

Abstract
Trace conditioning is a form of classical conditioning, where a neutral stimulus (conditioned stimulus, CS) is associated with a following appetitive or aversive stimulus (unconditioned stimulus, US). Unlike classical delay conditioning, in trace conditioning there is a stimulus-free gap between CS and US, and thus a poststimulus neural representation (trace) of the CS is required to bridge the gap until its association with the US. The properties of such stimulus traces are not well understood, nor are their underlying physiological mechanisms. Using behavioral and physiological approaches, we studied appetitive olfactory trace conditioning in honeybees. We found that single-odor presentation created a trace containing information about odor identity. This trace conveyed odor information about the initial stimulus and was robust against interference by other odors. Memory acquisition decreased with increasing CS–US gap length. The maximum learnable CS–US gap length could be extended by previous trace-conditioning experience. Furthermore, acquisition improved when an additional odor was presented during the CS–US gap. Using calcium imaging, we tested whether projection neurons in the primary olfactory brain area, the antennal lobe, contain a CS trace. We found odor-specific persistent responses after stimulus offset. These post-odor responses, however, did not encode the CS trace, and perceived odor quality could be predicted by the initial but not by the post-odor response. Our data suggest that olfactory trace conditioning is a less reflexive form of learning than classical delay conditioning, indicating that odor traces might involve higher-level cognitive processes.
Introduction
Animals can react to sensory stimuli either instantaneously or with a certain delay. Thus, sensory systems must keep a neural representation of a stimulus after its termination (i.e., a stimulus trace). Such stimulus traces can be revealed behaviorally by trace conditioning (Pavlov, 1927). In trace conditioning, a neutral conditioned stimulus (CS) is associated with a following meaningful, unconditioned stimulus (US). Unlike standard Pavlovian conditioning in which CS and US overlap, in trace conditioning the US occurs during a stimulus-free gap after the CS has terminated. This slight difference in temporal CS–US relation is thought to make trace and delay conditioning two fundamentally different phenomena. They differ with regard to the involved brain areas, as trace, but not delay, conditioning depends on the hippocampus (Solomon et al., 1986; Cheng et al., 2008). Moreover, trace, but not delay, conditioning is thought to represent higher-level cognitive processes, as it induces a form of declarative memory (Clark and Squire, 1998; Lovibond and Shanks, 2002). However, a mechanistic understanding of trace conditioning is still lacking (Christian and Thompson, 2003; Woodruff-Pak and Disterhoft, 2008). We therefore aimed to investigate the properties of stimulus traces and trace memory acquisition by studying appetitive olfactory trace conditioning in honeybees. Insects are important model organisms for studying the neural basis of learning and memory (Menzel, 2001; Giurfa, 2007; Gerber et al., 2009). Honeybees rapidly learn to associate the scent of flowers with food, and bees' odor–reward learning can easily be accessed by classical delay and trace conditioning of the appetitive proboscis extension reflex (Kuwabara, 1957; Bitterman et al., 1983). Physiological measurements of brain activity allow us to relate behavior to brain activity (Guerrieri et al., 2005). In the insect brain, the primary olfactory areas are the antennal lobes, which are structurally and functionally similar to the mammalian olfactory bulbs (Hildebrand and Shepherd, 1997); both are made up of glomeruli (Mori et al., 1999). The output neurons of the antennal lobe are projection neurons (PN; analogous to the vertebrate mitral/tufted cells), which send axons to higher brain areas such as the mushroom bodies (Menzel et al., 2005). Odor responses have been precisely characterized in insects (Galizia and Szyszka, 2008), but post-odor responses—a putative neural substrate for odor traces—are less well understood. We conducted a comprehensive analysis of trace conditioning in honeybees and searched for a neural substrate of odor traces within the antennal lobe by optically imaging PNs.
Here we show that odor traces decay in time, but can be extended by experience. Furthermore, bees, unlike mammals, form a trace after a single trace-conditioning event. Finally, we show that calcium activity in PNs, and hence spiking activity, does not represent the trace substrate. Rather, the trace must be searched for in other second messengers or in other brain areas, such as the mushroom bodies. These results open a fascinating new window into the diversity of memory processes in one of the most intelligent mini-brains, that of the honeybee.
Materials and Methods
Animals.
Experiments were performed with honeybee foragers (Apis mellifera). In winter, bees were taken from hives kept indoors with controlled temperature (16°C at night and 25°C during the day), light/dark cycle (12/12 h), and humidity (∼75%). In summer they were taken from outside hives.
Odor stimulation.
We used 1-nonanol, 1-octanol, 1-hexanol, 1-nonanone, 2-heptanone, octanal, and citral for odor stimulation (all from Sigm-Aldrich). The pure odorants were diluted to 10−2 in mineral oil (Sigma-Aldrich). Odorant dilutions were prepared freshly every 4 weeks. One or two hundred microliters of each odor were loaded onto a cellulose strip (Sugi, REF 31003; Kettenbach) located in a 3 ml syringe (Norm-Ject; Henke-Sass, Wolf), adjusted to 2.5 ml. Syringes were prepared every day. Odor stimuli were delivered as 0.5 or 6 s pulses with a custom-built computer-controlled olfactometer (Fig. 1A). The olfactometer was constructed to produce odor stimuli with highly controlled and reproducible dynamics. The olfactometer produced nearly rectangular odor pulses with steep odor onsets and offsets, as measured using a photoionization detector (Model 200a; Aurora Scientific) (Vetter et al., 2006) (Fig. 1B). Continuous air suction behind the bee cleared residual odor. Calcium responses in PNs confirmed the absence of contamination, as the initial responses to a 0.5-s-long odor pulse terminated within 0.7 ± 0.15 s after stimulus offset (mean ± SEM).

An olfactometer for highly controlled stimulus dynamics. A, Schematic of the olfactometer. Olfactometers were equipped with either three or six odor channels. Each odor channel consisted of two 3 ml syringes, one empty for equalizing air flow and one containing the odorant. One hundred or two hundred microliters of each odor were loaded onto a cellulose strip. Syringes were air supplied via a manifold that was designed to reduce cross-contamination to a minimum. The manifold (a) was a Teflon-septum equipped glass vial (20 ml headspace vial; Schmidlin Labor and Service), which was connected via injection needles (b; 1.20 × 40 mm, Sterican; Braun). The needles served as non-return valves. The volume of the glass vial reduced the flow speed and thus reduced the vacuum, which would suck back odor-loaded air from the syringe. The air stream through each channel was 150 (experiments in Fig. 3A–C) or 300 ml/min (all other experiments), each controlled by a flowmeter (Analyt-MTC). In each channel, a magnetic three-way solenoid valve (LFAA1200118H; Lee) controlled the odor pulses by diverting air from the empty syringe to the odorant syringe. The valves were switched with a spike-and-hold driver circuit to minimize opening time. Odors were injected into a continuous carrier air stream in a glass tube (k; 0.7 cm in diameter). A Teflon ring (l) inside the glass tube ensured complete mixing of the odor with the carrier air. The carrier air stream was adjusted to a total air stream of 3000 ml/min (1.3 m/s) and directed at the bee positioned ∼2 cm in front of the glass tube. B, The olfactometer was adjusted to produce odor pulses as rectangular as possible with steep odor onsets and offsets. The dynamic of the odor stimulus was measured with a photoionization detector. The detector was placed 20 mm in front of the olfactometer within the behavioral setup. One hundred microliters of 2-heptanone were used as tracer substance. The analog photoionization detector signal (0–10 V) was digitized at a sampling rate of 1 kHz. Traces represent mean ± SD of six (0.5 s stimulus) and 12 (6 s stimulus) measurements. Measurements were repeated every 20 s. ID, Inner diameter; PID, photoionization detector.
