

Abstract
Dopamine increases triggered by cocaine and consequent stimulation of dopamine receptors (including D1 and D2) are associated with its rewarding effects. However, while facilitation of D1 receptor (D1R) signaling enhances the rewarding effects of cocaine, facilitation of D2R signaling decreases it, which indicates that for cocaine to be rewarding it must result in a predominance of D1R over D2R signaling. Moreover, the transition to compulsive cocaine intake might result from an imbalance between D1R and D2R signaling. To test the hypothesis that chronic cocaine use unbalances D1R over D2R signaling during cocaine intoxication, we used microprobe optical imaging to compare dynamic changes in intracellular calcium ([Ca2+]i, marker of neuronal activation) to acute cocaine in striatal D1R-EGFP and D2R-EGFP-expressing neurons between control and chronically treated mice. Chronic cocaine attenuated responses to acute cocaine in D1R (blunting Ca2+ increases by 67 ± 16%) and D2R (blunting Ca2+ decrease by 72 ± 17%) neurons in most D1R and D2R neurons (∼75%). However, the dynamics of this attenuation during cocaine intoxication was longer lasting for D2R than for D1R. Thus, whereas control mice showed a fast but short-lasting predominance of D1R over D2R signaling (peaking at ∼8 min) during acute cocaine intoxication, in chronically treated mice D1R predominance was sustained for >30 min (throughout the measurement period). Thus, chronic cocaine use dramatically reduced cocaine-induced DA signaling, shifting the balance between D1R and D2R signaling during intoxication to a predominance of D1R (stimulatory) over D2R (inhibitory) signaling, which might facilitate compulsive intake in addiction.
Introduction
Dopamine (DA) increases triggered by cocaine and the consequent stimulation of DA receptors (including D1 and D2 receptors) in brain reward regions are associated with its rewarding effects (Koob and Bloom, 1988). However, while facilitation of D1 receptor (D1R) signaling enhances the rewarding effects of cocaine (Graham et al., 2007), facilitation of D2R signaling decreases cocaine reward (Lobo et al., 2010), which indicates that for cocaine to be rewarding it must result in a predominance of D1R over D2R signaling. Thus, an imbalance between the signaling through D1R (enhancing) and D2R (attenuating) resulting in enhanced D1R or attenuated D2R signaling during cocaine intoxication could increase its incentive salience as observed in cocaine addiction (Thompson et al., 2010). Preclinical studies have shown that the enhancement of D2R signaling decreases cocaine reward (Lobo et al., 2010) and interferes with cocaine self-administration (Thanos et al., 2008), whereas interfering with D2R signaling facilitates cocaine sensitization (Luo et al., 2011). This is in contrast with the increased sensitivity to cocaine reward reported with enhancement of D1R signaling (Lobo et al., 2010).
In the striatum, D1R and D2R are expressed in medium spiny neurons (MSNs) and exert opposing intracellular effects on cAMP signaling; D1R activates it, whereas D2R inhibits it (Sibley et al., 1993). Repeated cocaine use has been reported to upregulate the cAMP signaling pathway, which would be suggestive of enhanced D1R over D2R signaling (Anderson and Pierce, 2005). However, others have reported no changes or decreases in cAMP with chronic cocaine exposures (Crawford et al., 2004). Thus, the extent to which chronic cocaine use can modify the relative prevalence of D1R over D2R signaling is still unclear. Moreover, the effects of chronic cocaine use on the relative changes in D1R over D2R signaling during cocaine intoxication have not been investigated. In cocaine abusers, D2R signaling during intoxication with stimulant drugs (methylphenidate and amphetamine) is markedly attenuated when compared with controls (for review, see Volkow et al., 2011). Since the loss of control in cocaine-addicted individuals is triggered when they are exposed to cocaine or to cocaine cues, it is important to study the effects of chronic cocaine exposure on DA signaling during cocaine intoxication. Here, we tested the hypothesis that chronic cocaine exposure reduces cocaine-induced increases in DA signaling but enhances the predominance of D1R over D2R signaling during cocaine intoxication.
For this purpose, optical imaging was used to measure the effects of acute cocaine intake on the dynamic changes in [Ca2+]i (a marker of cell function) using Rhod2 (a fluorescent [Ca2+]i indicator) in striatal MSNs of transgenic mice that expressed EGFP under the control of either the D1R or the D2R gene (Gong et al., 2003). A custom epifluorescence microscope integrated with a microneedle endoscope [Luo et al., 2011; φ, 1 × 25 mm; 0.65 numerical aperture (NA)] allowed us to simultaneously image striatal EGFP in individual D1R- or D2R-expressing neurons and their [Ca2+]i changes in response to acute cocaine use (8 mg/kg, i.p.) in vivo in naive mice (referred to as “control mice”) and in mice chronically exposed to cocaine (30 mg/kg/d, 2 weeks; referred to as “chronic mice”).
Materials and Methods
Animals.
Drd1-EGFP and Drd2-EGFP bacterial artificial chromosome (BAC) transgenic mice generated by the Gensat BAC transgenic project were used (Gong et al., 2003). These animals were divided into different experimental groups (Table 1, Experiment no.). Protocols were performed in accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals and were approved by the Institutional Animal Care and Use Committees of Stony Brook University.
Summary of experimental groups
Surgical preparation.
