

Abstract
Drosophila multiple epidermal growth factor-like domains 8 (dMegf8) is a homolog of human MEGF8. MEGF8 encodes a multidomain transmembrane protein which is highly conserved across species. In humans, MEGF8 mutations cause a rare genetic disorder called Carpenter syndrome, which is frequently associated with abnormal left-right patterning, cardiac defects, and learning disabilities. MEGF8 is also associated with psychiatric disorders. Despite its clinical relevance, MEGF8 remains poorly characterized; and although it is highly conserved, studies on animal models of Megf8 are also very limited. The presence of intellectual disabilities in Carpenter syndrome patients and association of MEGF8 with psychiatric disorders indicate that mutations in MEGF8 cause underlying defects in synaptic structure and functions. In this study, we investigated the role of Drosophila dMegf8 in glutamatergic synapses of the larval neuromuscular junctions (NMJ) in both males and females. We show that dMegf8 localizes to NMJ synapses and is required for proper synaptic growth. dMegf8 mutant larvae and adults show severe motor coordination deficits. At the NMJ, dMegf8 mutants show altered localization of presynaptic and postsynaptic proteins, defects in synaptic ultrastructure, and neurotransmission. Interestingly, dMegf8 mutants have reduced levels of the Type II BMP receptor Wishful thinking (Wit). dMegf8 displays genetic interactions with neurexin-1 (dnrx) and wit, and in association with Dnrx and Wit plays an essential role in synapse organization. Our studies provide insights into human MEGF8 functions and potentially into mechanisms that may underlie intellectual disabilities observed in Carpenter syndrome as well as MEGF8-related synaptic structural and/or functional deficits in psychiatric disorders.
SIGNIFICANCE STATEMENT Carpenter syndrome, known for over a century now, is a genetic disorder linked to mutations in Multiple Epidermal Growth Factor-like Domains 8 (MEGF8) gene and associated with intellectual disabilities among other symptoms. MEGF8 is also associated with psychiatric disorders. Despite the high genetic conservation and clinical relevance, the functions of MEGF8 remain largely uncharacterized. Patients with intellectual disabilities and psychiatric diseases often have an underlying defect in synaptic structure and function. This work defines the role of the fly homolog of human MEGF8, dMegf8, in glutamatergic synapse growth, organization, and function and provide insights into potential functions of MEGF8 in human central synapses and synaptic mechanisms that may underlie psychiatric disorders and intellectual disabilities seen in Carpenter syndrome.
Introduction
Carpenter syndrome (CS), a rare genetic disorder with multiple congenital anomalies, was first described by George Carpenter (Carpenter, 1901). CS patients often present a variety of developmental defects at a very young age, including craniosynostosis, musculoskeletal abnormalities (mostly in fingers and toes), congenital heart disease, and intellectual disabilities (Taravath and Tonsgard, 1993; Hidestrand et al., 2009). Other clinical features, such as hypogenitalism, umbilical hernia, and obesity, have also been reported (Alessandri et al., 2010; Haye et al., 2014). Genome-wide sequence analyses identified three single nucleotide polymorphisms in the Multiple Epidermal Growth Factor-like Domains 8 (MEGF8) gene that showed association with the CS (Twigg et al., 2012). MEGF8 has also been implicated in neuropsychiatric disorders, such as schizophrenia (Bersani et al., 2003; Cox and Butler, 2015; Giacopuzzi et al., 2017).
Human MEGF8 is clinically relevant, and its homologs are highly conserved across species and encode a multidomain transmembrane protein (Fig. 1). Murine Megf8 is involved in bone morphogenetic protein (BMP) signaling and mutations in mice Megf8 disrupt axon guidance and lead to defects that phenotypically resemble human CS, including deformities of the limb, heart, and abnormal left-right patterning (Engelhard et al., 2013). Megf8, together with Mgrn1, catalyzes the ubiquitination and degradation of the Hedgehog pathway signaling molecules that coordinate cell–cell communication required for spinal cord and heart development (Jenkins et al., 2007; Pusapati et al., 2018; Kong et al., 2020). These findings suggest that Megf8 functions in multiple intracellular signaling pathways and may modulate different cellular processes during development. Despite these observations on the role of Megf8 in different developmental events and in neuronal function, it has remained unclear whether Megf8 has any role in synaptic development and function. Most importantly, intellectual disabilities seen in CS patients as well as the association of human MEGF8 with psychiatric disorders, such as schizophrenia, point toward a potential role of MEGF8 in synaptic processes as many of the learning disabilities and psychiatric disorders have underlying defects in synaptic development and/or function (Valnegri et al., 2012; Giacopuzzi et al., 2017; Obi-Nagata et al., 2019).
The Drosophila neuromuscular junctions (NMJs) have served as an ideal model for studying synapse development and functions (Wu et al., 2010), and have similarities with the mammalian central synapses. Since dMegf8 shows high amino acid sequence and domain homology with vertebrate Megf8 (Lloyd et al., 2018), studies on dMegf8 will be informative about the role of vertebrate Megf8 proteins. While not much is known about dMegf8, several genome-wide RNAi-screens found that dMegf8 knockdown resulted in cell death or reduced cell viability phenotype (Chittaranjan et al., 2009; Mummery-Widmer et al., 2009). In a machine-learning study, dMegf8 was found to be one of the putative synaptic genes indicating a potential involvement of dMegf8 in synapse assembly and function (Pazos Obregon et al., 2015). A recent report showed that dMegf8 mutants have disrupted larval denticle belts and bristle formation, and a delayed transition to third instar larvae associated with larval lethality (Lloyd et al., 2018). There are no published reports on any role of Megf8 in synapse organization or function in any species.
