times.
In some experiments the lifted cell technique (Vicini et al., 1998) was used to permit faster solution exchange times. For these experiments the HEK 293T cells were plated onto untreated glass coverslips. Once whole-cell access was obtained, negative pressure was reapplied, and the cell was lifted slowly into the solution flow. Small negative pressure was maintained throughout the experiment to prolong the duration of the experiment. The morphology of the cell was monitored continuously; if gross morphological changes occurred, the experiment was terminated.
All currents were recorded with an Axopatch 200 amplifier (Molecular Devices, Union City, CA) in voltage-clamp mode. The built-in series resistance correction and prediction circuitry were set to at least 80% in all experiments. Signals were low-pass filtered at 2.5 or 5 kHz (eight-pole Bessel; Warner Instruments) and sampled at 10–50 kHz. Where indicated, signals were refiltered at 1 kHz for display. All experiments were performed at room temperature.
Data analysis and curve fitting.
NMDA receptor-mediated current responses to depolarizing voltage jumps were corrected for leak and capacitive currents by subtracting the current response to an identical voltage jump in the absence of NMDA or glutamate, using pClamp 9.2 (Molecular Devices). When multiple sweeps of the same amplitude voltage jump were obtained, responses in the absence of NMDA or glutamate were averaged first and then subtracted from the agonist-induced ensemble average. Leak- and capacitive-subtracted currents (INMDA) then were fit with multi-exponential equations of the following form:  where C is the current level before the voltage jump and Ai is the amplitude of the exponential component, with time constant τi; n was adjusted between 1 and 3 as necessary to obtain quality fits. C, A, and τi were allowed to vary during fitting. The amplitudes were expressed as a percentage by dividing each Ai by the sum of the Ai values of all of the exponential components. In experiments in which voltage jumps to 35 mV were applied shortly after agonist application (see Fig. 7), currents were normalized before fitting to remove the effects of desensitization. Currents were normalized to the average of two responses (prevoltage jump and postvoltage jump) to agonist application while the cell was held at 35 mV to account for current rundown. All curve fitting was performed with Clampfit 9.2 (Molecular Devices) or Origin 7.0 (OriginLab, Northampton, MA). Data are expressed as the means ± SEM, and statistical analysis was performed by using Student's t tests unless otherwise noted.
where C is the current level before the voltage jump and Ai is the amplitude of the exponential component, with time constant τi; n was adjusted between 1 and 3 as necessary to obtain quality fits. C, A, and τi were allowed to vary during fitting. The amplitudes were expressed as a percentage by dividing each Ai by the sum of the Ai values of all of the exponential components. In experiments in which voltage jumps to 35 mV were applied shortly after agonist application (see Fig. 7), currents were normalized before fitting to remove the effects of desensitization. Currents were normalized to the average of two responses (prevoltage jump and postvoltage jump) to agonist application while the cell was held at 35 mV to account for current rundown. All curve fitting was performed with Clampfit 9.2 (Molecular Devices) or Origin 7.0 (OriginLab, Northampton, MA). Data are expressed as the means ± SEM, and statistical analysis was performed by using Student's t tests unless otherwise noted.
Results
NR2 subunit identity controls Mgo2+ unblocking kinetics
The rapid unblocking kinetics of Mgo2+ at the single-channel level (Nowak et al., 1984; Ascher and Nowak, 1988) predict that, during whole-cell recordings, membrane depolarization would induce rapid Mgo2+ unblock. However, recent data (Spruston et al., 1995; Vargas-Caballero and Robinson, 2003; Kampa et al., 2004) have shown that, after rapid membrane depolarization, Mgo2+ unblock from native NMDA receptors of an undefined subunit composition actually contains both fast (τ < 1 ms) and slow (τ = 3–20 ms) components. Because the identity of the NR2 subunit within a functional NMDA receptor influences many channel properties (Dingledine et al., 1999), we first investigated whether the kinetics of Mgo2+ unblock are also NR2 subunit-dependent. To obtain a homogenous population of NMDA receptors with a defined subunit composition, we cotransfected HEK 293T cells with the cDNA for the NR1-1a subunit and the cDNA for one of the four NR2 subunits (NR2A–NR2D). HEK 293T cells are well suited for these experiments because the cells are electrotonically compact, which eliminates any slow changes in current because of poor space clamp, as may be seen with neuronal cells (Vargas-Caballero and Robinson, 2003).
We first applied depolarizing voltage steps from −65 mV during long applications (15 s) of NMDA in the absence and presence of Mgo2+ (Fig. 1A,B). Long applications of NMDA allowed whole-cell currents to reach a steady-state level despite varying degrees of desensitization among NMDA subtypes (Dingledine et al., 1999). In the presence of physiological concentrations of Mgo2+ (1 mm), block of the NR1/2A and NR1/2B receptor-mediated currents was nearly complete at −65 mV (94.7 ± 0.6 and 96.1 ± 1.3%, respectively), whereas NR2C and NR2D receptors showed substantially less block (78.6 ± 0.4 and 77.9 ± 2.0%, respectively). This is consistent with previous reports (Monyer et al., 1994; Kuner and Schoepfer, 1996; Qian et al., 2005) showing that NR1/2C and NR1/2D receptors have a lower affinity for Mgo2+ than do NR1/2A and NR1/2B receptors.

Protocol used to investigate the kinetics of Mgo2+ unblock from recombinant NMDA receptors during steady-state response. NMDA (30 μm) and glycine (10 μm) (termed NMDA) was applied (top trace) in the absence (A) and presence (B) of 1 mm added Mgo2+ during standard whole-cell recordings from an HEK 293T cell expressing NR1/2B receptors. Once the current (bottom trace) had reached a stationary level, three depolarizing voltage jumps (here, from −65 to 35 mV; middle trace) were applied. After termination of the NMDA application, another identical set of depolarizing voltage jumps was applied to allow for off-line leak and capacitive subtraction. All three sweeps in the absence and presence of NMDA were averaged before subtraction. Current traces were refiltered at 1 kHz for display.