Mice (50% male and 50% female) were anesthetized with 2% isoflurane mixed in pure O2, and their heads were immobilized using a custom stereotaxic frame. A cranial window (φ, 1∼1.5 mm) was then created ∼2 mm lateral and 0.4 mm anterior to bregma and above the striatum (i.e., in the caudate–putamen nucleus region (Hof et al., 2000; Paxinos and Franklin, 2004). The [Ca2+]i fluorescence indicator Rhod2/AM (12.5 μg/100 μl; Invitrogen) was slowly infused into the brain using a microinfusion pump (3 μl/min; CMA 400, Carnegie Medicine). Image acquisition started after 1 h to ensure maximal intracellular uptake of Rhod2. The labeling distribution profile of Rhod2 was ∼1 mm φ around the loading spot.
Drug treatment.
Acute cocaine (cocaine HCl; 8 mg/kg, i.p.) was administered. Chronic cocaine (30 mg/kg, i.p.) was administered once a day for ∼2 weeks, and the same volume of saline (∼0.1 cc/10 g) was administered once a day for ∼2 weeks for the control group. In the article, we refer to the chronically cocaine exposed mice as chronic mice and to the mice treated chronically with saline as control mice. SCH23390 (75 μg/kg, i.p.; Sigma-Aldrich) and raclopride (3 mg/kg, i.p.; Sigma-Aldrich), both of which were administrated 30 min before acute cocaine injection, were used to assess the effects of D1R blockade and D2R blockade, respectively (Table 1).
In vivo microprobe fluorescence imaging.
The microprobe fluorescence imaging technique was applied to D1R-EGFP and D2R-EGFP transgenic mice (both control mice and chronic mice) to enable in vivo characterization of the dynamic [Ca2+]i changes in striatal D1R and D2R neurons in response to an acute cocaine challenge. The principle of the endomicroscopic imaging technique has been outlined previously (Jung et al., 2004; Luo et al., 2011), and Figure 1 illustrates the system setup used in this study, in which modifications have been implemented to improve fluorescence coupling and image throughput. For instance, additional Piezoelectrically activated focusing mechanism is used to enable instantaneous fine focal tracking to maximize [Ca2+]i-Rhod2 fluorescence detection within the field of view (FOV). After anesthesia and surgical preparation, the mouse along with the stereotaxic frame was mounted on a motorized 3D stage adapted to a modified upright fluorescence microscope (E800, Nikon) as shown in Figure 1a. Figure 1b illustrates a microneedle probe, a custom GRIN lens (φ, 1 × 25 mm; 0.65 NA) that relayed the image of mouse inner brain (e.g., striatum) at the distal end of the needle probe back to the focal plane of the microscope objective (e.g., PlanFluo 20×, 0.5 NA; Nikon). A motorized 3D microstage facilitated accurate light coupling and focal tracking between the microneedle probe and the microscope objective, allowing for accurate focusing on the striatal region where there is a high density of GFP-expressing neurons (e.g., at ∼3 mm below mouse cortical surface). Additionally, a custom microsleeve anchoring on mouse cortical bone reduces motion-induced artifacts and potentially enables implantable microprobe to be easily coupled with the microscope for repeated imaging studies on live animals (i.e., time course studies). Special attention was paid to avoid disruption of large vascular vessels during needle-probe injection, and minor bleeding was occluded by flushing saline solution. For dual-channel EGFP and Rhod2-Ca2+ fluorescence imaging, gated sequential illumination for the excitation of EGFP (λex = 460–500 nm) and Rhod2-Ca2+ (λex = 530–550 nm) was used to minimize photon bleaching. EGFP and Rhod2-[Ca2+]i fluorescence emissions of D1R or D2R neurons were detected by FITC/EGFP (excitation (EX): 460–500 nm, dichroic mirror (DM): 505 nm, barrier or emission filter (BA): 510–560 nm, Chroma #41001) and TRITC/Rhod (EX: 530–550 nm, DM: 565 nm, BA: 590–650 nm, Chroma #41002c) epifluorescence cubes, respectively. The images were acquired by an EM CCD camera (iXon+ 885; 1 m pixels, 8 μm/pixel; Andor) synchronized with the fluorescence excitation. Full-field microneedle images (FOV, φ 250 μm) were recorded continuously at 28 fps from baseline t ≈ −5 min to t ≈ 30 min after cocaine administration.

Microprobe fluorescence microscope for in vivo deep brain imaging. a, A modified fluorescence microscope enabling simultaneous EGFP and Rhod2-[Ca2+]i fluorescence imaging. b, Microneedle (μN) catheter, a 3D motorized microstage assembly, and a PZT-actuated ring holder for deep brain subcellular imaging. Obj, Objective; Ex, excitation; Em, emission. The dashed box illustrates the time base for dual-channel fluorescence illumination and imaging.
Image processing and statistical analysis.
The Rhod2-[Ca2+]i fluorescence image was coregistered with the corresponding EGFP image to identify [Ca2+]i of D1R or D2R EGFP-expressing neurons from non-EGFP cells. We quantified all of the D1R- or D2R-EGFP neurons in the field of view and measured their time course of [Ca2+]i changes (i.e., Δ[Ca2+]i) in response to acute cocaine for each neuron and normalized it to their mean basal levels (i.e., t < 0). Mean Δ[Ca2+]i as a function of time (Δ[Ca2+]i ∼ t curves) were computed by averaging the curves among all these neurons (Fig. 2a5,b5, black curves and bold red or blue curves, respectively). For statistical analysis, the Δ[Ca2+]i ∼ t curves of individual mice (e.g., m = 3–4) were further averaged to derive Δ[Ca2+]i ∼ t curves for each experimental group, and the results are summarized in Figure 3, in which the red and blue curves were the global Rhod2-[Ca2+]i fluorescence changes with their SDs in D1R and D2R neurons, respectively, for control mice (Fig. 3a) and for chronic mice (Fig. 3b). We also quantified the percentage of D1R-EGFP and D2R-EGFP neurons that showed attenuation of responses to cocaine within the field of view. A D1R-EGFP neuron from chronic mice was considered to have a downregulated response to cocaine if its increase was <50% of those in the D1R-EGF neurons of control mice; a D2R-EGFP neuron was considered downregulated if its decrease was <50% of those in the D2R-EGFP neurons of control mice. Similarly, a D1R-EGFP neuron or a D2R-EGFP neuron from chronic mice was considered to have an upregulated response if its increase or decrease was >50% that of control mice, respectively. The differences were assessed using a one-tailed Student's t test.