Here we report the generation of dMegf8 null mutants and show that dMegf8 is expressed both presynaptic and postsynaptically. dMegf8 is required for synaptic growth, and loss of dMegf8 leads to synaptic ultrastructural defects and reduced synaptic transmission. Additionally, genetic rescue experiments support that dMegf8 is necessary presynaptically for proper NMJ growth. However, synaptic ultrastructural defects resulting from dMegf8 loss are significantly rescued by dMegf8 presynaptic and/or postsynaptic expression. Importantly, dMegf8 shows genetic interactions with Drosophila neurexin (dnrx) and the BMP receptor wishful thinking (wit), and these proteins exist in a biochemical complex to coordinate proper NMJ development. Together, our studies establish the role of dMegf8 in NMJ synaptic development and function, and provide key insights into potential Megf8 functions in vertebrate synaptic mechanisms that underlie intellectual disabilities seen in CS patients.
Materials and Methods
Fly stocks
The Drosophila lines used in this study include w1118 Canton-S line (used as WT control; kind gift from Vivian Budnik), dMegf8Δ8 (Lloyd et al., 2018), dnrx273, UAS-dnrx (Li et al., 2007), and UAS-wit-GFP (a gift from Michael O'Connor). To generate UAS-dMegf8 flies, we obtained a partial dMegf8 cDNA clone from DGRC (clone #LD09511) that encodes a polypeptide containing only the C-terminal amino acids 2089-2892 of the full-length dMegf8RB protein. The remaining 2088 N-terminal amino acids were obtained by RT-PCR using Poly A+ mRNA isolated from adult fly brains. A full-length clone was assembled containing 10,201 nucleotides representing the largest dMegf8 isoform (dMegf8 RB), which was confirmed by sequencing and cloned into pUAS-attB vector to create transgenic flies. All other fly stocks, including Df(2L)7147, witA12, and witB11, and all Gal4 lines were obtained from the Bloomington Drosophila Stock Center. All flies were maintained at 25°C, 50% humidity and with a 12 h light/dark cycle.
Generation of dMegf8 mutants
To generate a targeted deletion in the dMegf8 locus that would produce a dMegf8 null allele, we used CRISPR/Cas9 methodology and procedures established at the GenetiVision company based on the previously reported strategies (Zhang et al., 2014). The targeting construct contained 5′ and 3′ homology arms flanking a GFP expression cassette that replaced the deleted sequences in the dMegf8 locus. The targeting construct was injected into embryos, and the transgenic flies were tested by PCR analysis using a combination of primers that distinguished the WT and dMegf8 null alleles (primer 1-5′-GCACGCTTCAGGTAAGTCGTA-3′, primer 2-5′-GCGGCTTGTGATCCGTAACCT-3′ and primer 3-5′-GATGGGACAAGTCGCCATGT T-3'). Further characterization of the dMegf8 alleles used standard methodologies.
Larval locomotion assays
The larval locomotion assay was performed as described previously (Banerjee and Riordan, 2018). Briefly, larvae were first washed in distilled water to remove any traces of food before performing various assays. Each larva was acclimatized to the test plate for 1 min before testing. Five trials per larvae were conducted, and the total number of larvae analyzed per genotype was 50.
For the grid crossing assay, individual larvae were placed in the center of a 145-mm-diameter Petri dish, with 2% non-nutritious agar previously poured and allowed to harden covering a graph paper at the bottom with 0.5 cm2 marked grids. The number of grid line crossings within a 30 s time window was recorded 5 times per larva.
For the central zone release assay, a circular white card 1.5 cm inch diameter was taped to the bottom of the dish to mark the central release zone. Five animals were placed at the center of the release zone. The time taken for each animal to exit the release zone was recorded.
For the peristalsis contraction assay, full body peristalsis contractions (full posterior to anterior movement = 1 contraction) were counted for each larva in 1 min while observing under a dissection microscope.
For the righting assay, larvae were turned on their dorsal surfaces with a fine brush and the time taken to return to their ventral crawling position was recorded.
Adult locomotion assay
To determine the locomotor ability of the flies, adult climbing assay was performed as described previously (Gargano et al., 2005; Xie et al., 2021). Briefly, 10 freshly eclosed male flies were collected in individual vials, and a total of 50 flies for each genotype were analyzed. The assay was started 24 h after CO2 anesthesia. Flies were gently taped down to the bottom of an empty clear vial, and the number of flies crossing a 10 cm mark drawn from the base of the vial within 10 s were recorded. Each assay was repeated 6 times with a recovery time of 1 min in between, and the mean was calculated. The climbing ability is shown as the percentage of mean number of flies crossing the mark.
Production of dMegf8 antibodies
To generate antibodies against the dMegf8 protein, a partial cDNA clone of dMegf8 that contained two-thirds of the coding sequences of dMegf8 from the 3′-end was obtained from the DGRC Center and further confirmed by DNA sequencing. A portion of the dMegf8 was subcloned (cDNA nucleotide numbers 6706-7323) into pET28 a(+) vector and expressed in Escherichia coli BL21DE3 followed by His-column affinity purification. The recombinant polypeptide was used as an antigen to generate rabbit and guinea pig polyclonal antibodies, which were further tested by immunostaining and immunoblotting methods.