A depolarizing voltage jump to 35 mV relieves the voltage-dependent block by Mgo2+ from all receptor subtypes and allows outward current flow. However, in response to a depolarizing voltage step, outward currents in the presence of 1 mm Mgo2+ from both NR1/A and NR1/2B receptors develop with both fast and slow components (Fig. 2A2,B2). The observed slow component is predominantly attributable to Mgo2+ unblock, because the same voltage jump performed in the absence of Mgo2+ results in a more rapid relaxation of outward current (Fig. 2A1,B1). Strikingly, after an identical depolarizing voltage jump, Mgo2+ unblocks from NR1/2C and NR1/2D receptors very rapidly, with no obvious slow component (Fig. 2C2,D2). The kinetics of outward currents from NR1/2C and NR1/2D receptors after a depolarizing voltage jump are also rapid in the absence of Mgo2+ (Fig. 2C1,D1). Thus, unlike NR1/2A and NR1/2B receptors, under these experimental conditions there is no slow Mgo2+ unblock from NR1/2C or NR1/2D receptors.

The kinetics of Mgo2+ unblock are NR2 subunit dependent. Whole-cell currents are shown from NR1/2A (A1, A2), NR1/2B (B1, B2), NR1/2C (C1, C2), and NR1/2D (D1, D2) receptors during a 500 ms depolarizing voltage step from −65 to 35 mV. The voltage steps were applied either in the absence (A1–D1) or in the presence of 1 mm Mgo2+ (A2–D2). A prominent slow relaxation of the outward current is present within records from NR1/2A and NR1/2B receptors when the depolarizing voltage jump is applied in the presence of 1 mm Mgo2+, indicating a slow component of Mgo2+ unblock. In contrast, the outward current from NR1/2C and NR1/2D receptors in response to a depolarizing voltage jump relaxes with a single, fast component both in the absence and presence of Mgo2+. NMDA indicates that the currents are recorded in the presence of 30 μm NMDA and 10 μm glycine. The timing of the voltage jump is indicated by the traces above A1 and A2, and the 0 current level is indicated by a horizontal dashed line. Double (A1–B2) or single (C1–D2) exponential fits are overlaid (gray lines). Note the different calibration in B1 and B2 because of the potentiation of NR1/2B receptors by Mgo2+ (Paoletti et al., 1995).
To quantify the kinetics of Mgo2+ unblock, we fit currents in response to voltage jumps from −65 to 35 mV with a single or double exponential equation (Fig. 2). In the absence of added Mgo2+ the current relaxation from all receptor subtypes was dominated by a single fast (sub-millisecond) component. However, we did observe a small slow component of outward current relaxation in the absence of added Mgo2+ from both NR1/2A and NR1/2B receptors (Fig. 2A1,B1). Slow relaxation of NMDA-mediated current after depolarizing voltage jumps in the absence of Mgo2+ has been reported previously (Benveniste and Mayer, 1995; Spruston et al., 1995) (but see Vargas-Caballero and Robinson, 2003; Kampa et al., 2004). In our hands the kinetics of current relaxation after a depolarizing voltage jump to 35 mV with zero-added Mgo2+ are dominated by a fast (sub-millisecond) component, with the slow component having a small percentage of amplitude (<15%) that in every case is significantly (p < 0.01) less than the amplitude of the slow component in the presence of Mgo2+ (supplemental Table 1, available at www.jneurosci.org as supplemental material).
In the presence of Mgo2+ the NR1/2A and NR1/2B receptor currents were well fit by a double exponential equation, whereas a single exponential equation provided excellent fits of currents from NR1/2C and NR1/2D receptors both in the presence and absence of Mgo2+ (Fig. 2). The tau of the fast component (τ1) did not differ significantly among any of the receptor subtypes or depend on the presence of Mgo2+, with a value of ∼0.5 ms under all conditions (supplemental Table 1, available at www.jneurosci.org as supplemental material). The tau of the slow component (τ2) of Mgo2+ unblock from NR1/2A and NR1/2B receptors was significantly (p < 0.0001) slower than τ1, with values of 4.80 ± 0.39 ms for NR1/2A (n = 6) and 9.15 ± 0.83 ms for NR1/2B receptors (n = 8). In addition, τ2 was significantly (p < 0.005) slower from NR1/2B than from NR1/2A receptors (see below). In both NR1/2A and NR1/2B receptors the slow component accounted for ∼40% of the total current.
It is possible that we missed a small slow component of Mgo2+ unblock from NR1/2C and NR1/2D receptors because they are blocked less effectively by 1 mm Mgo2+ than are NR1/2A and NR1/2B receptors (Monyer et al., 1994; Kuner and Schoepfer, 1996; Qian et al., 2005). To test this possibility, we raised Mgo2+ to 5 mm, a concentration at which the inhibition of NR1/2D receptor responses is comparable to inhibition of NR1/2B receptor responses by 1 mm Mgo2+. Even under these conditions there was no hint of a slow component of Mgo2+ unblock from NR1/2D receptors (Fig. 3). The kinetics of Mgo2+ unblock from NR1/2D receptors were similar in the presence of 1 and 5 mm Mgo2+, with time constants of 0.25 ± 0.01 and 0.30 ± 0.07 ms, respectively. The kinetics of Mgo2+ unblock from NR1/2A and NR1/2B receptors in the presence of 5 mm also were similar to those observed in the presence of 1 mm Mgo2+ (supplemental Table 1, available at www.jneurosci.org as supplemental material). Thus the kinetics of Mgo2+ unblock are determined by the identity of the NR2 subunit, with slow Mgo2+ unblock occurring only from NMDA receptors that contain either the NR2A or NR2B subunit.

