

Abstract
Food availability and nutritional status are important cues affecting behavioral states. Here we report that, in Caenorhabditis elegans, a cascade of dopamine and neuropeptide signaling acts to inhibit nociception in food-poor environments. In the absence of food, animals show decreased sensitivity and increased adaptation to soluble repellents sensed by the polymodal ASH nociceptors. The effects of food on adaptation are affected by dopamine and neuropeptide signaling; dopamine acts via the DOP-1 receptor to decrease adaptation on food, whereas the neuropeptide receptors NPR-1 and NPR-2 act to increase adaptation off food. NPR-1 and NPR-2 function cell autonomously in the ASH neurons to increase adaptation off food, whereas the DOP-1 receptor controls neuropeptide release from interneurons that modulate ASH activity indirectly. These results indicate that feeding state modulates nociception through the interaction of monoamine and neuropeptide signaling pathways.
Introduction
Feeding state and nutritional status guide many behaviors and physiological responses in all animals. Both internal cues, such as satiety or starvation, and external cues indicating the presence or absence of a food source strongly influence brain states and the properties of neuronal circuits. In the nematode Caenorhabditis elegans, food availability and nutritional status have strong effects on several behaviors. Within minutes of leaving food, animals change their speed and locomotion and inhibit pharyngeal pumping and egg laying (Trent et al., 1983; Avery and Thomas, 1997; Sawin et al., 2000; Hills et al., 2004). More prolonged starvation has additional effects on diverse behaviors, such as chemotaxis and foraging, and also affects gene expression (Rogers et al., 2006; Suo et al., 2006; Tomioka et al., 2006; Hukema et al., 2008; Zimmer et al., 2009).
In C. elegans, monoamines have been shown to be particularly important molecules in the control of food-related behavior. For example, dopamine appears to be directly released by food-sensing neurons and affects food-dependent changes in locomotion, gene expression, and sensory perception (Sawin et al., 2000; Chase et al., 2004; Hills et al., 2004; Suo et al., 2006; Kindt et al., 2007; Ezcurra et al., 2011). Serotonin is required for food-dependent modulation of pharyngeal pumping, locomotion, and olfaction (Horvitz et al., 1982; Zhang et al., 2005; Harris et al., 2009), whereas octopamine mediates the control of olfaction and gene expression by feeding state (Suo et al., 2006; Wragg et al., 2007; Mills et al., 2012). Finally, tyramine has been shown to affect the control of foraging, egg laying, and pharyngeal pumping by feeding state (Rex et al., 2004; Alkema et al., 2005; Rex et al., 2005).
In addition to classical neurotransmitters, C. elegans has a complex system of neuropeptides consisting of ∼100 peptide genes, encoding >250 peptides. Many of these are members of conserved families of bioactive peptides, including 40 insulin-like peptide genes and 34 of the FMRFamide-like peptide (FLP) family (Li and Kim, 2008). A few of these have been implicated in food-related behaviors (Harris et al., 2010), in particular INS-1 (Tomioka et al., 2006), FLP-18 (Cohen et al., 2009), and FLP-1 (Waggoner et al., 2000). Determining the function of many of these peptides has turned out to be a difficult task, because the precursor genes have overlapping functions and expression patterns, and neuropeptide receptors have the ability to bind to several peptides.
Here we report that neuropeptide signaling inhibits escape responses to noxious stimuli in C. elegans in the absence of food but not in the presence of food. Two receptors from the neuropeptide F/Y family, NPR-1 and NPR-2, act within the ASH nociceptor to inhibit avoidance by increasing sensory adaptation to repellents when food availability is low. Dopamine has the opposite effect of enhancing ASH responses and decreasing adaptation in the presence of food (Ezcurra et al., 2011). Our evidence suggests that the effects of dopamine are NPR-1 dependent and mediated indirectly through changes in neuropeptide signaling from the AUA interneurons. Thus, monoamine and neuropeptide modulators appear to interact to modify sensory perception in response to changes in environmental and behavioral states.
Materials and Methods
Strains and culture
Strains were maintained as described previously (Brenner, 1974). Hermaphrodites were used for all experiments. All strains were grown at 20°C on nematode growth medium (NGM) plates with OP50. The following strains were used: N2 (wild-type reference), CB1112 cat-2(e1112), CX4148 npr-1(ky13), DA609 npr-1(ad609), DA650 npr-1(g320), AQ2180 cat-2(e1112);ljEx95[sra-6::YC2.12;lin-15(+)], NY34 flp-8(pk360), RB982 flp-21(ok889), AX410 flp-18(db99), LX636 dop-1(vs101), and AQ1044 ljEx95 [sra-6::YC2.12;lin-15(+)]. The following strains were created: ljEx305[sra-6::npr-1;unc-122::gfp], AQ2803 npr-1(ky13);ljEx95 [sra-6::YC2.12;lin-15(+)], AQ2933 npr-1(ky13);ljEx95 [sra-6::YC2.12;lin-15(+)];ljEx305[sra-6::npr-1;unc-122::gfp], AQ2180 cat-2(e1112);ljEx95[sra-6::YC2.12;lin-15(+)], AQ2310 cat-2(e1112);npr-1(ky13);ljEx95[sra-6::YC2.12;lin-15(+)], AQ2928 ljEx466[dop-1::dop-1 RNAi;myo-2::gfp], AQ2929 ljEx467[flp-8::dop-1 RNAi;myo-2::gfp], AQ2931 ljEx469[flp-8::dop-1 cDNA;myo-2::gfp];dop-1 (vs101), AQ2899 dop-1(vs101);ljEx454[dop-1::dop-1;unc-122::gfp], AQ2897 dop-1(vs101);ljEx453[mec-7::dop-1;unc-122::gfp], AQ2942 dop-1(vs101);Ex [sra-6::dop-1;unc-122::gfp], AQ1364 npr-1(ky13);npr-2(ok419), AQ2193 npr-2(ok419);ljEx95[sra-6::YC2.12;lin-15(+)], AQ2056 npr-2(ok419), AQ2302 npr-1(ky13);npr-2(ok419);ljEx95[sra-6::YC2.12;lin-15(+)], AQ2387 npr-2(ok419);ljEx293[Psra-6::npr-2;Pelt-2::mCherry], AQ2171 ljEx243[Psra-6::npr-2::gfp, Punc-122::gfp] line 1, AQ2169, ljEx243[Psra-6::npr-2::gfp, Punc-122::gfp] line 2, GA1720 npr-1(ky13);wuEx285[gpa-13::npr-1;Pmyo-2::gfp], GA1721 wuEx286[Pflr-4::unc-31senseRNAi;Pflp-8::unc-31antisenseRNAi;Pmyo-2::gfp], GA1722 wuEx287[flp-8::unc-31;Pmyo-2::gfp], GA1723 npr-2(ok419);wuEx288[gpa-13::npr-2;Pmyo-2::gfp], and GA1724 wuEx289[C41G7.6::dop-1;Pmyo-2::gfp]; dop-1(vs101).