Immunohistochemistry
Wandering third-instar larvae from various genotypes were dissected and fixed in either Bouin's fixative or 4% PFA for 15 min and processed as previously described (Chen et al., 2012; Banerjee et al., 2017). Dnrx signal at NMJ was enhanced using previously described protocols (Li et al., 2007). Primary antibodies used were rabbit anti-dMegf8 (1:500, this study), FITC-conjugated anti-Hrp (1:250, Jackson ImmunoResearch Laboratories), mouse anti-GluRIIA (1:250) (Marrus et al., 2004; Chen et al., 2012), guinea pig anti-Dnrx (1:250) (Li et al., 2007), rabbit anti-PS1 (p-Mad) (1:500; a gift from P. ten Dijke), and rabbit anti-Smad (1:200, ab52903, Abcam). Anti-PS1 recognizes pMad at the NMJ and anti-Smad antibody, which is a recombinant anti-Smad3 [phospho S423 + S425], recognizes pMad in the ventral nerve cord (VNC) as well as detects pMad on immunoblots (Banerjee et al., 2017; Banerjee and Riordan, 2018). Mouse monoclonal anti-Dlg (1:500, 4F3), anti-BRP (1:250; NC82), and anti-Wit (1:25, 23C7) were obtained from Developmental Studies Hybridoma Bank (DSHB), University of Iowa. Secondary antibodies conjugated to Alexa-488, -568, and -647 (Invitrogen) were used at 1:400 dilution. Confocal images of all dissected larval tissues belonging to the same experimental group were imaged under identical settings with a Carl Zeiss LSM710 confocal microscope, and all image processing was done using Adobe Photoshop software.
Immunoblotting and immunoprecipitations (IPs)
The immunoblotting and IP experiments were performed as previously described (Banerjee et al., 2017). Briefly, for immunoblotting of dMegf8, fly heads of desired genotypes were homogenized using a glass homogenizer in a weight/volume ratio of 1:3 in ice-cold lysis buffer containing 50 mm HEPES, pH 7.5, 100 mm NaCl, 1 mm MgCl2, 1 mm CaCl2, 1% NP-40, and 0.5% deoxycholate with protease inhibitors. The lysates were kept on ice for 10 min and centrifuged at 20,000 × g for 15 min at 4°C. The resultant supernatant was centrifuged at 100,000 × g for 30 min at 4°C. The membrane pellet was further solubilized in 1 × PBS and used subsequently for IP and immunoblot analyses. For the immunoblotting of third instar larval musculature or VNC and brain lobes without any attached imaginal discs, tissues were homogenized in ice-cold RIPA buffer. The supernatants with equal amounts of proteins from each genotype were separated on SDS-PAGE followed by immunoblotting with appropriate antibodies. For IP studies, fly heads of the appropriate genotypes were processed according to previously described protocols (Banerjee et al., 2017). Each experiment was performed independently 3 times, and the most representative blots are presented. Primary antibodies used for immunoblotting were anti-dMegf8 (1:1000), anti-Dnrx (1:250) (Li et al., 2007), anti-Wit (1:1000, DSHB), anti-Smad (1:150, Abcam), anti-Trio (1:250, DSHB), and anti-β actin (1:10,000, 4967S, Cell Signaling).
Electron microscopy and morphometric analysis
Ultrastructural analyses of third-instar larval NMJs were processed for TEM as previously described (Banerjee et al., 2017). Briefly, third-instar larval fillets were dissected in ice-cold, Jan's 0.1 mm Ca2+ saline, pH 7.o2, and subsequently fixed in 4% PFA/1% glutaraldehyde/0.1 m cacodylic acid, pH 7.2, for 30 min at room temperature followed by overnight fixation at 4°C. The fixed fillets were rinsed 3× in 0.1 m cacodylic acid, pH 7.2, and postfixed in 2% aqueous osmium tetroxide for 1 h, followed by rinsing and dehydration in increasing ethanol concentrations. Samples were incubated for 1 h in propylene oxide and gradually infiltrated in increasing resin to propylene oxide ratio (1:2, overnight; 2:1, at least 6 h; and full resin for 36 h with constant agitation). Samples were embedded in flat silicone molds with Polybed resin and cured in the oven at 55°C for at least 36 h. Five larvae were processed for EM analysis from each of the genotypes. The number of boutons (n) analyzed for each genotype is indicated in respective figure legends. ImageJ was used for morphometric analysis of EM images of only Type Ib boutons from A2 and A3 as previously described (Chen et al., 2012; Banerjee et al., 2017).
Electrophysiology
Electrophysiological analysis of larval NMJ was performed as previously described (Shi et al., 2019). All third instar larvae were grown in an incubator at 25°C (65% humidity). All electrophysiological experiments were performed at room temperature, and all recordings were made only from abdominal segment A3, muscle 6 of third-instar larvae, in HL-3 solution (0.5 mm Ca2+ and 20 mm MgCl2) (Stewart et al., 1994). Electrophysiological signals were amplified with an Axoclamp 900A, under the control of Clampex 10 (Molecular Devices). Data were collected only when resting potential was <–63 mV. Excitatory junction potentials (EJPs) were evoked by applying currents of 6 ± 3 mA with fixed stimulus duration at 0.3 ms with 0.2 Hz of stimuli rate. Twenty evoked EJPs were recorded for each muscle for analysis. Miniature EJP (mEJP) events were collected for 2 min. The evoked EJP amplitude was corrected by using nonlinear summation (Feeney et al., 1998). The quantal content was calculated from individual muscles by ratio of the averaged EJP and averaged mEJP amplitudes. Statistical analyses of EJP and mEJPs between genotypes were done using Student's t test (SigmaPlot 10.0, Systat Software). Error bars indicate mean ± SEM.