Conditioning.
The day before conditioning, bees were caught, chilled on ice until they were immobilized, fixed in plastic tubes, and fed until satiation. They were kept in a moist box overnight. The next day, bees were tested for proboscis extension reflex (PER) by touching the antenna with a metal preparation needle soaked with sucrose solution (1 m in water). Only bees showing the PER were used for conditioning 10 min later. Bees received one or six training trials with the conditioned odor (CS). The intertrial interval was 10 min. Bees were moved in front of the olfactometer 15 or 20 s before odor stimulation to habituate them to the airflow. The CS was paired with sucrose stimulation (US) to the antenna and proboscis lasting for 3 s. In delay conditioning, CS and US overlapped, whereas in trace conditioning CS and US were separated by a stimulus-free gap (Fig. 2Ai). Apart from the experiments shown in Figure 3, Aii and Cii, the CS duration was equal during training and test. A proboscis extension reflex was counted as a conditioned response when the proboscis was extended horizontally in response to the odor stimulus. Memory retrieval was tested 15 min, 30 min, or 24 h after the last training trial. The intertrial interval was again 10 min and the sequence of CS and control odor stimulation was pseudorandomized. Whenever the response to CS and a new odor were compared (Figs. 3A–C, 4), hexanol and nonanol were equally often used as CS and new odor, and the sequence of CS and new odor stimulation was balanced. Whenever the effect of different experimental conditions was compared, the corresponding experiments were conducted in parallel.
Staining and preparation.
Each imaging experiment took 2 d. On the first day, bees were harnessed in Plexiglas stages by gluing their eyes to the stage with hard wax (Deiberit 502; Dr. Böhme und Schöps Dental). The mandibles were pushed open to each side and fixed to the stages. The antennae were fixed temporarily with n-eicosan (Sigma-Aldrich Chemie) to avoid touching them during the preparation. A window was cut into the head capsule and glands and tracheae were carefully removed. When necessary, a drop of Ringer's solution (in mm: 130 NaCl, 7 CaCl2, 6 KCl, 2 MgCl2, 160 saccharose, 25 glucose, 10 HEPES, pH 6.7, 500 mOsmol) was added to prevent the brain from drying. The PNs were stained with the calcium-sensitive dye Fura-2 dextran (Invitrogen) by inserting two dye-coated glass needles between the calyces of the mushroom bodies in each brain hemisphere (Fig. 5A). The head capsule was closed, the antennae were freed, and bees were fed with a 1 m sucrose solution until satiation and kept in a moist box until the next day (14–16 h). Then, the antennae were fixed pointing frontwards. The tracheae above the antennal lobes were removed carefully. To reduce movements, the esophagus and the surrounding muscles were lifted through a small opening in the clypeus and fixed with two-component silicon (Kwik-Sil; World Precision Instruments), and thorax and abdomen were gently pushed against the stage with a piece of foam (1 × 1 × 3 cm). The brain was covered with a thin layer of transparent Kwik-Sil. Within 10–30 min after the preparation, bees were put under the microscope and heated to ∼28°C with an infrared lamp. The antennal lobe that showed better staining and the stronger spontaneous activity was chosen for measuring.
Imaging.
PNs in the antennal lobe were imaged through a water-immersion objective (20×, numerical aperture 0.95; Olympus). The imaging system consisted of a fluorescence microscope (BX-50WI; Olympus), a light source (Polychrome IV; Till Photonics), and a CCD camera (Imago QE; Till Photonics). Images were binned on-chip, resulting in a resolution of 172 × 130 pixels (441 × 333 μm). Each recording lasted 29 s and consisted of 232 double frames recorded with 340 and 380 nm excitation light at a rate of 8 Hz. Excitation and emission light were separated with a 420 nm dichroic mirror and a 490–530 nm emission filter. Bees were stimulated with 1-nonanol, 1-octanol, 1-hexanol, 1-nonanone, 2-heptanone, octanal, and by touching the proboscis with a solution of 1 m sucrose. Odors were presented in a pseudorandomized order with an intertrial interval of 3 min. Odor stimulation was controlled by the acquisition software (Till Vision; Till Photonics).
Data analysis.
Imaging data were analyzed using custom-written programs in IDL (RSI). First, measurements were movement-corrected by aligning frames within and between measurements. Then, glomeruli were segmented with the help of an unsharp masked image of the raw fluorescence and a correlation image where the correlation of the signal traces between neighboring pixels was calculated (Fig. 5B). Glomeruli were identified by comparing glomerulus position and size with the morphological atlas of the honeybee (Galizia et al., 1999a) and the odor responses with the physiological atlas (Galizia et al., 1999b; Sachse et al., 1999). Signals were calculated as F340/F380. The baseline was shifted to 0 by subtracting the average signal before stimulation (frame 4, 69 for odor stimulation and 109 for sucrose stimulation). Activity patterns are shown as color-coded images of single measurements, which were filtered with a spatial low-pass filter of 5 × 5 pixels for better visualization (Fig. 5B). No filtering was used for quantitative analysis. Glomerulus responses were calculated from a 5 × 5 pixels square in the glomerulus center. Similarity between two odor responses was quantified by calculating Pearson's correlation between the two glomerular odor–response vectors (Figs. 5, 6), which can range from −1 (anticorrelated) to 1 (perfectly correlated).
Behavioral data were analyzed with an ANOVA. Although parametric ANOVA test is usually not allowed for dichotomous data, an empirical study has shown that ANOVA is appropriate under our conditions (Lunney, 1970). Statistical tests were performed with R (www.r-project.org), and Sigma Stat (SPSS).
Results
We performed trace-conditioning experiments in honeybees with odor as CS and sucrose reward as US to characterize two conceptually distinct aspects of trace conditioning: the properties of acquisition and retrieval of associative CS–US memories (trace memory), and the properties of the nonassociative CS memories themselves (trace). Learning and memory were assessed by conditioning the appetitive proboscis extension reflex. Moreover, we asked whether persistent odor responses in PNs, measured as calcium responses, may underlay odor traces.
Trace-memory acquisition decreases with increasing CS–US intervals
We first compared memory formation between trace and delay conditioning. We used a 3-s-long US either preceded by a 6-s-long CS (delay conditioning, CS and US overlap) or a 0.5-s-long CS (trace conditioning, CS and US separated by a gap) (Fig. 2Ai). In both groups, the CS–US interval, defined as the time between the CS onset and US onset, was 5 s (Fig. 2Aii). For trace conditioning, the proboscis extension rate increased from the first to the second training trial and then stagnated on a plateau at ∼33%. For delay conditioning, the proboscis extension rate increased from the first to the fourth training trial and reached an asymptotic level of ∼72%. Thirty minutes later, delay-conditioned bees showed the conditioned response nearly twice as often as trace-conditioned bees (70% vs 39%). Thus, trace conditioning is less efficient than delay conditioning.