Mgo2+ unblocks rapidly from NR2D receptors even at high [Mgo2+]. Shown are superimposed whole-cell currents during a voltage jump from −65 to 35 mV from NR1/2B receptors in the presence of 30 μm NMDA, 10 μm glycine, and 1 mm Mgo2+ (thin trace) or NR1/2D receptors in the presence of 30 μm NMDA, 10 μm glycine, and 5 mm Mgo2+ (thick trace). Currents are normalized to the steady-state outward current at 35 mV. Note the comparable level of inward current mediated by each of the two receptor isoforms. Voltage change is indicated by the top trace; the 0 current level is indicated by a dashed horizontal line.
Voltage dependence of slow Mgo2+ unblock
To characterize further the slow Mgo2+ unblock from NR1/2A and NR1/2B receptors, we applied depolarizing voltage jumps from rest to test voltages from −45 to 35 mV, using a protocol similar to that shown in Figure 1. The resulting currents were fit with single or double exponential equations. In the presence of 1 mm Mgo2+ a double exponential equation was required for adequate fits of currents in response to voltage jumps to each of the tested voltages, indicating that slow Mgo2+ unblock occurs throughout the physiological range of membrane voltages.
For both NR1/2B and NR1/2A receptors τ1 did not depend on the amplitude of the depolarization (p > 0.05; ANOVA), which enabled us to average the results across all test voltages. The averaged values for τ1 did not depend significantly on NR2 subunit identity or the presence of Mgo2+, with values of 0.40 ± 0.11 ms (n = 16) and 0.31 ± 0.02 ms (n = 19) for NR1/2A receptors and 0.52 ± 0.23 ms (n = 19) and 0.38 ± 0.13 ms (n = 15) for NR1/2B receptors in the presence and absence of Mgo2+, respectively. In contrast, τ2 from NR1/2B receptors was significantly slower than τ2 from NR1/2A receptors at most test voltages (Fig. 4A). Only in response to the smallest amplitude voltage jump (−65 to −45 mV) was τ2 not significantly different between NR1/2A and NR1/2B receptors. τ2 from both NR1/2A and NR1/2B receptors did show weak voltage dependence (p < 0.05; ANOVA), becoming slower as the amplitude of the depolarizing step increased (Fig. 4). The amplitudes of the slow component (Aslow) of Mgo2+ unblock from NR1/2A and NR1/2B receptors were not significantly different (Fig. 4B), carrying ∼40% of the total depolarization-induced current relaxation at all test voltages. Aslow did not display any significant voltage dependence (p > 0.05; ANOVA) in either NR1/2A or NR1/2B receptors, although there was a trend for Aslow to increase as the amplitude of the voltage jump increased.
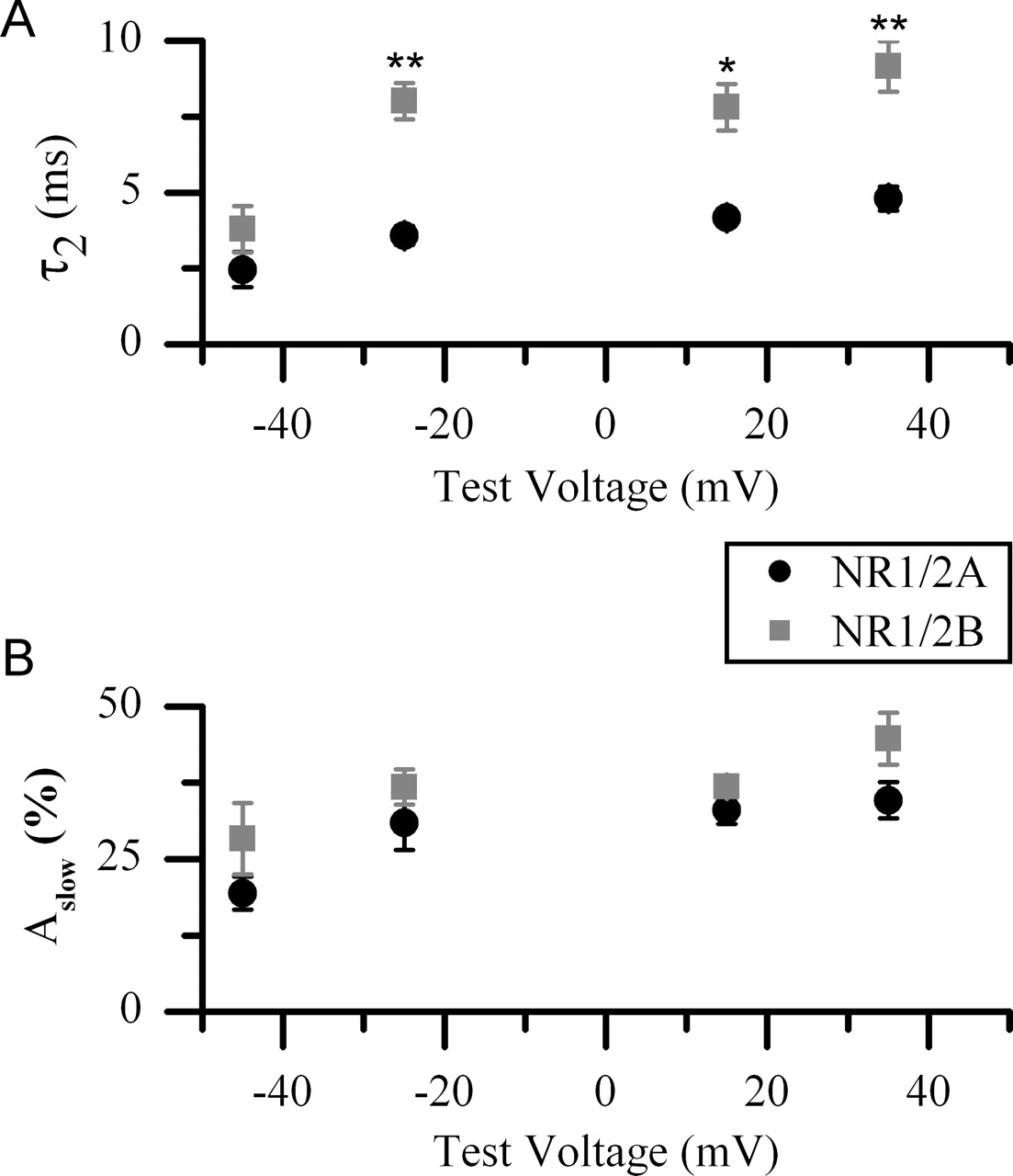
Quantification of the subunit and voltage dependence of Mgo2+ unblocking kinetics. Shown are results from double-exponential fits to NR1/2A (black circles) and NR1/2B (gray squares) receptor currents in response to a depolarizing voltage jump from rest (−65 mV) to the indicated test voltages in the presence of 30 μm NMDA, 10 μm glycine, and 1 mm Mgo2+. A, Pooled results of τ2. τ2 from fits to NR1/2B receptor currents was significantly slower than τ2 from fits to NR1/2A receptor currents at all test voltages except −45 mV. τ2 of both NR1/2A and NR1/2B receptors also showed weak voltage dependence, becoming significantly slower as the amplitude of the depolarizing voltage jump became larger. B, Pooled results of Aslow. Aslow did not differ significantly between NR1/2A and NR1/2B receptors and did not show any significant voltage dependence. Error bars indicate the means ± SEM. ∗p < 0.01; ∗∗p < 0.005.
