

Article Figures & Data
Figures
- Figure 1.
Experimental stimuli, subject reports, and performance. A, An epoch from an Illusory fading trial. Physical stimulus (top row; fixation dot not to scale), subject's perception of the stimulus (second row), and subject's report (third row). B, Distributions of the durations of fading and intensification percepts. C, Total amount of fading time across target eccentricities. D, Fading onset rate at each eccentricity. Asterisks indicate significantly higher fading onset rates at peripheral eccentricities for the pairs 0–3°, 0–6°, and 0–9° (one-tailed paired t test; *p = 0.012, **p = 0.0022, ***p = 0.0031). E, Subjects' performance in the Real fading condition. Shading and error bars indicate the SEM across subjects (n = 7).
- Figure 2.
Eye movements. A, Descriptive statistics for microsaccades (<1°), saccades (≥1°), drifts, and blinks. Numbers in parentheses indicate the SEM across subjects (n = 7). B, Magnitude distribution of microsaccades and saccades. Shading indicates the SEM across subjects (n = 7). C, Microsaccadic and saccadic peak velocity–magnitude relationship for all subjects combined. Each red dot represents a microsaccade or a saccade with peak velocity indicated on the y-axis and magnitude indicated on the x-axis.
- Figure 3.
Microsaccade rates relative to reported and perceptual transitions. A–E, Average microsaccade rates around reported transitions toward intensification versus fading in the Illusory fading condition. The solid vertical line indicates the reported transitions (t = 0). The horizontal dashed line indicates the average microsaccade rate. Target eccentricity is indicated at the top of each panel. The vertical dashed line and gray shading indicate the estimated time of the perceptual transitions (mean latency and 1 SD from the mean, extrapolated from the subject's detections of physical transitions in each inset). Insets, Reaction time distributions from correctly identified physical transitions (Real fading condition), for each eccentricity. In A, microsaccade rates depart from baseline ∼248 ms before the perceptual transitions (beginning of the vertical gray shading) and ∼764 ms before the reported transitions. Similar timing applies to each separate eccentricity in B–E. The gray dots and dashes (above the main red and blue traces) indicate the bins where microsaccade rates before transitions to intensification were significantly higher than microsaccade rates before transitions to fading (one-tailed paired t tests with Bonferroni correction, bin size = 20 ms, p value <0.01). Red and blue shading and error bars indicate the SEM across subjects (n = 7).
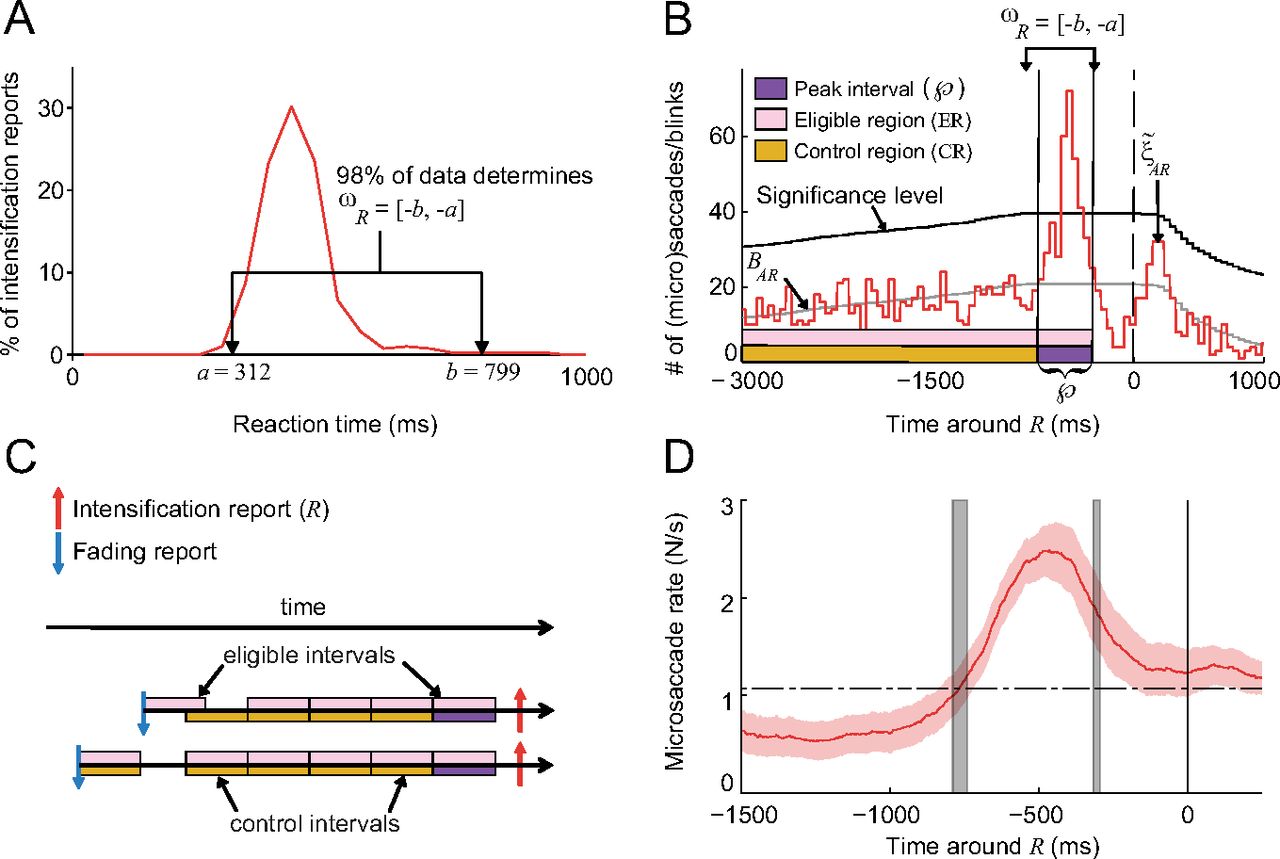
- Figure 4.
