

Abstract
We investigate stimulus specificity of repetition priming in a tractable model system; the feeding network of Aplysia. Previous studies primarily focused on an aspect of behavior that is altered during ingestive priming, radula opening. Priming of radula opening occurs when two modulatory peptides [feeding circuit activating peptide (FCAP) and cerebral peptide-2 (CP-2)] are released from the cholinergic command-like neuron cerebral buccal interneuron 2. Effects of FCAP/CP-2 on radula opening motor neurons are cAMP mediated. The present experiments sought to determine whether FCAP/CP-2 and cAMP are also involved in the priming of radula opening during an incompatible activity, i.e., during egestive motor programs. Egestive priming is induced when motor programs are triggered by afferents with processes in the esophageal nerve. We demonstrate that egestive priming is not FCAP/CP-2 mediated. Instead, it is induced by an unrelated peptide (small cardioactive peptide), which exerts PKC-mediated effects. Our data, therefore, suggest that different feeding motor programs are primed via actions of different sets of intercellular and intracellular substances. We suggest that this accounts for the stimulus specificity that can be characteristic of repetition priming. Different stimuli activate different central pattern generator inputs. These inputs release different modulators, which induce functionally distinct motor programs.
Introduction
Repetition priming is characterized by improved performance as behavior is repeated (Fowler et al., 1985; Cropper et al., 2014). Interestingly, work in both vertebrates and invertebrates has demonstrated that repetition priming is often stimulus specific (repeated presentation of a stimulus primes responses to that stimulus; Huettel et al., 2002; Proekt et al., 2004). How this is achieved at the network/cellular level remains an open question. One possibility is that it depends on chemical coding; the ability of different modulators to elicit distinct responses.
Behavioral sequences induced by modulators have been described. For example, egg laying hormone induces egg laying in Aplysia (Kupfermann, 1967, 1970; Heller et al., 1980) and eclosion hormone triggers molting in Manduca (Truman, 1978). Further, research at the network level has demonstrated that modulators can reconfigure activity to generate distinct motor programs (Marder and Bucher, 2007). A potential limitation of most of this research is that modulators were exogenously applied and “steady-state” effects characterized. This approach is relevant when modulators are blood borne and concentrations are relatively stable. We suggest that under other circumstances modulation will occur more dynamically. For example, modulators can be released from projection neurons that trigger episodic motor activity (Koh et al., 2003; Koh and Weiss, 2005) and modulatory effects may become cumulatively larger as behavior progresses (Cropper et al., 2014). This could lead to a stimulus-specific evolution of activity. Thus, a stimulus could activate a projection neuron that releases a specific modulator(s) thereby priming a specific type of motor activity. We present data in support of this idea.
Our experiments are conducted in the Aplysia feeding network. This network generates two incompatible behaviors; ingestion and egestion (Kupfermann, 1974). Earlier work focused on ingestion, demonstrating that feeding circuit activating peptide (FCAP) and cerebral peptide 2 (CP-2) play an important role in its induction (Koh et al., 2003; Koh and Weiss, 2007; Friedman and Weiss, 2010; Dacks and Weiss, 2013b). Both are present in an input to the feeding central pattern generator (CPG), the command-like neuron cerebral buccal interneuron 2 (CBI-2; Morgan et al., 2000; Koh et al., 2003). When motor programs are evoked by CBI-2, FCAP/CP-2 exert widespread effects, e.g., phase relationships of motor neurons change, and what was poorly defined activity becomes ingestive (Koh et al., 2003; Koh and Weiss, 2007; Friedman and Weiss, 2010; Dacks and Weiss, 2013b). This results in the priming of ingestion (e.g., progressive increases in bite strength). FCAP/CP-2 exert cAMP mediated effects (Friedman and Weiss, 2010).
Egestion also undergoes repetition priming (Proekt et al., 2004; Zhurov et al., 2005b), but for the most part underlying mechanisms are uncharacterized. We ask whether egestive priming is similarly induced by FCAP/CP-2. We demonstrate that it is not. It is induced by an unrelated peptide (small cardioactive peptideb; SCPb), which exerts PKC mediated effects. Our results provide support for the idea that there is chemical coding in the context of priming; i.e., different behaviors are primed via actions of different sets of intercellular and intracellular substances.
Materials and Methods
Animals.
Experiments were performed on Aplysia californica obtained from Marinus. Aplysia were maintained in circulating artificial seawater (ASW) made from Instant Ocean (Aquarium Systems), at 14°–15°C. Animals weighing 150–250 g were anesthetized by injection of approximately one-half their weight of isotonic MgCl2 (337 mm). Aplysia are hermaphrodites.
Cell identification.