If the slow component of Mgo2+ unblock resulted from slow unbinding of Mgo2+ from open NMDA receptor channels, τ2 should be strongly voltage-dependent and become faster with depolarization. Instead, we found that slow Mgo2+ unblock is only weakly voltage-dependent and becomes slower with depolarization (Fig. 4A), in agreement with previous data from native NMDA receptors (Vargas-Caballero and Robinson, 2003). Thus it is likely that the kinetics of slow Mgo2+ unblock arise from the slow exit of NMDA receptor channels from one or more closed block state(s) (see Discussion).
In the presence Mgo2+, returning the voltage to −65 mV caused rapid reblock of the channel (Fig. 5A,B). However, after depolarizations to −25, 15, and 35 mV a slow phase of Mgo2+ block was also present (Fig. 5A,B). In cells with large enough responses to allow adequate signal-to-noise ratios, we fit the slow portion of Mgo2+ reblock with a single exponential equation. The slow phase of Mgo2+ reblock of NR1/2B receptors was significantly slower than the slow phase of Mgo2+ reblock of NR1/2A receptors after repolarization to −65 mV from −25, 15, and 35 mV (Fig. 5C). In contrast, currents from NR1/2C and NR1/2D receptors showed very rapid reblock, which was well fit with a single exponential component with a τ of <1 ms (data not shown). Together, these data indicate that the kinetics of both Mgo2+ unblock and reblock during rapid voltage changes are strongly dependent on the identity of the NR2 subunit.

Slow reblock of Mgo2+. Shown are example traces from NR1/2A (A) and NR1/2B (B) receptors that follow a voltage return to −65 mV after a 500 ms depolarization to 35 mV in the presence of 30 μm NMDA, 10 μm glycine, and 1 mm Mgo2+. The majority of Mgo2+ reblock occurs in <1 ms (indicated by rapid Mgo2+ reblock). However, a slow phase of Mgo2+ reblock is present, which delays the return to baseline current (indicated by dashed black line). Insets, Single exponential fits of the slow phase of Mgo2+ reblock are overlaid as gray lines. C, Pooled results from fits to the slow phase of Mgo2+ reblock. The time constant (τ) of slow Mgo2+ reblock of NR1/2B receptors (gray squares) is significantly slower than the τ of slow Mgo2+ reblock of NR1/2A receptors (black circles) after repolarization to −65 mV from −25, 15, or 35 mV. Error bars indicate the means ± SEM. ∗p < 0.05; ∗∗p < 0.01.
Dependence of Mgo2+ unblocking kinetics on agonist concentration
Previous experiments investigating slow Mgo2+ unblock have disagreed as to the time course of slow unblock, varying from a single slow component with a τ of 14–23 ms (Vargas-Caballero and Robinson, 2003) to two slow components, one with a τ of ∼4 ms and one with a τ > 100 ms (Kampa et al., 2004). One prominent difference between these previous studies is the concentration of agonist used to activate NMDA receptors; Vargas-Caballero and Robinson (2003) used 25 μm NMDA, whereas Kampa and colleagues (2004) used 1 mm glutamate. To determine whether the kinetics of Mgo2+ unblock are dependent on the agonist concentration used to activate the receptors, we repeated the protocol described in Figure 1, using 1 mm glutamate or NMDA. Again, in response to a voltage step from −65 to 35 mV the currents from NR1/2A and NR1/2B receptors displayed a prominent slow component of Mgo2+ unblock (Fig. 6A,B). In contrast, Mgo2+ unblock from NR1/2C and NR1/2D receptors was dominated by a single fast component (τ < 1 ms) (supplemental Table 2, available at www.jneurosci.org as supplemental material), as observed previously in lower agonist conditions.

Kinetics of Mgo2+ unblock in high agonist concentration. A, B, Examples of the current response to a depolarizing voltage jump (from −65 to 35 mV) in the presence of 1 mm glutamate and 10 μm glycine (termed Glu) and 1 mm Mgo2+ from NR1/2A (A) or NR1/2B (B) receptors. C, Examples of double-(top) and triple-(bottom) exponential fits to NR1/2B receptor-mediated currents in response to the voltage jump. A triple-exponential equation also was required to obtain adequate fits of NR1/2A receptor-mediated currents (data not shown). The 0 current level is indicated by a horizontal dashed line. D, E, Pooled results from triple-exponential fits of whole-cell current responses to a depolarizing voltage jump from NR1/2A (n = 4; black bars) and NR1/2B (n = 4; gray bars) receptors. The kinetics of the prominent, intermediate slow component (τ2) of Mgo2+ unblock are significantly slower and the relative amplitude significantly larger from NR1/2B than from NR1/2A receptors. Error bars indicate the means ± SEM. ∗p < 0.01; ∗∗p < 0.0005.