The peak, control, and eligible intervals. A, One subject's reaction time distribution to physical intensifications during the Real fading condition, and ωR (first approximation of the peak interval) for that subject. The average ωR across subjects was [−801, −291] ± [21, 9] ms SEM. B, ξ̃AR is the correlogram of all eye movements with Rs for the same subject from A, BAR is the baseline, and the purple interval contained inside of ωR is the peak interval. C, Examples of the peak, eligible, and control intervals for two Rs. D, Correlation of microsaccades with Rs (as in Fig. 3A) with gray vertical shaded lines indicating the average location of
 ([−786, −312] ± [24, 17] ms SEM). Red shading indicates the SEM across subjects (n = 7).
([−786, −312] ± [24, 17] ms SEM). Red shading indicates the SEM across subjects (n = 7). - Figure 5.
Ocular events. Each row shows three 2 s epochs during the Illusory fading condition from one subject. The black trace indicates the eye's horizontal position. The blue and red pulses represent the subject's reports. The purple pulse indicates the location of the peak interval, in which ocular events can potentially cause Rs. Several types of ocular events are indicated above their peak intervals.
- Figure 6.
Contribution and efficacy for different ocular events. A, The bars and pie chart show P(E|
), the probability of an event E occurring within the peak interval, for each event E. Because P(DA|) = 17%, the upper bound on the contribution of DA is 17%. B, The contribution, 𝒞R(E), of each ocular event E. C, The efficacy, ℰR(E), of each ocular event E. Error bars indicate the SEM across subjects (n = 7 for A and B), see Table 1 for the number of subjects in each type of event in C). - Figure 7.
Microsaccade magnitude and single versus multiple microsaccades. A, Distributions of magnitudes of microsaccadic events in
and in the CR (inset bars are the averages of the subjects' mean microsaccadic event magnitude). Microsaccadic events during are significantly larger than those before (*one-tailed paired t test, p = 0.0002). B, Efficacy of microsaccadic events of different magnitudes and saccadic events. B, C, Mi denotes the microsaccadic event M whose magnitude falls in the interval [0.25(i − 1), 0.25i] degrees, for i = 1, …, 4. C, Contribution of microsaccadic events of different magnitudes and saccadic events, and their probability of occurring within . D, Contribution and efficacy of microsaccadic events M with one microsaccade (M1) versus two or more microsaccades (M≥2). Shading and error bars indicate the SEM across subjects (n = 7 in A and C and contribution plot in D; see Table 1 for the number of subjects in B and efficacy plot in D). - Figure 8.