Odor-trace memory formation. Ai, Experimental protocol used to compare memory acquisition between delay and trace conditioning. In delay conditioning, there is a temporal overlap between the CS and US. In trace conditioning, there is a stimulus-free gap between the CS and US. The onset between the CS and the US (CS–US interval) was always 5 s. Each bee received six training trials. Aii, Two groups of bees were trained with delay and trace conditioning (CS was nonanol). Percentage of bees showing the CS-evoked proboscis extension reflex within 5 s after odor onset during training and in a memory retrieval test 30 min after the last training trial. Proboscis extension rate in delay-conditioned bees reached a plateau at ∼72% after the third trial (one-way repeated-measurement ANOVA; Ftrial(5, 460) = 27.6, p < 0.001; Holm-Sidak post hoc tests; N = 93). Proboscis extension rate in trace-conditioned bees reached a plateau at ∼33% after the first trial (one-way repeated-measurement ANOVA; Ftrial(5, 545) = 5.4, p < 0.001; Holm-Sidak post hoc tests; N = 110). Different letters indicate significant differences. During the memory-retrieval test, the proboscis extension rate was higher in the delay-conditioning group than in the trace-conditioning group (t test, p < 0.001). Bi, Eight groups were conditioned in parallel with different intervals between the onset of the CS (octanol or heptanone) and the US stimulus. CS and US were presented either simultaneously (0 s), with a CS–US interval of 1, 2, 3, 6, 10, or 15 s (trace conditioning), or the CS was presented 6 s after the US (backward conditioning, −6 s). Bii, Memory retrieval was tested 15 min after the last training trial. Asterisks indicate a significant increase in proboscis extension rate compared with the spontaneous proboscis extension rate during odor presentation in the first training trial before sucrose stimulation (paired t tests, p < 0.05). At CS–US intervals of −6 and 0, the spontaneous proboscis extension rate could not be determined because the sucrose was presented before or together with the odor. Memory performance decayed with increasing CS–US intervals. The longest CS–US interval that produced memory was 6 s. Biii, Memory was tested in the same bees after 15 min (short-term memory) and again after 24 h (mid-term memory). These bees are a subset of the bees in Bii. In both memory tests, the longest effective CS–US interval was again 6 s. The CS–US interval functions did not differ for the 15 min and 24 h memory test (two-way repeated-measurement ANOVA with CS–US interval and test time as factors; FCS–US interval(9, 275) = 9.9, p < 0.001; Ftest time(1, 275) = 0.35, p = 0.6; FCS–US interval × test time(9, 275) = 1.5, p = 0.14). C, The perceived quality of an odor was compared between delay and trace conditioning (both groups were conditioned in parallel). Odor quality was measured by means of a generalization test. Each bee received six training trials with nonanol as CS. Left, Training protocols. Right, Memory test 15 min after training. Only bees that responded to the CS in the memory test were analyzed. Generalization differed between odors, but there was no difference between the generalization profiles after delay and trace conditioning (two-way ANOVA with odor and trace vs delay as factors; Fodor(4, 530) = 31.8, p < 0.001; Ftrace vs delay(1, 530) = 2.9, p = 0.09, Fodor × trace vs delay(4, 530) = 0.8, p = 0.53). Different letters mark significant differences between odors. n.s., Not significant.
We next determined the dependency of associative learning on the CS–US interval. During training, a 0.5-s-long CS was presented either simultaneously with the US (0 s CS–US interval), with a CS–US interval of 1, 2, 3, 6, 10, or 15 s (trace conditioning), or 6 s after the US (backward conditioning, −6 s) (Fig. 2Bi). Memory retrieval was tested 15 min after the sixth training trial (Fig. 2Bii). Simultaneous conditioning was most effective, with a 69% proboscis-extension rate. Memory decreased with increasing CS–US intervals (1–15 s) and was significant for all intervals up to 6 s. No bee responded to the CS in the backward-conditioning group (−6 s), indicating inhibitory learning.
Trace and delay conditioning share basic properties
The difference in memory acquisition between trace and delay conditioning may implicate different neural substrates of memory formation. In honeybees, the neural substrates for short-term and long-term memory formation differ (Müller, 2002). Thus, if the neural substrates for memory formation differ between trace and delay conditioning, then trace and delay conditioning may also differ with respect to short-term and long-term memory formation. We therefore compared memory after 15 min (short-term memory) and after 24 h (early long-term memory) (Fig. 2Biii). We found no differences between the CS–US interval functions in the 15 min and 24 h memory retrieval tests, and even the weak trace memory acquired at a 6 s CS–US interval was transformed into a long-term trace memory.
After olfactory conditioning, animals do not respond to the CS only, but they generalize to novel (odor) stimuli (Linster and Hasselmo, 1999; Guerrieri et al., 2005). Typically, generalization increases with perceived similarity. Thus, generalization can be used as a tool to measure the perceived quality of a CS in relation to other test odors. If trace and delay conditioning engage different brain areas, as is the case in mammals (Woodruff-Pak and Disterhoft, 2008), one would expect differences in the perceived odor quality between trace and delay conditioning, because neural odor representations change over time (Galán et al., 2004) and along different processing levels of the olfactory pathway (Linster et al., 2005; Szyszka et al., 2005). We therefore asked whether the perceived odor quality differs between trace memory and delay conditioning (Fig. 2C). We found no differences in the generalization profiles after trace and delay conditioning, indicating that an odor trace memory contains the same molecular identity information as the odor memory formed during delay conditioning.
A single CS–US pairing is sufficient for trace conditioning
Even though CS and US occurrence are non-overlapping, forming an association in the brain needs a temporal coincidence of CS and US representation in the neural network. This coincidence could be established in the brain by two nonexclusive mechanisms (Fig. 3Ai): by maintaining an odor representation (CS trace) or by shifting the US representation in time (US anticipation). In the latter case, bees should not learn after a single CS–US pairing, because US anticipation requires previous experience that a CS is followed by an US (Schultz et al., 1997). We therefore performed one-trial trace conditioning with a CS–US interval of 5 s (Fig. 3Aii). During the memory test, 30 min after training, the CS and a new odor were presented sequentially to the same bees. We used hexanol and nonanol equally often as CS and as new odor. The new odor served as a control for nonassociative effects of the training procedure. We found that a single CS–US trace conditioning event was sufficient to form an associative memory, showing that US anticipation is not necessary for trace conditioning and odor traces exist already after a first CS presentation.