Cerebral ganglia, buccal ganglia, and the attached buccal mass were removed with the cerebral-buccal connectives and buccal nerves intact. The buccal mass was separated from the distal digestive system at the buccal-esophageal junction and a portion of the buccal musculature was cut away to reveal the radula. Cerebral and buccal ganglia were pinned and desheathed in a Sylgard dish with the buccal mass pinned dorsal side down exposing the radula. B44 was identified by its location, and the nature of evoked movements (Church and Lloyd, 1994; Evans et al., 1996, 1999; Evans and Cropper, 1998; Jing and Weiss, 2001; Orekhova et al., 2001; Horn et al., 2004; Zhurov et al., 2005a).
After motor neuron identification, cerebral and buccal ganglia were transferred to a recording chamber with a volume of ∼1.5 ml ASW. Preparations were continuously superfused at 0.3 ml/min and maintained at 14°–17°C for 1 h before experiments. CBI-2 and B8 were identified based on location, size, and electrophysiological and morphological characteristics (Rosen et al., 1991; Church and Lloyd, 1994).
To summarize, identified neurons studied in these experiments are CBI-2 (a command-like neuron that triggers ingestive motor programs; Rosen et al., 1991), B8 (a radula closer motor neuron; Morton and Chiel, 1993a,b), B20 (a protraction phase interneuron that promotes egestive activity; Jing and Weiss, 2001), B40 (a protraction phase interneuron that promotes ingestive activity; Jing and Weiss, 2002), B48 (a radula opener motor neuron; Evans et al., 1996), and B65 (a protraction phase neuron that can promote egestive activity; Dacks and Weiss, 2013b).
Recording techniques.
Conventional intracellular and extracellular recording techniques were used. Intracellular electrodes were filled with 2 m K acetate and 100 mm KCl and beveled to 6–10 MΩ. Intracellular recordings were performed using either an Axoclamp 2B amplifier (Molecular Devices) or Getting 5A amplifier (Getting Instruments). Extracellular recordings from buccal nerves used suction electrodes constructed from polyethylene tubing. Signals were amplified by a CyberAmp 380 (Molecular Devices) and digitized using a Digidata 1200 (Molecular Devices) data acquisition system.
Excitability.
To measure excitability action potentials were elicited by injecting 3 s constant current depolarizing pulses with an interstimulus interval (ISI) of 60 s. Current size in individual experiments was selected to generate ∼10 spikes under control conditions. Preparations were superfused with a high-divalent ASW to reduce spontaneous activity and polysynaptic effects of peptides and other reagents. Six stable responses were obtained before each experiment. Washout periods were determined by the return to baseline activity and ranged from 20 to 60 min.
Motor program generation.
Motor programs were evoked by stimulating the esophageal nerve (EN), which elicits egestive motor programs both in semi-intact preparations and in the isolated nervous system (Chiel et al., 1988; Rosen et al., 1991; Jing and Weiss, 2002, 2005; Morgan et al., 2002; Due et al., 2004; Horn et al., 2004; Proekt et al., 2004; Zhurov et al., 2005a; Sasaki et al., 2008). To elicit programs the EN was stimulated continuously using 3 ms pulses at 2 Hz for 2 min. The pulse amplitude was adjusted to elicit approximately five successive cycles of activity. Two different stimulation paradigms were used. EN programs were elicited using either continuous stimulation at 2 Hz, which triggers programs with a short ISI (S-ISI), or were triggered with a long ISI (L-ISI; i.e., 3 min). Programs triggered with a S-ISI undergo repetition priming. Those triggered with a L-ISI do not. Programs evoked with a S-ISI were used to evaluate effects of endogenously released peptides. Those evoked with a L-ISI were used to evaluate effects of exogenous (bath applied modulators; e.g., to determine whether they mimicked effects of endogenous substances). Postpriming measurements were made 60 s after stimulation. After superfusion of reagents, data were collected 60 s after reagents filled the dish. Each experiment was performed in an individual preparation (i.e., without repeated stimulation or repeated application of reagents).
Motor program characterization.
Aplysia generate feeding-related behaviors that can be classified as ingestive, egestive, or intermediate activity (Kupfermann, 1974; Morton and Chiel, 1993a,b; Morgan et al., 2002). In all cases, the food-grasping organ (the radula) is initially protracted and then subsequently retracted. Behaviors differ in the relative timing of radula opening and closing. During ingestion the radula is open during protraction and closed during retraction, which pulls food into the buccal cavity (Morton and Chiel, 1993a,b). During egestion, the radula is closed during protraction and open during retraction. This pushes food out. The activity of the radula closer motor neuron B8 can be recorded both in vivo and in vitro. Consequently, it is used as a frame of reference to classify motor programs in vitro. During ingestive activity, B8 is predominantly active during retraction, whereas during egestive activity, it is predominantly active during protraction (Jing and Weiss, 2001, 2002; Morgan et al., 2002; Proekt et al., 2004; Jing et al., 2007; Friedman et al., 2009). In vitro, protraction is defined as the phase in which there is activity in the I2 nerve (which contains axons of radula protraction motor neurons; Hurwitz et al., 1996; Nargeot et al., 1999; Jing and Weiss, 2001, 2002; Morgan et al., 2002; Vilim et al., 2010). Retraction is defined as the phase in which high-frequency activity is recorded in buccal nerve 2 (after the cessation of activity in the I2 nerve; Morton and Chiel, 1993a,b; Nargeot et al., 1999; Sasaki et al., 2007, 2008, 2009; Wu et al., 2007). To classify motor programs we determined the B8 firing frequency during protraction and the B44 firing frequency during retraction (Morgan et al., 2002; Proekt et al., 2004, 2007; Friedman et al., 2009). With repeated S-ISI stimulation of the EN, intermediate programs become egestive as manifested by an increase in the B8 firing frequency during protraction, and an increase in the B44 firing frequency during retraction (Fig. 1A,B).