Unlike results using 30 μm NMDA, Mgo2+ unblock from NR1/2A and NR1/2B receptors in these high agonist concentrations required a triple exponential equation for an adequate fit (Fig. 6C). The tau of the fast component (τ1) of Mgo2+ unblock did not differ significantly between NR1/2A and NR1/2B receptors (Fig. 6D) or depend on the agonist concentration. In contrast, the tau of the dominant slow component (τ2) of Mgo2+ unblock from NR1/2A receptors was significantly (p < 0.005) faster in 1 mm glutamate as compared with the corresponding τ2 of Mgo2+ unblock in the presence of 30 μm NMDA. This was also true for the τ2 of Mgo2+ unblock from NR1/2B receptors. It seems reasonable to compare these two components because the third component observed only in high agonist conditions was the same in NR1/2A and NR1/2B receptors, had a time constant (τ3) of several hundred milliseconds, and had a small (<10%) amplitude (Fig. 6D,E). Despite showing faster kinetics at high agonist concentrations, the τ2 of Mgo2+ unblock was still NR2 subunit-dependent; NR1/2B receptor τ2 was significantly slower and had a significantly larger amplitude than the corresponding τ2 from NR1/2A receptors (Fig. 6D,E).
For both NR1/2A and NR1/2B receptors the main difference between the kinetics of Mgo2+ unblock in 30 μm NMDA versus 1 mm glutamate was an acceleration of the prominent slow component of unblock (termed τ2 in both agonist conditions). This could be an agonist-specific difference, but this is unlikely because similar results to those shown with 1 mm glutamate were obtained by using 1 mm NMDA (supplemental Table 2, available at www.jneurosci.org as supplemental material). Instead, this difference may result from an increase in channel open probability (Popen) that results from increasing agonist concentration. The hypothesis that a larger Popen leads to a faster τ2 of Mgo2+ unblock is consistent with our observation that τ2 is faster from NR1/2A than from NR1/2B receptors (Fig. 4); NR1/2A receptors have a higher Popen than NR1/2B receptors (Chen et al., 1999; Erreger et al., 2005).
On the basis of this hypothesis, any manipulation that raises the Popen of the receptor should accelerate the slow component of Mgo2+ unblock. To test this hypothesis, we repeated the protocol in Figure 1 with NR1/2B receptors at pH 8.2, which reduces proton inhibition (Giffard et al., 1990; Tang et al., 1990; Traynelis and Cull-Candy, 1990) and hence raises Popen. Under these conditions τ2 was significantly (p = 0.02) accelerated from 9.15 ± 0.93 ms at pH 7.2 to 3.93 ± 0.77 ms at pH 8.2. Aslow also was reduced from 44.7 ± 4.8% at pH 7.2 to 32.7 ± 1.3% at pH 8.2, although this difference did not reach statistical significance. These data support the hypothesis that the differences in Mgo2+ unblock from NR1/2A and NR1/2B receptors are attributable to differences in Popen.
Effect of the timing of membrane depolarization on Mgo2+ unblocking kinetics
The relative timing of synaptic NMDA receptor activation and postsynaptic depolarization plays a critical role in determining whether Ca2+ influx via NMDA receptors will lead to long-term changes in synaptic strength (Magee and Johnston, 1997; Markram et al., 1997; Bi and Poo, 1998). Previous work has shown that the kinetics of Mgo2+ unblock from native NMDA receptors also depend on the relative timing of receptor activation and postsynaptic depolarization; unblock is slower after prolonged receptor activation (Kampa et al., 2004). We next determined whether the kinetics of Mgo2+ unblock are NR2 subunit-dependent when unblock is stimulated shortly after receptor activation.
For these experiments we used lifted HEK 293T cells, which allowed for rapid exchange of the external solution (90% complete in <1 ms). To verify that glutamate application to lifted HEK 293T cells transfected with NR1/2A or NR1/2B receptors was rapid, we measured the rise time of NMDA currents. The 10–90% rise times of currents activated by 1 mm glutamate in the presence of 1 mm Mgo2+ at a constant membrane potential of −65 mV was, for NR1/2A receptors, 4.87 ± 0.61 ms (n = 6) and, for NR1/2B receptors, 9.95 ± 1.39 ms (n = 5). These are similar to previously published values for the rise times of NR1/2A and NR1/2B receptor currents in the absence of Mgo2+ (Chen et al., 1999; Erreger et al., 2005).
In the presence of 1 mm Mgo2+ the rapid glutamate applications at 35 mV resulted in large outward currents, whereas current responses to glutamate applications at −65 mV mainly were blocked in both NR1/2A and NR1/2B receptors (Fig. 7A,B). Voltage jumps from −65 to 35 mV applied after a ∼15 ms delay that followed glutamate application resulted in outward currents that relaxed toward the current observed during glutamate application with the cell held at 35 mV. However, the current in response to the voltage jump did not immediately join the current during the glutamate application at 35 mV, indicating a slow component of Mgo2+ unblock (Fig. 7A,B). The voltage jump was time-locked to barrel movement signal, which resulted in some jitter between receptor activation and membrane depolarization because of variability in the distance between the barrels and the lifted cell. However, the average timing between receptor activation and membrane depolarization was similar in NR1/2A (15.0 ms) and NR1/2B (14.3 ms) receptors.

Mgo2+ unblock near the time of onset of NMDA receptor activity. Shown are example traces during a rapid application of 1 mm glutamate in the presence of 1 mm Mgo2+ while the cell was held either at 35 mV (thin trace) or during a voltage jump from −65 to 35 mV (thick trace) from NR1/2A receptors (A) and NR1/2B receptors (B). The timing of the voltage jump and the barrel movement are indicated by the middle and bottom traces, respectively. Current traces were refiltered at 1 kHz for display.