Quantification
Bouton number quantifications (n = number of larvae analyzed) were performed from muscles 6/7 of abdominal segment 3 (A3) by staining of the body wall musculature preparations with anti-Hrp and anti-Dlg. Fluorescence intensity measurements for dMegf8, Dnrx, Wit, and pMad were quantified from confocal slices of Z stack images compressed using maximum projection functions, which were stained in combination with either NC82 or anti-Hrp antibodies. Same ROIs were selected for each channel and used for assessment and quantification of fluorescence intensity using ImageJ. At least 30 NMJ branches from 8 larvae were analyzed for various genotypes. All genotypes listed under the same quantification groups were stained and processed for imaging and quantified under identical parameters and settings. ImageJ (National Institutes of Health) was used for quantification of band intensities of immunoblots from three independent experiments. The intensity of the bands of interest was divided by their respective actin protein blots to control for any possible unequal loading.
Experimental design and statistical analysis
All experiments described above, including microscopy, image processing, and quantification, were performed by trained researchers. All experiments used both sexes, except for adult locomotion assay, in which only male flies were used to exclude the influence of female's oviposition. Larvae and adult flies from the control and experimental groups were reared in the same media and maintained at the same temperature (25°C) and humidity (50%) and were processed identically.
All statistical analyses were performed using the GraphPad Prism software, and data are presented as mean ± SEM. Statistical significance was determined by one way ANOVA followed by post hoc Tukey's multiple comparison test and Student's t test. Specifically, the adult locomotion assay used two-way ANOVA as there were two independent variables (age and climbing index, Fig. 1J). Error bars indicate mean ± SEM. Wherever possible, exact p values are provided in the figure legends. For Figure 1F, we analyzed the data using Welch's Student's t test, assuming unequal variances and unequal sample sizes. For Figure 4G, instead of using normal one-way ANOVA, we used Brown-Forsythe and Welch ANOVA, which takes care of unequal sample size issues. Additional information about each statistical test, including degrees of freedom and other statistic-specific values, are included in the figure legends. For all quantifications, the statistical significance immediately above the bars is with respect to the control genotype for that experimental group.
Results
Generation of dMegf8 null mutants
The primary amino acid sequence of human MEGF8 protein revealed it as a transmembrane protein with a complex domain structure in its extracellular region, including CUB domains, multiple EGF repeats, KELCH domains, and laminin-EGF-like domains (Twigg et al., 2012). The mouse Megf8 protein also revealed a conserved domain structure similar to the human MEGF8 (Engelhard et al., 2013). The Drosophila Megf8, referred as dMegf8 (Lloyd et al., 2018), also revealed a complex domain structure with similar domains as the vertebrate Megf8 with some variations, but is the closest homolog of the vertebrate Megf8 family (Fig. 1A). dMegf8 shares >2400 amino acid sequence homology with human MEGF8 and close to 33% amino acid identity (Lloyd et al., 2018). In our studies, the dMegf8 locus was identified in an ethyl methane sulfonate chemical mutagenesis screen that was designed to uncover behavioral mutants with motor coordination deficits (A.V. and M.A.B., unpublished data). To explore the in vivo functions of dMegf8, and whether it has a role in neuronal and synaptic functions, we proceeded to generate a dMegf8 null allele. The dMegf8 locus (Fig. 1B) is composed of 6 exons with the ATG codon in exon 1 and the termination codon in exon 6. We used the CRISPR-Cas9 gene editing strategy (Zhang et al., 2014) and inserted a GFP cassette to replace ∼4062 bp starting from nucleotide sequence 5′-AAGCTATGGGGTCTG-3′ in exon 3 and ending in nucleotide sequence 5′-GATCGAATGCCTCTT-3′ in exon 4 (for details, see Fig. 1B). We conducted PCR amplification using fly genomic DNA with a primer combination that distinguished the WT allele (primers 1 + 2 = 407 bp) (Fig. 1B,C) from the dMegf8 mutant allele (primers 1 + 3 = 505 bp) (Fig. 1B,D). The PCR amplification confirmed the deletion of the sequences, and this dMegf8 allele is referred as dMegf8HSC. Next, we generated polyclonal antibodies against dMegf8 polypeptide to determine the relative molecular weight of dMegf8 and also to establish that dMegf8HSC is indeed a protein null allele. We also generated UAS-dMegf8 transgenic flies to express dMegf8 using cell-specific Gal4 drivers. We performed immunoblot analysis of adult head lysates from WT (+/+), dMegf8HSC and elav-Gal4;UAS-dMegf8 (elav>dMegf8) using anti-dMegf8 antibodies. As shown in Figure 1E, WT and elav>dMegf8 lysates showed three major protein bands ranging from 250 to >300 kDa, which were absent from dMegf8HSC head lysates, confirming the specificity of the dMegf8 antibodies and dMegf8HSC as a protein null allele. Using pan-neuronal expression of dMefg8 (elav>dMegf8), the dMegf8 protein is expressed at higher levels than in the WT control adult heads, indicating overexpression from the dMegf8 transgene. The dMegf8HSC null allele reported here showed no significant embryonic lethality and only had <10% larva to adult lethality. We next assayed third instar larvae for various larval behavioral and coordination defects that included general motor activity using grid crossing assay (Fig. 1F), central zone release assay (Fig. 1G), peristalsis assay which analyses the muscle contraction waves (Fig. 1H), and larval righting assay (Fig. 1I). In all these assays, dMegf8HSC larvae showed significant deficits compared with WT control larvae. We also conducted adult locomotor assay that uses negative geotaxis (Gargano et al., 2005), as part of motor coordination and climbing activity. dMegf8HSC adult flies showed a significant weakness in their climbing activity, which declined progressively as flies aged (Fig. 1J), indicating that loss of dMegf8 leads to severe motor deficits in third instar larvae as well as adults. Another dMegf8 null allele (dMegf8Δ8) was reported previously, which showed larval lethality with denticle belt formation defects (Lloyd et al., 2018). It is not known whether variable genetic background contributes to early developmental lethality or whether there are any second site mutations associated with the dMegf8Δ8 allele (Lloyd et al., 2018). Together, these data show that dMegf8HSC is indeed a null allele that is viable and that loss of dMegf8 is associated with severe motor deficits in both the larvae and adult flies.