Feeding motor programs induced by EN stimulation. A, Plot illustrating progressive changes in the average firing frequency of B8 and B44 when five programs were generated by stimulating the EN with a S-ISI. The initial program triggered (Program 1) has intermediate characteristics. Programs 4 and 5 are egestive (as determined by monitoring the average B8 firing frequency during protraction and the average B44 firing frequency during retraction. B, The first and fifth program induced by stimulating the EN with a short ISI (after (Friedman et al., 2009).
Reagents.
Most experiments were conducted in ASW with the following composition (in mm: 460 NaCl, 10 KCl, 55 MgCl2, 11 CaCl2, and 10 HEPES buffer, pH 7.6). Excitability and occlusion experiments were conducted in high divalent ASW, (in mm: 368 NaCl, 10 KCl, 101 MgCl2, 13.8 CaCl2, and 10 HEPES, pH 7.6; Trudeau and Castellucci, 1992).
FCAP (SynPep; Sweedler et al., 2002), CP2 (SynPep; Phares and Lloyd, 1996; Vilim et al., 2001), and SCPb (Lloyd et al., 1986; Anaspec) were superfused at 10−6 m, a concentration that produces maximum effects (Koh et al., 2003). In all experiments, peptides were superfused at a rate of 0.3 ml/min.
Chelerythrine was dissolved in dimethyl sulfoxide (DMSO) and was then superfused at 10−5 m (a concentration used in other studies in Aplysia; Manseau et al., 1998; Zhou et al., 2014). A 0.2% DMSO ASW solution was used as a control. Phorbol ester TPA was first dissolved in DMSO, and then superfused at a relatively high concentration (10−5 m). Comparisons of different concentrations of phorbol esters in Aplysia have demonstrated that they are more potent at high concentrations but effects are still specific (Critz and Byrne, 1992). Inactive 4-α-phorbol ester was used as a control.
Statistics.
Digitized electrophysiological recordings were plotted using Corel DRAW v13 and data are expressed as mean ± SEM. Two-group statistical comparisons were performed using t test statistics. Data with more than two groups were first analyzed using an appropriate ANOVA. Subsequent comparisons used Bonferroni's multiple-comparison tests. The significance level was set at p < 0.05 (***p < 0.001; **p < 0.01; *p < 0.05; n.s., p > 0.05). All statistical tests were performed using SigmaPlot (Systat Software).
Results
If chemical coding is used to obtain input specificity of repetition priming we would expect that ingestive and egestive priming would be mediated by two different peptides or sets of peptides. During ingestive motor programs,priming is induced by FCAP and CP-2 (Koh et al., 2003; Koh and Weiss, 2007; Friedman and Weiss, 2010; Dacks and Weiss, 2013b). We sought to determine whether these peptides are also involved in egestive priming. Ingestive and egestive behavior differ in the phasing of the activity of radula opener and closer motor neurons (e.g., during ingestion the radula opens during protraction, whereas during egestion it opens during retraction; Morton and Chiel, 1993a,b). This study focuses on the control of radula opening. Radula opening is mediated by two different motor neurons, i.e., there is an ingestive motor neuron (B48) and an egestive motor neuron (B44; Friedman et al., 2009). Experiments conducted in parallel focused on B48 and demonstrated that FCAP/CP-2 produce ingestive priming by inducing an excitability increase in this cell (Friedman and Weiss, 2010). In principle, FCAP/CP-2 could also induce an excitability increase in B44, which would not translate into an increase in firing frequency during ingestion due to a lack of excitatory synaptic input. To determine whether FCAP/CP-2 affects the excitability of B44 we used procedures developed to study peptide effects on B48. We injected constant current pulses into B44 and adjusted pulse size so that ∼10 action potentials were evoked (i.e., either increases or decreases in excitability would be apparent). The preparation was superfused with a high divalent solution to reduce polysynaptic effects, and FCAP (Fig. 2A) and CP-2 (Fig. 2B) were exogenously applied. In the five preparations tested under each condition, peptide perfusion did not significantly change the number of evoked action potentials. Before FCAP, 11.2 ± 0.9 spikes were induced. After FCAP, 11.7 ± 1.4 spikes were induced (t(4) = 2.10, p = 0.21). Spikes (8.7 ± 0.7) were induced before CP-2 and spikes (10.2 ± 0.6) were induced after CP-2 (t(4) = 2.74, p = 0.10). Thus, although we cannot rule out a modest peptide effect that might be detected with a larger sample size, we concluded that FCAP/CP-2 were not likely to be the major peptides impacting B44 excitability.