The initial part of an odor stimulus induces an odor trace. Ai, Two possible mechanisms for establishing the overlap between the neural CS and US representation necessary for associative CS–US learning: Top, The CS leaves a sensory trace that overlaps with the US (CS trace). Bottom, In the course of repeated CS–US pairings, the neural US representation shifts backward in time (US anticipation). Aii, One-trial trace conditioning. Memory retrieval was tested 30 min after training by presenting a 6-s-long CS and new odor sequentially to the same bees. Trace conditioning worked after a single CS–US pairing with a 0.5-s-long CS and a 4.5-s-long gap between CS offset and US onset (one-way repeated-measurement ANOVA, Fodor(51, 102) = 19.3, p < 0.001). Thus, odor traces were present during the first CS presentation and US anticipation is not necessary for trace conditioning. B, Trace conditioning did not work with a 6-s-long CS and the same 4.5 s gap (one-way repeated-measurement ANOVA, Fodor(50, 100) = 2.2, p = 0.11). Thus, the late phase of a continuing odor stimulus does not induce an odor trace. This experiment was performed in parallel with the one in Aii. C, Identical procedure as in Aii and B, but tests used 0.5 s stimuli instead of 6 s stimuli to ensure that test-stimulus length did not influence the result. The conclusion was identical: trace conditioning did not occur for long odor stimuli if the gap between odor onset and US onset became too long. Ci, Trace conditioning worked after a single CS–US pairing with a 0.5-s-long CS and a 4.5-s-long gap between CS offset and US onset (one-way repeated-measurement ANOVA, Fodor(96, 192) = 18.8, p < 0.001). Cii, Trace conditioning did not work with a 6-s-long CS and the same 4.5 s gap (one-way repeated-measurement ANOVA, Fodor(96, 192) = 2.4, p = 0.09). D, The conditioned response occurred later after trace conditioning than after simultaneous conditioning, but earlier than the predicted reward. Each bee received six training trials with nonanol as CS. Left, Training protocols. Right, Memory test 15 min after training (simultaneous conditioning, 1.3 ± 0.1 s; trace conditioning, 2 ± 0.1 s, mean ± SEM; t test, p < 0.001).
Does an odor trace bridge the stimulus-free time gap between CS offset and US onset or does it bridge the interval between CS onset and US onset (CS–US interval)? We compared memory acquisition in bees that were trace conditioned with a 0.5-s-long CS and a 5 s CS–US interval (Fig. 3Aii) with bees that were conditioned with a 6-s-long CS and a 10.5 s CS–US interval (Fig. 3B). The stimulus-free gap was 4.5 s in both groups. Bees of the 5 s CS–US interval group acquired a trace memory, whereas bees of the 10.5 s CS–US interval group did not. Odor stimulus length during testing did not influence this result (Fig. 3C). Thus, the initial part of an odor stimulus induces an odor trace, whereas the late phase of a continuing odor stimulus does not.
In classical conditioning, animals often not only learn what to respond to, but also when to respond (Gormezano and Moore, 1969). Accordingly, the peak amplitude of a conditioned response often occurs at the expected US onset time (Pavlov, 1927; Ebel and Prokasy, 1963). We asked whether honeybees also learn when to extend their proboscis, i.e., whether their response delay corresponds to the timing of the CS–US interval in trace conditioning (Fig. 3D). Bees exhibited conditioned responses later after trace conditioning than after simultaneous conditioning, but earlier than the predicted reward. Thus, the odor trace memory either did not contain information about the CS–US timing or timing information was not behaviorally relevant.
The CS–US interval can be extended by experience
It is known from trace autoshaping experiments in pigeons (Lucas et al., 1981), visual trace conditioning in honeybees (R. Menzel, personal communication), and olfactory trace conditioning in Drosophila (Galili et al., 2011) that the maximum learnable CS–US interval is not fixed, but can be extended by experience. We therefore asked whether the maximum learnable CS–US interval of trace conditioning is experience-dependent too. We measured 10 s trace (CS10) memory in bees that were trained with a 5 s trace conditioning (CS5) 10 min before (Fig. 4Ai) and compared this to bees that experienced the reversed order (Fig. 4Aii). Hexanol and nonanol were equally often used as CS5 and CS10 during the test, and each bee received a different odor for CS5 and CS10. We found that bees with previous CS5 experience learned the CS10-trace task (Fig. 4Ai). In contrast, bees that received the 10 s trace conditioning before the 5 s trace conditioning did not learn the CS10 task (Fig. 4Aii). Thus, experience of a single trace-conditioning trial extended the learnable CS–US interval in subsequent trace conditioning, meaning that the maximum learnable CS–US interval is not fixed but is experience-dependent. A simultaneous conditioning (CS0) was not sufficient to extend the permissive gap (Fig. 4B). Thus, it is specifically the experience of a trace-conditioning event that extends the effective CS–US interval.

Plasticity of trace memory acquisition. A, Two groups of bees were trained in parallel with two successive trace-conditioning trials. Odor stimuli were always 0.5 s long. The two training trials were 10 min apart and differed with respect to the CS–US interval, which was either 5 s (CS5) or 10 s (CS10), and with respect to the odor that was used (hexanol or nonanol, both were used equally often as CS5 and CS10). The sequence of CS5 and CS10 stimulation in the memory test was balanced. Ai, Bees were first conditioned with the 5 s and then with the 10 s CS–US interval. These bees acquired both a CS5- and a CS10-trace memory (one-way repeated-measurement ANOVA; Fodor(3, 144) = 4.9, p = 0.003, N = 49). Aii, Bees were first conditioned with the 10 s and then with the 5 s CS–US interval. These bees acquired a CS5- but not a CS10-trace memory (one-way repeated-measurement ANOVA, Fodor(3, 153) = 5.6, p < 0.001, N = 52). Different letters indicate significant differences. B, Experiments were performed as in A, however, unlike in Aii, bees in Bii were first conditioned with overlapping CS and US (simultaneous conditioning, CS0) and then with a 10 s trace conditioning. Bi, Bees that were first conditioned with the 5 s CS–US interval were able to acquire both a CS5- and a CS10-trace memory (one-way repeated-measurement ANOVA, Fodor(3, 248) = 17.1, p < 0.001, N = 63). Bii, Bees that were first trained in a simultaneous conditioning trial acquired a CS0- but not a CS10-trace memory (one-way repeated-measurement ANOVA, Fodor(2, 177) = 48.8, p < 0.001, N = 52). During training, the CS0-evoked proboscis extension could not be recorded because bees extended their proboscis in response to the simultaneous sucrose (US) presentation. Different letters indicate significant differences. C, The effect of an additional 0.5-s-long odor pulse during the stimulus-free gap between the CS and US was tested by comparing memory acquisition between 5 s trace conditioning with (Ci) and without (Cii) an additional odor pulse (CS2) 2 s before the US. Both groups were conditioned in parallel. Ci, Bees acquired a CS5-specific trace memory after trace conditioning with an additional CS2 stimulus (one-way repeated-measurement ANOVA, Fodor(3, 264) = 9.4, p < 0.001). Cii, Memory performance after 5 s trace conditioning without additional odor presentation (one-way repeated measurement ANOVA, Fodor(3, 255) = 5.2, p < 0.01). Bees in Ci and Cii were conditioned in parallel to allow comparison (the experimental procedure in Cii was the same as in Fig. 3Ci. The weaker memory performance in Cii, compared with Fig. 3Ci, reflects seasonal variability). Ciii, Comparison of the CS5-specific associative memory retrieval between the CS5 and CS2 group and the CS5 group. Associative memory retrieval was higher in the CS5 and CS2 group than in the CS5 group (t test, p < 0.05). Thus, the stimulation with an additional odor (CS2) improved CS5-specific trace memory acquisition.