Generation of dMegf8 mutants. A, Protein domain structure of human MEGF8, mouse Megf8, and Drosophila dMegf8. Green bar represents the antibody region in dMegf8. B, Genomic structure of dMegf8 showing exons 1-5. The targeting construct using CRISPR/Cas9 for recombination and the final targeted allele of dMegf8HSC is shown. Blue boxes next to loxP sites on both sides represent phage C31 integration sites referred to as attP sites in the targeting vector. Red arrows with numbers indicate location of the primers for genotyping that differentiate the WT and the dMegf8HSC mutants. C, D, PCR confirmation of the targeted deletion using primer combinations 1 + 2 for WT and 1 + 3 for dMegf8HSC mutants in B. E, Immunoblot analysis of dMegf8 showing presence of dMegf8 in WT, loss of dMeg8 in dMegf8HSC mutants, and overexpression of dMegf8 in elav>dMegf8. The blot was probed for actin as loading control. F–I, WT (+/+) and dMegf8HSC larval locomotor behaviors assayed by measuring the number of 0.5 cm2 grids crossed in 30 s (F), time taken in seconds for larvae to exit a circle of 1.5 cm in diameter (G), number of full body peristaltic contractions in 1 min (H), and time taken in seconds for larvae to right themselves when turned on their dorsal surface (I). n = 50 larvae. Data are mean ± SEM. t(46.49) = 18.22, ****p < 0.0001 (Welch's Student's t test) (F), t(78) = 4.833, ****p < 0.0001 (unpaired Student's t test) (G), t(78) = 5.753, ****p < 0.0001 (unpaired Student's t test) (H), and t(78) = 6.863, ****p < 0.0001 (unpaired Student's t test) (I). J, Adult locomotion assay using climbing ability of WT (+/+) (black) and dMegf8HSC mutants (red) flies. n = 50 flies. Data are mean ± SEM. Interaction: F(9,80) = 4.579, ****p < 0.0001 (two-way ANOVA test).
dMegf8 localizes to synaptic terminals and is required for synaptic growth
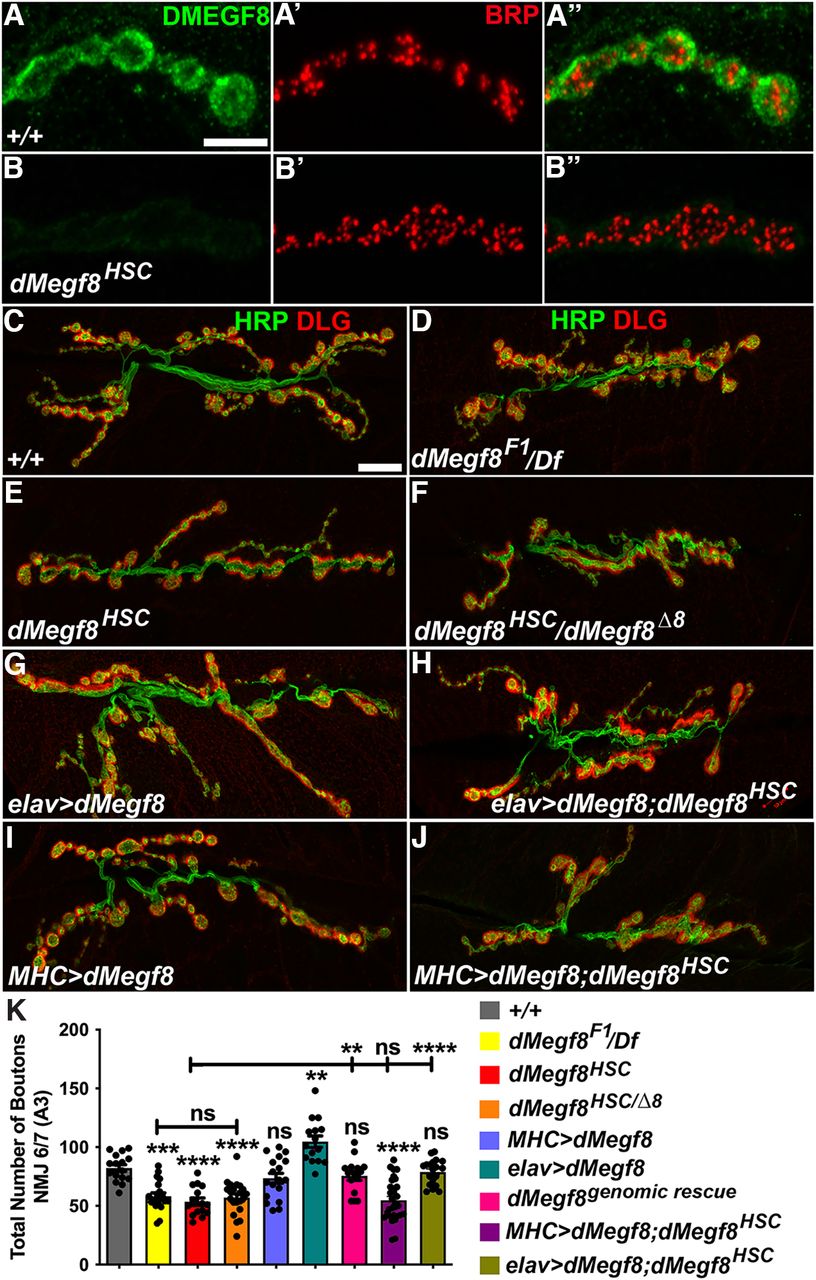
Since mutations in human MEGF8 have been associated with developmental malformations (Twigg et al., 2012; Pan et al., 2014; Giacopuzzi et al., 2017), and the murine Megf8 null allele showed developmental malformations and embryonic lethality (Engelhard et al., 2013), we wanted to examine where dMegf8 was expressed and whether loss of dMegf8 had any consequences on synaptic development at the NMJ, as these synapses are generally considered to have a functional relevance to mammalian glutamatergic central synapses (Ruiz-Canada and Budnik, 2006; Xue et al., 2009). We first examined the endogenous expression of dMegf8 at the third instar larval NMJs by performing immunostaining for dMegf8 in combination with antibodies against the presynaptic active zone protein, Bruchpilot (Brp). We found that dMegf8 localized to the larval NMJ synapses in WT (Fig. 2A-A′′) when labeled with anti-dMegf8 (Fig. 2A,A′′) with respect to Brp (Fig. 2A′,A′′) (Wagh et al., 2006; Weyhersmuller et al., 2011). dMegf8 was not detected in the larval NMJ synapses in dMegf8HSC mutants (Fig. 2B,B′′), which were clearly highlighted by Brp immunostaining (Fig. 2B′,B′′), indicating that dMegf8 localizes to larval NMJ synapses. We next investigated the consequences of dMegf8 loss on the NMJ synaptic growth. We immunostained the larval NMJ of specified genotypes, including several alleles of