

Article Figures & Data
Figures
- Figure 1.
tmc1/2a/2b triple-mutant larvae have auditory/vestibular defects. A, Truncation mutant alleles are shown for tmc1 (frame shift), tmc2a (frame shift), and tmc2b (point mutation) genes. A diagram of each gene is included. Vertical bars represent exons. Horizontal lines indicate introns. 1 cm = 3 kb. Dotted lines indicate the target locus for each gene. Mutant sequences are aligned to corresponding WTs. Dashes represent deleted bases. Spaces represent inserted bases. For tmc1, the 20 bp Cas9 target sequence (bold) and adjacent PAM sequence (underlined) are indicated. For tmc2a, the TALEN-deleted sequence is indicated (bold). For tmc2b, a G > T point mutation yields a TGA stop codon (bold). All tmc1/2a/2b double- and triple-mutant larvae include the tmc1Ex3, +1bp(1) allele; however, the tmc1Ex3, +1bp(2) allele also appears in tmc1/2a mutants used for microphonics assays. The tmc1 single mutants are homozygous for the tmc1Ex3, −5bp allele. B, The AEBR is absent in tmc1/2a/2b triple-mutant larvae (n = 9) at 5 dpf, suggesting complete deafness (WT = 70 ± 20.5%; mutant = 0 ± 0%; t = 10.26; df = 8.0). C, *tmc1/2a/2b triple-mutant larvae show characteristics of vestibular dysfunction. They rest on their sides or backs in a Petri dish without regard for gravitational direction, and they fail to inflate their swim bladders. A WT sibling is included for comparison. D, VIEM in tmc1/2a/2b triple mutants at 5 dpf is lacking, further demonstrating the absence of vestibular function (WT = 1.0 ± 0.22 AU; mutant = 0.1 ± 0.057 AU; t = 7.88; df = 3.26).
- Figure 2.
Gross morphology of stereociliary bundles remains intact in tmc1/2a/2b triple mutants. Hair cell-specific expression of Tg(myo6b:β-Actin-mEGFP-pA) fluorescently labels actin-filled stereocilia in tmc1/2a/2b triple mutants and WT siblings. A, In vivo confocal microscopy of lateral cristae (side view) reveals no bundle splaying or other obvious defects of bundle morphology in tmc1/2a/2b triple-mutant larvae (5 dpf). Scale bar, 2.5 μm. B, Three-dimensional surface rendering of confocal z stacks demonstrates two distinct bundle height groups in centrally located mature hair cells, regardless of genotype. These bundle heights correspond to two hair-cell types in the cristae. Fluorescent patches at the base of bundles are labeling of actin filaments in the cuticular plate. Scale bar, 2 μm.
- Figure 3.
MET components are unaffected in the inner ear (cristae) of tmc1/2a/2b triple mutants (5 dpf). A, Immunohistochemistry shows comparable localization of Pcdh15a (magenta in merge panel) at the tips of stereocilia (green) in tmc1/2a/2b triple mutants. Scale bar, 2 μm. C, E, In vivo fluorescence imaging similarly reveals similar patterns of localization of mEGFP-Lhfpl5a (C) or Tmie-mEGFP (E). Scale bar, 2 μm. B, D, F, Quantification of fluorescence shows no significant differences in expression levels of all three proteins relative to WT levels (ns indicates p > 0.1). B, WT = 504 ± 88.7 AU; mutant = 587 ± 132 AU; t = 1.49; df = 13.79. D, WT = 2927 ± 222 AU; mutant = 2618 ± 132; t = 1.66; df = 7.86.F, WT = 1404 ± 388; mutant = 1449 ± 518; t = 0.175; df = 9.2.
- Figure 4.
Hair cells in tmc1/2a/2b triple mutants lack mechanosensitivity. A, The amplitude of microphonic potentials correlates with stimulus intensity in WT siblings; however, tmc1/2a/2b triple-mutant larvae (3 dpf) have no detectable microphonic potentials, even with high-intensity stimuli (n = 9 for each genotype; F(1,16) = 32.3). B, Otocyst injection of FM 1-43 vital dye, which labels hair cells with intact MET channels, does not label inner ear hair cells in tmc1/2a/2b triple-mutant larvae (6 dpf), further demonstrating a uniform lack of hair-cell activity in all end organs. Side view of inner ear is shown. *Injection damage. AC, Anterior crista; LC, lateral crista; PC, posterior crista; AM, anterior macula; PM, posterior macula. Scale bar, 50 μm.
- Figure 5.