To compare the dynamic differences of Δ[Ca2+]i in D1R and D2R neurons between drug-naive and chronic mice, linear regression with least-squares fitting was applied to the Δ[Ca2+]i ∼ t curves of each group to determine their slopes (Luo et al., 2010; k = Δ[Ca2+]i/Δt), such as k(D1R) or k(D2R) and their ratio, k(D1R)/k(D2R) or Δ[Ca2+]i(D1R/D2R). We had previously shown that drug-naive mice exhibited biphasic Δ[Ca2+]i increases in D1R-EGFP neurons; thus, we used piecewise linear regression (e.g., two-piece fitting) to calculate the fast ka1(D1R) [phase 1, turning point (tp) ≤ 10 min] and the slow ka2(D1R) (phase 2, tp > 10 min) sections on the Δ[Ca2+]i ∼ t curve. The data collected before and after the tp for D1R-EGFP were used to quantify the slope ratio between the two phases r  k2/k1. This slope ratio r
k2/k1. This slope ratio r  k2/k1 was used to statistically evaluate whether the curves for D1R-EGFP and D2R-EGFP followed uniphasic (r → 1) or biphasic (r → 0) dynamics.
k2/k1 was used to statistically evaluate whether the curves for D1R-EGFP and D2R-EGFP followed uniphasic (r → 1) or biphasic (r → 0) dynamics.
Baseline [Ca2+]i assessment and correction for changes induced by chronic cocaine.
To assess whether baseline [Ca2+]i within D1R- and D2R-EGFP neurons changed after chronic cocaine exposure, we performed ex vivo imaging of [Ca2+]i fluorescence on brain slices (striatal region) labeled with Rhod2 in Drd1-EGFP or Drd2-EGFP mice (m = 3 for each group) with chronic cocaine exposures (30 mg/kg/d, i.p.; Table 1, Experiment 2) and those (m = 3) exposed to saline (0.1 cc/10 g/d, i.p.; Table 1, Experiment 3) for 2 weeks. The mice underwent the same surgical procedure as described above and were loaded with Rhod2/AM for intracellular Ca2+ labeling. After 1 h to allow for cellular uptake of Rhod2, the mice were perfused with 20 ml of PBS followed by 20 ml of formaldehyde (4%). Their heads were embedded in 4% formaldehyde for 24 h, and then the brains were removed and placed in sucrose solution (30%) until the solution was fully absorbed. The striatal region of the mouse's brain was cryosectioned to ∼10 μm slices and imaged by the dual-wavelength microscope setup (Fig. 1) except that the microprobe was removed. [Ca2+]i-Rhod2 fluorescence within D1R-EGFP or D2R-EGFP neurons was assessed over background autofluorescence using a custom MATLAB program, by which the influences of image location or setup parameters (e.g., exposure time) were eliminated (Pan et al., 2010). Differences in [Ca2+]i fluorescence intensities between the experimental groups were analyzed using Student's t test.
The percentage change of baseline [Ca2+]i induced by chronic cocaine exposure was defined as the coefficient γ, which was empirically determined through the ex vivo experiments (Experiments 2 and 3) by computing the ratio of baseline [Ca2+]i between chronic mice and control mice using the following linear equation:
 where Δ[Ca2+]t,bo % was the intracellular calcium change over the baseline of control mice before cocaine challenge (i.e., t ≤ 0 min), whereas Δ[Ca2+]t,b1 % was the intracellular calcium change over its baseline in chronic mice before cocaine challenge. γ was used to eliminate the influence of the baseline [Ca2+]i difference in chronic mice versus control mice for the quantification of the effects of cocaine challenge on Δ[Ca2+]i.
where Δ[Ca2+]t,bo % was the intracellular calcium change over the baseline of control mice before cocaine challenge (i.e., t ≤ 0 min), whereas Δ[Ca2+]t,b1 % was the intracellular calcium change over its baseline in chronic mice before cocaine challenge. γ was used to eliminate the influence of the baseline [Ca2+]i difference in chronic mice versus control mice for the quantification of the effects of cocaine challenge on Δ[Ca2+]i.
Results
The detailed animal experimental groups are categorized in Table 1, and the quantitative results of the in vivo imaging studies are summarized in Table 2 and Table 3.
