

Article Figures & Data
Figures
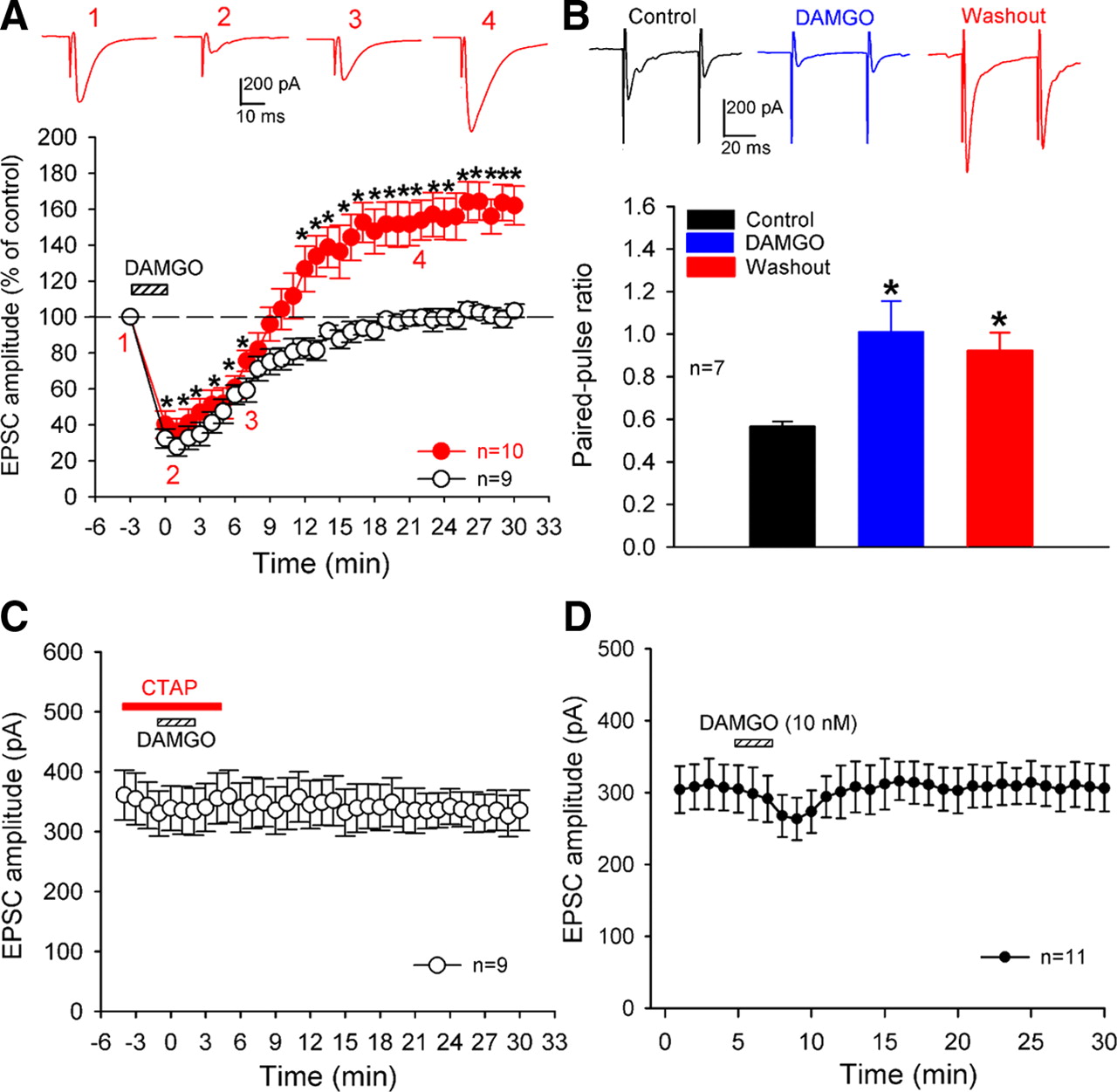
- Figure 1.
Brief μ-opioid receptor activation induces presynaptic LTP in lamina II neurons. A, Mean time courses of 10 lamina II neurons in which bath application of 1 μm DAMGO for 3 min produced initial inhibition and LTP upon washout (red solid circles) and nine neurons in which DAMGO produced acute depression only (black open circles). Insets show individual EPSC traces of one neuron recorded at the time points indicated. B, DAMGO-induced acute depression and LTP were associated with an increased paired-pulse ratio (n = 7 cells). Insets show individual traces of one neuron recorded during the control period, DAMGO application, and 20 min after DAMGO washout. C, Bath application of CTAP (1 μm) blocked the DAMGO-induced initial inhibition and LTP in nine lamina II neurons. D, Mean time courses of 11 lamina II neurons where bath application of 10 nm DAMGO for 3 min produced only a small inhibition of monosynaptic EPSCs evoked from the dorsal root but did not induce LTP upon washout. Data are presented as mean ± SEM. *p < 0.05 compared with the pre-DAMGO control value.