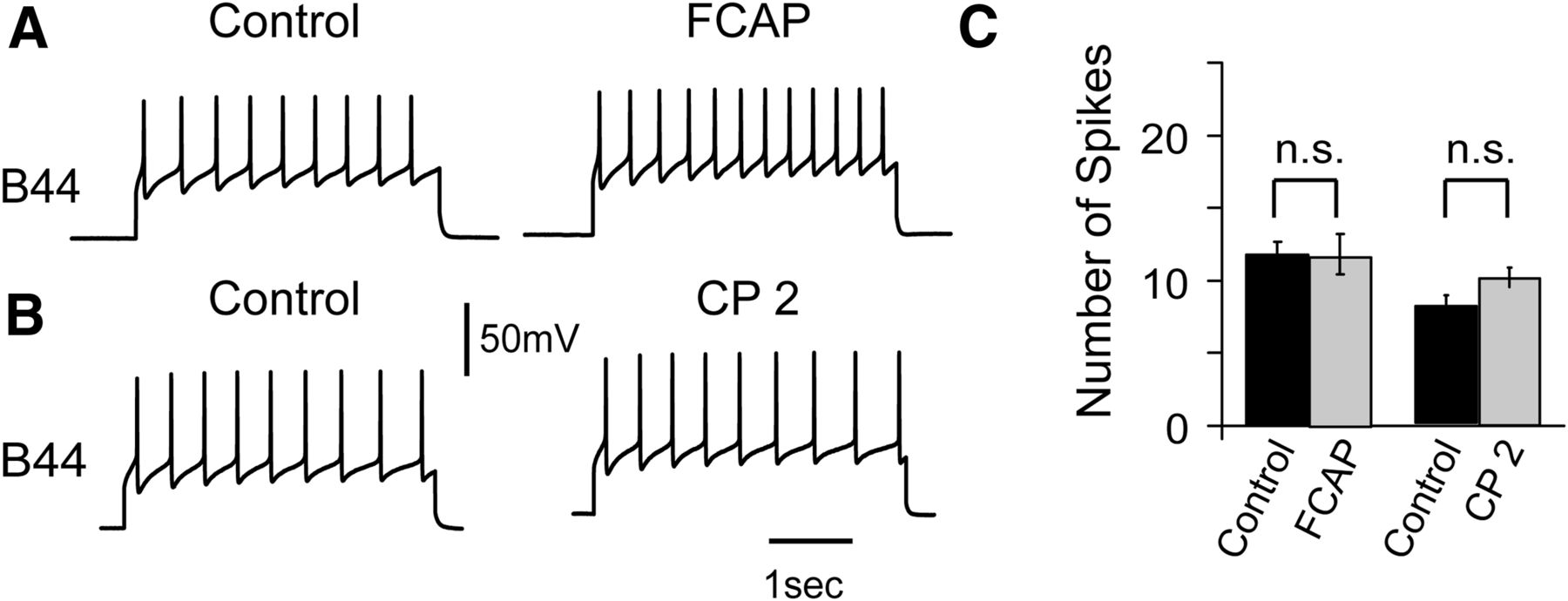
Effect of FCAP and CP2 on B44 excitability (as measured by injecting a 3 s depolarizing current pulse into B44 every 60 s). Experiments were conducted in a Hi-Di saline. A, FCAP (10−6 m) had no significant effect on the number of induced spikes. B, CP-2 (10−6 m) also had no effect on spike number. C, Group data (n = 5) from the experiments shown in A and B.
To identify a peptide that might contribute to egestive priming we focused on SCPb. Egestive priming is induced by EN stimulation, which releases SCPb (Wu et al., 2010). Further, under steady-state conditions exogenous SCPb can convert CBI-2 induced ingestive programs into egestive activity (Wu et al., 2010). To determine whether exogenous SCPb could mimic effects of EN-induced egestive priming, we stimulated the EN with a L-ISI. Under these conditions program reconfiguration does not occur, i.e., motor programs consistently have intermediate characteristics (Jing and Weiss, 2001; Morgan et al., 2002). SCPb superfusion modified characteristics of EN induced motor programs, making them egestive (Fig. 3A). SCPb superfusion preferentially increased the protraction phase firing frequency of the radula closing motor neuron B8 (Fig. 3 compare B1, B2). The firing frequency before SCPb was 2.4 ± 0.3 Hz, whereas after SCPb it was 5.8 ± 1.0 Hz. A repeated-measures one-way ANOVA revealed a significant overall difference between protraction phase firing before SCPb perfusion, in the presence of SCPb, and after SCPb washout (F(2,8) = 17.25; p < 0.01). Individual comparisons showed that in the presence of SCPb, the protraction phase firing frequency of B8 was higher than either the control or washout conditions (B8 protraction: control vs SCPb: t(4) = 6.05, p < 0.01; SCPb vs washout: t(4) = 8.19, p < 0.01; Fig. 3B1). In contrast SCPb had no significant effect on the B8 firing frequency during retraction as determined by a one-way ANOVA (F(2,8) = 0.69, p = 0.52) and individual comparisons (B8 retraction: control vs SCPb: t(4) = 1.05, p = 0.94; SCPb vs washout: t(4) = 0.98, p = 1; Fig. 3B2). The firing frequency before SCPb was 0.9 ± 0.4 Hz, whereas after SCPb it was 0.4 ± 0.2 Hz.