Generation of transgenic animals
Cameleon strains were generated by crossing ljEx95[sra-6::YC2.12] (Hilliard et al., 2005) into npr-1(ky13) and cat-2(e1112). npr-1(ky13) was confirmed by PCR, followed by digest of the PCR product using DraI. cat-2(e1112) was confirmed by PCR, followed by digest using StyI.
Rescue plasmids.
Plasmids were made using the MultiSite Gateway Three-Fragment Vector Construction kit (Invitrogen). Promoters were inserted into pDONR P4-P1R. Genomic DNA of npr-1 and npr-2 cDNA were inserted into pDONR 221. A pENTRY P2R-P3 containing the unc-54 3′UTR (kindly provided by Ithai Rabinowitch, MRC-LMB, Cambridge, UK) was used. npr-1 and npr-2 rescue constructs were generated by recombining the pENTRY and pDESTR4-R3 vectors using LR Clonase. sra-6 pEntry was kindly provided by Emanuel Busch (MRC-LMB, Cambridge, UK) and contains a 3 kb region of the sra-6 promoter. dop-1 rescue constructs were made by PCR fusion of the promoter, the genomic DNA, and the unc-54 3′UTR. The flp-8::dop-1 rescue was made with cDNA instead of genomic DNA, and the cDNA was synthesized using SuperScript III One-Step RT-PCR System with Platinum Taq High Fidelity (Invitrogen). All rescue constructs were injected (Mello et al., 1991) at 50 ng/μl with coinjection marker unc-122::gfp at 50 ng/μl.
Rescue constructs by PCR fusion.
Rescue constructs using the gpa-13 and C41G7.6 promoters were made using PCR fusion (Hobert, 2002) and injected at 50 ng/μl with myo-2::gfp at 50 ng/μl as injection marker.
RNAi constructs and strains.
dop-1 RNAi lines were generated as described previously (Esposito et al., 2007), and each RNAi strand was injected at 100 ng/μl with either unc-122::gfp at 50 ng/μl or myo-2::gfp at 5 ng/μl as coinjection marker. flp-8::unc-31 expressing strains were constructed using the same method, and each strand was injected at 50 ng/μl with myo-2::gfp at 50 ng/μl as injection marker. The AUA-specific unc-31 RNAi strain was made as described previously (Hamakawa et al., 2015). The unc-31 RNAi sense strand was expressed using the flr-4 promoter, and the antisense strand was expressed under the flp-8 promoter. Each strand was injected at 50 ng/μl with myo-2::gfp at 20 ng/μl as injection marker.
Drop test assays
For all behavioral assays, L4 stage animals were picked 20 h before the assay. The drop test and the adaptation drop test were performed on NGM plates and prepared for off-food and on-food assays as follows. For on-food plates, 25 μl of overnight culture of OP50 in Luria broth [LB; absorbance (A) = 0.5] was spread on each plate. For off-food plates, 25 μl of LB was spread on each plate. All plates were allowed to dry for 1 h without lids and used after 1 additional hour for assays. Behavioral assays can be sensitive to environmental factors, such as humidity. We have noticed that the humidity levels of the plates have a strong effect on the adaptation phenotype, and animals adapt faster on wetter plates than on dry plates. Therefore, baseline levels of adaptation differ between experiments. Animals were picked off the culture plate using an eyelash pick and placed on a plate without food for a few seconds to prevent food from being transferred to the assay plates. The animals were then placed on the assay plate. Animals were allowed to settle for 10 min and then assayed using a capillary to deliver the repellent drop as described previously (Hilliard et al., 2002). For the drop test, 10 animals were assayed on each plate. All animals were stimulated every 60 s, and the fraction of worms reversing was recorded. In the adaptation drop test, a single animal was picked to each plate and assayed every 10 s, 20 times. The response to every drop was recorded, and the avoidance index was calculated by dividing the number of animals reversing with the number of animals tested. The tested repellents were CuCl2 (copper(II)chloride dehydrate; Sigma) and glycerol (Thermo Fisher Scientific). Repellents were dissolved and diluted in M13 buffer. Each experiment was performed blinded and on more than 2 different days.
For transgenic strains, we generated two independent transgenic lines from each transgene and analyzed at least 15 animals from each line, averaging the results from both lines.
Calcium imaging experiments
Sample preparation and delivery of stimulation.