- Figure 2.
DAMGO induces LTP from primary afferent terminals in the spinal cord. A, Blocking the postsynaptic opioid action with GDP-β-S and Cs2+ failed to block LTP induction by DAMGO in nine lamina II neurons without affecting the initial depression (black solid circles). In another nine neurons, DAMGO produced only acute depression (black open circles). Insets show individual EPSC traces of one neuron recorded at the time points indicated. *p < 0.05 compared with the pre-DAMGO control value. B, Representative traces show that blocking of postsynaptic opioid signaling with intracellular dialysis of 1 mm GDP-β-S abolished the DAMGO-induced outward GIRK currents elicited by bath application of 1 μm DAMGO in the lamina II neuron.
- Figure 3.
Opioid-induced LTP in lamina I neurons originates from primary afferent terminals in the spinal cord. A, Mean time courses of 11 lamina I neurons in which bath application of 1 μm DAMGO for 3 min produced acute inhibition and LTP upon washout (red solid circles) and 10 neurons in which DAMGO produced acute depression only (black open circles). B, Blocking the postsynaptic opioid signaling with GDP-β-S and Cs2+ failed to block the DAMGO-induced LTP in 12 lamina I neurons without affecting the initial depression (red solid circles). In another 10 neurons, DAMGO produced acute depression only (black open circles). Insets show individual EPSC traces of one neuron recorded at the time points indicated. Data are presented as mean ± SEM. *p < 0.05 compared with the pre-DAMGO control value.
- Figure 4.
Opioid-induced presynaptic LTP originates from TRPV1-expressing primary afferents in the spinal cord. A, Treatment with the ultrapotent TRPV1 agonist RTX ablates dorsal root ganglion neurons. Representative confocal images from one RTX-treated and one vehicle control rat show TRPV1-immunoreactive and Griffonia simplicifolia isolectin B4 (IB4, a marker of unmyelinated neurons)-positive neurons in the dorsal root ganglion. Colocalization of TRPV1 immunoreactivity and IB4 labeling is indicated in yellow when the two images are digitally merged. All images are single confocal optical sections. B, RTX treatment not only abolished LTP but also prolonged inhibition of the amplitude of evoked monosynaptic EPSCs by DAMGO in all of the 16 lamina II neurons tested. Insets show individuals traces of evoked EPSCs of one neuron recorded at the time points indicated. Data are presented as mean ± SEM. *p < 0.05 compared with the pre-DAMGO control value.
- Figure 5.
Opioid-induced synaptic glutamate release originates from TRPV1-expressing primary afferent terminals. A, Mean time courses of nine lamina II neurons where bath application of 1 μm DAMGO for 3 min produced acute inhibition followed by potentiation of the frequency of mEPSCs upon washout (black solid circles) and another nine neurons in which DAMGO acutely inhibited mEPSCs only (black open circles). B, RTX treatment not only abolished LTP but also prolonged inhibition of the frequency of mEPSCs by DAMGO in all of the 16 lamina II neurons tested. Insets show individuals traces of mEPSCs of one neuron recorded at the time points indicated. Data are presented as mean ± SEM. *p < 0.05 compared with the pre-DAMGO control value.