Effect of exogenous SCPb on L-ISI elicited EN programs. A, L-ISI programs were elicited by stimulating the EN every 3 min in the absence (control) and presence of 10−6 m SCPb. Black and white bars indicate the protraction and retraction phase of the motor program respectively. SCPb application increased the B8 firing frequency during protraction, increased the B44 firing frequency during retraction, and decreased the delay between the onset of retraction and the first action potential induced in B44. The latter is indicated by the double-headed arrows. B, Group data (n = 5) from the experiment shown in A showing the preferential increase in the B8 firing frequency during protraction (B1 vs B2) and the increase in the B44 firing frequency during retraction (B3).
To determine whether SCPb superfusion also modified activity of motor neurons that open the radula, we monitored changes in the activity of B44 (Fig. 3A,B3). There was a significant increase in the B44 firing frequency during retraction (F(2,8) = 15.97, p < 0.001). Thus the B44 firing frequency in the presence of SCPb was significantly different from the firing frequency under control or washout conditions (B44 retraction: control vs SCPb: t(4) = 4.60, p < 0.001; SCPb vs washout: t(4) = 8.41, p < 0.001; control vs washout: t(4) = 0.56, p = 0.59; Fig. 3B3). The firing frequency before SCPb was 3.8 ± 0.6 Hz, whereas after SCPb it was 8.3 ± 0.9 Hz. In addition, with SCPb application there was a significant decrease in the delay between the onset of retraction and the first action potential induced in B44. The delay was 4.4 ± 0.4 s before SCP, and 1.3 ± 0.2 s after SCP (t(4) = 6.17, p < 0.001; Fig. 3A). This indicates that SCP will decrease the latency of B44 activation, an effect also seen during egestive priming (Fig. 1B; Friedman et al., 2009).
We hypothesized that effects of EN stimulation were due to a direct effect on B44's excitability. To test this possibility we induced spiking in B44 by injecting depolarizing constant current pulses (as described above). After 5 min of EN stimulation with 3 ms current pulses at 3 Hz the number of B44 action potentials increased from 9.2 ± 2.0 to 15.4 ± 3.2 (Fig. 4A1). To determine whether this increase is statistically significant, a one-way ANOVA was performed (B44: F(2,8) = 6.0, p < 0.05). Individual comparisons revealed that B44 excitability was significantly higher following EN stimulation (control vs post-EN: t(4) = 4.2, p < 0.05; post-EN vs recovery: t(4) = 4.23, p < 0.05; control vs recovery: t(4) = 0.47, p = 0.67). In addition to its effect on excitability we also found that EN stimulation altered (increased) the input resistance of B44 (Fig. 4A2). For example, at 5 nA we observed a depolarization of 59.7 ± 2.4 mV before EN stimulation, and a depolarization of 78 ± 3.1 mV after EN stimulation (Wilcoxon sign rank test: W = 21, p = 0.034). Further, significantly smaller current injections were required to elicit spiking. For example at 6 nA 0.7 ± 0.3 spikes were triggered before EN stimulation whereas 3.8 ± 0.4 spikes were triggered after EN stimulation (Wilcoxon sign rank test: W = −21, p = 0.034).

Effects of EN stimulation and SCPb superfusion on the excitability and input resistance of B44. B44 excitability was determined by injecting a 3 s current pulse every 60 s (A1). To determine changes in input resistance B44 was held at −80 mV and a series of 2 s pulses of 1, 2, 5, and 6 nA were injected (e.g., A2). A, EN stimulation increased the B44 excitability (A1; n = 6) and input resistance (A2; n = 6). Further, the current needed to induce spiking was reduced, i.e., the 6 nA current injection did not induce spiking in the control condition but did after EN stimulation (A2). B, Effects of SCPb superfusion at 10−6 m. SCPb also increased the B44 excitability (B1; n = 6) and input resistance (B2; n = 6). Again, the current needed to induce spiking was also reduced, i.e., the 6 nA current injection did not induce spiking in the control condition but did after SCPb (B2).