Tg(myo6b:tmc2b-mEGFP-pA) expression rescues vestibular function in tmc1/2a/2b triple-mutant larvae (5 dpf). A, Side view of lateral cristae. Tmc2b-mEGFP localizes to hair bundles and rescues FM 4-64 hair-cell labeling in the tmc1/2a/2b triple mutants (WT, n = 7; mutant, n = 8). Scale bar, 5 μm. B, Tmc2b-mEGFP rescues the VIEM deficit in tmc1/2a/2b triple mutants (mutant = 0.099 ±.0.057 AU; mutant with transgene = 1.26 ± 0.35 AU; t = 6.6; df = 3.1). C, Tmc2b-mEGFP rescues FM 4-64 hair-cell labeling in mutant tmc1/2a/2b anterior maculae (side view) (WT, n = 7; mutant, n = 7). Scale bar, 5 μm.
- Figure 6.
Tg(myo6b:tmc2b-mEGFP-pA) expression rescues hearing in tmc1/2a/2b triple-mutant larvae (5 dpf). A, The AEBR deficit is rescued by Tmc2b-mEGFP in tmc1/2a/2b triple-mutant larvae (mutant = 0 ± 0%; mutant with transgene = 56.5 ± 37.9%; t = 3.65; df = 5.0). B, Tmc2b-mEGFP restores microphonic potentials in tmc1/2a/2b triple mutants (n = 8 rescued larvae; n = 9 for other genotypes; F(2,23) = 14.68). p values (blue) compare tmc1/2a/2b triple mutants with and without Tg(myo6b:tmc2b-mEGFP-pA). C, Tmc2b-mEGFP restores FM 4-64 labeling of hair cells in the posterior maculae (top down view) of tmc1/2a/2b triple mutants (WT, n = 5; mutant, n = 5). Scale bar, 10 μm.
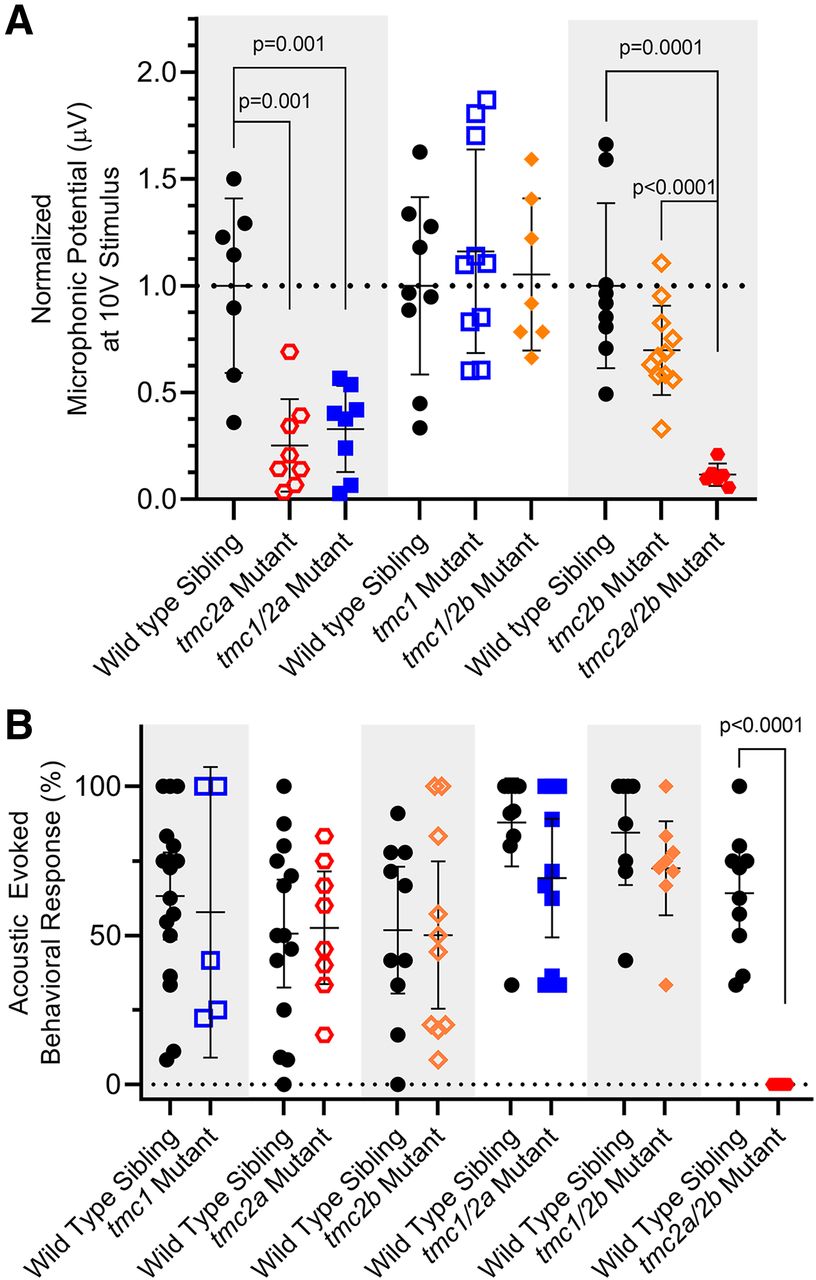
- Figure 7.