dMegf8 (dMegf8F1 -from the ethyl methane sulfonate screen, dMegf8HSC, and dMegf8Δ8; Fig. 2C–K) (Lloyd et al., 2018) using anti-Hrp (green, Fig. 2C–J) to label neuronal membranes and anti-Discs large (Dlg) (red, Fig. 2C–J) to label Type I boutons (Lahey et al., 1994; Budnik et al., 1996). Compared with WT (Fig. 2C, quantified in Fig. 2K), all dMegf8 mutant alleles showed synaptic undergrowth with fewer boutons (Fig. 2D–F,K). dMegf8F1 also showed reduced synaptic growth comparable to dMegf8HSC (Fig. 2D,K). To ensure the dMegf8HSC homozygous phenotype was not caused by any other mutations in cis, we analyzed the dMegf8HSC allele over a deficiency chromosome (Df(2L)7147) that uncovered the dMegf8 locus. Mutant larvae of dMegf8HSC/Df(2L)7147 revealed NMJ growth phenotype that was not significantly different from dMegf8HSC/dMegf8HSC homozygous mutants (data not shown), indicating that dMegf8HSC allele can be used for NMJ phenotypes in a homozygous state. While the dMegf8HSC homozygous null flies were viable and fertile, an independently generated dMegf8 allele, also thought to be null, was reported to be larval lethal (Lloyd et al., 2018). We used one of the reported dMegf8 mutant alleles, dMegf8Δ8 (Lloyd et al., 2018), to analyze the synaptic growth in trans-allelic combination with dMegf8HSC. dMegf8HSC/dMegf8Δ8 larvae showed similar synaptic undergrowth as the dMegf8HSC homozygous null mutants (Fig. 2F,K). Since dMegf8Δ8 allele was also generated using CRISPR/Cas9 deletion, it remains to be determined whether there is a second site mutation associated with this allele. Together, these data show that dMegf8 is expressed at the larval NMJ and that loss of dMegf8 causes a significant reduction in bouton growth at the NMJ.

dMegf8 is expressed in synaptic terminals and required for synaptic bouton growth. A–B′′, Confocal images in (A) +/+ and (B) dMegf8HSC mutant third instar larvae NMJ Type Ib boutons at muscles 6/7 labeled with anti-dMegf8 (green) and anti-Brp (red). C–J, Confocal images in (C) dMegf8HSC, (D) dMegf8 point mutant dMegf8F1/Df, (E) dMegf8 null mutant (dMegf8HSC), (F) dMegf8HSC/dMegf8Δ8 trans-allelic combination, (G) presynaptic overexpression of dMegf8 (elav>dMegf8), (H) presynaptic rescue (elav>dMegf8;dMegf8HSC), (I) postsynaptic overexpression of dMegf8 (MHC>dMegf8), and (J) postsynaptic rescue (MHC>dMegf8;dMegf8HSC) third instar larvae NMJ at muscles 6/7 labeled with the presynaptic marker Hrp (green) and the postsynaptic marker Dlg (red). K, Quantification of total bouton numbers in indicated genotypes. K, Data are mean ± SEM (one-way ANOVA test with Tukey's multiple comparisons): F(8,160) = 22.3064, +/+ versus F1/Df: ***p = 0.0001, +/+ versus dMegf8HSC: ****p < 0.0001, +/+ versus dMegf8HSC/Δ8: ****p < 0.0001, +/+ versus MHC>dMegf8: p = 0.7496, +/+ versus elav>dMegf8: **p = 0.0014, +/+ versus Genomic Rescue: p = 0.9467, +/+ versus MHC>dMegf8; dMegf8HSC: ****p < 0.0001, +/+ versus elav>dMegf8; dMegf8HSC: p = 0.9992, dMegf8HSC versus F1/Df: p = 0.9915, dMegf8HSC versus dMegf8HSC/Δ8: p = 0.9989, dMegf8HSC versus genomic Rescue: **p = 0.0027, dMegf8HSC versus MHC>dMegf8; dMegf8HSC: p > 0.9999, dMegf8HSC versus elav>dMegf8; dMegf8HSC: ****p < 0.0001. Scale bars: A–B′′, 5 μm; C–J, 20 μm.