To quantify the kinetics of Mgo2+ unblock, we normalized the current in response to the voltage jump to the current evoked by glutamate application while the cell was held at 35 mV. The normalized currents in response to these voltage jumps that occurred shortly after receptor activation were well fit with a double exponential equation. A triple exponential equation was not needed because a very slow component (τ of several hundred milliseconds), which was observed when a prolonged application of 1 mm glutamate preceded the voltage jump, was absent. The time constant of the fast component (τ1) of unblock did not differ significantly between NR1/2A and NR1/2B receptors, with values of 0.22 ± 0.04 and 0.24 ± 0.06 ms, respectively. In contrast, the time constant of the slow component (τ2) of Mgo2+ unblock from NR1/2B receptors, 3.69 ± 0.38 ms, was significantly (p < 0.01) slower than from NR1/2A receptors, 2.13 ± 0.27 ms. In these experiments the amplitude of the slow component was similar for NR1/2A and NR1/2B receptors, with values of 32.2 ± 8.3 and 44.1 ± 7.8%, respectively. Thus our data suggest that the kinetics of Mgo2+ unblock depend slightly on the timing between receptor activation and membrane depolarization, with a very slow component (τ > 100 ms) occurring only after prolonged receptor activation. However, in all situations Mgo2+ unblock proceeds more rapidly from NR1/2A than from NR1/2B receptors.
Effect of varying the duration of depolarization on Mgo2+ unblock
Ca2+ influx via NMDA receptors plays a critical role in many physiological processes. The kinetics of Mgo2+ unblock during membrane depolarization determine how many NMDA receptors become unblocked and hence are available to pass inward current after membrane repolarization. The repolarization-induced peak of inward current (Ipeak) is of particular interest because it is the point at which the largest NMDA receptor-mediated Ca2+ influx occurs (Garaschuk et al., 1996). During brief depolarizations slow Mgo2+ unblock should reduce the fraction of NMDA receptors from which Mgo2+ unblocks and thus the fraction of receptors available to pass inward current during repolarization. In addition, brief depolarizations should be less effective at stimulating Mgo2+ unblock from NR1/2B receptors (from which Mgo2+ unblock occurs more slowly) than from NR1/2A receptors. Thus the Ipeak after brief depolarizations should be reduced as compared with the Ipeak after long depolarizations, and the reduction should be greater for NR1/2B than for NR1/2A receptors. To test these hypotheses, we applied rapid voltage jumps from rest to 35 mV of varying duration (1, 2, 5, 10, 50 ms) in the presence of 1 mm each glutamate and Mgo2+. Because there is not a strong effect of the timing between receptor activation and membrane depolarization on the kinetics of Mgo2+ unblock (see above), depolarizations were applied during prolonged agonist application (similar to Fig. 1). This allowed multiple voltage jumps to be applied during a single agonist application. Depolarizations were separated by 100 ms intervals, which is >10-fold longer than the τ of the slow component of Mgo2+ reblock, allowing steady-state block to be reached between each depolarization.
As expected, longer duration depolarizations resulted in a larger Ipeak (Fig. 8A). To quantify the relationship between depolarization duration and Ipeak, we normalized all Ipeak values to the Ipeak after a 50 ms depolarization. In experiments from both NR1/2A and NR1/2B receptors the 1 ms depolarizations induced an Ipeak that was <70% of the Ipeak after a 50 ms depolarization (Fig. 8B). Only after the depolarization duration was increased to 10 ms was the Ipeak not significantly smaller than the Ipeak after a 50 ms depolarization. In addition, after depolarizations of <10 ms, the normalized Ipeak from NR1/2A receptors was significantly larger than the normalized Ipeak from NR1/2B receptors (Fig. 8B). Thus slow Mgo2+ unblock blunts inward currents via NR1/2A and NR1/2B receptors after brief depolarizations. However, brief depolarizations (<10 ms) are able to stimulate more Mgo2+ unblock from NR1/2A than from NR1/2B receptors.

Brief depolarizations are more effective at stimulating Mgo2+ unblock from NR1/2A than from NR1/2B receptors. A, Superimposed currents from NR1/2A (black) and NR1/2B (red) receptors (bottom traces) during depolarizations of varying duration (1, 2, 5, 10, and 50 ms; indicated by top traces) in the presence of 1 mm glutamate, 10 μm glycine, and 1 mm Mgo2+. Currents are normalized to the Ipeak value after a 50 ms depolarization. The value of Ipeak after the 50 ms depolarization is indicated by a dotted line. Inset, Enlargement of the normalized inward peaks in response to the briefest depolarizations (1, 2, and 5 ms). B, Comparison of the Ipeak values after depolarizations of varying duration. Results are from NR1/2A (n = 5; black) and from NR1/2B (n = 4; red) receptors. Ipeak values were normalized to the Ipeak after a 50 ms depolarization. Error bars indicate the means ± SEM; ∗∗p < 0.0005. C, Superimposed currents (bottom traces) from NR1/2A (black) and NR1/2B (red) receptors during three APs (somatic, 5 ms extended, and 25 ms extended; top traces) in the presence of 1 mm glutamate, 10 μm glycine, and 1 mm Mgo2+. Traces are normalized to the Ipeak,AP induced by the 25 ms extended AP. D, Pooled ratios of Ipeak,AP values in response to the somatic or 5 ms extended APs normalized to the Ipeak,AP value in response to the 25 ms extended AP. Results are from NR1/2A (black bars) and NR1/2B (red bars) receptors. Error bars indicate the means ± SEM; ∗p < 0.01.
Although square voltage jumps are easily applied experimentally, physiological voltage changes occur with slower rising and falling phases. Recently, there has been a strong interest in back-propagating APs (b-APs) invading the dendritic tree and providing the depolarization needed for Mgo2+ unblock of NMDA receptors during synaptic activity (Magee and Johnston, 1997; Markram et al., 1997; Bi and Poo, 1998). Within dendrites near the soma the b-APs are rapid, often having a half-width of <2 ms. Farther out in the dendritic tree the b-AP half-width can be >10 ms (Bernard and Johnston, 2003). In all instances the rising and falling phases of b-AP waveforms are slower than the square pulses we have used above.