[Ca2+]i dynamics in D1R-EGFP and D2R-EGFP neurons in chronic mice
Numbers of D1R cells and D1R cells downregulated by chronic cocaine exposure
Chronic cocaine changes the dynamic profile of [Ca2+]i in D1R but not in D2R
The dynamic features of Δ[Ca2+]i in response to acute cocaine exposure (8 mg/kg, i.p.) differed between control mice and chronic mice in D1R-EGFP neurons but not in D2R-EGFP neurons (Table 2). Figure 2a (top panels) shows dual-channel fluorescence images of Rhod2-[Ca2+]i (Fig. 2a1,a2) and D1R-EGFP (Fig. 2a3), and their merged image (Fig. 2a4), which allowed us to trace the temporal Δ[Ca2+]i profile (Fig. 2a5) of individual D1R-EGFP neurons (Fig. 2a5, black curves; n = 6) and their averaged changes (Fig. 2a5, bold red curve); the same strategy was used for D2R-EGFP neurons (Fig. 2b, bottom panels). Interestingly, whereas in control mice (dashed red curve) the Δ[Ca2+]i increase in D1R-EGFP neurons was biphasic, with an initial rapid increase to 7.6 ± 2.3% within tp = 8.8 ± 1.1 min of cocaine injection followed by a plateau, in chronic mice (Fig. 2a, bold red curve) the Δ[Ca2+]i increase was monophasic starting at t = 4 min after cocaine injection and gradually increasing throughout the recording period (t ≥ 30 min; Fig. 2a). In contrast, the dynamics of [Ca2+]i in D2R-EGFP neurons with acute cocaine was similar in control mice (Fig. 2b, dashed blue curve) and in chronic mice (Fig. 2b, bold blue curve), showing a progressive decrease in [Ca2+]i (Fig. 2b).

a, b, [Ca2+]i changes induced by acute cocaine injection (8 mg/kg, i.p.) in striatal D1R-expressing (a) and D2R-expressing (b) neurons in mice chronically exposed to cocaine (30 mg/kg/d, i.p., for 2 weeks), which are referred to as chronic mice. 1, 2, Δ[Ca2+]i images at t = −5 min and t = 25 min before and after cocaine injection, respectively. 3, EGFP image to identify striatal D1R-/D2R-expressing neurons. 4, [Ca2+]i overlapped with EGFP image to identify Δ[Ca2+]i from D1R/D2R neurons. 5, Time course of Δ[Ca2+]i increases within D1R neurons (black curves, n = 6) and their mean (bold red curve) or Δ[Ca2+]i decreases within D2R neurons (black curves, n = 6) and their mean (bold blue curve) in response to acute cocaine exposure (also included are dashed curves of previously reported control mice with no history of cocaine exposures before the acute administration (Luo et al., 2011) for comparison. 6, Comparison of mean [Ca2+]i changes at t = 25 min versus baseline (t ≤ 0 min) among the groups, showing less Δ[Ca2+]i increase for D1R in chronic mice (3.79 ± 1.32%) than in control mice (10.6 ± 3.2%) and less Δ[Ca2+]i decrease for D2R in chronic mice (−3.85 ± 1.55%) than in control mice (−10.4 ± 5.8%). In vivo simultaneous EGFP and Rhod2-[Ca2+]i imaging of striatal neurons was enabled by a microneedle approach (Fig. 1) that relayed the subcellular images in the mouse's striatum at the distal end of a microendoscope (φ, 1 × 25 mm; 0.65 NA) back to the focal plane of a modified epifluorescence microscope (E800, Nikon) to be acquired by a time-sharing EM CCD (i885, Andor).
To quantify the dynamics of the [Ca2+]i responses to acute cocaine, linear-curve fitting was applied to the temporal [Ca2+]i profile (i.e., Fig. 2a, bold red curve) of each group (i.e., averaged over m = 3–4 mice) to compute the slope (i.e., k = Δ[Ca2+]i/Δt). In control mice, the response was biphasic in striatal D1R-EGFP neurons (Fig. 3a, red curve) and was characterized by the following two different linear fittings (slopes): ka1(D1R) for phase 1 (t ≤ tp) and ka2(D1R) for phase 2 (t > tp). [Ca2+]i first increased rapidly with ka1(D1R) = 0.77 ± 0.2%/min in phase 1 (t ≤ tp = 8.8 ± 1.1 min, m = 4), and then plateaued with ka2(D1R) = 0.05 ± 0.05%/min in phase 2 (t > tp, m = 4) for the remaining recording period (∼22 min; Luo et al., 2011). In chronic mice, the temporal response to acute cocaine exposure in D1R-EGFP neurons showed an ∼4 min delay and then a uniphasic continuous [Ca2+]i increase with kc1(D1R) ≈ kc2(D1R) = 0.44 ± 0.02% (m = 3) for the remaining recording period (Fig. 3b, red curve).

a, b, Dynamic changes in Δ[Ca2+]i induced by acute cocaine in striatal D1R-expressing neurons (red curve), D2R-expressing neurons (blue curve), and their ratios [Δ[Ca2+]i(D1R/D2R); bold black curves] in control mice (a) and chronic mice (b). Least-squares fitting (dashed lines) is applied to analyze the slope differences [i.e., k = d(Δ[Ca2+]i)/dt]. In control mice (a), Δ[Ca2+]i increases in striatal D1R neurons are biphasic (pink lines): an initial steep rise with ka1(D1R) in phase 1 (t ≤ 8.3 min) followed by a long-lasting, slow increase with ka2(D1R) in phase 2 (t > 8.3 min) and the Δ[Ca2+]i in striatal D2R neurons are uniphasic: a gradual decrease with ka(D2R) over t > 30 min. In chronic mice (b), Δ[Ca2+]i increases in D1R neurons have changed to uniphasic, with a gradual but long-lasting rise with kc(D1R), and those of D2R neurons remain uniphasic with a gradual decrease. As a result, Δ[Ca2+]i(D1R/D2R) in control mice rapidly increases to a climax at t ≈ 5–7 min, but it then returns to baseline; whereas in chronic mice Δ[Ca2+]i(D1R/D2R) progressively increases and does not return to baseline.