Animals were glued on 2% agarose pads made with neuronal buffer using cyanoacrylate glue (Nexaband S/C; Abbott Laboratories). The pad was placed briefly on ice during the gluing to constrict the animal's movements. The animals were placed under the microscope in a perfusion chamber (RC-26GLP; Warner Instruments) under constant flow rate (0.4 ml/min) of neuronal buffer using a perfusion pencil (AutoMate). Outflow was regulated using a peristaltic pump (Econo Pump; Bio-Rad). Repellents were delivered using the perfusion pencil and automated valves (EW-98302-20; Cole Parmer), controlled by Motorway software 2.5. The neuronal buffer contained 40 mm NaCl, 10 mm HEPES-NaOH, pH 7.1, and 1 mm MgSO4, and the osmolarity was adjusted to 350 mOsm using glycerol. Copper and glycerol were dissolved in M13 buffer. For recordings with food, OP50 was grown overnight in LB (A = 0.5), and 50 ml cultures were pelleted by centrifugation. The pellets were dissolved in neuronal buffer or repellent, to a final volume of 25 ml, and used within 1 h. Adaptation was tested by first giving the animals a 3 s prechronic stimulation, followed by a 15 s chronic stimulation and a 2 min rest. Animals were then given a 3 s postchronic stimulation. The prechronic and postchronic responses were analyzed.
Calcium imaging and image analysis.
Optical recordings were performed on a Zeiss Axioskop 2 upright compound microscope using a 63× Zeiss Achroplan water-immersion objective. Filter/dichroic pairs were as follows: excitation, 420/40 nm; excitation dichroic, 455 nm; CFP emission, 480/30 nm; emission dichroic, 505 nm; and YFP emission, 535/30 nm (Chroma Technology). The microscope was fitted with a Hamamatsu Orca ER CCD camera, a Hamamatsu W-View emission image splitter, and a Uniblitz Shutter (Vincent Associates). Images were acquired at 10 Hz using MetaVue 4.6 (Universal Imaging). Image analysis was performed as described previously (Kerr et al., 2000) using a custom program written in Java, parameterized, and averaged using scripts written in MATLAB 6.5.1 (MathWorks). The ASH cell body was identified based on its position and tracked as described using custom software (Hilliard et al., 2005).
In vitro receptor activation assays
Activation of NPR-2 by candidate peptide ligands was monitored using an in vitro calcium mobilization assay as described previously (Janssen et al., 2008). Chinese hamster ovary (CHO) K1 cells stably overexpressing the mitochondrial-targeted apo-aequorin and the human Gα16 subunit were cultured and transfected with pcDNA3.1D::npr-2-cDNA. Cells for negative control experiments were transfected with an empty pcDNA3.1D vector. A peptide library of 262 synthetic C. elegans peptides was composed based on in silico predictions and in-house peptidomics data and custom synthesized by Thermo Fisher Scientific, GL Biochem, and Eurogentec. Receptor activating peptides were HPLC purified and tested in final concentrations from 10−4 to 10−10 m. Calcium responses were monitored as described previously (Janssen et al., 2008) for 30 s on a Mithras LB 940 luminometer (Berthold Technologies). The half-maximal effective concentration (EC50) values of peptide ligands were determined from concentration–response curves, which were constructed using a computerized nonlinear regression analysis with a sigmoidal concentration–response equation (Prism 6; GraphPad Software).
Results
The neuropeptide receptor NPR-1 promotes adaptation to chemical repellents off food
We demonstrated previously that escape responses to soluble repellents adapt more slowly in animals tested in the presence of a bacterial food source than in animals tested in the absence of food. This effect requires dopamine, which is released by neurons that directly sense bacteria in the environment (Ezcurra et al., 2011). To investigate whether other molecules also mediate the effect of feeding state on repellent adaptation, we screened through candidate monoamine and neuropeptide receptor genes for abnormal adaptation to copper. We found that animals carrying a null mutation in the neuropeptide receptor npr-1, which is implicated in aggregation and other food-modulated behaviors (de Bono and Bargmann, 1998), adapted normally to repeated stimulation with 10 mm CuCl2 in the presence of food but showed reduced adaptation to copper in the absence of food (Fig. 1A,B). We observed the same phenotype for other chemical repellents, including 1 m glycerol and HCl at pH 3.5 (Fig. 1C–F). Thus, npr-1 loss of function caused food-deprived animals to show behavior characteristic of feeding animals. This adaptation phenotype was rescued by expression of an npr-1(+) transgene in ASH under the sra-6 promoter (Fig. 1A), which drives expression in ASH and other neurons. To ensure that NPR-1 functions in ASH, we also constructed a transgene under the gpa-13 promoter, which only overlaps with sra-6 in ASH. This transgene also rescued the adaptation phenotype (see Fig. 3D), suggesting that NPR-1 functions in ASH to modulate repellent sensation in the absence of food.

The neuropeptide receptor NPR-1 promotes sensory adaptation off food. A–J, Fraction of animals responding to repeated drops of chemical repellent. A, npr-1(ky13) adapts slower than wild type to 10 mm CuCl2 off food. n = 40. Transgenic animals expressing genomic npr-1 DNA under the sra-6 promoter (ASH, ASI, and PVQ) adapt like wild type. Two stable independent lines were analyzed, and the average of both lines is shown. n = 40. B, npr-1(ky13) animals adapt like wild type to 10 mm CuCl2 on food. n = 24. npr-1(ky13), wild-type and rescued animals adapt at the same rate on food. n = 21. C, npr-1(ky13) adapts slower than wild type to HCl, pH 3.5, off food. n = 38. D, npr-1(ky13) animals adapt like wild type to HCl, pH 3.5, on food. n = 19. E, npr-1(ky13) adapts slower than wild type to 1 m glycerol off food. n = 37. F, npr-1(ky13) animals adapt like wild type to 1 m glycerol on food. n = 22. G, npr-1(ad609) adapts slower than wild type off food. n = 34. H, npr-1(ad609) animals adapt like wild type on food. n = 20. I, npr-1(g320) adapts like wild type off food. n = 29. J, npr-1(g320) animals adapt like wild type on food. n = 19. Behavioral data were analyzed using Student's t test. *p < 0.05, **p < 0.01, ***p < 0.001.
