

Abstract
Different peripheral nerve injuries cause neuropathic pain through distinct mechanisms. Even the site of injury may impact underlying mechanisms, as indicated by the clinical finding that the antiseizure drug carbamazepine (CBZ) relieves pain because of compression injuries of trigeminal but not somatic nerves. We leveraged this observation in the present study hypothesizing that because CBZ blocks voltage-gated sodium channels (VGSCs), its therapeutic selectivity reflects differences between trigeminal and somatic nerves with respect to injury-induced changes in VGSCs. CBZ diminished ongoing and evoked pain behavior in rats with chronic constriction injury (CCI) to the infraorbital nerve (ION) but had minimal effect in rats with sciatic nerve CCI. This difference in behavior was associated with a selective increase in the potency of CBZ-induced inhibition of compound action potentials in the ION, an effect mirrored in human trigeminal versus somatic nerves. The increase in potency was associated with a selective increase in the efficacy of the NaV1.1 channel blocker ICA-121431 and NaV1.1 protein in the ION, but no change in NaV1.1 mRNA in trigeminal ganglia. Importantly, local ICA-121431 administration reversed ION CCI-induced hypersensitivity. Our results suggest a novel therapeutic target for the treatment of trigeminal neuropathic pain.
SIGNIFICANCE STATEMENT This study is based on evidence of differences in pain and its treatment depending on whether the pain is above (trigeminal) or below (somatic) the neck, as well as evidence that voltage-gated sodium channels (VGSCs) may contribute to these differences. The focus of the present study was on channels underlying action potential propagation in peripheral nerves. There were differences between somatic and trigeminal nerves in VGSC subtypes underlying action potential propagation both in the absence and presence of injury. Importantly, because the local block of NaV1.1 in the trigeminal nerve reverses nerve injury-induced mechanical hypersensitivity, the selective upregulation of NaV1.1 in trigeminal nerves suggests a novel therapeutic target for the treatment of pain associated with trigeminal nerve injury.
Introduction
The premise for this study is the clinical observation that pain associated with compression injuries to trigeminal nerves most dramatically manifest in the form of trigeminal neuralgia (TN), is relatively well managed in a subpopulation of patients for at least a limited period of time by the anti-seizure drug carbamazepine (CBZ; Panczykowski et al., 2020), while the same drug has failed in a number of well controlled clinical trials in patients with neuropathic pain arising from somatic structures (Grosskopf et al., 2006; Salinas et al., 2012; Demant et al., 2014), including compression injuries of somatic nerves such as ulnar nerve entrapment and carpel tunnel syndrome (Dosenovic et al., 2017). That there might be a differential response to injury that contributes to this difference in therapeutic efficacy is suggested by evidence that there are developmental (O'Rahilly and Muller, 2007; Steventon et al., 2014) and structural (Oyagi et al., 1989; Chai et al., 2014) differences between trigeminal ganglia (TGs) and dorsal root ganglia (DRGs). There are also differences in gene expression that persist into adulthood documented by comparative RNA sequencing analyses in mouse (Manteniotis et al., 2013), rat (Goswami et al., 2014; Sapio et al., 2016), and human (LaPaglia et al., 2018). And there are differences in the patterns of genes expressed in response to nerve injury (Korczeniewska et al., 2020).
Because CBZ is a voltage-gated sodium channel (VGSC) blocker (Rush and Elliott, 1997), we hypothesized that the differences in the therapeutic efficacy of this drug were because of differences between trigeminal and somatic nerves with respect to injury-induced changes in the biophysical properties, expression, and/or distribution of VGSCs. While CBZ appears to have little VGSC subtype selectivity among heterologously expressed channels (Hinckley et al., 2021), differences in the pharmacological properties of channels in native and heterologous expression systems have been well described (Jarvis et al., 2007). Thus, in contrast to the heterologous expression data, CBZ has subtype selectivity for VGSCs present in sensory neurons (Rush and Elliott, 1997). Furthermore, injury-induced changes in VGSCs have been well described in TGs (Bongenhielm et al., 2000; Liu et al., 2019) and DRGs (Waxman et al., 1994; Novakovic et al., 1998; Gold et al., 2003; Lindia et al., 2005), and available evidence supports a critical role for VGSCs in many chronic pain syndromes (Lai et al., 2004). Importantly, because an increase in VGSC density may increase neuronal excitability to a point sufficient for the emergence of spontaneous activity (Amir et al., 2002), we predicted that the nerve injury-induced change in VGSCs would not only underlie the therapeutic specificity of CBZ, but the pain and hypersensitivity associated with the nerve injury.
To test this hypothesis, we assessed the impact of chronic constriction injury (CCI) of the infraorbital nerve (ION) and sciatic nerve (SN) in male and female rats on the potency and efficacy of CBZ on ongoing pain behavior, as well as CCI-induced mechanical hypersensitivity. With evidence of a selective therapeutic effect on pain behaviors associated with trigeminal nerve injuries, we assessed the extent to which this difference might be because of a differential effect on action potential (AP) propagation in the peripheral nerve. Finally, with evidence that the selective therapeutic effect was due, at least in part, to VGSCs in the peripheral nerve, we assessed the basis for this difference. Pharmacological and protein analysis data pointed to an increase in the relative density in NaV1.1 in trigeminal nerves contributing to both the mechanical hypersensitivity and therapeutic selectivity of CBZ and suggests a novel therapeutic target for the treatment of neuropathic pain associated with trigeminal nerve injury.
Materials and Methods
Experimental design.
The general flow of the experiments in this study was as follows. Rats were purchased as young adults (weight, ∼125 g) and allowed to habituate to the animal facility for at least 3 d before any manipulation. They were then habituated to handling, and any testing apparatus with the experimenter in the room. Rats were then randomized to surgery groups [naive, sham, or surgery (SN or ION)], surgery performed, and then monitored postsurgery for 14 d. Rats were then further randomized to treatment groups (saline, vehicle, or drug), and then used in the different behavioral assays (described below). Different groups of rats were used for control experiments such as the rotarod and the calibration of mechanical stimuli used in the orofacial stimulus test. All behavioral tests were conducted blinded to treatment. After the behavioral testing (∼26 d postinjury), rats were killed, and tissue was collected for biochemical, molecular, and in vitro experiments.
Group sizes were determined by power analysis based on preliminary results, with the general goal of enabling the detection of a 50% difference between groups with a power of 0.8. While not originally powered to detect the influence of sex, sex was included as a factor in the analyses.
Animals.
Adult (weight, 125–150 g; age, 6–7 weeks) male and female Sprague Dawley rats (Envigo) were used in this study. Rats were housed by sex, two per cage in a temperature- and humidity-controlled, Association for Assessment and Accreditation of Laboratory Animal Care International-accredited animal housing facility on a 12 h light/dark schedule. Food and water were available ad libitum. All procedures were approved by the University of Pittsburgh Institutional Animal Care and Use Committee and were performed in accordance with National Institutes of Health Guide for the Care and Use of Laboratory Animals. In addition, every effort was made to both minimize the number of rats used in this study, and any pain or distress associated with the procedures performed.
Peripheral nerve injury.
To enable comparisons between animals with a compression injury to the trigeminal nerve with those with a compression injury to a somatic nerve, the CCI model of compression nerve injury was applied to either the ION or the SN. CCI of the ION was performed as previously described (Kernisant et al., 2008). Briefly, rats were anesthetized with a combination of ketamine (55 mg/kg), xylazine (5.5 mg/kg), and acepromazine (1.1 mg/kg). After sterile preparation of the surgical site, an incision was made above the left eye, the fascia and muscle bluntly dissected from the bone, and the contents of the orbit were gently retracted laterally to free and expose the ION. Once exposed, two pieces of chromic gut 6–0 suture were tied loosely around the ION, tight enough to just constrict blood flow in the perineural vasculature. Sham surgeries were performed in the same way except that the ION was not manipulated. Rats failing to eat, and groom normally would have been removed from the study, but none met these criteria for removal.
CCI of the SN was also performed as described previously (Bennett and Xie, 1988). Rats were anesthetized and the surgical site was prepared as with the ION-CCI, and an incision was made in the back of the thigh. Using blunt dissection, the hamstring muscle along the plane of the muscle was separated, and the SN freed and exposed. Two pieces of chromic gut 6-0 suture were loosely tied around the SN with each loop separated by ∼2 mm, tight enough to constrict perivascular blood flow as with the ION-CCI. Sham operations were performed in the same way except that the SN was not manipulated.
Assessment of orofacial mechanical nociception.
To evaluate changes in the sensitivity to mechanical stimulation of the face, we used the Orofacial Pain Test (catalog #31300, Ugo Basile). This apparatus consists of a plastic cage with 4 × 3 cm portal at one end, through which rats could access the spout of a drinking bottle containing a 5% sucrose solution (Fig. 1). The distance from the portal to the bottle spout was set at 14 mm. The amount of time the rat spent with its face through the portal was quantified with an infrared photograph beam on the spout side of the portal that would be broken when the rat face was extended into the portal. The time rats spent in contact with the drinking spout was also quantified by enabling contact with the spout to close a circuit.