In mammals, trace conditioning becomes more effective when an additional stimulus is placed within the CS–US gap (Kamin, 1965; Kaplan, 1984). We tested whether this is the case in bees as well by performing a 5 s trace conditioning (CS5) and presenting a pulse of citral (CS2) 2 s before US onset (Fig. 4Ci). We compared the memory retrieval of this group with the memory retrieval of bees conditioned without the additional citral pulse (Fig. 4Cii). In both groups, bees acquired a CS5-specific trace memory, but this memory was higher in the group with the additional CS2 presentation (Fig. 4Ci). This cannot be attributed to an unspecific effect such as sensitization or generalization, because the effect was specific to the response to the CS5 and not to the new odor (Fig. 4Ciii). Thus, an additional odor stimulus during the stimulus-free gap strengthens the CS–US association.
PN activity during the gap does not represent the odor information in trace conditioning
Where in the brain is the physiological correlate of the odor trace? One possibility would be that olfactory PN in the antennal lobe encode the trace in the form of persistent neural activity. Previous studies have found persistent PN responses after odor offset (Sachse and Galizia, 2002; Galán et al., 2004); however, these post-odor responses have not yet been precisely characterized. Three conditions should be fulfilled for PNs to encode the odor trace: (1) post-odor response patterns should persist as long as the effective gap between CS and US (at least 6 s after odor onset), (2) properties of post-odor response patterns should include odor specificity and reliability, and (3) physiological and perceived similarity profiles should correspond to each other. To test these three criteria, we recorded odor responses in uniglomerular PNs by means of calcium imaging (Fig. 5).
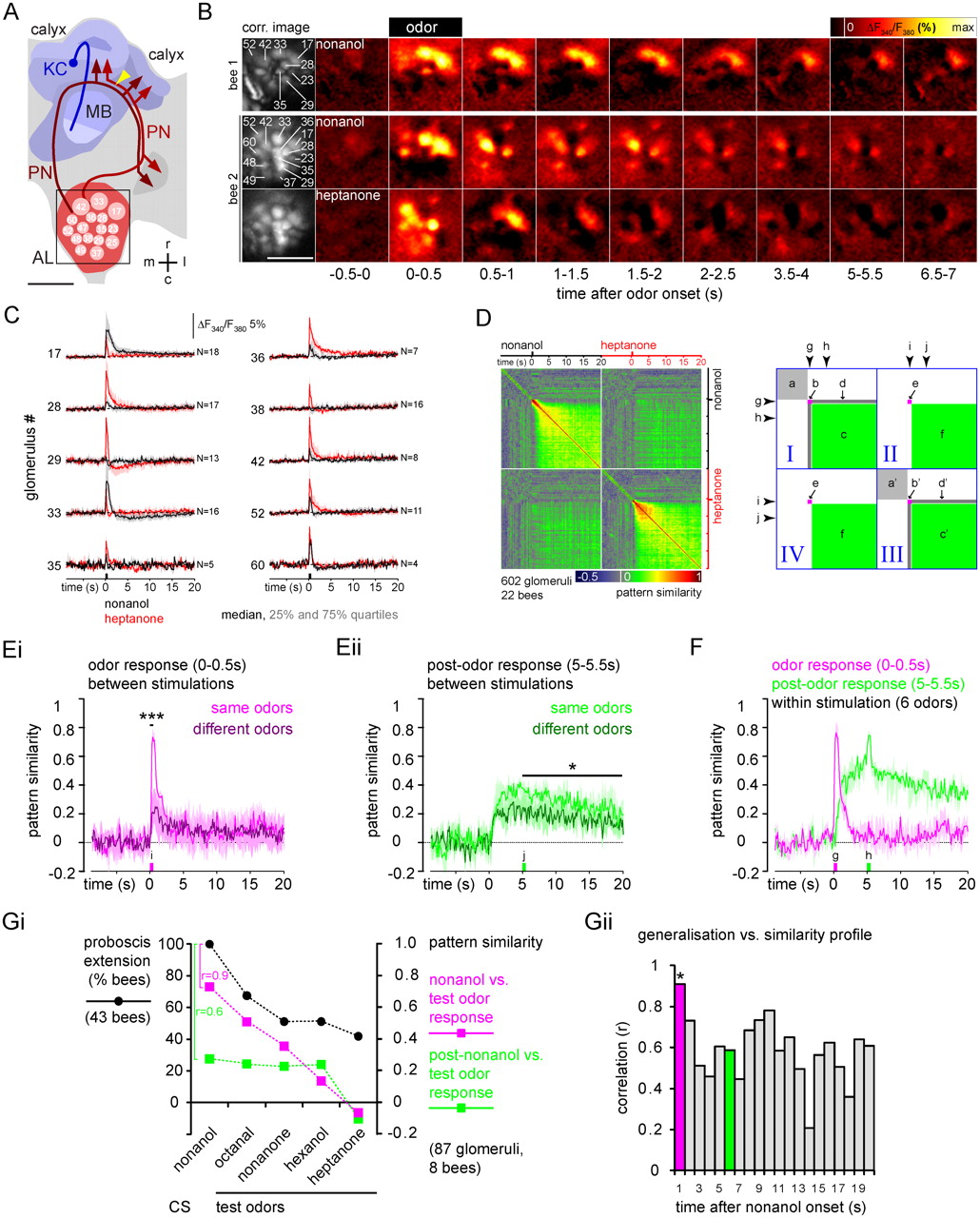
Trace memory relates to initial odor responses and not to post-odor responses in the antennal lobe. PNs were selectively stained with the calcium indicator Fura-2 dextran and their odor responses were recorded in the antennal lobe. A, Frontal view of the honeybee central olfactory system (left hemisphere). Olfactory receptor neurons terminate in the glomeruli (identity marked) of the antennal lobe (AL, red). Uniglomerular PNs transmit odor information from the antennal lobe to the mushroom body (MB, blue). PN axons project to the lateral and medial calyces where they form synapses with Kenyon cells (KC). PNs were stained by injecting Fura-2 between the calyces (yellow arrowhead). The square indicates the imaged area. Scale bar, 200 μm. r, Rostral; c, caudal; m, medial; l, lateral. B, Calcium responses of PNs in the antennal lobes of two bees. Left, Correlation (corr.) images used for glomerulus segmentation. Numbers indicate glomeruli that were unambiguously identified. Color-coded images show the signal changes (averaged over 0.5 s windows) before, during, and after stimulation with nonanol or heptanone. Scale bar, 200 μm. C, PN responses to nonanol (black) and heptanone (red) in identified glomeruli (median of responses from 4–18 bees; shading indicates the 25 and 75% quartiles). Odor stimuli induced a fast signal (initial odor response) followed by a slower positive or negative signal that lasted for several seconds (post-odor response). Both, initial odor and post-odor response patterns, were odor specific. D, Left, Similarity matrix shows the time-resolved similarity within (quadrants I and III) and between (quadrants II and IV) odor-response patterns. The similarity between activity patterns (vectors with signal values of all 602 glomeruli recorded in 22 bees) was quantified for each time point of the measurement by the Pearson correlation. Correlation values were color coded. Each pixel represents the similarity between two response patterns. The line of pixels at the time of nonanol presentation (g, arrowhead) represents the similarity between the initial nonanol response pattern and the response patterns at all time points within the same nonanol measurement (quadrant I) and the heptanone measurement (quadrant II). The diagonal (quadrant I and III) represents the autocorrelation at the respective time point. Right, Letters indicate areas showing the similarity within measurements before (a), during (b), and after (c) odor stimulation, between odor and post-odor response (d), across measurements between odor responses (e), and post-odor responses (f). E, Time-resolved similarity between repeated stimulation with same odors (six odors, first vs second stimulation, light traces) or with different odors (15 odor pairs, dark traces; median, 25% and 75% quartiles) for initial odor responses (0–0.5 s; Ei) and for post-odor responses (5–5.5 s; Eii). Asterisks represent significant differences between pattern similarity for same and different odors, averaged over the time indicated by the bars. ***p < 0.001, *p < 0.05 (t test). F, Time-resolved similarity between the initial odor response pattern (0–0.5 s after stimulus onset, magenta) and the entire measurement, and between the post-odor response pattern (5–5.5 s after stimulus onset, green) and the entire measurement (median, 25% and 75% quartiles for six odors; nonanol, heptanone: 602 glomeruli in 22 bees; hexanol, octanol, octanal, and nonanone: 87 glomeruli in 8 bees). g, Initial odor response pattern; h, post-odor response pattern. Gi, Comparison of the generalization profile after 5 s trace conditioning with nonanol as CS (black), and the similarity profiles for the physiological PN response patterns. Magenta, Similarity between the initial nonanol and test odor responses (first second after odor onset); green, similarity between the post-nonanol response (fifth second after odor onset) and the initial test odor responses. The similarity profile for the initial nonanol response correlated with the generalization profile (r = 0.9, p = 0.03), whereas that of the post-nonanol response did not (r = 0.6, p = 0.3). We therefore conclude that the post-odor response does not convey odor information in trace conditioning. Gii, Correlations between the generalization profile and pattern similarity profiles between initial test-odor responses (first second after odor onset) and post-nonanol responses for different time points after odor onset. Magenta and green bars represent the correlation between the generalization and pattern similarity profiles shown in Gi. The asterisk marks the only significant correlation (p = 0.03). The p values of the remaining correlations ranged between 0.12 (10 s) and 0.74 (14 s).