The npr-1 allele found in the standard C. elegans strain N2 differs from the allele found in naturally occurring C. elegans populations (McGrath et al., 2009). The N2 allele, which is thought to have arisen during cultivation in the laboratory, encodes a receptor with a valine residue at position 215; this npr-1(215V)-encoded variant has heightened activity relative to the npr-1(215F) allele found in wild C. elegans populations carrying a phenylalanine residue at position 215. Because the low-activity npr-1(215F) strains share behavioral and other phenotypes with npr-1 null mutants (de Bono and Bargmann, 1998; Styer et al., 2008), we wondered whether the effect of food on repellent adaptation might be specific to strains carrying the laboratory-selected npr-1(215V) allele. To examine this possibility, we compared the escape responses to copper on and off food of npr-1(g320), which harbors the npr-1(215F) allele in an otherwise N2 background, with N2. We observed no significant differences in copper adaptation between this strain and N2 either on or off food (Fig. 1G,H), indicating that food modulation occurs in both npr-1(251V) and npr-1(215F) strains. In contrast, a second npr-1 null allele (ad609) showed the same decreased adaptation off food that was observed in the npr-1(ky13) null mutant (Fig. 1G,H). Thus, both the 215V and 215F variants of NPR-1 appear to mediate enhanced adaptation to soluble repellents in the absence of food.
NPR-1 affects sensory adaptation in ASH chemosensory neurons
The ASH neuron has been shown to play a critical role in the sensation of multiple soluble repellents, including copper, glycerol, and acidic pH. To determine whether NPR-1 affects the repellent responses of the ASH neurons themselves, we performed calcium imaging experiments using the genetically encoded calcium indicator cameleon. In these calcium imaging experiments, we observed that npr-1(ky13) animals showed normal initial CuCl2-evoked calcium responses in ASH in both the presence and absence of food (Fig. 2B,D,E). We then tested adaptation after prolonged exposure to copper. We found that, in wild-type animals, the ASH neurons adapted more strongly in the absence of food than in the presence of food (Fig. 2A,D,E). When we tested npr-1(ky13) animals, we found that they adapted less than wild type in the absence of food (Fig. 2A,B,D,E) but adapted to the same degree as wild type in the presence of food. The calcium imaging phenotype, like the behavioral defect, was rescued by expression of an npr-1(+) transgene in ASH under the sra-6 (Fig. 2C–E) or the gpa-13 promoter. Together, these results indicate that NPR-1 functions in ASH to modulate repellent responses in the absence but not the presence of food. This suggests that the NPR-1 receptor responds to a ligand (or ligands) released under conditions in which food is absent.

The neuropeptide receptor NPR-1 acts in ASH to increase adaptation to repellents off food. A–C, Average neuronal responses in ASH before and after chronic treatment with 10 mm CuCl2. Shown are responses on and off food. Gray bars indicate the duration of the stimulus. A, Off food, wild type completely adapts after the chronic treatment and shows no postchronic responses. On food, wild type still responds after the chronic treatment. B, npr-1(ky13) shows postchronic responses both off and on food. C, Transgenic animals expressing genomic npr-1 DNA under the sra-6 promoter display wild-type responses. Data from one stable transgenic strain are shown. D, E, Quantification of ASH responses on and off food in wild-type, npr-1(ky13), and rescued animals before and after chronic treatment. Blue circles represent individual animals assayed. Black diamonds indicate average value. D, Postchronic responses in npr-1(ky13) are significantly larger than in wild-type and rescued animals off food. E, Postchronic responses are not significantly different on food. Data were analyzed using the Mann–Whitney rank-sum test. *p < 0.05, **p < 0.01, ***p < 0.001.
NPR-1 and NPR-2 act in the same pathway to affect adaptation in ASH
In addition to NPR-1, we found a closely related neuropeptide receptor that also shows defective adaptation to noxious stimuli. Animals carrying a null mutation in npr-2, a deletion that takes out most of the coding sequence, have a phenotype similar to npr-1 mutants; they respond normally to initial stimulation with 10 mm CuCl2 but adapt slower to repeated stimulation in the absence of food (Fig. 3A). Furthermore, calcium imaging experiments showed that, in npr-2(ok419) animals, the ASH neuron shows normal initial responses and suppressed adaptation to repeated stimulation with CuCl2 in the absence of food but not in the presence of food (Fig. 3E–G). NPR-2 is widely expressed in the nervous system, including in the ASH neuron (Luo et al., 2015). We expressed an npr-2(+) transgene under either the sra-6 or gpa-13 promoter and found that the behavioral and calcium imaging adaptation phenotypes of npr-2(ok419) were rescued (Fig. 3A,C,E,F). This suggests that both NPR-1 and NPR-2 act in the ASH neuron to increase adaptation in the absence of food.