Rats were trained to access the sucrose solution over 5 consecutive days. The first 2 d of the training, food was removed from cages for 12 h before training to increase food-seeking behavior. Training sessions consisted of a 10 min habituation period in the testing chamber followed by 10 min of access to the sucrose solution. Rats were then returned to their home cages. To assess changes in mechanical sensitivity, access to the sucrose solution was impeded by inserting a module with nine wires on each side of the portal. The wires were positioned to ensure contact with the face of the rat when the rat accessed the sipper tube. The number of wires was used to regulate the intensity of the mechanical stimulus, and a wire number was used that produced an ∼50% decrease in the cumulative time the naive rats spent in contact with the drinking tube.
Assessment of mechanical allodynia of the hindpaw.
The von Frey test was used to assess changes in mechanical sensitivity of the hindpaw. Briefly, animals were placed in acrylic boxes with a mesh grid floor and were allowed to acclimate for a minimum of 30 min before testing. Calibrated von Frey filaments (Stoelting) were used to determine the 50% paw withdrawal threshold using the up-down method (Chaplan et al., 1994).
Rats were randomly assigned to ION-CCI, SN-CCI, or their respective sham control groups by cage. They were also randomly assigned to CBZ or vehicle groups, both of which were administered to all animals in a random order 1 d apart. Rats in the ION-CCI/sham groups received an intraperitoneal injection of CBZ or vehicle 30 min before being given access to the sucrose solution. In the SN-CCI/sham group, baseline mechanical threshold was assessed before intraperitoneal administration of CBZ/vehicle and then over the next 3 h. The investigator testing the animals was blinded to the treatment group.
Conditioned place preference test.
Previous studies have suggested the conditioned place preference (CPP) test can be used to assess the presence of tonic or ongoing pain as well as the efficacy of therapeutic interventions for the treatment of ongoing pain (King et al., 2009). Consequently, we used the CPP test to determine whether CBZ could be used to block ongoing pain associated with ION-CCI or SN-CCI. The CPP testing apparatus consisted of two conditioning chambers distinguished by visual and tactile cues that were connected by a middle (neutral) chamber. Before surgery, rats underwent 3 d of preconditioning consisting of full access to all chambers for 30 min/d. On the last of these 3 d, behavior was recorded for 15 min to determine whether rats had a chamber preference. Animals spending >80% (>720 s) of the total time in one chamber were eliminated from further testing (nine rats screened were eliminated based on this criterion). Fourteen days after the respective surgeries, rats were subjected to 6 d of conditioning consisting of alternating treatment–chamber pairings. Rats were placed in one chamber or the other 15 min after receiving an intraperitoneal injection of vehicle, CBZ, or saline. Rats received vehicle–CBZ chamber pairing on odd days and saline–CBZ chamber pairing on even days. Rats spent 15 min in the paired chamber with no access to the other chambers before being returned to their home cage. Drug and chamber pairing were counterbalanced. On the test day, rats were placed in the neutral chamber with access to all chambers during a 15 min observation period. The amount of time spent in each chamber was recorded via video for off-line analysis.
Assessment of motor activity.
Motor coordination was assessed in the rotarod system according to methods previously used (Pineda-Farias et al., 2013). Animals were acclimated for 2 d on the apparatus, which was set to accelerate from 4 to 40 rpm over 10 min. Animals were placed back on the wheel a maximum of five times during this acclimation period. On the third day, animals were tested 30 min after receiving CBZ or vehicle (intraperitoneally). The dependent measure was the latency to fall off of the rotarod. The mean of three trials was used for each rat.
Compound action potential recording from isolated nerves.
Rats were anesthetized and the ipsilateral trigeminal or sciatic nerve was quickly dissected and transferred into a dish containing cold Krebs' solution (∼4°C) with the following composition (in mm): 136 NaCl, 5.6 KCl, 14.3 NaHCO3, 1.2 NaH2PO4, 2.2 CaCl2, 1.2 MgCl2, and 11 glucose oxygenated continuously with carbogen (O2 95%/CO2 5%) to achieve a pH ranging from 7.2 to 7.4.
L5 DRGs in continuity with a section of central process (extending from the T junction toward the dorsal horn of the spinal cord) were also collected from human organ donors with the consent of family members for the use of their tissue for research purposes. The protocol for the collection and study of tissue from organ donors has been approved by the University of Pittsburgh Committee for Oversight of Research and Clinical Training Involving Decedents. The isolated central process from L5 segment was transferred to cold Krebs' solution and processed in the same way as the rat nerves.
The isolated nerve was carefully trimmed of excess connective tissue under a binocular microscope, cut in segments of ∼2 cm. and then transferred to a recording chamber. The endings were mounted in suction electrodes with the peripheral end for electrical stimulation and the central end for recording. The compound AP (CAP) was evoked by a rectangular current pulse (duration, 0.1–0.5 ms) at 0.03 Hz. The electrical stimulus intensity was determined as that necessary to evoke the maximal amplitude of either the A- or C-fiber components of the CAP. The recording electrode was connected to a differential preamplifier (0.1–10 kHz; model DAM-80, WPI), and the recording was sampled at 20 kHz via an Axon Digidata analog-to-digital converter, and acquired and analyzed using pCLAMP version 10 for MS Windows (Molecular Devices). The nerve was superfused continuously with oxygenated Krebs' solution [at room temperature (22–25°C)], and the drugs were delivered via a perfusion pump (Masterflex C/L, model 77120–70, Masterflex). The CAP was recorded before and after drug application. Cumulative concentration–response curves were generated for each drug tested to be able to estimate the potency (equivalent to half-maximal block) and efficacy (maximal block) of each drug, as described below. Each new concentration of drug was applied for 10 min or until the CAP stabilized. CAP data were rectified, and the average of 10 consecutive CAPs was integrated to quantify A- and C-fiber components of the CAP as the area under the curve (AUC).
Quantitative real-time PCR.
Total RNA from ipsilateral TGs and L4-L5 DRGs was isolated by the guanidinium-thiocyanate method with TRIzol reagent according to manufacturer instructions (Thermo Fisher Scientific). Concentration and purity of the RNA were determined by measuring the absorbance at 260 and 280 nm. cDNA was reverse transcribed from equal amounts (1 μg) of total RNA as per SuperScript II First Strand Synthesis System for RT-PCR (Thermo Fisher Scientific), and Oligo(dT)20 primers. Real-time PCR was performed using the iTaq Universal SYBR Green Supermix (BIO-RAD) with a primer final concentration of 300 nm. The thermal cycling protocol used was initial denaturation at 95°C for 30 s, followed by 40 cycles of denaturation at 95°C for 5 s, and an annealing/extension step at 60°C for 30 s. In addition, the analysis of the melting curve was performed to ensure false-positive results, such as those because of primer dimers. Primers were designed and screened for target specificity using Primer-Blast (National Center for Biotechnology Information). The primer sequences used in the present study are shown in Table 1. Several housekeeping genes were included in initial experiments to identify genes that showed no change in CT values as a function of nerve (SN vs ION) or injury (naive, sham, or CCI). 18S met this criterion and was used as an internal control (housekeeping gene) for subsequent analysis of VGSC expression.
PCR Primers used for semiquantitative analysis of voltage-gated Na+ channel α-subunits
The method of Pfaffl (2001) was used to determine the relative expression levels of NaV subunits and 18S. Amplification efficiency (E) was determined for each primer set and was calculated from the slope in the CFX Maestro software (BIO-RAD).
Western blotting.
The ipsilateral trigeminal and sciatic nerves from rats were homogenized in ice-cold RIPA lysis buffer (Thermo Fisher Scientific) containing 1× Halt Protease and Phosphatase Inhibitor Cocktail (Thermo Fisher Scientific). Homogenates were centrifuged at 4°C for 10 min at 14,000 rpm, and the supernatant fraction was used to measure protein concentration by a BCA Protein Assay Kit (Thermo Fisher Scientific) according to manufacturer instructions. A total protein concentration of 200 µg was separated by NuPAGE 4–12% Bis-Tris gels (Thermo Fisher Scientific) using MOPS Buffer (Thermo Fisher Scientific), and then transferred to nitrocellulose membranes. Membranes were blocked with 5% blotting-grade blocker (BIO-RAD) in 1× PBS at pH 7.4 (Sigma-Aldrich) with Tween 0.05%, and they were incubated at 4°C overnight with rabbit anti-NaV1.1 (1:150; catalog #ASC-001, Alomone Labs), rabbit anti-NaV1.3 (1:200; catalog #ASC-004, Alomone Labs), rabbit anti-NaV1.5 (1:200; catalog #ASC-005, Alomone Labs), rabbit anti-NaV1.6 (1:200; catalog #ASC-009, Alomone Labs), rabbit anti-NaV1.7 (1:200; catalog #ASC-008, Alomone Labs), rabbit anti-NaV1.9 (1:200; catalog #ASC-017, Alomone Labs), or a mouse monoclonal antibody directed against β III tubulin (1:1000; catalog #ab78078, Abcam). Horseradish peroxidase-conjugated secondary antibodies (1:9000; anti-rabbit, catalog #111–036-144; 1:3000; anti-mouse, catalog #115–036-146, Jackson ImmunoResearch Laboratories) were applied for detecting the primary antibody signal using an enhanced chemiluminescence detection system (GE Healthcare). β-Tubulin was used as a loading control to normalize protein levels. Imaging of the membranes was performed in an image analysis system (LAS-3000, Fujifilm), and the bands were quantified by densitometry using an image analysis program (LabWorks, UVP). β-Tubulin was used as loading control because it displayed a consistent expression across the groups. β-Actin was assessed as well, but we found that β-actin levels changed as a function of nerve injury. In addition, control peptides for all the NaV subtypes were used to determine the specificity of the immunoreactivity and to identify any bands that were not eliminated when primary antibodies were preadsorbed with the corresponding antigenic peptides.