On the contrary, presynaptic expression of dMegf8 in elav>dMegf8 larvae led to increased growth of synaptic boutons (Fig. 2G,K), suggesting that dMegf8 overexpression caused enhanced bouton growth at the NMJ. Next, we wanted to assess whether the reduced synaptic growth seen in dMegf8HSC mutants could be rescued by presynaptic or postsynaptic expression of dMegf8. We found that presynaptic expression of dMegf8 in dMegf8HSC mutants (dMegf8HSC/HSC;elav-Gal4/UAS-dMegf8) restored synaptic growth to WT levels (Fig. 2H,K); however, the postsynaptic expression of dMegf8 using MHC-Gal4 (MHC>dMegf8) (Fig. 2I,K) did not show any changes in synaptic bouton growth compared with WT (Fig. 2C,K). The postsynaptic expression of dMegf8 in dMegf8HSC mutants (dMegf8HSC/HSC; MHC-Gal4>dMegf8) failed to rescue the reduced bouton growth in dMegf8HSC mutants (Fig. 2I,K). These data show that presynaptic dMegf8 expression is able to promote synaptic growth at NMJ and that the synaptic growth function of dMegf8 is mostly presynaptic as the postsynaptic expression neither promoted nor rescued the synaptic growth at the NMJ.
Loss of dMegf8 affects the distribution of presynaptic and postsynaptic proteins
Since NMJ bouton growth was affected in dMegf8 mutants, we wanted to determine whether loss of dMegf8 would have any consequences on the proper assembly of presynaptic and postsynaptic proteins which display a stereotypic localization at the NMJ synapses with respect to assembly and distribution. We first studied the localization of the well-characterized active zone protein, Brp (Wagh et al., 2006) (green, Fig. 3A, A′′; Fig. 3B,B′′) with respect to the postsynaptic glutamate receptor subunit, GluRIIA (Marrus et al., 2004) (red, Fig. 3A′,A′′; Fig. 3B′,B′′) in WT (Fig. 3A-A′′) and dMegf8HSC mutant (Fig. 3B′,B′′) larval NMJ. In WT larvae (Fig. 3A–A′′), active zones labeled with anti-Brp (Fig. 3A,A′′) were juxtaposed to the GluRIIA punctae (Fig. 3A′,A′′), which was similar in dMegf8HSC mutant synapses (Fig. 3B-B′′), indicating that the alignment of the presynaptic BRP and postsynaptic GluRIIA was not significantly affected. Next, we analyzed the number of Brp puncta in the WT (Fig. 3A-A′′) and dMegf8HSC mutants (Fig. 3B-B′′) to determine whether there were any differences in the number of active zones per bouton area. dMegf8HSC mutants showed a significant increase in the number of Brp-positive puncta as quantified and normalized to the bouton area in dMegf8HSC mutants compared with WT (Fig. 3C). GluRIIA puncta also showed similar increase as the Brp puncta in dMegf8 mutants compared with the WT (data not shown). These data indicate that there is increase in the number of Brp-positive active zones in dMegf8HSC mutant NMJ synapses.

dMegf8HSC mutants show altered localization of presynaptic/postsynaptic proteins. A–B′′, Confocal images in (A) +/+ and (B) dMegf8HSC mutant third instar larval NMJ Type Ib boutons at muscles 6/7 labeled with the presynaptic protein Brp (green) and the postsynaptic protein GluRIIA (red). C, Quantification of Brp puncta/bouton area (μm2) in WT and dMegf8HSC mutants. D–E′, Confocal images in (D) +/+ and (E) dMegf8HSC mutant third instar larvae NMJ labeled with the presynaptic marker Hrp (green) and the postsynaptic protein Dlg (red). C, Quantification data are mean ± SEM; t(62) = 11.14. ****p < 0.0001 (unpaired Student's t test). F–G′, Dlg localization in dMegf8HSC mutants that have either presynaptic expression (elav>dMegf8) (F,F′) or postsynaptic expression (MHC>dMegf8) (G–G′). Scale bars: A–B′, F, G, 5 μm; D–E′, F′, G′, 20 μm.