The neuropeptide receptor NPR-2 acts in ASH to increase adaptation in the absence of food. A–D, Fraction of animals responding to repeated drops of chemical repellent. For transgenic strains, the average values of two stable independent strains are shown. A, npr-2(ok419) adapts slower than wild type to 10 mm CuCl2 off food. Transgenic animals expressing npr-2 cDNA under the sra-6 promoter (ASH, ASI, and PVQ) adapt like wild type. B, npr-1(ky13);npr-2(ok419) adapts like the npr-1(ky13) and npr-2(ok419) single mutants. C, Transgenic animals expressing genomic npr-1 DNA under the gpa-13 promoter display wild-type responses. D, Transgenic animals expressing npr-2 cDNA under the gpa-13 promoter display wild-type responses. For transgenic strains, the average values of two independent strains are shown. E, Average neuronal responses in ASH after chronic treatment with 10 mm CuCl2. Shown are responses on and off food. Gray bars indicate the duration of the stimulus. Off food, wild type completely adapts after the chronic treatment and shows no postchronic responses. On food, wild type still responds after the chronic treatment. npr-2(ok419) shows postchronic responses both off and on food. Transgenic animals expressing npr-2 cDNA under the sra-6 promoter display wild-type responses. Off food, npr-1(ky13);npr-2(ok419) has similar postchronic responses to the npr-1(ky13) and npr-2(ok419) single mutants. F, G, Quantification of ASH responses off food before and after chronic treatment. Black triangles represent individual animals assayed. Straight lines indicate average value. F, Prechronic responses off food are similar in all genotypes. G, Postchronic responses are significantly larger in npr-2(ok419) than in wild type. Transgenic animals expressing npr-2 cDNA under the sra-6 promoter display wild-type responses off food. Data from one stable transgenic strain are shown. Behavioral data were analyzed using Student's t test. Calcium imaging data were analyzed using the Mann–Whitney rank-sum test. *p < 0.05, **p < 0.01, ***p < 0.001.
NPR-1 and NPR-2 could either be acting in parallel, separate pathways to modulate cellular responses in ASH or in the same signaling pathway. We performed behavioral tests and calcium imaging experiments, which showed that double mutants carrying both the npr-1(ky13) and npr-2(ok419) mutations do not have a stronger defect than the single npr-1(ky13) and npr-2(ok419) mutants (Fig. 3B,E–G). These results suggest that NPR-1 and NPR-2 act in the same pathway to increase ASH adaptation.
The NPR-1 ligands FLP-18 and FLP-21 do not increase ASH adaptation
To identify relevant neuropeptide receptor ligands, we surveyed known C. elegans neuropeptides using in vivo and in vitro approaches. Previous in vitro studies have identified the FMRFamide peptides FLP-18 and FLP-21 as ligands of NPR-1 (Rogers et al., 2003), suggesting that these peptides could be functioning to increase adaptation through NPR-1. We performed similar in vitro screens for NPR-2 ligands, testing >260 known C. elegans neuropeptides for activation of the two NPR-2 isoforms, NPR-2a and NPR-2b. Only one peptide encoded by flp-21 (GLGPRPLRFamide) showed evidence for activating NPR-2 at physiological concentrations (Fig. 4A). Thus, the FLP-21 neuropeptide appears to be a ligand of NPR-2, as well as of NPR-1.

FLP-21 peptides are ligands for NPR-2. A, Concentration-dependent calcium responses evoked by FLP-21 (GLGPRPLRFamide) in CHO cells expressing NPR-2a or NPR-2b. The log(EC50) values ± SEM of FLP-21 is −7.464 ± 0.147 for NPR-2a and −7.459 ± 0.110 for NPR-2b. Each point ± SEM represents the average of two independent experiments performed in triplicate. Concentration–response data are shown as relative (percentage) to the highest value (100% activation) after normalization to the total calcium response (n ≥ 6). B, C, Fraction of animals responding to repeated drops of chemical repellent. flp-21(ok889) adapts like wild type and flp-18(db99) adapts faster than wild type (B) to 10 mm CuCl2 off food. For transgenic strains, the average values of two independent strains are shown. Behavioral data were analyzed using Student's t test. *p < 0.05, **p < 0.01, ***p < 0.001.
Therefore, we tested null mutants for flp-21 and flp-18 for ASH adaptation phenotypes. We found that flp-21(ok889) adapts like wild type and that flp-18(db99) adapts more rapidly (Fig. 4B), indicating that the peptides encoded by these genes are not required for adaptation in ASH. To test whether these previously identified NPR-1 ligands might act redundantly with other peptides, we also made lines overexpressing flp-18 and flp-21 in wild-type background. None of these lines displayed altered responses to acute or repeated repellent stimulation (Fig. 4C). Thus, FLP-21 and FLP-18 peptides do not appear to be necessary for activating NPR-1 or NPR-2 in ASH. We also tested deletion mutants of other FMRFamide-related peptide genes (flp-1–flp-3, flp-6–flp-13, flp-15–flp-17, flp-19, flp-20, flp-25, flp-28, and flp-33) and found that none of these strains exhibit an adaptation defect similar to npr-1 and npr-2 (data not shown). Together, these results suggest that the relevant ligand (or ligands) that activates NPR-1 and NPR-2 to promote ASH adaptation is likely to be an unknown peptide or that a more complex network of peptidergic interactions is at play.
Neuropeptide signaling mediates the effect of dopamine on adaptation
NPR-1 and dopamine signaling have opposing effects on adaptation to repellents: NPR-1 increases adaptation in unfed animals, whereas dopamine decreases adaptation in well fed animals. We showed previously that cat-2(e1112) animals, which carry a nonsense allele in the gene encoding tyramine hydroxylase and are defective in dopamine biosynthesis (Lints and Emmons, 1999), adapted rapidly in the presence of food, suggesting that endogenous dopamine is required for the inhibitory effect of food on adaptation (Ezcurra et al., 2011).
To determine how these opposite pathways interact, we analyzed copper adaptation in cat-2; npr-1 double mutants. In the absence of food, in both behavioral and calcium imaging experiments, we found that cat-2; npr-1 animals showed decreased adaptation compared with wild type (Fig. 5A,C,E), a phenotype comparable with that of the npr-1 single mutant. Likewise, in the presence of food, cat-2; npr-1 animals showed slow adaptation (Fig. 5B,D,F), resembling wild type and the npr-1 single mutant, but not the cat-2 single mutant. Thus, npr-1 appeared completely epistatic to cat-2 with respect to ASH adaptation phenotypes. These results indicate that dopamine acts in the same pathway as NPR-1 to inhibit ASH adaptation and imply that dopamine somehow inhibits NPR-1 activity. Because NPR-1 is a neuropeptide receptor, this suggests that the effects of dopamine on adaptation could be mediated indirectly through changes in neuropeptide signaling.