Drugs.
Carbamazepine was purchased from Sigma-Aldrich and dissolved in Tween 80. Tween 80 (1% in saline) was used as the vehicle for systemic administration, or ≤1% dimethylsulfoxide (DMSO) for the electrophysiological recordings. 4,9-Anhydrotetrodotoxin (4,9-anhydro-TTX) was purchased from Alomone Labs, dissolved in ≤1% ethanol, and tested at concentrations between 1 and 100 nm. ICA-121431, PF-05089771, and A-803467 were also purchased from Alomone Labs, but were dissolved in ≤1% DMSO, and tested at concentration ranges between 3 and 1000 nm, 3 and 100 nm, and 1 and 1000 nm, respectively. Phrixotoxin-3 was purchased from Tocris Bioscience, dissolved in water, and tested at concentration ranges between 0.5 and 1000 nm.
Data analysis.
For the electrophysiological recordings, concentration–response curves were constructed by plotting the fractional inhibition (1 – drug/baseline) versus concentration and the experimental points fitted with a modified Hill equation, enabling estimation of potency (EC50), efficacy (Emax), and the Hill coefficient (n). Because these values were obtained for each nerve with each drug tested, statistical comparisons were made between groups defined by nerve (ION vs SN), and injury (naive, sham, CCI).
Statistics.
Statistical differences between two groups were determined by the Student's t test. One-, two-, three-, or four-way ANOVA, followed by Student–Newman–Keuls or Bonferroni test, were used to compare differences between more than two groups, with two- to four-way ANOVAs used to assess the presence of significant main effects and/or interactions between factors including sex, nerve, injury, and drug administration. Analyses were performed with GraphPad Prism (version 8.0; GraphPad) and SigmaPlot (version 14.0; Systat Software) software packages. Differences were considered statistically significant at < 0.05. Unless otherwise noted, data are plotted as the mean ± SEM with data for individual animals overlaid.
Results
Characterization of the Orofacial Pain Test
While there have been a number of modifications to the application of the von Frey hair test for the assessment of craniofacial hypersensitivity (Ren, 1999; Kernisant et al., 2008), the response quantified is subjective, and approaching the face with a von Frey hair may be just as likely to evoke an attack response as a withdrawal (Kernisant et al., 2008). Thus, we sought to use a more objective measure of mechanical hypersensitivity of the face. The Orofacial Pain Test (Ugo Basile) is a forced-choice assay, where rats must endure mechanical stimulation of the face to access a 5% sucrose solution (Fig. 1). An advantage of the testing apparatus is that it is possible to assess the impact of experimental manipulations on hedonic drive or depression associated with chronic pain as well as the impact of experimental manipulations on hedonic drive, as measured by sucrose consumption in the absence of mechanical stimulation. Thus, we first assessed the impact of CCI on sucrose consumption in the absence of mechanical stimulation (Fig. 1B,C). There was no detectable influence of CCI on sucrose consumption as measured by total contact time with the sipper tube over the 10 min test interval (Fig. 1C). Nor was there a detectable influence of CBZ (50 mg/kg, i.p.) on sucrose consumption (Fig. 1D), suggesting that any changes in drinking behavior associated with CBZ were because of an influence on mechanical sensitivity per se.

The Orofacial Pain Test. A, Setup for the Orofacial Pain Test. Top row, Pictures of the testing chamber and the port providing access to the sucrose solution. Bottom row, rat accessing the sipper tube, with box highlighting mechanical stimulation of the face, and then the different inserts providing incrementally more mechanical stimulation as the number of wires increases from 12 to 24. The amount of mechanical stimulation can also be modified by changing the distance between the window and the sipper tube. Drinking behavior is quantified by the number of nose pokes through the window (via infrared beam breaks) and via the number and duration of contacts with the sipper tube. Five days of training was generally sufficient to generate a consistent pattern of drinking behavior across the 10 min test period and required no additional food or water restriction. B, The cumulative contact time was comparable in male and female rats. Total contact time proved to be the most sensitive variable with which to assess the impact of mechanical stimuli and nerve injury. Data are the mean ± SEM for naive female (n = 18) and male (n = 24) rats. C, The impact of CCI of the ION on sucrose consumption was assessed with no wires creating a barrier to the 5% sucrose. Data were analyzed with a two-way ANOVA (sex × injury) but revealed no significant main effects or a significant interaction between the two. D, The impact of carbamazepine (50 mg/kg, i.p.) on sucrose consumption was assessed and analyzed as in A. No significant differences between groups were detected. E and F, To determine the impact of mechanical stimulation (number of wires in the module between the rat and the sipper tube) on drinking behavior, we assessed the impact of the number of wires on total contact time in groups of rats defined by nerve injury (naive, sham, CCI) and sex. Data were analyzed with a three-way ANOVA. Significant main effects and/or interactions are indicated above the graphs, and the results of post hoc comparisons are indicated within the graphs. Data in all panels and subsequent figures are plotted as the mean ± SEM for five or more animals per group, and points over the bars represent data for individual animals. In this and subsequent figures: *p < 0.05; **p < 0.01.
We next sought to identify a level of mechanical stimulation sufficient to attenuate drinking behavior in naive and sham rats enough to be able to detect an analgesic effect, but not so much that we could not detect the influence of nerve injury. The Ugo Basile apparatus comes with three interchangeable mechanical stimulation modules, where mechanical stimulation is varied by the number of wires on each side of the module, with 6, 9, or 12 wires on each side, for a total of 12, 18, and 24 wires. Mechanical stimulation could also be varied by altering the distance from the nose poke port to the sipper tube. This variable was fixed at 14 mm and was not altered in the present study. The 12-wire module reduced contact time to ∼50% of that without wires in naive and sham rats (Fig. 1A,E). The 18-wire module reduced drinking behavior by ∼65%, while the 24-wire module reduced drinking behavior by >80% (Fig. 1E). In CCI animals, the 12-wire module reduced drinking behavior by 8.75 ± 5.43%, the 18-wire module by 85.97 ± 2.43%, and the 24-wire module by 28.32 ± 12.51% (Fig. 1E), relative to the contact times observed in naive and sham rats with each wire module. Because of the reduction in drinking behavior associated with the 24-wire module in naive/sham rats, the relative impact of nerve injury was dramatically reduced. And while it was possible to detect the presence of CCI-induced hypersensitivity with the 12-wire module, the reduction was relatively small and as a result, it would have been more difficult to detect an antinociceptive effect of experimental manipulations. Thus, we used the 18-wire module for subsequent experiments.
Effect of CBZ on perioral/facial mechanical sensitivity
There is disagreement in the literature as to the therapeutic efficacy of CBZ in preclinical models of nerve compression injuries (Idänpään-Heikkilä and Guilbaud, 1999; Decosterd et al., 2004; Lau et al., 2013; Deseure and Hans, 2017). Nevertheless, because CBZ has failed in clinical trials of neuropathic pain in which other VGSC blockers have been shown to have efficacy, we sought to minimize potential differences between these previous studies by assessing the impact of CBZ on hypersensitivity associated with trigeminal and somatic nerve injuries induced and assessed by the same investigator. Ninety-four animals (47 males, 47 females) were used in the Orofacial Pain Test. None were excluded because of the failure to reach criteria (cumulative contact time, >150 s) in the assay.