Detection of sound by single and double tmc1/2a/2b mutants (5 dpf). Shaded areas in graphs group each mutant genotype with its corresponding siblings. A, Microphonic potentials are significantly reduced in larvae carrying the tmc2a allele (single or double mutants), with the greatest reduction in tmc2a/2b double-mutant larvae. Normalized amplitudes using 10 V stimulus are shown. For tmc2a and tmc1/2a, WT = 1.0 ± 0.41 μV; tmc2a = 0.25 ± 0.22 μV; tmc1/2a = 0.33 ± 0.2 μV. Compared with WT, tmc2a t = 4.34; df = 8.36 and tmc1/2a t = 3.94; df = 8.48. For tmc2a/2b, WT = 1.0 ± 0.39 μV; tmc2a/2b = 0.12 ± 0.05 μV; t = 6.77; df = 8.43. B, Concurrent mutation of tmc2a/2b is sufficient and necessary to eliminate AEBR relative to corresponding WT siblings (WT = 64.2 ± 20.6%; tmc 2a/2b mutant = 0 ± 0%; t = 9.86; df = 9.0).
- Figure 8.
FM labeling of the posterior macula in single and double tmc1/2a/2b mutants (5 dpf). Otocysts of Tg(myo6b:β-Actin-mEGFP-pA) larvae of each genotype were injected with FM 4-64 vital dye. Representative top down views of the posterior macula are shown. GFP-positive hair bundles (green) were overlaid with FM 4-64 label (magenta) in the merge panels. tmc2a/2b double mutants show the strongest reduction in FM 4-64 label throughout the neuroepithelium, with the exception of a subset of hair cells at the periphery of the dorsal posterior region. We examined multiple specimens per genotype (WT, n = 8; tmc1, n = 8; tmc2a, n = 7; tmc2b, n = 5; tmc1/2a, n = 7; tmc1/2b, n = 5; tmc2a/2b, n = 5). Scale bar, 10 μm.
- Figure 9.
Vestibular function in single and double tmc1/2a/2b mutants (5 dpf). Gray shading represents cohorts of siblings. A, Normalized VIEM in single and double mutants. For tmc1/2b comparison, WT = 1.0 ± 0.31 AU; tmc1/2b = 0.34 ± 0.20 AU; t = 4.4; df = 5.3. For tmc2a/2b, WT = 1.0 ± 0.56 AU; 2a/2b = 0.23 ± 0.27 AU. B, Confocal images of hair-cell labeling in the anterior macula (side view) of FM 4-64-injected larvae in the Tg(myo6b:β-Actin-mEGFP-pA) background. GFP-positive hair bundles (green) were overlaid with FM 4-64 label (magenta) in the merge panels. The tmc2a/2b double-mutant larvae show a loss of FM 4-64 hair-cell labeling relative to all other genotypes (background signal is associated with the otolithic membrane). We examined multiple specimens per genotype (WT, n = 8; tmc1, n = 7; tmc2a, n = 5; tmc2b, n = 5; tmc1/2a, n = 6; tmc1/2b, n = 5; tmc2a/2b, n = 5). Scale bar, 10 μm.
- Figure 10.
tmc1/2a/2b single and double mutants reveal molecular differences between two morphologically distinct hair-cell types in lateral cristae (5 dpf). Otocysts of Tg(myo6b:β-Actin-mEGFP-pA) larvae of each genotype were injected with FM 4-64 vital dye. GFP-positive hair bundles (green) were overlaid with FM 4-64 label (magenta) in the merge panels. Type I-like “teardrop”-shaped cells are located in the upper layer, whereas Type II-like, “gourd”-shaped cells are present in the lower layer of the neuroepithelium. *Centrally located mature hair cells that lack FM labeling. We examined multiple specimens per genotype (WT, n = 8; tmc1, n = 7; tmc2a, n = 7; tmc2b, n = 5; tmc1/2a, n = 10; tmc1/2b, n = 5; tmc2a/2b, n = 7). Scale bar, 5 μm.














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}