Post-odor response patterns
Each odor induced an odor-specific initial pattern of activated and inhibited glomeruli (Fig. 5B,C) that corresponded to the previously described glomerular response patterns (Galizia et al., 1999b; Sachse and Galizia, 2003). After odor offset, activity changed into a prolonged pattern of activated and inhibited glomeruli (post-odor response) that was different from the initial odor response. Post-odor responses of individual glomeruli were not predictable from their odor response. For example, glomerulus 33 showed a positive initial response to nonanol and heptanone, but it showed a prolonged negative post-odor response for nonanol only (>20 s) (Fig. 5C).
Properties of post-odor response patterns
We next analyzed the dynamics and odor specificity of odor response and the post-odor response patterns by measuring the activity pattern similarity within and between measurements (Fig. 5D). Similarity between activity patterns was measured as frame-by-frame correlation (r) between glomerular response vectors. This analysis yielded a similarity matrix in which each pixel depicts the correlation between the patterns at one versus another time point. Time goes from left to right and from top to bottom, and the diagonal is always 1 (self-similarity). Before odor onset (Fig. 5D, a), the correlation fluctuated ∼0 (−0.01 ± 0.13, mean ± SD), indicating that background activity was random. Odor stimulation (0.5 s) gave rise to an initial response that was characterized by an increased similarity for ∼2 s (nonanol, r = 0.86 ± 0.08; heptanone, r = 0.62 ± 0.27) (Fig. 5D, b). Post-odor response is visible in a homogeneous increase in similarity (nonanol and heptanone, r = 0.4 ± 0.14) (Fig. 5D, c) with no similarity to the initial odor response (Fig. 5D, d) (nonanol, r = 0.03 ± 0.18; heptanone, r = 0.01 ± 0.15). Nonanol and heptanone evoked different patterns from each other, both for the initial odor responses (r = 0.03 ± 0.09) (Fig. 5D, e) and for the post-odor responses (r = 0.14 ± 0.08) (Fig. 5D, f). Initial odor and post-odor response patterns were reproducible (Fig. 5E), indicating that both the initial odor and post-odor responses were odor-specific. The transition from the odor response to the post-odor response pattern did not occur exactly at odor offset, but rather between 0.5 and 1.4 s after odor onset (0.7 ± 0.15 s, mean ± SEM) (Fig. 5F).
Physiological and perceived similarity profiles
Even though initial odor and post-odor responses differ, the post-odor response could still serve as a substrate of the odor trace. A previous study has shown that the perceived odor similarity correlates well with the physiological similarity between odor-evoked glomerular activity patterns in the honeybee (Guerrieri et al., 2005). We argue that, if the neural substrate of an odor trace would be in the post-odor PN activity, then the perceived odor similarity should also match post-odor physiological similarity. We quantified perceived nonanol similarity as the generalization profile after a 5 s trace conditioning. Post-odor physiological similarity (Fig. 5Gi) was calculated between post-nonanol responses (5–6 s after odor onset) and the initial test odor responses (0–1 s after odor onset). We chose the initial test odor response patterns, because in the test situation bees typically responded within 1.5 s after odor presentation (Fig. 3D). The similarity profile of the post-nonanol response did not correlate significantly with the perceived similarity profile (r = 0.6, p = 0.3). In contrast, the similarity profile of the initial nonanol response correlated significantly with the generalization profile (r = 0.9, p = 0.03). Figure 5Gii shows the correlation between the generalization profile and the similarity profile of the nonanol-evoked activity patterns at different times after stimulus onset. Again, the only significant correlation between the generalization profile and the physiological similarity profile was for the initial nonanol response. Thus, the initial odor response of PNs, rather than the post-odor response, represents perception-like odor information in trace conditioning.
A previous imaging study in honeybees suggested that odor stimulation results in a Hebbian-like strengthening between coactivated glomeruli, creating a transient network attractor (Galán et al., 2006). This attractor could mediate an odor trace by becoming active during US presentation only. In insects, the US (sucrose reward) is encoded by the octopaminergic VUMmx1 neuron, which projects to the antennal lobe (Hammer, 1993). Octopamine has excitatory effects on neural activity (Flecke and Stengl, 2009). Thus, octopamine release during US presentation may reactivate the odor trace, and thus allow for coincident activity necessary for associations. To test this idea, we compared PN responses to nonanol alone and to nonanol with subsequent sucrose stimulation of the proboscis (Fig. 6A). The odor–sucrose pairing was done as in trace conditioning with a 5 s CS–US interval. However, different from the behavioral experiments, sucrose was applied to the proboscis only and not to the antenna, because sucrose-stimulation of the antenna induces its own activity pattern that might override trace-related activity (Peele et al., 2006; Szyszka et al., 2008). Sucrose stimulation of the proboscis, however, had no visible effect on PN activity patterns across identified glomeruli. To test for more subtle across-glomeruli reward effects, we calculated changes in the similarity of the entire across-glomerular response pattern (Fig. 6B). Again, there was no consistent effect of sucrose on the odor-induced glomerular activity patterns; in particular, the post-odor response pattern did not become more similar to the initial odor response. This was not because the sucrose reward to the proboscis failed to induce activity in the antennal lobe; even though no clear sucrose responses were visible in the averaged PN responses (Fig. 6C), sucrose stimulation evoked weak but reproducible response patterns across PNs (Fig. 6D). We thus reject the hypothesis that an odor trace becomes activated in the presence of the reward and we conclude that the odor trace is not encoded in PN calcium or spiking activity within the antennal lobe.