Consistent with our pilot experiments with the 18-wire module in place, CCI of the ION was associated with a significant reduction in total contact time with the sipper tube (contact time) in both males and females, and the CBZ vehicle had no impact on this change (Fig. 2). Furthermore, CBZ, at the highest dose tested (50 mg/kg), had no significant influence on contact time in either male or female sham or naive rats (Fig. 2). However, the 50 mg/kg dose partially reversed the CCI-induced suppression of contact time in males, and completely reversed this suppression in females. Data were analyzed with a three-way ANOVA and revealed a significant interaction among sex, drug, and nerve injury (p < 0.01). Post hoc analysis confirmed that the difference between the males and females with respect to the response to CCI in the CBZ groups was significant (p < 0.01), while the CBZ-induced reversal of CCI-induced hypersensitivity was significant in both males and females. Finally, having demonstrated complete recovery in CCI females with 50 mg/kg CBZ, another group of female rats was tested with 25 mg/kg. In contrast to the first experiment where rats were randomized to vehicle or CBZ groups and received a single injection, in this second experiment rats were tested twice and received either vehicle or CBZ in a blinded manner. This dose was also associated with a significant elevation in contact time (Fig. 2). Together, these observations not only support the utility of the Orofacial Pain Test for the objective assessment of mechanical sensitivity in the perioral/facial area of rats, but also indicate that the mechanical hypersensitivity associated with a trigeminal nerve injury in the rat is sensitive to CBZ.

Impact of systemic carbamazepine (CBZ) on mechanical sensitivity assessed with the Orofacial Pain Test. Mechanical nociception in the orofacial region was assessed in the Orofacial Pain Test as described in Materials and Methods and Figure 1. Groups were defined by injury [naive, sham, chronic constriction injury (CCI) of the infraorbital nerve (ION)], drug administration [none, vehicle (Veh), or CBZ, where the systemic dose of CBZ in milligrams per kilogram administered intraperitoneally is indicated by the numbers], and sex (female and male). In this and subsequent figures, significant main effects and/or interactions are indicated above the plots, and the results of post hoc comparisons are indicated in the plots.
Effect of CBZ on hindpaw mechanical sensitivity
The von Frey hair test was used to assess the impact of CBZ on the mechanical hypersensitivity associated with CCI of the SN. Forty-eight rats (24 males and 24 females) were used for this experiment. The impact of CBZ on changes in mechanical sensitivity associated with CCI of the SN was assessed with a four-way mixed design (repeated-measures) ANOVA (sex × nerve injury × drug × time). Results of this analysis revealed significant main effects of nerve injury (p < 0.01), drug (p < 0.05), and time (p < 0.01), as well as a significant interaction between drug and time (p < 0.01), but no effect of sex or significant interaction between sex, drug, and/or nerve injury (Fig. 3A,B). The absence of a significant interaction between nerve injury and drug appeared to be because of a CBZ-induced increase in withdrawal threshold in both sham and CCI animals. To further explore this effect, data were analyzed as an area under the curve defined by the withdrawal threshold over time. Because there was no significant influence of sex, data from male and female rats were pooled (Fig. 3C). These data were analyzed with a two-way ANOVA, which again supported the presence of significant main effects associated with nerve injury and drug, but no significant interaction between the two. Thus, as expected, CCI was associated with a significant reduction in mechanical threshold. Similarly, there was a small but significant elevation of mechanical threshold in animals receiving CBZ but not vehicle.

Impact of systemic carbamazepine on mechanical sensitivity assessed with the hindpaw von Frey Hair test. A and B, Mechanical nociception in the ventral surface of the hindpaw was assessed with the von Frey Hair test. Groups tested are indicated, where CCI-SN indicates CCI of the sciatic nerve. CBZ (50 mg/kg, i.p.) was administered 15 min before the start of testing, and behavior was assessed over 3 h, as indicated. Statistical analysis of these data with a three-way ANOVA revealed a significant main effect of injury, and a significant interaction between drug and time, but no effect of sex. C, To more easily visualize the influence of injury and drug on mechanical threshold assessed with von Frey hairs, data were analyzed as an AUC defined by the paw withdrawal threshold over the 3 h of testing. Furthermore, because there was no influence of sex, data from male and female rats were pooled. Analysis of these data with a two-way ANOVA revealed the main effects of injury and drug, but no significant interaction between the two. D, The impact of CBZ on gross motor function was assessed with the rotarod test. The performance score for each animal on each test day was generated from the mean of three trials. Groups of male and female rats were tested on 2 separate days (3 d apart) 15 min after receiving an injection of CBZ (50 mg/kg, i.p.) or vehicle. Drugs were given in a random order after 3 d of training on the apparatus. Data were analyzed with a two-way ANOVA, which revealed a significant influence of CBZ on motor performance (but not influence of sex or a significant interaction between sex and treatment).
While the timing of the tests for changes in mechanical sensitivity of the face and hindpaw were similar, the sedation associated with CBZ administration was more apparent during von Frey hair testing. As this assay depends on a motor response, we next sought to determine whether motor impairment could have contributed to the von Frey hair results. The rotarod test was used for this purpose. Consistent with the presence of motor impairment, the performance on the rotarod test was significantly (main effect of treatment, p < 0.01, two-way ANOVA) impaired in both male and female rats 30 min after an injection of CBZ (50 mg/kg, i.p.; Fig. 3D).
It is worth noting that because the differences between the results obtained with the Orofacial Pain Test and von Frey hair test may have been because of differences in the assays, subsequent experiments were designed to directly address this concern. Nevertheless, in contrast to CCI of the ION, the influence of CBZ was dependent on neither sex nor the presence of SN injury.
Effect of CBZ in the conditioned place preference test
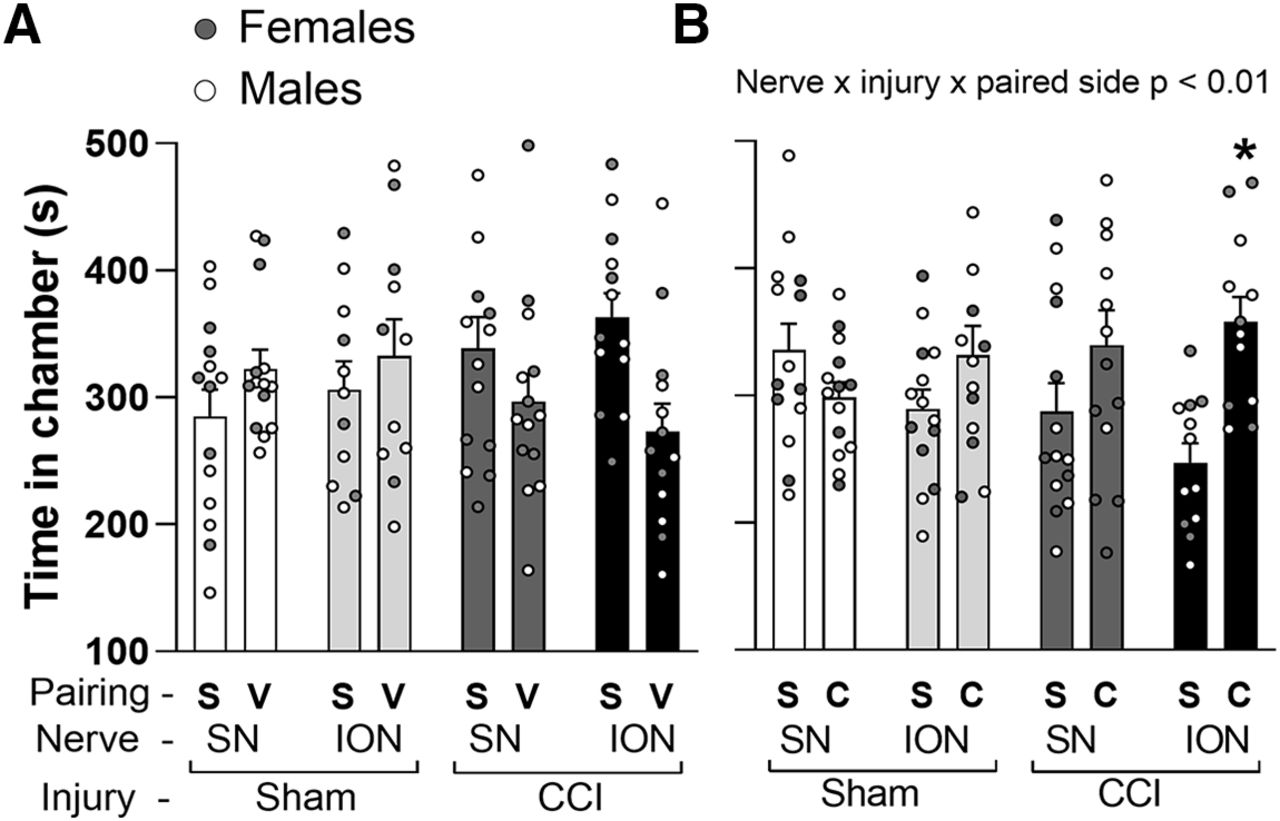
Having obtained evidence of a selective influence of CBZ on mechanical hypersensitivity associated with CCI of the ION, we next sought to determine whether there was a differential influence of CBZ on ongoing pain behavior in rats with CCI of the ION or SN. One hundred twenty-three animals (68 males, 55 females) were used in the CPP assay. Nine were excluded because they demonstrated a place preference in the baseline screening phase. Male and female rats were randomized to ION-CCI/sham and SN-CCI/sham groups. We first assessed the impact of CBZ vehicle in the CPP assay, and data were analyzed with a three-way ANOVA with groups defined by sex (male vs female), site of manipulation (ION vs SN), and injury (CCI vs control). The results of this analysis indicated that there were neither significant main effects nor interactions among any of the three factors (p > 0.05; Fig. 4A). In other words, CBZ vehicle was associated with no place preference, regardless of sex, site of injury, or the presence of injury. The same analysis was then performed on rats that received CBZ paired with one chamber in the testing apparatus and saline paired with the other. The results of this analysis revealed a significant interaction between the side of pairing, site of injury, and the presence of injury (p < 0.01; Fig. 4B), but no influence of sex. That is, both male and female rats with CCI of the ION but not SN exhibited a small, but significant, preference for the CBZ paired side.