If EN induced increases in B44 excitability are SCPb mediated, we would expect exogenous SCPb to mimic effects of EN stimulation. We found that this was the case. SCPb increased the number of B44 action potentials induced by injection of depolarizing pulses from 9.7 ± 1.3 to 16.6 ± 1.1 (Fig. 4B1). To determine whether changes in the number of action potentials were significant, we performed a one-way ANOVA (B44: F(2,8) = 7.53, p < 0.05). Individual comparisons revealed that B44 excitability was significantly higher in the presence of SCPb (control vs SCPb: t(4) = 3.68, p < 0.05; SCP vs recovery: t(4) = 3.04, p < 0.05; control vs recovery: t(4) = 0.67, p = 0.54). Additionally we found that SCPb superfusion increased the B44 input resistance. For example, at 5 nA we observed a depolarization of 26.2 ± 3.1 mV before application of SCPb, whereas after SCP we observed a depolarization of 42.7 ± 5.3 mV (Wilcoxon sign rank test; W = 21, p = 0.036). We also observed a decrease in the amount of current needed to induce spiking (Fig. 4B2). For example, at 5 nA 0.8 ± 0.02 spikes were induced before SCPb perfusion and 1.7 ± 0.3 were induced afterward.
Further, we would expect that exogenous SCPb would occlude effects of EN stimulation (Swensen et al., 2000; Koh and Weiss, 2005, 2007). To test this possibility SCPb was exogenously applied, which increased excitability (as expected; Fig. 5A1,B). In the continued presence of SCPb, we then reduced the size of the current pulse so that ∼10 action potentials were induced (excitability was similar to the control condition; Fig. 5A2, left). We then stimulated the EN (using parameters that increase B44 excitability in the absence of SCPb). In the presence of SCPb EN stimulation had no effect (i.e., 12.2 ± 0.7 spikes were induced before EN stimulation, whereas 12.4 ± 2.4 were induced after EN stimulation (t(8) = 1.96, p = 0.09; Fig. 5A2, right, B). Together these data indicate that EN induced increases in B44 excitability result from SCPb release.

Effect of EN stimulation on B44 excitability is occluded in the presence of SCPb. A, B44 excitability was determined by injecting a 3 s current pulse every 60 s. SCPb superfusion increased the B44 excitability (A1). To prevent a ceiling effect, the size of the current pulse was decreased (I ↓) to reduce the number of evoked spikes to control levels (A2, left). EN stimulation no longer increased the B48 excitability (A2, right). B, Group data (n = 9) for the experiment shown in A.
To determine whether effects of SCPb are like those of FCAP/CP-2 in that they are cAMP mediated we performed experiments in which we injected 8-br-cAMP into B44 and measured the change in excitability using protocols previously developed in experiments with B48 (Friedman and Weiss, 2010). In these experiments, B44 was impaled with three separate electrodes. The first electrode was used to inject a depolarizing 3 s constant current pulse. The second electrode monitored the resulting change in membrane potential, and the third electrode was used to iontophoretically inject 8-br-cAMP (10 mm) for 15 min. Intracellular injection of 8-br-cAMP had no significant effect on excitability (data not shown; t(4) = 1.5, p = 0.21). Namely, 10.4 ± 1.8 spikes were induced before injection, and 12.7 ± 3.9 were induced afterward.
Subsequent experiments sought to determine whether SCPb effects might be mediated by a different second messenger. Several lines of evidence implicate PKC. First, we found that perfusion of the PKC activator, phorbol ester TPA mimicked effects of SCPb (Fig. 6A1,A3). We demonstrated this in experiments in which we used the excitability paradigm described above and bath applied TPA. This significantly increased the number of action potentials evoked by depolarizing pulses (t(4) = 9.76, p < 0.001). Namely, 10.0 ± 0.5 spikes were triggered before TPA, whereas 16.0 ± 1.3 were triggered after TPA. In contrast, in four preparations inactive phorbol ester did not alter the B44 excitability (t(3) = 0.45, p = 0.66; data not shown). Further, we used the occlusion paradigm described above, and demonstrated that TPA occludes effects of SCPb superfusion (Fig. 6A2,A3), i.e., SCPb did not produce a significant increase in the number of action potentials induced by constant current injection when it was applied in the presence of phorbol-ester-TPA (t(4) = 0.44, p = 0.57). Thus, 10.2 ± 0.4 spikes were induced before SCPb was applied, and 10.8 ± 1.7 were induced in the presence of SCPb. Finally, to confirm that the PKC pathway was necessary as well as sufficient for both SCPb and EN induced changes in B44 excitability, we performed experiments in the presence of chelerythrine, a PKC inhibitor. Under these conditions EN stimulation and SCPb application both had no effect on excitability (Fig. 6B1,B2,B3,B4; for EN stimulation: t(4) = 0.62, p = 0.57; for SCPb superfusion; t(4) = 0.48, p = 0.66). Thus, 8.8 ± 1.9 spikes were triggered before EN stimulation and 8.7 ± 1.3 were triggered following EN stimulation. Before SCPb, 9.4 ± 0.7 spikes were triggered. After SCPb, 9.8 ± 1.7 spikes were triggered. Together these data indicate that EN (and SCPb) effects on B44 excitability are PKC mediated.