In control mice, the response to acute cocaine exposure in D2R-EGFP neurons (Fig. 3a, blue curve) was uniphasic, showing a continuous decrease (over t > 30 min) with ka(D2R) = −0.40 ± 0.13%/min. In chronic mice (Fig. 3b), the dynamic features of Δ[Ca2+]i in D2R-EGFP neurons was also uniphasic with kc(D2R) = −0.12 ± 0.09%/min (m = 3) following cocaine injection and did not differ from that in control mice (Fig. 3a).
Chronic cocaine exposure reduces baseline [Ca2+]i fluorescence in D2R-EGFP neurons
To assess differences in baseline [Ca2+]i between control mice and chronic mice, we measured baseline [Ca2+]i fluorescence levels ex vivo in the striatal D1R-EGFP and D2R-EGFP neurons (Table 1, Experiments 2 and 3). Baseline [Ca2+]i-Rhod2 fluorescence in striatal D1R-EGFP neurons (Fig. 4, red bar) was similar in control mice ([Ca2+]i/b = 1.41 ± 0.08, m = 3) and chronic mice ([Ca2+]i/b = 1.29 ± 0.06; m = 3, p = 0.3), but in D2R-EGFP neurons (Fig. 4, blue bar) it was significantly higher in control mice ([Ca2+]i/b = 1.78 ± 0.15; m = 3) than in chronic mice ([Ca2+]i/b = 1.35 ± 0.24, m = 3, p = 0.04).
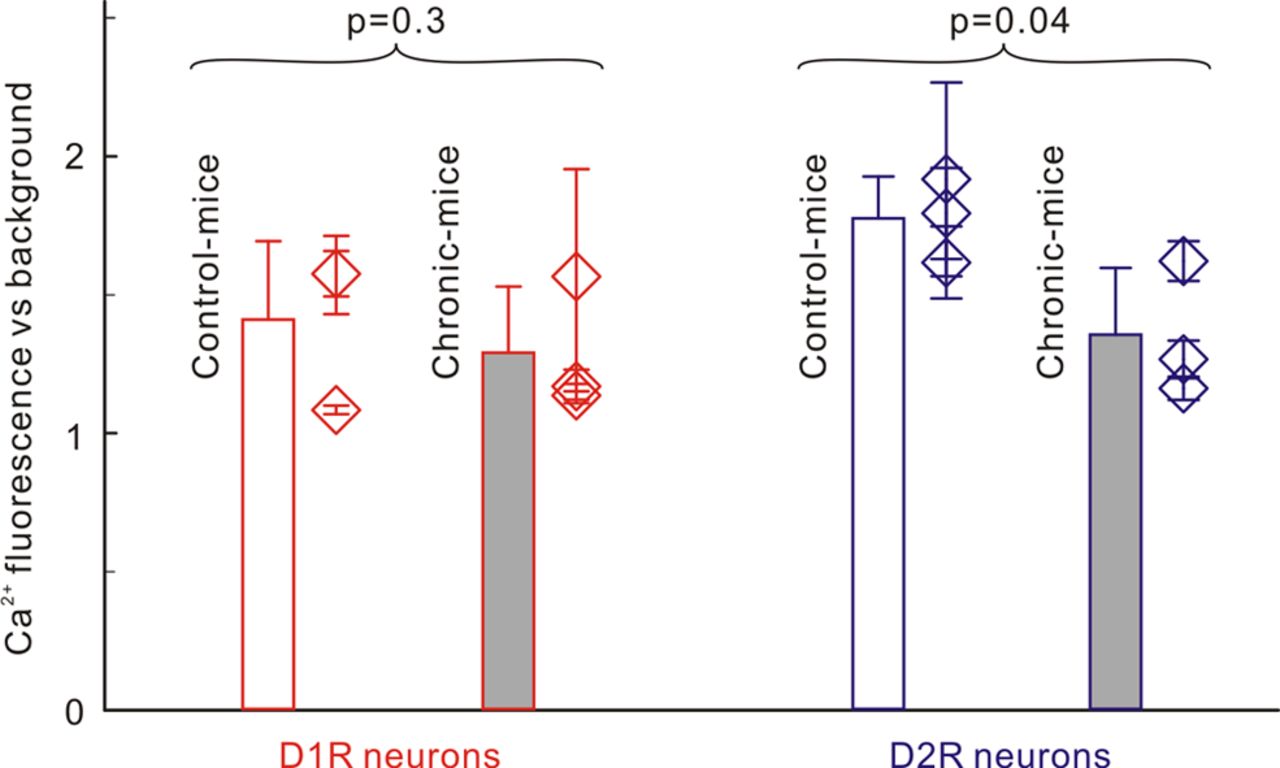
Comparison of baseline [Ca2+]i-Rhod2 fluorescence levels (∝[Ca2+]i) between control mice and chronic mice. Red/blue lines are for D1R/D2R neurons, and white/gray bars are for saline- versus cocaine-treated groups. [Ca2+]i in D1R neurons did not differ significantly (p = 0.3) between control mice (1.41 ± 0.08) and chronic mice (1.29 ± 0.06), whereas [Ca2+]i in D2R neurons in chronic mice (1.35 ± 0.24) was significantly lower (p = 0.04) than in control mice (1.78 ± 0.15).
Chronic cocaine exposure reduces acute cocaine-induced Δ[Ca2+]i amplitudes in D1R- and D2R-EGFP neurons
To compare the amplitudes of [Ca2+]i changes (i.e., Δ[Ca2+]i) induced by acute cocaine exposure between control mice and chronic mice, we measured the mean Δ[Ca2+]i at t = 25 min after cocaine injection (Fig. 2). In D1R-EGFP neurons, the [Ca2+]i increases induced by acute cocaine exposure were significantly higher in control mice (Δ[Ca2+]i = 10.6 ± 3.2%, m = 4) than in chronic mice (Δ[Ca2+]i = 3.79 ± 1.32%, m = 4, p = 0.009). In D2R-EGFP neurons, the [Ca2+]i decreases induced by acute cocaine were significantly larger in control mice (Δ[Ca2+]i = −10.4 ± 5.8%, m = 4) than in chronic mice (Δ[Ca2+]i = −3.85 ± 1.55%, m = 3, p = 0.005).