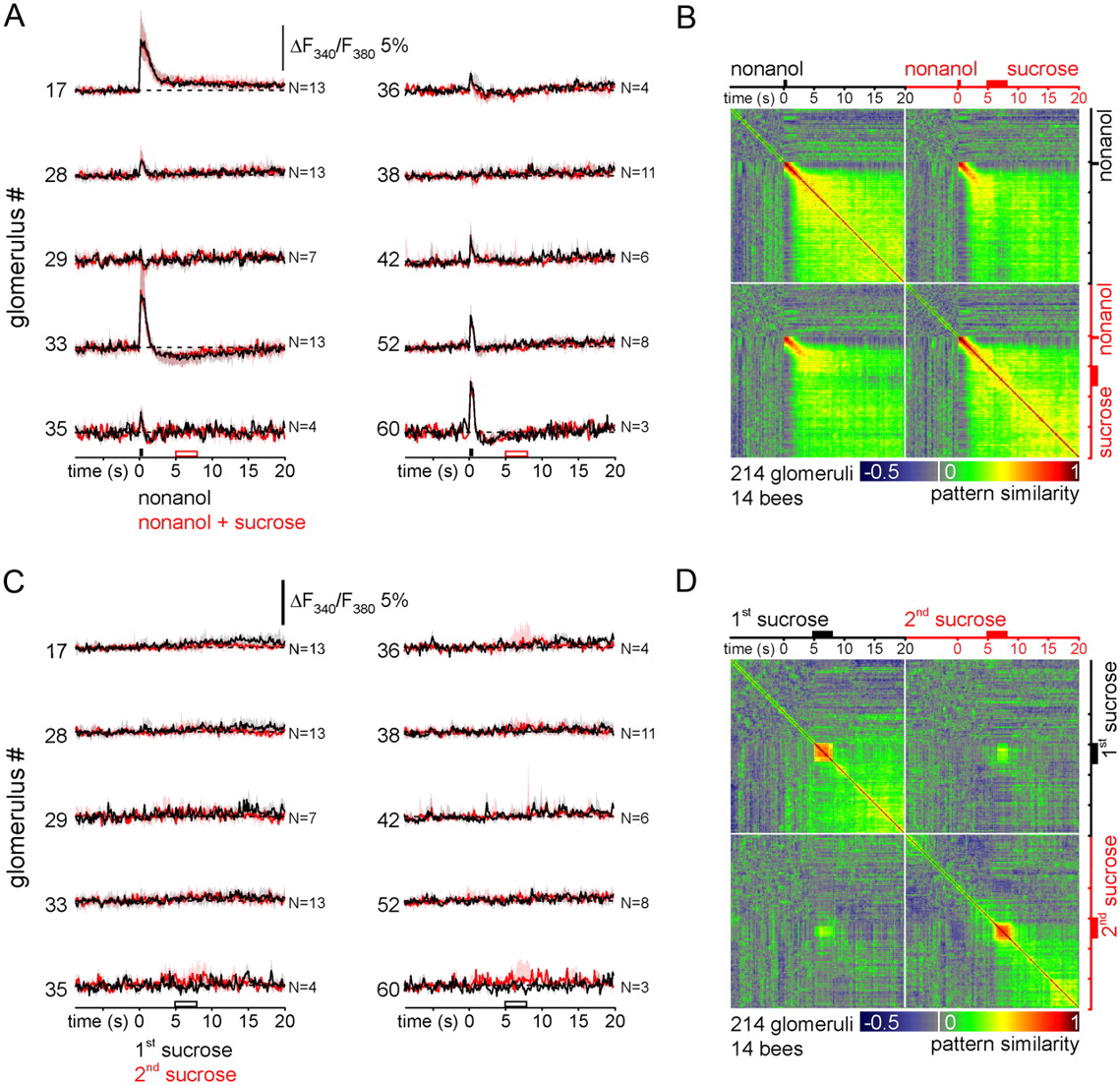
Odor–reward pairing in trace conditioning has minor effects on PN′s post-odor responses. A, PN responses to nonanol alone (black) and to the pairing of nonanol and a sucrose stimulation of the proboscis (red) in identified glomeruli (median, 25% and 75% quartiles of responses from 3–13 bees). The sucrose stimulus 5 s after odor onset did not induce visible signals. B, Time-resolved similarity within and between response patterns. Both nonanol and nonanol plus sucrose stimulation induced initial odor and post-odor responses. These were reproducible, as seen by the high correlation in quadrants II and IV. Sucrose stimulation had a minor effect on the pattern similarity and did not increase the similarity between the initial response and the post-odor response pattern. C, PN activity during sucrose-only stimulation (black, first stimulation; red, second stimulation). The sucrose stimulus did not induce consistent responses in identified glomeruli. D, Time-resolved similarity within and between PN activity patterns revealed sucrose-induced response patterns, which were reproducible (see quadrants II and IV).
Discussion
Taking advantage of the honeybee's exceptional learning capabilities, we conducted the first comprehensive characterization of olfactory trace conditioning, in which an odor (CS) was paired with a temporally separated sucrose reward (US).
Features of trace-memory acquisition
Olfactory trace conditioning differed from delay conditioning as its learning rate and asymptotic level were lower and memory acquisition decreased with increasing CS–US intervals (Fig. 2A,B). This finding is in accordance with a previous study in honeybees (Menzel and Bitterman, 1983) and with trace conditioning in mammals (Weisz and Walts, 1990); however, it differs from the moth, in which trace conditioning is more effective than delay conditioning (Ito et al., 2008).
The differences between trace and delay memory acquisition may indicate that their neural mechanisms for memory formation differ. Indeed, depending on the number and spacing of trials, associative conditioning induces multiple memory phases, which correspond to different molecular mechanisms (Menzel, 2001; Müller, 2002). We therefore wondered whether memory consolidation differs for conditioning with different CS–US intervals. However, the lack of a difference between short-term and early long-term memory for varying CS–US intervals (Fig. 2Biii) indicates that trace and delay memories use the same molecular mechanisms.