Impact of systemic carbamazepine (CBZ) on ongoing pain assessed with the conditioned place preference assay. A, There was no influence of CBZ vehicle on conditioned place preference, as assessed with the time spent on paired chambers. B, However, when chambers were paired with saline (S) and CBZ (C) in sham and chronic constriction injury (CCI) animals studied 28–35 d postsurgery, there was a significant interaction among nerve, injury, and paired chamber because of a place preference in the CCI-Infraorbital nerve (ION) group but not in the CCI-sciatic nerve (SN) group.
Effect of CBZ on the A and C components of the CAP
Because CBZ administered systemically could have been attenuating mechanical hypersensitivity at sites throughout the neuraxis, we next determined whether the apparent nerve injury-induced increase in potency, at least as observed with CCI of the ION was associated with a change in the potency of CBZ-induced block of the peripheral nerve. Fifty-two rats (26 male and 26 female) were used for this part of the study. It was possible to obtain a sufficient length of both the ION and SN to clearly resolve the more rapidly conducting A-fiber component (A-wave) and more slowly conducting C-fiber component (C-wave) of the CAP (Fig. 5A,B). Cumulative concentration–response data were collected from both the ION and SN, 26–30 d after sham or CCI surgery (Fig. 5B–D). Because it was possible to generate estimates of potency (EC50) for each nerve, groups were statistically compared based on this parameter. Data were analyzed with a three-way ANOVA [sex (male vs female) × nerve (ION vs SN) × injury (sham or CCI)]. Data for the A-wave (Fig. 5E) and the C-wave (Fig. 5F) were analyzed separately. The results of this analysis revealed a significant (p < 0.01) interaction between nerve and injury for both components of the CAP, and an interaction (p < 0.01) among sex, nerve, and injury for the C-wave. Strikingly, there was no influence of nerve injury on the potency of CBZ-induced block of the SN, while there was a dramatic increase in potency in the block of the ION. The effect was even more pronounced for the block of the C-wave, which was sex dependent because of the larger increase in potency associated with CCI of the ION observed in females. These changes are consistent with the behavioral results and suggest that direct block of the ION may underlie the therapeutic actions of CBZ in the presence of CCI.

Impact of nerve injury the potency of carbamazepine (CBZ)-induced block of action potential propagation in isolated peripheral nerves. A, Diagram of the setup used to evoke and record compound action potentials (CAPs) in isolated peripheral nerves. B, Typical example of a CAP evoked in an isolated sciatic nerve, where the more rapidly conducting A-wave (Aβ) can easily be distinguished from the more slowly conducting C-wave (C) based on the latency to arrive at the recording electrode. Insets, Typical examples of the impact of increasing the concentration of CBZ on the A-wave recorded from an infraorbital nerve (ION) obtained from animals ∼28 d after sham (SH) surgery or injury (chronic constriction injury (CCI)). C, D, Pooled concentration response data for the C-wave recorded in ION (C) and sciatic nerve (SN) (D) isolated from sham and CCI female rats. E, F, CBZ was bath applied, and the CAP was recorded until the amplitude of the waveform had stabilized. EC50 data for the A-wave (E) and C-wave (F) are plotted.
To determine the extent to which the differences between the ION and SN observed in the rat could account for differences in therapeutic efficacy of CBZ for the treatment of neuropathic pain in humans, we assessed the potency of CBZ-induced block of action potential propagation in human peripheral nerves. The central process of L5 ganglia (Fig. 6A) were obtained from seven organ donors (five males and two females). The average age of the donors was 48 years with a range of 23–66 years. The central process of five trigeminal nerves (Fig. 6B) was obtained from cadavers (two males and three females) within 8 h of death. The average age of the cadavers was 67 years with a range of 33–85. The CAP was evoked as described for rat nerves (Fig. 6C), and cumulative concentration–response data were generated (Fig. 6D–G). The EC50 values for block of both the A-wave (Fig. 6D,E) and the C-wave (Fig. 6F,G) in the L5 central process were comparable to that obtained for block of the rat SN. In contrast, the potency of CBZ-induced block of the C-wave in human trigeminal nerve (Fig. 3C) was higher than that observed in the central process of the L5 ganglia (Fig. 6D–G). There was also considerably more variability between trigeminal nerves, with two exhibiting a potency comparable to that observed following CCI of the rat ION. Of note, the A-wave on the first four trigeminal nerves recovered was not stable enough to generate a complete CBZ concentration–response curve. Thus, the CBZ effect on the A-wave from only one nerve is plotted in Figure 6, D and E.

Effect of carbamazepine (CBZ) on human peripheral nerves. A, L5 ganglion recovered from an organ donor, with the spinal nerve, ganglia (DRGs), and central roots indicated. B, Top, Photograph of a trigeminal ganglia in situ, just before recovery from a cadaver enrolled in a rapid recovery program. Bottom, Trigeminal ganglia after recovery, with the central root process for recording indicated. C, Examples of CAP recorded from human trigeminal nerve (TN) (top trace) and central process of the L4 ganglia (somatic nerve (SN); bottom trace). D, Impact of increasing concentrations of CBZ on the A-wave evoked from somatic (n = 7) and TNs (n = 1). Data were analyzed and pooled as in Figure 2. E, Pooled EC50 data for the nerves plotted in D, where data for each nerve illustrate the variability in the EC50 values obtained. Of note, the A-wave from four of the five TN donors was not stable enough to obtain a full concentration–response curve, so only a single data point is plotted. F, Impact of increasing concentrations of CBZ on the C-wave evoked from SNs (n = 7) and TNs (n = 5). Data were analyzed and pooled as in Figure 2. G, Pooled EC50 data for the nerves plotted in F, where data for each nerve illustrate the variability in EC50 values obtained.
Effect of NaV blockers on the A and C components of the CAP
Given that CBZ is thought to be a VGSC blocker, we next sought to determine whether there was a nerve injury-induced shift in the channel subtypes underlying the propagation of action potentials in the ION and SN that could account for the ION-induced increase in CBZ potency. Forty-eight animals (24 males and 24 females) were used for this part of the study. Concentration–response data were collected for a panel of VGSC blockers applied to ION and SN from rats with sham and CCI, enabling comparisons of changes in both efficacy (maximal suppression) and potency (EC50; Fig. 7A–L). A plot of the maximum fractional block (Emax) for the A- and C-waves of the SN and ION from sham and CCI groups revealed marked differences between nerves and fiber types in the absence of injury as well as in the response to injury (Fig. 8A,B). Interestingly, in contrast to the injury-induced changes in blocker sensitivity observed in the ION, there was little detectable change in blocker sensitivity in the SN.

Impact of nerve injury on the potency of voltage-gated sodium channel (VGSC) blockers on action potential propagation in isolated peripheral nerves. Compound action potential recording was performed as described in Figure 5 on infraorbital nerve (ION) and sciatic nerve (SN) isolated from rats with sham or chronic constriction injury (CCI) surgery. A, B, The A-wave (A) and C-wave (B) were analyzed separately. Concentration–response data were collected for each VGSC blocker on different nerves, and the fractional block produced by a saturating concentration for each was calculated. Examples of the concentration–response data obtained with ICA-121431 on the ION from sham and CCI rats. IC50 (potency) was determined from the concentration–response curve fitted with a modified Hill equation. The average potency for each blocker tested is plotted, where data from individual nerves from females (closed symbols) and males (open symbols) are overlaid on the bar graphs. C–L, The blockers assessed included the following: ICA-121431 (NaV1.1 and 1.3; C, D); 4,9-anhydro-TTX (NaV1.6; E, F); phrixotoxin-3 (NaV1.2, NaV1.3, and NaV1.5; G, H), PF-05089771 (NaV1.7; I, and J); and A-803467 (NaV1.8; K, L). Data were analyzed with a three-way ANOVA (nerve × injury × sex). The presence of a significant interaction is indicated, as are significant differences between groups as determined with post hoc testing.