NPR-1 is required for dopamine modulation of ASH adaptation. A, B, Fraction of animals responding to repeated drops of 10 mm CuCl2. A, cat-2(e1112);npr-1(ky13) animals adapt like npr-1(ky13) in the absence of food. B, cat-2(e1112);npr-1(ky13) animals adapt like npr-1(ky13) in the presence of food. C, D, Average neuronal responses in ASH off food after chronic treatment with 10 mm CuCl2. Gray bars indicate the duration of the stimulus. C, Wild-type and cat-2(e1112) animals completely adapt after the chronic treatment on food. npr-1(ky13) and cat-2(e1112);npr-1(ky13) animals adapt less and show postchronic responses. D, Wild-type, npr-1(ky13), and cat-2(e1112);npr-1(ky13) animals show postchronic responses on food. cat-2(e1112) animals completely adapt on food. E, F, Quantification of ASH responses on and off food. Blue circles represent individual animals assayed. Black diamonds indicate average value. E, Wild-type and cat-2(e1112) completely adapt after the chronic treatment off food. npr-1(ky13) and cat-2(e1112);npr-1(ky13) respond. F, Wild-type, npr-1(ky13), and cat-2(e1112);npr-1(ky13) respond after the chronic treatment on food. cat-2(e1112) has completely adapted. Behavioral data were analyzed using Student's t test. Calcium imaging data were analyzed using the Mann–Whitney rank-sum test. *p < 0.05, **p < 0.01, ***p < 0.001.
Dopamine inhibits adaptation through the dopamine receptor DOP-1
To learn more about the role of dopamine in ASH adaptation, we analyzed the repellent adaptation phenotypes of dopamine receptor mutants. C. elegans has four G-protein-coupled dopamine receptors, dop-1 to dop-4, and we tested deletion mutants of each for food-dependent abnormalities in copper adaptation. Previously, we showed that DOP-4 is required for the acute response but not for adaptation in ASH in the presence of food (Ezcurra et al., 2011). We found that the dop-1(vs101) deletion allele has an adaptation phenotype identical to cat-2: mutant animals adapt faster than wild type on food, but not off food (Fig. 6A,B), This result suggested that DOP-1 acts in the presence of food to inhibit adaptation.

The dopamine receptor DOP-1 inhibits adaptation on food. A–D, Fraction of animals responding to repeated drops of 10 mm CuCl2. A, dop-1(vs101) adapts like wild-type off food. B, dop-1(vs101) adapts faster than wild type on food. C, Rescue of dop-1 using the dop-1 promoter rescues the dop-1 phenotype, but expression under the mec-7 promoter does not. D, Rescue of dop-1 using the sra-6 promoter does not rescue the phenotype. Behavioral data were analyzed using Student's t test. *p < 0.05, **p < 0.01, ***p < 0.001. For transgenic strains, the average values of two independent strains are shown.
To determine whether DOP-1 acts cell autonomously in ASH to modulate adaptation to repellents, we tested whether the dop-1 mutant phenotype could be rescued by expressing a dop-1(+) transgene in ASH. We observed that an ASH-expressing sra-6::dop-1 transgene failed to rescue the dop-1 adaptation phenotype (Fig. 6D); in contrast, expression of dop-1 under its own promoter rescued the adaptation phenotype (Fig. 5C). Thus, we reasoned that DOP-1 might function in neurons extrinsic to ASH, perhaps to control release of NPR-1 ligands.
DOP-1 has been reported to be expressed in a subset of neurons, including the ALM, AVM, and PLM gentle touch neurons, cholinergic motor neurons in the ventral cord, and the RIS, RIM, AUA, RIB, PHC, and PVQ interneurons (Tsalik et al., 2003; Chase et al., 2004; Sanyal et al., 2004). Expressing dop-1(+) under a touch neuron-specific promoter, mec-7 (shown previously to rescue mechanosensory adaptation phenotypes; Kindt et al., 2007) failed to rescue the copper adaptation phenotype of dop-1 (Fig. 6C), but expressing dop-1(+) under the flp-8 promoter, which expresses in the touch neurons as well as several additional neuron classes, fully rescued the dop-1 adaptation phenotype (Fig. 7A). We also generated cell-specific RNAi lines expressing dop-1 sense and antisense sequences under the flp-8 promoter (referred to as dop-1 RNAi), along with control RNAi lines expressing the same dop-1 sequences under the dop-1 promoter. Behavioral phenotyping revealed that expression of dop-1 RNAi under the dop-1 and flp-8 promoters leads to increased adaptation on food compared with wild type (Fig. 7B,C). One candidate site for DOP-1 function are the AUA neurons, because they are in the expression domains of dop-1 and flp-8 but not mec-7. To determine whether DOP-1 expression in AUA is required for modulation of adaptation, we constructed a dop-1 rescue strain using the C41G7.6 promoter, which only overlaps with flp-8 and dop-1 in AUA. Driving expression of dop-1 under C41G7.6 rescued the adaptation phenotype (Fig. 7D), suggesting that DOP-1 acts in the AUA neurons to decrease ASH adaptation in the presence of food, potentially by negatively regulating the release of neuropeptides that act through the NPR-1 pathway.