Effect of EN stimulation and SCPb are PKC mediated. A, Experiments with phorbol ester-TPA. Phorbol ester-TPA superfusion at 10−5 m increases the B44 excitability (A1; n = 5). Further, it occludes effects of SCPb superfusion (A2). To demonstrate occlusion, the size of the current pulse injected was decreased (I ↓) to reduce the number of evoked spikes to control levels (A2, left). SCPb no longer produced an excitability increase (A2, right). Group data (n = 5) are shown in A3. B, EN stimulation (B1, B2; n = 5) and SCPb superfusion (B3, B4; n = 5) did not produce increases in B44 excitability in the presence of 10−5 m chelerythrine.
Discussion
This study focuses on egestive priming in the feeding network, which for the most part has not been studied at the cellular/molecular level. A question we address is as follows: are effects of egestive priming similar to those of ingestive priming in that they are FCAP/CP-2 and cAMP mediated? To address this issue we focused on an aspect of behavior where effects of FCAP/CP-2 have been studied, radula opening. We demonstrate that FCAP/CP-2 do not significantly modify the excitability of the opener motor neuron used during egestion, B44. Instead excitability is increased by SCPb. SCPb is unrelated to either FCAP or CP-2. The peptides are structurally dissimilar (Lloyd, 1982; Morris et al., 1982; Phares and Lloyd, 1996; Sweedler et al., 2002). Further, they derive from different precursor proteins (Mahon et al., 1985; Vilim et al., 2001; Sweedler et al., 2002). We show that egestive priming during EN stimulation is SCPb-mediated. Further, we demonstrate that SCPb acts via PKC. We conclude that ingestive and egestive priming of radula opening are mediated by different intercellular and intracellular signaling molecules.
Will these findings apply to other loci in the feeding network?
When Aplysia feed there are two components to radula movements (Kupfermann, 1974). The radula protracts and retracts, and opens and closes. Comparisons of ingestion versus egestion have demonstrated that the two forms of “feeding” differ in the phasing of radula opening and closing (Morton and Chiel, 1993a,b). Radula protraction precedes radula retraction in both cases. However, during egestion the radula is closed as it protracts, and opens as it retracts. This pushes food out of the buccal cavity. During ingestion, the radula is open as it protracts and closed as it retracts. This pulls food in. Thus, because feeding motor programs differ in the timing of opening and closing movements, studies that have sought to determine how ingestive versus egestive activity is generated have focused on this component of behavior.
Although this study focuses on radula opening, data suggest that a similar difference in signaling systems may be responsible for the control of radula closing. Exogenous FCAP induces motor programs that have an ingestive pattern of activity in radula closer motor neurons (i.e., the B8 neurons; Sweedler et al., 2002). In contrast, exogenous SCPb converts ingestive B8 activity into egestive activity (Wu et al., 2010). Radula closing is more complex than radula opening in that B8 activity is at least in part under interneuronal control (Jing and Weiss, 2001, 2002; Jing et al., 2003; Dacks and Weiss, 2013a; Sasaki et al., 2013; Svensson et al., 2014; Wu et al., 2014). The neuron B20 produces an egestive pattern of B8 activity, and the neuron B40 produces an ingestive pattern of activity. Exogenous SCPb increases the excitability of B20 and potentiates B20-B8 synaptic transmission (Wu et al., 2010). In contrast, SCPb decreases the excitability of B40 and depresses B40 induced PSPs in B8 (Wu et al., 2010). Thus, although the quoted studies have been done under “steady-state conditions” (and not during repetition priming), data suggest that our results may generalize to radula closing, the other aspect of feeding that is reconfigured to generate ingestive versus egestive activity.
Chemical coding in the feeding network
Our results are consistent with the idea that a form of “chemical coding” is used in the selection of feeding motor programs. The idea that an endogenous chemical can trigger a complex behavior dates back to at least the 1970s. For example, research of this period demonstrated that egg laying in Aplysia could be triggered by injection of bag cell extracts (Kupfermann, 1967, 1970). Subsequent work identified the active ingredient in these extracts as egg laying hormone (Heller et al., 1980). Truman (1978) demonstrated that molting motor programs in the hawkmoth Manduca could be triggered by eclosion hormone. Decades of research on the crustacean stomatogastric nervous system have demonstrated that different modulatory neurotransmitters select different functional circuits in the stomatogastric ganglion (STG; Marder et al., 2014).