Impact of nerve injury on the relative efficacy of voltage-gated sodium channel (VGSC) blockers on action potential propagation in isolated peripheral nerves. Efficacy (maximal fractional block) was determined from the concentration–response data as described in Figure 7. A, B, To illustrate differences between the ION and the SN with respect to the response to different VGSC blockers, pooled data for each blocker on the A-wave (A) and C-wave (B) from nerves from each group of rats are plotted. Of note, dotted lines connect data from each group of rats to more clearly illustrate differences between groups with respect to the pattern of block. As noted, however, blockers were applied to different nerves and not in sequence to the same nerve. C–L, The potency data for each blocker assessed on the A-wave (left panels) and C-wave (right panels) are plotted. As with the potency data plotted in Figure 7, data were analyzed with a three-way ANOVA (nerve × injury × sex), and the presence of significant main effects and/or interactions are indicated above the graphs, while significant post hoc comparisons are indicated in the graphs.
The impact of each blocker was analyzed with a three-way ANOVA (sex × nerve × injury). There was a significant interaction between nerve and injury (p < 0.01) with respect to the block of the A-wave (Fig. 8C) and C-wave (Fig. 8D) by ICA-121431. The efficacy was increased in CCI-ION but not CCI-SN. In contrast, while there was a significant interaction between nerve and injury (p < 0.01) with respect to the block of the A-wave by 4,9-anhydro-TTX (Fig. 8E), there was no influence of nerve injury on the actions of this blocker against the C-wave (Fig. 8F). Of the other blockers tested (Fig. 8G–L), the only one in which significant differences were detected was for phrixotoxin-3, which produced a greater block of the ION than SN, regardless of sex or injury for both the A- and C-wave.
While several of the blockers used, such as 4,9-anhydro-TTX, PF-05089771, and A-803467 are reported to be highly selective for NaV1.6 (IC50 = 7.8 nm; Rosker et al., 2007), NaV1.7 (IC50 = 11 nm; Alexandrou et al., 2016), and NaV1.8 (IC50 = 8 nm; Jarvis et al., 2007), respectively, ICA-121431 (NaV1.1/ NaV1.3; IC50 = 19 nm; McCormack et al., 2013) and phrixotoxin-3 [NaV1.2 IC50 = 0.6 nm (Bosmans et al., 2006); NaV1.6 IC50 ∼100 nm (Yin et al., 2017)] have activity at several subunits. Thus, a change in the relative contribution of a subunit might result in a shift in the apparent potency of either of these blockers. To assess this possibility, we analyzed the impact of nerve and injury on blocker potency as estimated from the concentration–response data. Consistent with this prediction, there was a significant interaction between nerve and injury (p < 0.01) with respect to the potency of ICA-121431, at least for block of the C-wave (Fig. 7D): the potency was higher in ION-CCI nerves (from male and females) relative to sham surgery, but not in SN nerves. No changes in potency were detected for phrixotoxin-3 or any of the other channel blockers screened (Fig. 7E–L).
Modulation of the NaV subunit levels in ION- and SN-injured rats
Our pharmacological data suggested that compression injury of at least the ION was associated with changes in the relative proportion of functional NaV subunits along peripheral axons. To determine whether these apparent changes in functional channels were associated with a change in subunit mRNA, we used semi-quantitative PCR to assess the impact of compression injury on subunit mRNA levels in the TGs (Fig. 9A) and DRGs (Fig. 9B) from naive rats, and those with sham injury or CCI. Similarly, Western blot of the peripheral nerves was used to quantify changes in subunit protein.

Impact of nerve injury on NaV mRNA levels in the DRGs and TGs. mRNA levels from naive, sham, and CCI groups were first normalized levels of the housekeeping gene 18S. Relative expression, calculated as described in Materials and Methods, is plotted for each transcript. A, B, While data for TGs (A) and DRGs (B) are plotted, data were analyzed with a three-way ANOVA (sex × ganglia × nerve manipulation). There was a significant interaction between ganglia and nerve manipulation for NaV1.3, which was because of the selective injury-induced increase in NaV1.3 mRNA in DRGs.
While changes in mRNA NaV1.3, NaV1.6, and NaV1.9 were detected in association with CCI, none were consistent with the pharmacological results obtained. That is, there was a significant interaction between nerve and injury (p < 0.01) for the expression of NaV1.3, but this was because this subunit was increased in injured SN but not ION (Fig. 9B). There were also significant effects of nerve (p = 0.01) and injury (p < 0.01) for the expression of NaV1.6, but this was because of the relatively lower levels of NaV1.6 expression in the TGs, and a decrease in both the TGs and DRGs associated with nerve injury (Fig. 9 compare A, B). The only significant interaction between sex and injury was for NaV1.9 (p < 0.05). But this was because of a decrease in subunit expression in male, but not female, TGs and DRGs associated with CCI (Fig. 9A,B). Of note, while data from TGs and DRGs were plotted in different panels, statistical comparisons were made between nerves and, thus, between panels.
In contrast to the PCR results, there was at least one change in NaV subunit protein levels in the peripheral nerve that were consistent with the pharmacological data. That is, there was a significant interaction between nerve and injury (p < 0.01), because of the relative increase in NaV1.1 in the ION but not SN in association with nerve injury (Fig. 10A,B). There was a significant interaction between nerve and injury (p < 0.01) for NaV1.3, but this was because of the increased levels of NaV1.3 in the SN but not in the ION in association with nerve injury (Fig. 10A,B). The only other changes in nerve NaV protein levels observed was a significant main effect of injury (p < 0.01) for NaV1.7 because of the decrease in protein associated with nerve injury in both the ION and the SN (Fig. 10B). Together, these results suggest that both the behavioral and CAP data with CBZ are due, at least in part, to an upregulation in an ICA-sensitive current, presumably NaV1.1.

Impact of nerve injury on levels of voltage-gated sodium channel (VGSC) subunit-like immunoreactivity in peripheral nerves. A, Raw blots of total protein extracted from the infraorbital nerve (ION) and the sciatic nerve (SN) recovered from naive rats, as well as rats subjected to sham or chronic constriction injury (CCI) probed with antibodies against VGSC subunits. Blots were stripped and reprobed with an antibody against β-tubulin (β-Tub), used as a loading control. All bands were eliminated when antibodies preabsorbed with the VGSC subunit-specific peptide (PEP; left column). B, Pooled data from male and female rats were normalized to the loading control and then to normalized values from naive animals. Data generated with each antibody was analyzed with a three-way ANOVA (nerve × injury × sex). The only significant differences between groups were for NaV1.1 and NaV1.3. For both, there was a significant nerve × injury interaction (p < 0.01). The results of post hoc comparisons are indicated.
Effect of ICA-121431 on the orofacial mechanical hyperalgesia
To directly test the role of an ICA-121431-sensitive current in the mechanical hypersensitivity associated with CCI of the ION, we returned to the Orofacial Pain Test with local administration of ICA-121431 on the ION. Twenty-three rats (11 males and 12 females) were used for this experiment. Consistent with the isolated nerve data, ICA-121431, but not vehicle, reversed the attenuation of sucrose consumption associated with mechanical stimulation in the CCI group (Fig. 11A). The results of this analysis revealed a significant interaction between injury and drug (p < 0.01) because of the selective increase in drinking behavior associated with ICA-121431 administration in the CCI group. However, also consistent with the isolated nerve data, there was no significant impact of sex on the effects of ICA-121431 (p > 0.05). Importantly, and consistent with the selective upregulation of NaV1.1 in the ION, ICA-121431 locally applied to the SN, had no detectable influence on von Frey hair withdrawal thresholds in rats with a CCI of the SN (Fig. 11B).

Impact of the local administration of ICA-121431 on mechanical nociception. ICA-121431 (200 μl of 1 μm) was injected percutaneously around the infraorbital nerve (ION) or sciatic nerve (SN) the site exposed for chronic constriction injury (CCI). A, Mechanical sensitivity in the face was assessed with the Orofacial Pain Test. Pooled total contact time for male and female rats are plotted. To illustrate the impact of CCI on total contact time, baseline data for all animals was included in the first two bars of the graph. B, The impact of ICA-121431 (200 μl of 1 μm) applied to the sciatic nerve of CCI-SN rats (n = 6 male; n = 6 female rats) on von Frey hair withdrawal threshold. As with the data plotted in A, ICA-121431 and vehicle were tested on the same group of animals on separate data, with drugs administered in a random order. There was no detectable influence of ICA-121431 on the CCI-induced hypersensitivity of the hindpaw.