DOP-1 acts in FLP-8-expressing neurons to inhibit adaptation on food. A–D, Fraction of animals responding to repeated drops of 10 mm CuCl2. A, Rescue of dop-1 using the flp-8 promoter rescues the adaptation phenotype on food. B, Knockdown of dop-1 in dop-1-expressing neurons increases adaptation on food. C, Knockdown of dop-1 in flp-8-expressing neurons increases adaptation on food. D, Rescue of dop-1 using the C41G7.6 promoter rescues the adaptation phenotype on food. For transgenic strains, the average values of two independent strains are shown. Behavioral data were analyzed using Student's t test. *p < 0.05, **p < 0.01, ***p < 0.001. For transgenic strains, the average values of two independent strains are shown.
Neuropeptide release from AUA increases adaptation
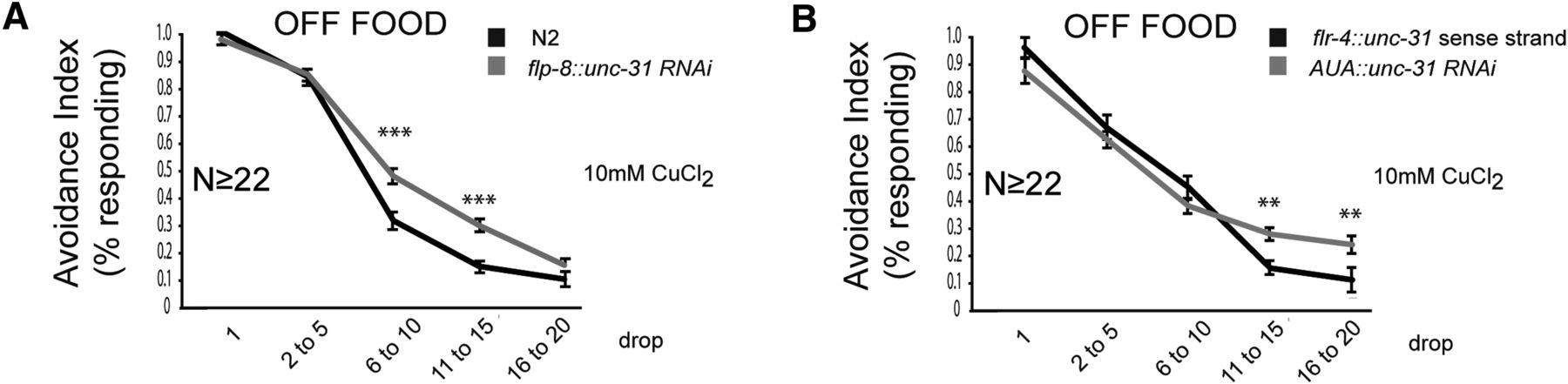
We then tested whether neuropeptide release from flp-8-expressing neurons suppresses adaptation. unc-31 encodes the dense-core vesicle priming proteins CAPS, which is essential for the release of dense core vesicles (Hammarlund et al., 2008; Speese et al., 2007). Expression of unc-31 RNAi under the flp-8 promoter suppressed adaptation (Fig. 8A), suggesting that peptidergic signaling from flp-8-expressing neurons function to increase adaptation. To determine whether neuropeptide signaling from AUA specifically is required for modulation of adaptation, we constructed a strain expressing unc-31 RNAi in AUA only, by expressing the sense strand under the flp-8 promoter and the antisense strand under the flr-4 promoter. These two promoters overlap in AUA but not in other neurons, resulting in expression of double-stranded unc-31 RNAi specifically in AUA. This transgene resulted in decreased adaptation compared with the unc-31 sense strand alone expressed under the flr-4 promoter (Fig. 8B), suggesting that peptidergic signaling from AUA results in increased adaptation and in line with DOP-1 acting in AUA to negatively regulate neuropeptide release.

Neuropeptide release from AUA increases adaptation. A, B, Fraction of animals responding to repeated drops of 10 mm CuCl2. A, Knockdown of unc-31 RNAi in flp-8-expressing neurons suppresses adaptation. B, Knockdown of unc-31 RNAi in AUA suppresses adaptation. Behavioral data were analyzed using Student's t test. **p < 0.01, ***p < 0.001. For transgenic strains, the average values of two independent strains are shown.
Discussion
We have shown here that sensory adaptation of the ASH nociceptors to soluble repellents is controlled by different food-dependent neuromodulatory pathways. In the presence of food, dopamine acts via the D1-like receptor DOP-1 to inhibit adaptation, whereas in the absence of food, neuropeptides act through the NPY-like receptor NPR-1 to increase adaptation. These pathways are not independent; rather, dopamine modulation of adaptation is dependent on the cell-autonomous activity of NPR-1 in the ASH neurons. Because both DOP-1 activity and neuropeptide release are both required for the adaptation phenotype in the presence of food and both appear to function cell specifically in AUA, we conclude that dopamine acts via DOP-1 to negatively regulate the release of neuropeptides from the AUA interneurons (Fig. 9). In addition to NPR-1, another neuropeptide receptor, NPR-2, also increases adaptation in the absence of food. NPR-1 and NPR-2 are not independent and act in the same pathway to increase ASH adaptation. Thus, the control of food-dependent behavioral states appears to involve a cascade of neuromodulatory peptides and amines that modifies the properties of target neurons such as ASH.

Proposed model for modulation of avoidance on and off food. In the presence of food (right), dopamine acts via DOP-1 to inhibit neuropeptide release from AUA, resulting in low activity of NPR-1 in ASH and allowing ASH to continuously respond to repeated stimulation. In the absence of food (left), DOP-1 is not activated by dopamine, resulting in the release of dense-core vesicles from AUA. Neuropeptides of unknown identity act directly or indirectly to activate NPR-1, which inhibits ASH responses to continued stimulation, resulting in adaptation.