To test whether the NR2 subunit-dependent differences in Mgo2+ unblocking kinetics also impact NMDA receptor activity during physiological depolarizations, we voltage-clamped transfected HEK 293T cells to AP waveforms. Three waveforms were used: an unmodified AP recorded from the soma of a hippocampal CA1 pyramidal cell (termed somatic AP) and two APs that we modified so the duration at the most depolarized voltage was 5 ms (5 ms extended AP) or 25 ms (25 ms extended AP). The ability of each waveform to generate inward current via NMDA receptors after Mgo2+ unblock was quantified by measuring the peak inward current during the repolarizing phase of the APs (Ipeak,AP). The 25 ms extended AP should give maximum Mgo2+ unblock from both NR1/2A and NR1/2B receptors. Thus Ipeak,AP induced by the somatic AP and by the 5 ms extended AP was normalized to Ipeak,AP induced by the 25 ms extended AP. The normalized values of Ipeak,AP for the 5 ms AP were close to 1 for both NR1/2A and NR1/2B receptors (Fig. 8C,D). This suggests that the 5 and 25 ms extended APs induced a similar amount of Mgo2+ unblock. In contrast, the normalized values of Ipeak,AP for the somatic AP were significantly (p < 0.001) <1 for both NR1/2A and NR1/2B receptors (Fig. 8C,D). Thus the somatic AP was too brief to stimulate full Mgo2+ unblock from either NR1/2A or NR1/2B receptors. The normalized value of Ipeak,AP for the somatic AP was also significantly smaller for NR1/2A than for NR1/2B receptors (Fig. 8D). This suggests that somatic APs are more effective at stimulating Mgo2+ unblock from NR1/2A than from NR1/2B receptors. Thus during synaptic activity postsynaptic APs coincident with presynaptic glutamate release are more effective at stimulating Mgo2+ unblock from NR1/2A than from NR1/2B receptors.
Discussion
Many pharmacological and biophysical properties of NMDA receptors are, at least in part, determined by the identity of the NR2 subunit (Dingledine et al., 1999). Our results show that Mgo2+ unblocking kinetics are also NR2 subunit-dependent. Mgo2+ unblocks from NR1/2A and NR1/2B receptors with a prominent slow component that is not present when Mgo2+ unblocks from NR1/2C or NR1/2D receptors (Fig. 2). In addition, the slow component of Mgo2+ unblock from NR1/2A and NR1/2B receptors is not equivalent; Mgo2+ unblocks from NR1/2B receptors more slowly than from NR1/2A receptors (Fig. 4). These NR2 subunit differences in Mgo2+ unblock render brief depolarizations, including APs, more effective at stimulating Mgo2+ unblock from NR1/2A than from NR1/2B receptors (Fig. 8).
Comparison with previous studies
Our data are consistent with previous studies describing slow Mgo2+ unblock from native hippocampal and cortical NMDA receptors (Spruston et al., 1995; Vargas-Caballero and Robinson, 2003; Kampa et al., 2004) in that the hippocampus and cortex predominantly express the NR2A and NR2B subunits (Monyer et al., 1994). Our data also provide potential explanations for discrepancies between previous studies of the kinetics of Mgo2+ unblock. When depolarizations occurred during prolonged receptor activation, Vargas-Caballero and Robinson (2003) reported that slow Mgo2+ unblock had a single component (τs1 > 10 ms), whereas Kampa and colleagues (2004) reported that slow Mgo2+ unblock contained two components (τs1 ∼4 and τs2 ∼300 ms). One experimental difference between these two studies is the concentration of agonists used to activate NMDA receptors. Vargas-Caballero and Robinson (2003) used 25 μm NMDA, whereas Kampa and colleagues (2004) used 1 mm glutamate. We find that at high agonist concentrations the main component of slow Mgo2+ unblock is accelerated (from τ2 between 5 and 9 ms to a t2 between 2 and 5 ms), and an additional very slow component (τ = 200–400 ms) is discernible (Fig. 6). It is also likely that the τs1 reported by Vargas-Caballero and Robinson (2003) was slower than the τs1 reported by Kampa and colleagues (2004), because Vargas-Caballero and Robinson (2003) used younger animals than Kampa and colleagues (2004), which should express a higher percentage of NR1/2B receptors.
The additional very slow component of Mgo2+ unblock is not present when depolarizations occur shortly after agonist application (Fig. 7). This is in agreement with Kampa and colleagues (2004) and shows that the kinetics of Mgo2+ unblock depend on the relative timing of receptor activation and membrane depolarization. However, after a prolonged glutamate application, we find that the very slow component of Mgo2+ unblock accounts for <10% of the total current rather than ∼30% of the total current reported previously (Kampa et al., 2004). The source of this discrepancy is not clear. Resolution of this conflict is important because changes in the kinetics of Mgo2+ unblock with prolonged receptor activation influence the time window for STDP induction (Kampa et al., 2004).
Several previously published models of NMDA receptor function (Jahr and Stevens, 1990; Antonov and Johnson, 1999; Sobolevsky and Yelshansky, 2000) do not predict a slow component of Mgo2+ unblock in response to a depolarizing voltage jump (data not shown). In order to explain the surprising observation of slow Mgo2+ unblock, two models have been proposed (Kampa et al., 2004; Vargas-Caballero and Robinson, 2004). These models suggest that occupation of the channel by Mgo2+ moderately accelerates only the rate of channel closure (Vargas-Caballero and Robinson, 2004) or accelerates rates of channel closure and agonist unbinding, along with increasing the occupancy of desensitized states (Kampa et al., 2004). These models predict accurately the appearance of slow component or components of Mgo2+ unlock. They also are supported by the observed reduction in single-channel burst duration of cortical NMDA receptors in the presence of Mgo2+ (Ascher and Nowak, 1988), as would be predicted if Mgo2+ accelerates channel closure. These models, however, are not fully consistent with other observations. If Mgo2+ accelerates channel closure, the IC50 for Mgo2+ inhibition of whole-cell currents should be lower than the Mgo2+ KD calculated from single-channel measurements, a prediction that disagrees with previous measurements (Qian et al., 2002). Additional relevant data involve the organic NMDA receptor channel blocker amantadine, which unblocks from open channels even faster than Mgo2+. Burst analysis demonstrates that amantadine does accelerate channel closure moderately (Blanpied et al., 2005). After a depolarizing voltage step, amantadine, like Mgo2+, exhibits a slow component of channel unblock. However, the slow component of unblock is much slower for amantadine than for Mgo2+ (Blanpied et al., 2005). These data suggest that a complete understanding of the mechanistic basis of the slow component of Mgo2+ unblock awaits additional research.