Effect of ICA-121431 CBZ-induced block of the CAP
Finally, because of the increase in efficacy of the ICA-induced block of the CAP associated with CCI of the ION, we sought to determine whether ICA and CBZ were acting on the same target. To address this issue, we tested the effect of increasing concentrations of CBZ in the presence of a fixed concentration of ICA-121431 on the CAP evoked in the ION from rats that received sham surgery or CCI. Consistent with our initial set of experiments, CCI was associated with an increase in the fractional block of the CAP by ICA-121431 (13.1 ± 1.2% vs 37.3 ± 2.2% for the A-wave and 18.0 ± 2.1 vs 45.1 ± 2.7% for the C-wave, for sham (n = 13) and CCI (n = 11) groups, respectively). In the presence of ICA-121431, there was also no significant difference between CCI and sham groups with respect to the potency of CBZ for the block of the A-wave (Fig. 12A,B) and a small, but significant decrease in potency for the block of the C-wave (two-way ANOVA revealed a significant main effect of injury (p = 0.02). However, the absence of an effect on the A-wave, and the small difference in the C-wave appeared to be because of an increase in the potency of CBZ when in the presence of ICA-121431, as observed in nerves from sham animals, rather than a block of the CCI-induced increase in potency in nerves from CCI animals (Fig. 12A,B).

Impact of nerve injury on the potency of carbamazepine (CBZ)-induced block of action potential propagation in the presence of ICA-121431 in isolated nerves. Compound action potentials were recorded from isolated infraorbital nerves (IONs) recovered from sham or CCI rats as described in Figure 2. A, Data from the A-wave (left panels) and C-wave (right panels) were analyzed separately. B, ICA-121431 (ICA; 1 μm) was applied after stable recordings were obtained, and then increasing concentrations of CBZ were applied. Concentration–response curves were generated for each nerve, fitted, and pooled.
Discussion
The purpose of this study was to test the hypothesis that differences in the therapeutic efficacy of CBZ for the treatment of trigeminal versus somatic neuropathic pain are because of differences between trigeminal and somatic nerves in the injury-induced changes in the biophysical properties, expression, and/or distribution of VGSCs in sensory neurons. Our behavioral data indicated that CBZ was more effective at reducing ongoing pain behavior and mechanical hypersensitivity associated with CCI of the ION than the SN. Consistent with the behavioral data, CCI of the ION, but not of the SN, was associated with a significant increase in the potency of CBZ-induced block of the CAP. Interestingly, this effect was greater in females than males. Pharmacological data not only revealed differences between the ION and SN with respect to the pattern of block by subtype-selective NaV channel blockers, but differences in CCI-induced changes in the relative efficacy of blockers. Most notable among these was an increase in the efficacy of ICA-121431. While there were no detectable CCI-induced changes in NaV subunit mRNA levels consistent with the changes in CBZ potency or the efficacy of NaV channel blockers, a CCI-induced increase in NaV1.1 protein was detected in the ION but not in the SN. Finally, local administration of ICA-121431 on the ION alleviated the mechanical hypersensitivity in rats with ION-CCI, but appeared to increase the potency of CBZ on the inhibition of the CAP. These data are consistent with our initial hypothesis and suggest that an increase in NaV1.1 may contribute to the therapeutic selectivity of CBZ.
Our behavioral results are consistent with our initial hypothesis and the clinical literature with respect to the efficacy of CBZ for the treatment of neuropathic pain. Unfortunately, the picture emerging from the preclinical literature with this compound in models of persistent pain is far from clear. That is, there are previous reports that CBZ has antinociceptive efficacy (Ardid et al., 2003; De Vry et al., 2004) as well as those in which limited to no efficacy was detected (Fox et al., 2003; Decosterd et al., 2004; Lau et al., 2013) in preclinical models of somatic nerve injury. There are also conflicting reports of antinociceptive efficacy in models of trigeminal nerve injury (Idänpään-Heikkilä and Guilbaud, 1999; Deseure and Hans, 2017). The basis for the discrepant results is not clear, but may reflect the way antinociceptive efficacy was assessed, the nerve injury model used, whether motor deficits were accounted for, as well as the strain of the rats used. For example, while others have used the von Frey hair test on the hindpaw (Ardid et al., 2003; De Vry et al., 2004) and face (Idänpään-Heikkilä and Guilbaud, 1999; Deseure and Hans, 2017), we are the first to use a more objective method for the assessment of nerve injury-induced changes in the efficacy of CBZ for orofacial hypersensitivity. We are also the first to assess the relative efficacy of CBZ in an assay for ongoing pain. Importantly, the results with both of these more objective assays were consistent. With respect to the nerve injury model, CCI is thought to generate hypersensitivity because of the nerve compression as well as the neuritis associated with the chromic gut (Kajander et al., 1996). In contrast, the extent of neuritis associated with the spinal nerve ligation model used in other studies appears to depend on the experience of the surgeon and the approach used (Djouhri et al., 2006). That CBZ may influence hypersensitivity associated with inflammatory processes is suggested by results demonstrating efficacy in the formalin model (Ghorbanzadeh et al., 2019), as well as other models involving inflammatory cascades (Iwamoto et al., 2011). With respect to the side effects associated with CBZ, motor deficits were clearly present following systemic administration of CBZ (Fig. 3C) that we suggest contributed to the elevated withdrawal thresholds in the von Frey hair test. CNS sites of action of CBZ, in particular those associated with memory (Shannon and Love, 2007) may have also impacted conditioning, contributing to the relatively small effects observed in the CPP. Finally, with respect to strain effects, a genetic polymorphism has been shown to impact the efficacy of CBZ with respect to the block of VGSCs (Tanaka et al., 2016; Adi et al., 2018), so strain differences in the efficacy of the drug should not be surprising. Regardless of the basis for these differences in the literature, it is worth pointing out that even studies in which positive results were reported with models of somatic nerve injury, the effects were, in general, relatively small, reaching just 60% of the maximal possible effect (De Vry et al., 2004). This small effect is consistent with our results with the von Frey hair test and with the CAP inhibition with CBZ on the sciatic nerve.
The sex difference observed in the relative efficacy of CBZ was surprising. That is, in contrast to other pain syndromes, such as migraine, temporomandibular disorder, and irritable bowel syndrome, that are more prevalent in women than in men, sex differences in neither prevalence nor severity are generally reported for neuropathic pain (Berkley, 1997). The exception is trigeminal neuralgia, which has a higher prevalence in women than in men (Sekula et al., 2011). Thus, because CCI of the ION has been used as a preclinical model of TN, we included sex as a biological variable in the present study, acknowledging that this pain syndrome is generally manifest in older adults and only young adult rats were used in the present study. Nevertheless, we anticipated, if anything, a more severe and/or longer lasting hypersensitivity in females than in males, as previously reported in response to an infraorbital, but not a sciatic, nerve partial injury (Dominguez et al., 2009). But this is the first demonstration of a sex difference in the pharmacological response to injury of the ION. While the basis for this sex difference has yet to be determined, it raises the intriguing possibility that the mechanisms contributing to this sex difference in therapeutic efficacy in the preclinical model could contribute to the higher prevalence of trigeminal neuralgia in women.
This study is the first to assess the impact of injury and sex on the potency and efficacy of CBZ on the CAP evoked in the ION. These electrophysiological results are consistent with our behavioral data and suggest that while CNS and/or other peripheral targets may contribute to the antinociceptive activity of CBZ, changes in the peripheral nerve critical to action potential propagation are likely to contribute to the therapeutic effects as well. A previous study implicated a gain-of-function mutation in NaV1.6 in the manifestation of trigeminal neuralgia in the patient in which the mutation was identified, because the mutation was associated with an increase in neuronal excitability. More interestingly, the mutation increases the potency of CBZ-induced block of the channel and, therefore, may account for the therapeutic efficacy of CBZ in this patient (Tanaka et al., 2016). A comparable mutation in the peripherally restricted NaV1.7 is also associated with an increase in the potency of CBZ (Adi et al., 2018), suggesting that the therapeutic efficacy of CBZ may be determined by genetics rather than the type or site of injury. In contrast, our results suggest that injury of the trigeminal nerve is sufficient to increase the potency and efficacy of CBZ in the periphery. That said, our data from human trigeminal and somatic nerves suggest that injury-induced changes in the peripheral nerve may not be necessary to account for the therapeutic selectivity of CBZ, given that even in the absence of evidence of neuropathic pain, the potency of CBZ-induced block of the trigeminal nerve was greater than that of somatic nerves.
A key prediction arising from our central hypothesis is that there would be differences between trigeminal and somatic nerves with respect to the effects of VGSC blockers. Consistent with this prediction, we not only observed baseline differences between the SN and ION with respect to the relative efficacy of VGSC blockers, but differences in the response to nerve injury. These results highlighted differences between trigeminal and somatic nerves with respect to channels underlying AP propagation as well as differences between A-fibers and C-fibers with respect to the VGSC underlying AP propagation. These results provide further justification for what has remained a relatively elusive goal of the development of VGSC subtype-selective blockers for therapeutic purposes, in particular for pain management.