We have shown previously that dopamine has at least two food-related effects on ASH chemosensation: it enhances acute responses to repellents and decreases sensory adaptation (Ezcurra et al., 2011). The effect on acute responses requires the activity of the DOP-4 receptor in the ASH neurons themselves, implying that it is a direct consequence of modulation of the ASH neurons by dopamine. In contrast, because npr-1 is epistatic to dopamine signaling genes with respect to adaptation, the effect of dopamine on adaptation appears to occur through indirect inhibition of an “off-food” neuropeptide signal.
Interactions between neuromodulator systems play a critical role in the control of behavioral states in many organisms, including mammals. In C. elegans, dopamine has been shown to negatively regulate release of octopamine from neurons controlling CREB-dependent gene expression in interneurons (Suo et al., 2006), a process implicated in the response to starvation. Likewise, octopamine acts via the SER-6 receptor to promote the release of peptides that antagonize avoidance responses to octanol (Mills et al., 2012). Finally, under hypoxic conditions, serotonin acts via the SER-7 receptor to promote the release of the NPR-1 ligand FLP-21, thereby enhancing soluble chemotaxis (Pocock and Hobert, 2010). Future studies of these processes in C. elegans may provide insight into conserved mechanisms by which feeding state modifies neural circuits controlling complex behaviors.
This study is not the first to implicate the NPR-1 receptor in the modulation of neuronal activity in response to feeding state. npr-1 was first defined genetically because of its role in aggregation, a behavior strongly influenced by feeding state (de Bono and Bargmann, 1998). npr-1 has been shown subsequently to affect other behaviors directly related to feeding, such as dispersal on a bacterial lawn (Gloria-Soria and Azevedo, 2008), food leaving (Milward et al., 2011), and avoidance of pathogenic bacteria (Styer et al., 2008; Reddy et al., 2009). Interestingly, for these other behaviors, the phenotype of the N2 allele 215V, a gain-of-function variant that arose during cultivation in the laboratory (McGrath et al., 2009), differs from both null alleles and the naturally occurring 215F allele. In contrast, we observed that both npr-1(215V) and npr-1(215F) strains showed “wild-type” adaptation to copper in the absence of food, whereas npr-1 nulls showed abnormal adaptation. Thus, our results indicate that the naturally occurring form of NPR-1 as well as the laboratory-selected form are involved in feeding state-dependent modulation of the ASH neurons. Because NPR-1 shows a high degree of sequence similarity to mammalian NPY receptors, this may suggest a deeply conserved role for these receptors in the control of feeding-related behavioral states. NPR-1 and NPR-2 have been implicated in avoidance responses to methyl salicylate (Luo et al. 2015) but act in a different set of neurons to mediate these responses, suggesting a complex role for these neuropeptide receptors in mediating responses to different noxious stimuli through different neurons.
Our data suggest a model by which dopaminergic signaling regulates neuropeptide signaling to modulate aversive behaviors depending on the feeding state of the animal. Rescue and RNAi data suggest that the AUA neurons act as candidate cells linking dopaminergic and neuropeptide signaling, and we speculate that activation of DOP-1 in these cells results in the inhibition of neuropeptide release. The relevant peptides whose release from AUA might modulate ASH are currently undetermined. The known NPR-1 and NPR-2 ligands FLP-18 and FLP-21 are not required to increase adaptation, suggesting that other peptide ligands may function as NPR-1 and/or NPR-2 ligands to modulate adaption in ASH. Alternatively (or additionally), a more complex network of peptidergic interactions may be involved. Both flp-21- and flp-18-encoded peptides are known to act on multiple receptors in addition to NPR-1 and NPR-2, and several of these target receptors are known to have additional ligands (Peymen et al., 2014). In such a network of complex and overlapping ligand–receptor interactions, individual ligand knock-outs might not be expected to phenocopy knock-outs of individual targets. Moreover, if NPR-1 and NPR-2 function as a dimer (a possibility consistent with our double-mutant analysis), the heteromeric receptor might have distinct signaling properties from either homomeric receptor. Future genetic and biochemical studies may allow us to address these possibilities.
According to the C. elegans wiring diagram, there are no synaptic connections between the dopamine neurons and AUA, nor between AUA and ASH. Thus, our model implies that a sequence of extrasynaptic modulatory connections in the C. elegans nervous system are involved in signaling food availability. Many other examples of amine and neuropeptide signaling in C. elegans (Chase et al., 2004; Kindt et al., 2007; Ezak and Ferkey, 2010; Ezcurra et al., 2011) and mammals (Agnati et al., 1986; Marcellino et al., 2012) are known or suspected to occur extrasynaptically. Thus, neuromodulator signaling forms a wireless network overlaid on the synaptic wiring diagram that is particularly critical for the control of behavioral states.
Footnotes
This work was supported by National Institutes of Health (NIH) Grant R01 DA018341, Medical Research Council Grant MC-A022-5PB91, and Wellcome Trust Investigator Award WT103784MA (W.R.S.) and from the Swedish Research Council, the Torsten Söderberg and Åhlén Foundations, and the Karolinska Institute Strategic Neurosciences Program (P.S.). I.B. is supported by a postdoctoral fellowship of the Research Foundation–Flanders. Some nematode strains used in this work were provided by the Caenorhabditis Genetics Center, which is funded by the NIH National Center for Research Resources. We thank Robyn Branicky and Andre Brown for comments on this manuscript and Emanuel Busch and Mario de Bono for sharing strains and constructs.
This is an Open Access article distributed under the terms of the Creative Commons Attribution License Creative Commons Attribution 4.0 International, which permits unrestricted use, distribution and reproduction in any medium provided that the original work is properly attributed.
- Correspondence should be addressed to William R. Schafer, Medical Research Council Laboratory of Molecular Biology, Cambridge CB2 0QH, UK. wschafer{at}mrc-lmb.cam.ac.uk
This article is freely available online through the J Neurosci Author Open Choice option.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}