To correct for the baseline [Ca2+]i level differences (Fig. 4) between control mice and chronic mice (8.5 ± 9.5% lower for D1R-EGFP neurons and 24.2 ± 18.1% lower for D2R-EGFP neurons), we recomputed the Δ[Ca2+]i changes induced by chronic cocaine use by taking into account these lower [Ca2+]i baseline levels in chronic mice, according to Equation 1. This revealed that the attenuation of Δ[Ca2+]i amplitude corresponded to 67 ± 16% in D1R-EGFP neurons (i.e., from 10.6 ± 3.2% to 3.47 ± 1.21%; m = 3) and to 72 ± 17% in D2R-EGFP neurons (i.e., from −10.4 ± 5.8% to −2.93 ± 1.18%, m = 3).
We also quantified the percentage of D1R-EGPF and D2R-EGFP neurons in the striatum of the chronic mice that showed attenuated responses to cocaine versus the total number of D1R-EGFP and D2R-EGFP neurons within the field of view. These results showed that ∼75% of neurons showed attenuated responses to cocaine in D1R-EGFP neurons (i.e., 76.4 ± 15.5%) and in D2R-EGPF neurons (i.e., 74.7 ± 17.5%), and the percentages did not differ between these two neuronal populations (Table 3). No neurons with enhanced responses were detected.
Chronic cocaine unbalances the D1R/D2R Δ[Ca2+]i signaling ratio
To assess the dynamic interaction of D1R and D2R signaling in response to acute cocaine, we computed the ratio of Δ[Ca2+]i between D1R-EGFP and D2R-EGFP neurons (i.e., Δ[Ca2+]i(D1R/ D2R) in striatum as a function of time after acute cocaine injection. In control mice, Δ[Ca2+]i(D1R/D2R) initially increased and reached a peak at t ≈ 6–8 min (Fig. 3a, bold black curve), after which it returned to baseline at t ≈ 17–20 min after cocaine administration. In contrast, in chronic mice Δ[Ca2+]i(D1R/D2R) continuously increased in response to acute cocaine challenge (Fig. 3b, bold black curve). Thus, chronic cocaine exposure shifted the fast and short-lasting predominance of D1R signaling (<6–8 min followed by a balanced D1R/D2R signaling, D1R = D2R) during cocaine intoxication observed in control mice, into a sustained predominance of D1R signaling (>30 min after acute cocaine injection).
D1R and D2R antagonists suppress Δ[Ca2+]i in D1R- and D2R-EGFP neurons in chronic mice
To assess whether blockade of D1R interfered with cocaine-induced [Ca2+]i increases, we pretreated D1R-EGFP chronic mice with SCH23390 (0.075 mg/kg, i.p.), a D1R selective antagonist, at t = −30 min before acute cocaine injection (Table 1, Experiment 4). SCH23390 blocked cocaine-induced [Ca2+]i increases in D1R-EGFP neurons in striatum (i.e., from 3.79 ± 1.32% to 0.27 ± 1.95%, m = 3, p = 0.03; Fig. 5a,c, top panels), thus further confirming that cocaine-induced [Ca2+]i increases reflected activation of D1R-EGFP neurons in striatum. In addition, to analyze potential cross-talk effects from D2R antagonism on striatal D1R-EGFP neurons in chronic mice, we pretreated D1R-EGFP mice with raclopride (3 mg/kg, i.p.) at t = −30 min before cocaine administration. Raclopride also inhibited cocaine-induced [Ca2+]i increases in D1R-EGFP neurons (i.e., from 3.79 ± 1.32% to 0.46 ± 1.96%, m = 4, p = 0.02; Fig. 5b,c, top panels).

I, Top panels, II, Right panels, Dynamic [Ca2+]i changes induced by acute cocaine (8 mg/kg, i.p.) in striatal D1R-expressing neurons (I, top panels) and D2R-expressing neurons (II, right panels), with D1R antagonist (SCH23390, i.p.; a) and with D2R antagonist (raclopride, i.p.; b) in chronic mice. 1, 2, Δ[Ca2+]i images at t = −5 min and t = 25 min before and after cocaine injection, respectively. 3, EGFP image to identify striatal D1R-expressing (I) or D2R-expressing (II) neurons. 4, [Ca2+]i overlapped with EGFP image to identify Δ[Ca2+]i from D1R (I) or D2R (II) neurons. 5, Δ[Ca2+]i increases within D1R (I) or decreases within D2R (II) neurons (black curves, n = 5, 6, respectively) and their mean time course (bold red or blue curves) in response to acute cocaine. c, Comparison of mean Δ[Ca2+]i increases at t = 25 min versus baseline (t ≤ 0 min) among groups. D1R (I) shows less Δ[Ca2+]i increase in chronic mice (3.79 ± 1.32%) than in the control mice (10.6 ± 3.2%), which was further reduced by D1R and D2R antagonists to 0.2 ± 1.8% and 0.46 ± 1.96%, respectively. D2R (II) shows less Δ[Ca2+]i decrease in chronic mice (−3.85 ± 1.55%) than in control mice (−10.4 ± 5.8%), which was further reduced by D2R and D1R antagonists to −0.23 ± 0.23% and −1.07 ± 1.26%, respectively.