We detected nerve type- and nerve injury-specific increases in the relative efficacy of two VGSC blockers consistent with those observed with CBZ: ICA-121431 and 4,9-anhydro-TTX. However, because the nerve injury-induced increase in CBZ potency was larger for the block of C-fibers than A-fibers, and the increase in the efficacy of 4,9-anhydro-TTX was restricted to A-fibers, we focused on ICA-121431. Since ICA-121431 is a selective blocker for NaV1.1 and NaV1.3 subunits, the increase in relative efficacy could reflect an increase in either of the channel subtypes. NaV1.3 is by far the more extensively studied of these two channel subtypes in the context of nerve injury (Waxman et al., 1994). However, while an increase in NaV1.3 has previously been reported in response to injury to the trigeminal nerve as well (Eriksson et al., 2005; Liu et al., 2019), phrixotoxin-3 is reported to be a NaV1.3 blocker with activity at NaV1.2 and NaV1.5 channels. The limited change in the relative efficacy of phrixotoxin-3 argues against NaV1.3 as the channel underlying the increase in efficacy of ICA-121431. Consistent with this interpretation, we detected an increase in NaV1.1 protein, but not in NaV1.3 protein, in the ION associated with CCI. Thus, while the nerve injury-induced increase in the relative efficacy of 4,9-anhydro-TTX suggests that an increase in NaV1.6 channels may contribute to the increase in the potency of CBZ-induced block of the CAP A-wave, our results are most consistent with a dominant role for NaV1.1 in the nerve injury-induced increase in CBZ potency.
Our pharmacological results suggesting that NaV1.6 is the VGSC subunit primarily responsible for AP conduction in both myelinated and unmyelinated axons in the sciatic nerve are consistent with those of several previous reports (Black et al., 2002; Wilson et al., 2011; Gasser et al., 2012; Qin et al., 2017). NaV1.6 has been shown to play a particularly important role in AP conduction in A-fibers (Wilson et al., 2011), despite evidence of NaV1.1 and NaV1.7 expression in this subpopulation of sensory neurons (Fukuoka et al., 2008; Ho and O'Leary, 2011; Usoskin et al., 2015) and anatomic evidence of NaV1.7 protein at nodes of Ranvier in human dental nerves (Luo et al., 2008). Thus, we would have predicted a larger maximal effect of 4,9-anhydro TTX on the A-wave in the sciatic nerve. While limited access of the toxin to NaV1.6 in our preparation could explain this effect, we were able to completely block the A-wave with 500 nm TTX (data not shown). On the other hand, there is evidence from pharmacological experiments (Schmalhofer et al., 2008; Alexandrou et al., 2016; Hoffman et al., 2018) that other channels contribute to AP conduction in A-fibers. Interestingly, in contrast to data suggesting that NaV1.6 contributes to AP conduction in C-fibers (Black et al., 2002) and the data generated with PF-05089771 (Alexandrou et al., 2016), there is a report that AP conduction in C-fibers is entirely dependent on NaV1.7 (Schmalhofer et al., 2008). The basis for these discrepant results is not clear, but may reflect limited selectivity of the compounds used (Zhang et al., 2017), the impact of compensatory changes in the knock-out mouse, and/or the part of the nerve assessed (i.e., the impact of the PF-05089771 compounds on AP propagation was assessed on the central roots; Alexandrou et al., 2016). Furthermore, our failure to detect an impact of the putative NaV1.7-selective blocker PF-05089771 on the CAP C-wave may have reflected state-dependent properties of this compound, which is more effective at inactivated channels (Alexandrou et al., 2016).
Given the compelling evidence from a number of laboratories (Waxman et al., 1994; Lindia et al., 2005) as well as from our own expression data, indicating that somatic nerve injury is not only associated with an increase in NaV1.3 mRNA, but aslo in functional channels (Samad et al., 2013), we were surprised by the limited impact of nerve injury on the relative efficacy of phrixotoxin-3. Our negative results with this toxin, at least in the sciatic nerve from control rats, is consistent with previous data suggesting a minimal contribution of NaV1.1, NaV1.2, and NaV1.3 to AP propagation in the sciatic nerve (Wilson et al., 2011). That said, despite evidence that NaV1.3 contributes to the pain association with nerve injury (Samad et al., 2013), we are unaware of any reports supporting a role for this subunit in AP propagation in peripheral nerves. Thus, if the increase in NaV1.3 mRNA detected results in an increase in functional protein, these channels do not appear to contribute to AP propagation. Regardless, our results with ICA-121431 and phrixotoxin-3 constitute the first evidence of functional NaV1.1, NaV1.2, or NaV1.3 channels in a peripheral nerve, and support the suggestion that the VGSCs underlying AP propagation in trigeminal nerves are distinct from those underlying propagation in somatic nerves.
While our pharmacological data implicated an increase in functional NaV1.1 channels in the ION, this increase did not appear to be because of an increase in subunit mRNA, at least as assessed in the whole ganglia. The absence of significant changes in any of the VGSC subunits assessed in the TGs differ from those of previous studies in which downregulation of NaV1.7, NaV1.8, and NaV1.9 or upregulation of NaV1.3 mRNA have been reported (Liu et al., 2019, 2020) after CCI. This discrepancy could be because of differences in the time of evaluation (28 d vs 6 and 9 d). Nevertheless, that we were able to detect changes in NaV subunit expression following SN injury comparable to those previously reported, supports the suggestion that the increase in NaV1.1 protein detected in the ION is not because of a sustained increase in NaV1.1 mRNA.
In addition to the injury-induced increase in the relative efficacy of ICA-121431 in the ION but not SN, and the injury-induced increase in NaV1.1 protein in the ION but not SN, we obtained two additional lines of evidence in support of the conclusion that the upregulation of this subunit in trigeminal nerve fibers contributes to the therapeutic selectivity of CBZ. First, and most importantly, local administration of ICA-121431 attenuated the mechanical hypersensitivity observed following CCI of the ION but not the SN. These observations suggest that local block of ICA-121431-sensitive channels is sufficient to recapitulate the therapeutic effects of systemic CBZ. Second, pretreating the isolated ION with ICA-121431 attenuated the injury-induced increase in the potency of CBZ. This latter observation was somewhat equivocal, however, in that rather than simply eliminating the injury-induced increase in CBZ potency, resulting in a CBZ potency comparable to that observed in the absence of nerve injury (i.e., approximately >1 mm), the presence of ICA-121431 increased the potency of CBZ, even in the absence of nerve injury. This observation suggests that ICA-121431 may drive a conformational change in the VGSC α-subunit that facilitates CBZ binding to the channel. Thus, we suggest that the selective upregulation of NaV1.1 following CCI of the ION accounts, at least in part, for the therapeutic selectivity of CBZ.
While the changes described are compelling, and support the suggestion that it may be possible, if not necessary, to treat the pain associated with trigeminal nerve injury differently than that associated with somatic nerve injury, a number of interesting questions were raised by our results that will require further investigation. For example, in the absence of an increase in NaV1.1 mRNA (as least as assessed in the TGs), it will be important to determine the basis for the increase in NaV1.1 protein in the ION. In light of evidence in support of axonal translation of a number of proteins (Price and Géranton, 2009), as well as injury-induced changes in protein trafficking (Scheff and Gold, 2015) and protein stabilization (Iwata et al., 2004; Gomez et al., 2021), there are at least three viable mechanisms worth exploring. Another of these questions is the basis for the sex difference in the impact of nerve injury on the potency of CBZ, as noted above. In the context of the sex differences observed, it is important to point out that while NaV1.1 appears to be a target for CBZ, there was no evidence of a sex difference in results related to NaV1.1. That is, there was no sex difference in the relative efficacy of ICA-121431, in the impact of nerve injury on NaV1.1 protein level, or in the impact of local administration of ICA-121431 on nociception or on the interaction between ICA-121431 and CBZ in the isolated nerve. These observations suggest that an additional target contributes to the therapeutic selectivity of CBZ, and identification of that target may have important therapeutic implications.
While our results are consistent with our initial hypothesis and suggest that the therapeutic selectivity of CBZ during TN is related to a differential regulation of the VGSC, they suggest that additional mechanisms may contribute. Nevertheless, our results suggest that it may be possible to develop novel strategies for the treatment of TN by targeting the selective pattern of changes in the NaV1.1 subtype.
Footnotes
This work was supported by generous donations from trigeminal neuralgia patients and family members as well as by National Institutes of Health Grant R01-NS-064988 (M.S.G.), R01 NS122784 (M.S.G and R.F.S.), and P30-AG066468-02 (UPMC Neuropathology Core, Dr. Julia Kofler, PI), and the Bloch Foundation (M.S.G.). J.B.P.-F. received a Support for Postdoctoral Stays Abroad from Conacyt CVU 331856. We thank members of the Gold laboratory and the Pittsburgh Center for Pain Research for feedback in writing this manuscript; and Dr. Jamie Moy for the photograph of the human DRG. In addition, we thank the Center for Organ Recovery for tissue recovery. Finally, we thank the next of kin of tissue donors for their generous consent to the use of the donor's tissue for research purposes.
The authors have no competing financial interests.
- Correspondence should be addressed to Michael S. Gold at msg22{at}pitt.edu or Raymond F. Sekula Jr at sekularf{at}upmc.edu





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}