Why then, is memory acquisition weaker in trace than in delay conditioning? In classical conditioning, memory acquisition decreases with decreasing associative strength, which is a function of CS and US salience (Rescorla and Wagner, 1972). Thus, increasing the CS–US gap might decrease the salience of the CS, the US, or both. The CS salience may decrease with increasing CS–US intervals, just as decreasing odor concentration reduces its saliency and reduces learning (Pelz et al., 1997). Alternatively, or additionally, CS onset may influence the salience of a following US, and this effect may decrease with increasing CS–US intervals. In Drosophila, the habituated response to a US (electroshock) can be dishabituated by odor stimulation, showing that odors can indeed increase US salience (Acevedo et al., 2007). Such a CS-mediated increase in the US salience would also explain the improved memory acquisition that we found when a second odor stimulus was placed into the CS–US gap (Fig. 4C).
Single-trial acquisition reveals a sensory odor trace
Olfactory trace conditioning in honeybees shares features with trace eyeblink conditioning in mammals, where the CS is a tone and the US an aversive air puff to the eye. Perhaps the most striking difference to mammalian trace conditioning is that honeybees acquire odor trace memories during a single learning trial (Fig. 3Aii), whereas trace eyeblink conditioning requires several hundred training trials (Woodruff-Pak and Disterhoft, 2008). Honeybees' ability to acquire a trace memory after only a single trial requires the existence of a US-independent CS trace. In mammals, in contrast, it is not clear whether the CS leaves a sensory trace containing the stimulus-specific information that is then associated with the US, or whether the neural US representation shifts backward in time until it overlaps with the CS (US anticipation) (Fig. 3Ai). The latter view is supported by the finding that US-encoding dopaminergic neurons in primates change their response onset from just after the US to the CS onset after multiple appetitive conditioning trials (Schultz et al., 1997). Accordingly, in trace eyeblink conditioning, US-induced activity in hippocampal neurons shifts backward in time and finally coincides with the CS onset (McEchron and Disterhoft, 1997). It is possible, however, that training induces a US shift in addition to an existing CS trace in honeybees.
The olfactory system maintains a trace of the odor onset
Honeybees' odor traces were odor-specific and the perceived odor similarity profiles did not differ between trace and delay conditioning (Fig. 2C). These results were similar to findings in Drosophila (Galili et al., 2011) and may indicate that the neural substrate of odor memories is the same for trace and delay conditioning. Odor information from the initial part of an odor stimulus was transformed into the odor trace, whereas the late phase of a continuing odor stimulus did not influence the odor trace (Fig. 3). This resembles the situation in mammals, where the CS onset contributes mostly to the trace memory (Kehoe et al., 2009). Parallel to response strength, the response latency is a measure of associative strength, as it decreases with acquisition (Rehder, 1987). The latency is also a function of task difficulty (Wright et al., 2009). Since in trace conditioning, the acquisition rates were lower than in delay conditioning, the longer latency likely reflects a weaker associative strength and more difficult task, rather than acquired knowledge about the CS–US timing. In our experiments, bees showed the conditioned response later after trace conditioning, but earlier than the predicted reward (Fig. 3D). We thus conclude that US-timing information is either not learned or not behaviorally relevant.
Neural substrate of the odor trace: a place for Kenyon cells?
We have shown that odor traces are not stored in PN calcium activity (Fig. 5G). This is surprising, given that Galán et al. (2006) found a sensory odor memory in the form of increased stochastic coupling between coactivated glomeruli. It thus remains to be shown whether the odor-specific post-odor response patterns that we found have a functional relevance or whether they are a mere byproduct of odor processing in the antennal lobe. The absence of odor-trace-related calcium activity in PNs, however, does not exclude the possibility that PNs encode the trace in form of biochemical changes. For example, the initial odor response may elevate PNs' cAMP level, a mechanisms that has been discussed to underlay odor traces in Drosophila mushroom body neurons (Tomchik and Davis, 2009).
The mushroom body, in fact, is a plausible candidate for encoding the odor trace. This higher brain area is involved in learning and memory (Menzel, 2001; Heisenberg, 2003) and the mushroom body intrinsic Kenyon cells feature some properties that make them well suited for encoding odor traces. For example, they convey information about the initial PN response only (Szyszka et al., 2005), just as an odor trace does. Normally, Kenyon cells' odor responses are short and go back to baseline within a few seconds, even in the presence of an odor (Perez-Orive et al., 2002; Szyszka et al., 2005; Ito et al., 2008; Turner et al., 2008). However, during the pairing of an odor with a US (sucrose), odor-activated Kenyon cells become reactivated (Szyszka et al., 2008). In Szyszka et al. (2008), CS and US overlapped; it remains to be shown whether odor-responsive Kenyon cells can also be reactivated by the US after CS offset. Moreover, odors activate primarily non-overlapping Kenyon cell ensembles. This would allow parallel representation of multiple odor traces (Heisenberg, 2003, Szyszka et al., 2005) and would explain the robustness of odor traces against the interference by other odors (Fig. 4C).
Furthermore, there is evidence that the mushroom body is particularly involved in solving ambiguous learning tasks, as it is required for reversal learning (Devaud et al., 2007). Trace conditioning represents an ambiguous task: animals must distinguish between the CS–US and the US–CS association, since the stimuli do not co-occur (Beylin et al., 2001). Thus, the mushroom body might equally be involved in resolving the ambiguous CS–US relationship in trace conditioning. Interestingly, the mammalian hippocampus is specifically needed for reversal learning (Berger and Orr, 1983), and it is thought to resolve the temporally ambiguous stimulus relationship in trace conditioning (Beylin et al., 2001; Bangasser et al., 2006).
Relevance of olfactory traces
In which situations could animals use odor traces? Odor traces might be involved in higher-level cognitive processes, as has been implicated for trace conditioning in mammals, which is thought to induce a basal form of declarative memory (Clark, 2011). Odor traces could, for example, serve as short-term olfactory storage for working memory (Baddeley, 1992). In honeybees, olfactory and visual working memory have been approached experimentally in the delayed matching-to-sample paradigm, in which animals learn to respond to a matching stimulus (Giurfa et al., 2001; Menzel, 2009). In this paradigm, a bee has to keep a transient nonassociative memory of the sample stimulus and then apply a learned rule to choose the right stimulus. The largest effective time interval between the sample was ∼5 s (Zhang et al., 2005), a value close to the 6 s odor trace found here (Fig. 2B). Our present results cannot directly identify whether olfactory trace memories in honeybees have features of working memories. However, our finding that bees can extend their trace time span by experience (Fig. 4) is reminiscent of attention-like processes, and might implicate higher-level cognitive processes in honeybee trace conditioning that may not be necessary for delay conditioning.
Footnotes
This work was supported by Bundesministerium für Bildung und Forschung (01GQ0771 to C.G.G. and 01GQ0931 to P.S. and C.G.G.) and Deutsche Forschungsgemeinschaft (GA524/8-1 and GA524/12-1 to C.G.G.).We thank Hiromu Tanimoto, Dana Galili, Andreas V.M. Herz, and Randolf Menzel for sharing their ideas and for critical discussion of the manuscript; Nadine Treiber for help with the behavioral experiments; and Missanga van de Sand, the entire Galizia group, and three anonymous reviewers for valuable input.
- Correspondence should be addressed to Paul Szyszka, Department of Biology, University of Konstanz, D-78457 Konstanz, Germany. paul.szyszka{at}uni-konstanz.de





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}