The same pharmacological interventions were applied to D2R-EGFP chronic mice to assess the effects of D2R and D1R antagonism on cocaine-induced [Ca2+]i decreases in D2R-EGFP neurons (Table 1, Experiment 5). Raclopride blocked cocaine-induced [Ca2+]i decreases from −3.85 ± 1.55% to −0.23 ± 0.23%, m = 3, p = 0.009; Fig. 5b,c, bottom panels). Pretreatment with SCH23390 partially inhibited cocaine-induced [Ca2+]i decreases from −3.85 ± 1.55% to 1.07 ± 1.26% (m = 3, p = 0.02; Fig. 5a,c, bottom panels).
Comparisons of the cross-talk responses in control mice and in chronic mice showed that there were no differences in the responses to raclopride in D1R-EGFP neurons between the control mice (from 10.6 ± 3.2% to 2.7 ± 0.3%) and the cocaine mice (from 3.79 ± 1.32% to 0.46 ± 1.96%); similarly, the blockade by SCH23390 of cocaine-induced [Ca2+]i decreases in D2R-EGFP neurons did not differ in chronic mice (from −3.85 ± 1.55% to −1.07 ± 1.26%) and in control mice (from −10.4 ± 5.8% to −0.5 ± 0.4%).
Discussion
Here we show that chronic cocaine exposure is associated with attenuated intracellular Ca2+ changes (reflecting neuronal activity) induced by acute cocaine exposure in both D1R- and D2R-EGFP neurons in striatum, and that it changes the dynamic responses to cocaine in D1R- but not D2R-EGFP neurons. In D1R-EGFP neurons, cocaine-induced Ca2+ increases were attenuated and the dynamics changed from that of a fast but short-lasting increase into a slower but long-lasting increase. In D2R-EGFP neurons, the marked attenuation of Ca2+ decreases was stable throughout the time of measurement. The dynamic changes in D1R-EGFP neurons resulted in a shift from short-lasting predominance of striatal D1R over D2R signaling (<8 min) in control mice into a sustained predominance of D1R over D2R signaling (>30 min) after cocaine injection in chronic mice (Fig. 3).
To our knowledge, this is the first report of decreases in activity of both striatal D1R- and D2R-expressing neurons during cocaine intoxication in vivo in mice exposed chronically to cocaine. Our findings of attenuated D1R signal with chronic cocaine exposure differ from prior studies reporting enhanced striatal D1R signaling, an effect associated with cocaine sensitization (Pascoli et al., 2012). This discrepancy is likely to reflect methodological differences; specifically, we measured the attenuation of D1R by cocaine (monitoring changes in intracellular Ca2+) as opposed to electrophysiological responses of D1R-exressing neurons to high-frequency stimulation. Also, sensitization responses were measured several days after the last cocaine administration and are associated with changes in synaptic expression of AMPA receptors in MSNs (Boudreau et al., 2007; Kourrich et al., 2007), whereas our measurements were performed while mice were on cocaine and 24 h after the prior cocaine dose. On the other hand, our findings are consistent with those from a study that reported reduced membrane excitability of D1R-expressing MSNs in a brain slice preparation in the nucleus accumbens in mice previously exposed to repeated cocaine injections (5 days), though the membrane excitability of D2R-expressing MSNs was not affected (Kim et al., 2011) when compared with drug-naive mice. Also, in nonhuman primates repeated cocaine exposure was associated with reductions in D1R levels in striatum (Moore et al., 1998), though human studies failed to observe this (Martinez et al., 2009), and thus it is unclear whether chronic cocaine exposure reduces the levels of D1R in striatum.
While we predict that D2R-expressing neurons are downregulated by chronic cocaine exposure, the basal calcium levels in D2R-expressing neurons are downregulated. This is likely to reflect the regulation of calcium levels by other receptors in addition to D2R (e.g., A2A receptors, which colocalize in D2R-expressing neurons opposing D2R signaling; Ferré et al., 1993; Svenningsson et al., 1999).
Our results in D2R-expressing neurons are consistent with studies showing decreases in striatal D2R signaling with chronic cocaine exposure (baseline studies), including preclinical studies reporting reductions in striatal D2R levels (Nader et al., 2006; Thanos et al., 2007) and clinical studies showing reductions in striatal D2R in cocaine abusers (for review, see Volkow et al., 2009). In human addicts, the reduction in striatal D2R has been implicated in compulsive drug intake and impulsivity (Volkow et al., 1993, 2006, 2007). Similarly, an in vitro study reported attenuation of cocaine-induced Ca2+ decreases in D2R-expressing neurons in nucleus accumbens with chronic cocaine exposure (Perez et al., 2011), and imaging studies have reported marked reductions in stimulant-induced DA increases in cocaine abusers (for review, see Volkow et al., 2009).
Since activation of striatal D1R- or inhibition of striatal D2R-expressing neurons enhances cocaine reward (Hikida et al., 2010; Lobo et al., 2010; Ferguson et al., 2011) and upregulation of D2R interferes with cocaine consumption (Thanos et al., 2008), an imbalance between the stimulatory direct D1R pathway and the inhibitory D2R indirect pathway during cocaine intoxication would be expected to facilitate compulsive cocaine intake even when DA signaling is markedly attenuated (for review, see Volkow et al., 2009). This would be akin to the improvement in movement observed in patients with Parkinson's disease after surgical lesioning of the subthalamic nucleus (to remove control from the D2R-modulated indirect pathway, which inhibits locomotor activity), even when they have very low DA levels (Alvarez et al., 2009). So, while we showed a significant decrease in signaling through the striatal D1R direct pathway with chronic cocaine exposure, the longer-lasting inhibition of the striatal D2R indirect pathway during cocaine intoxication could analogously be expected to allow the direct pathway to drive the behavior of cocaine reward and consumption.