Efficacy of microsaccades as a function of their direction. A, θC is the angle between the microsaccade direction and the compass position of the Gabor. θO is the angle between the microsaccade direction and the orientation of the Gabor. B, Efficacy of microsaccades did not vary as a function of their direction relative to the compass position of the Gabor (two-tailed paired t tests with Bonferroni correction and a family significance level α = 0.05). MCi denotes the microsaccadic event M whose θC falls in the interval [10(i − 1), 10i] degrees, for i = 1, …, 18. C, Efficacy of microsaccades did not vary as a function of their direction relative to the orientation of the Gabor (two-tailed paired t tests with Bonferroni correction and a family significance level α = 0.05). MOi denotes the microsaccadic event M whose θO falls in the interval [10(i − 1), 10i] degrees, for i = 1, …, 9. Error bars indicate the SEM across subjects (see Table 1 for the number of subjects in each type of event in B and C).















Tables
Ocular event (E) Number of subjects with ≥6 eligible events Average number (±SEM) of eligible events M 7 667 ± 199 S 5 17 ± 5 B 4 25 ± 8 MS 4 33 ± 9 MB 4 17 ± 2 M1 (magnitude in [0, 0.25] degrees) 6 122 ± 47 M2 (magnitude in [0.25, 0.5] degrees) 7 350 ± 112 M3 (magnitude in [0.5, 0.75] degrees) 7 191 ± 57 M4 (magnitude in [0.75, 1] degrees) 7 42 ± 12 M1 (one microsaccade) 7 530 ± 179 M≥2 (two or more microsaccades) 6 157 ± 31 DA (drift alone) 7 969 ± 248 MC1 (θC in [0 10] degrees) 6 27 ± 6 MC2 (θC in [10 20] degrees) 6 34 ± 10 MC3 (θC in [20 30] degrees) 6 34 ± 12 MC4 (θC in [30 40] degrees) 6 36 ± 9 MC5 (θC in [40 50] degrees) 6 39 ± 12 MC6 (θC in [50 60] degrees) 6 35 ± 12 MC7 (θC in [60 70] degrees) 6 32 ± 12 MC8 (θC in [70 80] degrees) 6 38 ± 10 MC9 (θC in [80 90] degrees) 6 47 ± 14 MC10 (θC in [90 100] degrees) 6 43 ± 11 MC11 (θC in [100 110] degrees) 6 42 ± 12 MC12 (θC in [110 120] degrees) 6 39 ± 12 MC13 (θC in [120 130] degrees) 7 44 ± 15 MC14 (θC in [130 140] degrees) 6 46 ± 13 MC15 (θC in [140 150] degrees) 7 40 ± 12 MC16 (θC in [150 160] degrees) 7 41 ± 17 MC17 (θC in [160 170] degrees) 6 42 ± 16 MC18 (θC in [170 180] degrees) 6 33 ± 7 MO1 (θO in [0 10] degrees) 6 32 ± 8 MO2 (θO in [10 20] degrees) 7 66 ± 19 MO3 (θO in [20 30] degrees) 7 88 ± 24 MO4 (θO in [30 40] degrees) 7 104 ± 33 MO5 (θO in [40 50] degrees) 7 133 ± 45 MO6 (θO in [50 60] degrees) 7 98 ± 33 MO7 (θO in [60 70] degrees) 6 77 ± 24 MO8 (θO in [70 80] degrees) 7 70 ± 14 MO9 (θO in [80 90] degrees) 6 24 ± 6 The efficacy calculations in Figs. 6D; 7B,D; and 8B,C required that each subject had a minimum of six occurrences per ocular event type.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}