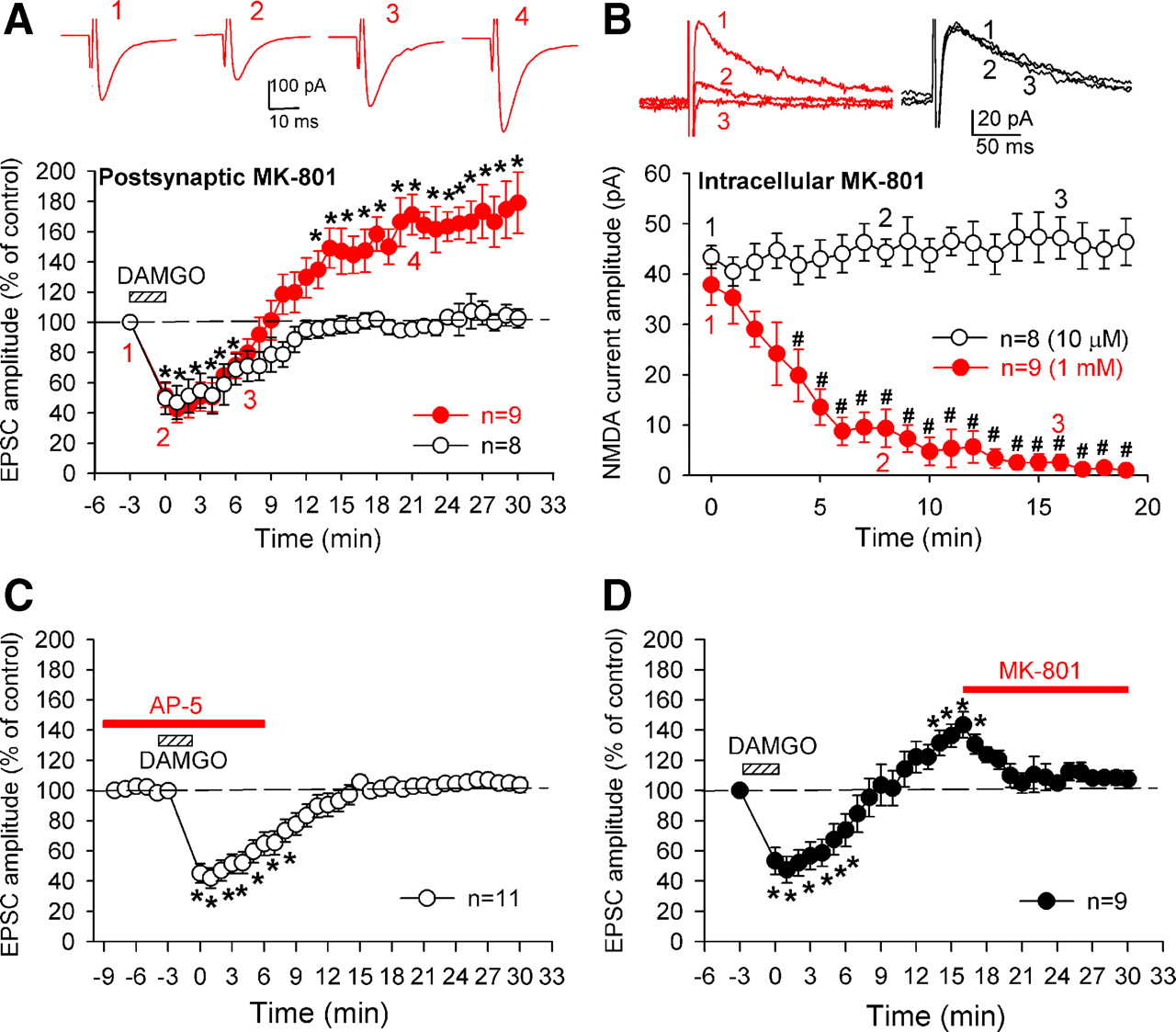
- Figure 6.
Presynaptic NMDA receptors contribute to opioid-induced LTP in the spinal cord. A, Postsynaptic dialysis with 1 mm MK-801 failed to block LTP induction by 1 μm DAMGO in nine lamina II neurons (red solid circles). Insets show individual EPSC traces of one neuron recorded at the time points indicated. B, Mean time courses of lamina II neurons where intracellular dialysis of 1 mm (n = 9 neurons, red solid circles), but not 10 μm (n = 8 neurons, black open circles), of MK-801 blocked postsynaptic NMDA currents evoked from the dorsal root. Insets show individual NMDA current traces of neurons recorded at the time points indicated. C, Bath application of 50 μm AP-5 before DAMGO application blocked LTP induction without affecting the initial depression by DAMGO. D, Bath application of 10 μm MK-801 during DAMGO-induced LTP reversed LTP in nine lamina II neurons. Data are presented as mean ± SEM. *p < 0.05 compared with the pre-DAMGO control value; #p < 0.05 compared with the control value at time 0.
- Figure 7.
Increased intracellular calcium is involved in opioid-induced presynaptic LTP in the spinal cord. A, Postsynaptic dialysis with 30 mm BAPTA failed to block DAMGO (1 μm)-induced LTP in nine lamina II neurons (black solid circles). DAMGO produced acute inhibition only in another eight neurons dialyzed with BAPTA (black open circles). B, Bath application of 50 μm BAPTA-AM abolished DAMGO-induced LTP in 18 lamina II neurons without affecting the initial depression. Insets show individual monosynaptic EPSC traces of one neuron recorded at the time points indicated. Data are presented as mean ± SEM. *p < 0.05 compared with the pre-DAMGO control value.











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}