In this study, we also showed that chronic cocaine exposure disrupted the dynamics of cocaine-induced Ca2+ increases in D1Rs from an abrupt increase followed by a leveling off to a delayed and progressive increase in D1R signaling. Though its behavioral significance is unclear, we postulate that it might result in an attenuation of reward, since the speed at which drugs stimulate DA signaling modulates their rewarding effects (Beardsley and Balster, 1993; Volkow et al., 1995, 2000; Samaha and Robinson, 2005). We also postulate that the long-lasting predominance of D1R over D2R signaling might result in enhanced motivational drive for the drug despite a decrease in reward intensity. This is because D1R, which is stimulatory and has a low affinity for DA, will be activated when exposed to large DA increases, as occur during cocaine intoxication, whereas D2R, which is inhibitory and has a high affinity for DA, will be activated both by the sharp DA increases as well as by the lower tonic DA levels that follow (Beaulieu and Gainetdinov, 2011). Thus, the relative predominance of D1R over D2R following cocaine administration (as seen in control mice) probably contributes to the dynamics of the behavioral effects of cocaine (Volkow et al., 1997a). That is, immediately after cocaine injection, full signaling from D1R and D2R is expected, but as DA levels start to decline then D1R signaling would decrease faster than D2R, eventually resulting in a D2R predominance and a consequent decrease in the rewarding effects of cocaine. Indeed, in control mice, acute cocaine exposure abruptly stimulated striatal D1R-expressing neurons, whereas it slowly and continuously inhibited D2R-expressing neurons. Thus, the persistent predominance of D1R signaling seen in the chronic mice might sustain the incentive and motivation for cocaine intake without the opposing effects of striatal D2R signaling.
The molecular mechanisms underlying the attenuated responses to cocaine in D2R- and D1R-expressing MSNs during intoxication are unclear. They could reflect blunted cocaine-induced DA increases, as shown by neuroimaging studies reporting reduced DA increases to stimulant drugs in cocaine abusers (Volkow et al., 1997b, 2011; Martinez et al., 2007). In D2R-expressing MSNs, they could also reflect downregulation of D2R levels with chronic cocaine exposure, as reported by prior studies (for review, see Volkow et al., 2012). For D1R-expressing MSNs, this is less clear since studies have reported increases, decreases, or no changes in D1R levels with chronic cocaine exposure (for review, see Volkow et al., 2012). As for the mechanisms underlying the differential effects of chronic cocaine use on the dynamic changes in D1R versus D2R signaling, we can only speculate that they could reflect interactions with other receptors (i.e., D3R, which heteromerizes with D1R; Marcellino et al., 2008), that they are upregulated with chronic cocaine use (Staley and Mash, 1996), and that differential changes occur in synaptic plasticity in D1R versus D2R-expressing MSNs with chronic cocaine use.
The cross talk between D1Rs and D2Rs observed with the administration of the D1R antagonist in D2R-EGFP neurons and the D2R antagonist in D1R-EGFP neurons is consistent with synergism between D1R- and D2R-expressing neurons in striatum (LaHoste et al., 2000), including interactions of both receptors within striatal neurons that coexpress D1R and D2R (Lester et al., 1993; Bertran-Gonzalez et al., 2008; Matamales et al., 2009, Rashid et al., 2007; Aizman et al., 2000). Interestingly, raclopride was more effective in blocking cocaine-induced D1R signaling than SCH23390 was in blocking D2R signaling. Though the mechanisms are unclear, we postulate that they might reflect the blockade by raclopride of D3R (Malmberg et al., 1994), which heteromerizes with D1R (Marcellino et al., 2008).
Limitations of this study include measurements in dorsal striatum rather than in nucleus accumbens, which is the brain region implicated in drug reward (Koob and Bloom, 1988). However, the dorsal striatum is implicated in the neuroadaptations associated with the transition from controlled to compulsive cocaine intake (for review, see Belin and Everitt, 2008; Volkow et al., 2012), and thus our findings are pertinent to addiction. In this study, we measured the dynamic changes over a 30 min period, but further studies are needed to determine whether dynamics in D2R-expressing MSN are affected by chronic cocaine exposure over a longer imaging period.
Here we show that chronic cocaine exposure markedly decreased the responses to acute cocaine exposure both in D1R- and D2R-expressing MSNs and changed the dynamics of their signaling during cocaine intoxication. Chronic cocaine exposure resulted in a sustained predominance of D1R over D2R signaling during cocaine intoxication, which, by removing the reward-decreasing effects of D2R stimulation, might leave the D1R direct pathway unopposed.
Footnotes
This research was supported in part by National Institutes of Health (NIH) Grants K25-DA021200 (C.D.), 1RC1DA028534 (Y.P., C.D.), R21DA032228 (Y.P., C.D.), and R01DA029718 (Y.P., C.D.); and by NIH intramural programs (N.D.V.).
The authors declare no financial conflicts of interest.
- Correspondence should be addressed to either of the following: Dr. Nora D. Volkow, Director, National Institute on Drug Abuse, National Institutes of Health, Bethesda, MD 20892, nvolkow{at}nida.nih.gov; or Dr. Congwu Du, Associate Professor, Department of Biomedical Engineering, Stony Brook University, Stony Brook, NY 11794, congwu.Du{at}stonybrook.edu





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}