In some situations where a behavior (or motor program) is specified by chemical input, the substance in question is blood borne. The arrangement described in the present study obviously differs, i.e., FCAP/CP2 are released from the command-like neuron CBI-2 (Koh et al., 2003), and SCPb is released from afferents with processes in the EN (Wu et al., 2010). This also occurs in other systems, e.g., octopamine triggers flight in locusts, presumably via release from the dorsal unpaired median cells (Sombati and Hoyle, 1984). However, a further distinction arises from the fact that FCAP/CP2 and SCPb are present as cotransmitters in the inputs to the feeding CPG (in this case cotransmitter meaning that a “classical” or “fast acting” neurotransmitter is also present). For example, CBI-2 is a cholinergic neuron (Hurwitz et al., 2003). Again, the feeding system is not unique in this respect, e.g., some of the modulatory input to the STG is provided by input from projection neurons that contain multiple neurotransmitters, e.g., the GABAergic neurons MCN1 and MPN both also contain the peptide proctolin, and MCN1 additionally contains Cancer borealis tachykinin-related peptide (CabTRP; Nusbaum and Marder, 1989; Blitz et al., 1999). These STG neurons elicit different motor programs (as do the ingestive and egestive inputs to the feeding CPG). MCN1 elicits gastric rhythms whereas MPN does not (Blitz and Nusbaum, 1997, 1999; Wood et al., 2000).
The fact that CBI-2 and EN afferents make “fast” synaptic connections is of interest since these connections are not identical. For example, CBI-2 produces one-for-one fast EPSPs in an interneuron (B40), which plays a key role in patterning ingestive activity (Jing and Weiss, 2002). In contrast B40 is not activated by EN stimulation (Proekt et al., 2007). CBI-2 produces fast IPSPs in B65, an interneuron (Jing and Weiss, 2005) that receives excitatory input during EN induced motor programs (Proekt et al., 2007). In principal, therefore, motor program selection could simply result from differences in fast synaptic transmission. It is likely, however, that the dynamics of program selection in this type of situation would differ from the dynamics that are actually observed. Thus, if motor program selection were entirely a consequence of fast synaptic activity it presumably would occur relatively quickly. This is not what has been reported in previous studies of feeding motor program induction.
Feeding motor programs that are triggered after a period of inactivity are poorly defined (although unprimed CBI-2 induced activity does differ from EN induced activity, presumably as a result of differences in fast synaptic connections; Proekt et al., 2004; Friedman et al., 2009). With repeated input activation there are progressive alterations in evoked activity, but these changes occur relatively slowly (Proekt et al., 2004, 2007; Friedman et al., 2009; Dacks et al., 2012). Programs assume their final character after induction of multiple cycles (e.g., ∼9–10 cycles in the case of ingestive priming). Presumably, the relatively slow dynamics that are observed are a consequence of the role of modulatory neurotransmitters. Effects of modulators are second messenger mediated, and their effects persist beyond periods of actual neural activity (Friedman and Weiss, 2010). Consequently, when cycles of motor programs are repetitively induced, modulatory effects carry over from one cycle to the next and gradually summate. This type of gradual selection process is likely to be beneficial in the situation where stimuli of a given nature tend either to be clustered or persist. It will be advantageous because fully articulated responses will only be generated if a stimulus is repeatedly encountered. Stimuli that trigger feeding behaviors in Aplysia are likely to fall into this category (e.g., seaweed tends to grow in clumps).
The fact that there are differences in intercellular and intracellular signaling processes used to induce ingestive versus egestive activity may be important because it influences the specific dynamics of the “selection” process. Further, it may influence the associated duration of the persistence of priming. Thus, although it is not necessarily the case, effects that are exerted via different signaling pathways tend to have different temporal characteristics. Consistent with this idea, experiments that have studied the induction and persistence of ingestive and egestive priming in vitro have noted differences in observed dynamics. Precise values noted obviously depend on the stimulation parameters used to induce priming. In all reports to date, however, buildup during ingestive priming develops more slowly and its effects outlast those of egestive priming (Proekt et al., 2004, 2007; Friedman et al., 2009; Dacks et al., 2012). In future work it will be of interest to determine whether these different dynamics are also observed in vivo.
In summary, in this report we present data that suggest that a form of chemical coding is used to induce ingestive versus egestive patterns of activity in the motor neurons that open the radula. Namely, FCAP/CP-2/cAMP induces ingestive activity, and SCPb/PKC induces egestive activity. FCAP/CP-2 and SCPb are both present as cotransmitters in inputs to the feeding CPG. We suggest that the use of two different signaling systems results in a situation where priming is induced with behavior specific dynamics.
Footnotes
This work was supported by National Institutes of Health Grants NS066587, NS070583, and MH051393.
The authors declare no competing financial interests.
- Correspondence should be addressed to Dr Elizabeth Cropper, Department of Neuroscience, Box 1065, Mt Sinai School of Medicine, 1 Gustave Levy Place, New York, NY 10029. elizabeth.cropper{at}gmail.com





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}