We also have described here a slow component of Mgo2+ reblock of NR1/2A and NR1/2B receptors (Fig. 5). This contrasts with previous studies (Spruston et al., 1995; Vargas-Caballero and Robinson, 2003; Kampa et al., 2004) that report near-instantaneous Mgo2+ reblock of native NMDA receptors. Because the slow phase of Mgo2+ reblock observed here is small, it is possible that it was not observed in previous studies that used nucleated patches (Spruston et al., 1995; Vargas-Caballero and Robinson, 2003; Kampa et al., 2004), which typically yield relatively small currents. We were able to quantify slow Mgo2+ reblock only from whole-cell records that had large currents (>100 pA) at −65 mV in the presence of 1 mm Mgo2+. Although of small amplitude, the slow phase of Mgo2+ reblock provides a relatively long window during which highly significant Ca2+ influx could occur after a postsynaptic AP that coincides with synaptic input.
We have studied NMDA receptors containing one of the four NR2 subunits. However, there is evidence that triheteromeric receptors, which contain more than one type of NR2 subunit, exist within many brain regions, including the cortex (Sheng et al., 1994; Didier et al., 1995; Chazot and Stephenson, 1997; Luo et al., 1997), cerebellum (Chazot et al., 1994; Cathala et al., 2000; Brickley et al., 2003), and substantia nigra (Jones and Gibb, 2005). Coexpression of the NR1, NR2A, and NR2D subunits in Xenopus oocytes yields a receptor with many characteristics intermediate between NR1/2A and NR1/2D receptors (Cheffings and Colquhoun, 2000). It is tempting to speculate that triheteromeric receptors would show intermediate Mgo2+ unblocking kinetics. However, testing this hypothesis would be challenging because it is difficult to isolate triheteromeric receptors within heterologous systems (Vicini et al., 1998).
Implications for synaptic plasticity
Ca2+ influx via NMDA receptors during coincident synaptic activity and postsynaptic depolarization underlies long-term changes in synaptic strength at many synapses (Bliss and Collingridge, 1993). However, not all NMDA receptors are equivalent. Because NR1/2C and NR1/2D receptors have a lower affinity for Mgo2+ (Monyer et al., 1994; Kuner and Schoepfer, 1996; Qian et al., 2005), they allow significant Ca2+ influx near typical resting membrane potentials where NR1/2A and NR1/2B receptors are >95% blocked. In addition, we show here that the relatively weak Mgo2+ block of NR1/2C and NR1/2D receptor currents is relieved more rapidly after depolarization than is the Mgo2+ block of NR1/2A and NR1/2B receptor currents (Fig. 2). Thus Mgo2+ block has a stronger influence on NR1/2A and NR1/2B receptor currents than NR1/2C and NR1/2D receptor currents, rendering NR1/2A and NR1/2B receptors more effective coincidence detectors.
Several recent studies have suggested that selective NR1/2A receptor activation leads preferentially to long-term potentiation (LTP), whereas selective NR1/2B receptor activation leads to long-term depression (LTD) (Liu et al., 2004; Massey et al., 2004; Kim et al., 2005; Mallon et al., 2005). Based on proposed differences in the Ca2+ threshold for LTP and LTD induction (Bienenstock et al., 1982; Cormier et al., 2001), our data suggest a mechanism that could help to differentiate the physiological impact of NR1/2A and NR1/2B receptor activation. In response to brief membrane depolarizations, the relatively faster unblock of Mgo2+ from NR1/2A receptors would result in greater Ca2+ influx (Fig. 8), favoring LTP, whereas the relatively slower unblock of Mgo2+ from NR1/2B receptors would result in lower Ca2+ influx, favoring LTD. It should be noted, however, that it is unlikely that there is an exclusive association of NR1/2A receptors with LTP and of NR1/2B receptors with LTD (Tang et al., 1999; Berberich et al., 2005; Toyoda et al., 2005; Weitlauf et al., 2005; Neyton and Paoletti, 2006).
In many studies of the NR2 subunit dependence of synaptic plasticity, induction protocols that involve the pairing of presynaptic stimulation with prolonged postsynaptic depolarization are used. Such induction protocols reduce the relevance of the kinetics of Mgo2+ unblock. We also expect slow Mgo2+ unblock to have limited impact during relatively small synaptic depolarizations: slow unblock time constants are faster, amplitudes are smaller, and NR2 subunit-dependent differences are smaller at more hyperpolarized potentials (Fig. 4). The kinetics of Mgo2+ unblock are most clearly of critical importance when plasticity is induced with STDP protocols, which pair synaptic input with postsynaptic APs (Magee and Johnston, 1997; Markram et al., 1997; Bi and Poo, 1998). The NR2 subunit differences in Mgo2+ unblock kinetics reported here should help to determine the magnitude of Ca2+ influx mediated by NR1/2A and NR1/2B receptors in response to brief, large-amplitude depolarizations such as APs.
Footnotes
-
This work was supported by National Institute of Mental Health Grants MH045817 (J.W.J.) and T32 MH18273 (R.C.). We thank Xixi Chen and Dan Johnston for the somatic AP waveform.
- Correspondence should be addressed to Jon W. Johnson, Department of Neuroscience, 446 Crawford Hall, University of Pittsburgh, Pittsburgh, PA 15260. Email: johnson{at}bns.pitt.edu





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}