Next, we determined whether dMegf8HSC mutants caused any aberrations in the organization or differentiation of postsynaptic specializations. We studied the localization of Discs large (Dlg, red, Fig. 3D–E′), which has been shown to function both presynaptically and postsynaptically in the proper assembly of the subsynaptic reticulum (SSR) in Type Ib boutons (Lahey et al., 1994; Budnik et al., 1996) together with the neuronal membrane marker, Hrp (green, Fig. 3D′,E′). In WT (Fig. 3D,D′), Dlg localization is typically in the periphery of the bouton circumference and mostly excluded from the core of the boutons. However, in dMegf8HSC mutants (Fig. 3E,E′), there was a more diffuse Dlg distribution throughout the bouton; and there was no clear separation in the presynaptic Hrp areas, as is observed in the WT boutons (Fig. 3D,D′), suggesting that loss of dMegf8 affected the localization of Dlg at the NMJ. Next, we tested whether the mislocalization of Dlg in the dMegf8HSC mutant boutons would be rescued by presynaptic or postsynaptic expression of dMegf8. As shown in Figure 3F and at a higher magnification in Figure 3F′, elav>dMegf8;dMegf8HSC NMJs showed improved Dlg localization that was more in the bouton perimeter similar to the WT boutons (Fig. 3D) than that displayed by MHC>dMegf8;dMegf8HSC NMJs (Fig. 3G, higher magnification in Fig. 3G′), indicating that presynaptic expression of dMegf8 restored Dlg localization better than the postsynaptic dMegf8 expression. Together, these data show that dMegf8 is required for proper presynaptic and postsynaptic protein assembly and distribution at the NMJs, and that presynaptic expression was able to restore Dlg localization at the NMJ compared with dMegf8HSC mutants.
Presynaptic and postsynaptic ultrastructural abnormalities in dMegf8 mutants
The reduced synaptic growth and altered subcellular localization of Dlg observed in dMegf8 mutants led us to analyze the ultrastructure of the synaptic boutons in these mutants to examine the organization of the overall synaptic architecture and how that compared with WT controls. The WT boutons are characterized by morphologically distinct and closely apposed presynaptic and postsynaptic membranes (Fig. 4A). The presynaptic compartment also contains synaptic vesicles, apart from organelles, such as mitochondria. The presynaptic membrane has active zones that are composed of electron dense structures called T-bars (Fig. 4A) (Wagh et al., 2006). The Type Ib boutons at the postsynaptic muscle are surrounded by elaborate membrane invaginations, the SSR, which occupies a large area on the postsynaptic side (Fig. 4A,B) (Budnik et al., 1996; Jia et al., 1993). We performed serial sectioning of boutons from WT, dMegf8F1, and dMegf8HSC mutants and subjected them to morphometric analyses to determine any presynaptic and/or postsynaptic defects. In addition, we also assessed whether any of the synaptic defects displayed by dMegf8HSC mutants would be rescued by expressing dMegf8 either presynaptically or postsynaptically.

Loss of dMegf8 causes synaptic ultrastructural defects. A–E′, TEM images of cross sections through Type Ib boutons in (A) +/+, (B) dMegf8HSC mutant, (C) dMegf8 point mutant dMegf8F1/Df, (D) presynaptic rescue (elav>dMegf8;dMegf8HSC), and (E) postsynaptic rescue (MHC>dMegf8;dMegf8HSC) at low magnification (A–E) and high magnification (A′–E′). Arrows indicate the active zones (AZs). Arrowheads indicate the PSDs in A–E. F–I, Quantification in (F) total bouton area, (G) number of AZs, (H) total PSD length/perimeter (%), and (I) normalized SSR width in represented genotypes. F–I, Data are mean ± SEM (one-way ANOVA test with Tukey's multiple comparisons). F, F(4,183) = 0.4529, +/+ versus dMegf8HSC: p = 0.7990, +/+ versus F1/Df: p > 0.9999, +/+ versus elav>dMegf8; dMegf8HSC: p = 0.9994, +/+ versus MHC>dMegf8; dMegf8HSC: p > 0.9999. G, F(4.000,159.4) = 25.97, +/+ versus dMegf8HSC: ****p < 0.0001, +/+ versus F1/Df: ****p < 0.0001, +/+ versus elav>dMegf8; dMegf8HSC: p = 0.9999, +/+ versus MHC>dMegf8; dMegf8HSC: ****p < 0.0001 (Brown-Forsythe and Welch ANOVA with Games-Howell's multiple comparisons specifically). H, F(4,179) = 18.18, +/+ versus dMegf8HSC: ****p < 0.0001, +/+ versus F1/Df: *p = 0.0292, +/+ versus elav>dMegf8; dMegf8HSC: p > 0.9999, +/+ versus MHC>dMegf8; dMegf8HSC: ****p < 0.0001. I, F(4,169) = 16.03, +/+ versus dMegf8HSC: ****p < 0.0001, +/+ versus F1/Df: *p = 0.0136, +/+ versus elav>dMegf8; dMegf8HSC: ****p < 0.0001, +/+ versus MHC>dMegf8; dMegf8HSC: p = 0.1136, dMegf8HSC versus elav>dMegf8; dMegf8HSC: p = 0.2102, dMegf8HSC versus MHC>dMegf8; dMegf8HSC: ****p < 0.0001. Scale bars: A–E, 600 nm; A′–E′, 200 nm.
We did not observe any significant changes in the overall area of the boutons in dMegf8HSC (Fig. 4B, quantified in Fig. 4F) and dMegf8F1 mutants (Fig. 4C,F) compared with WT (Fig. 4A,F) or any of the other genotypes analyzed (Fig. 4D–F). It is important to note that, although many dMegf8HSC mutant boutons had larger bouton areas, there was also a significant variability. The number of active zones (arrows) and total PSD length (arrowheads) showed a significant increase in dMegf8HSC (Fig. 4B, quantified in Fig. 4G and Fig. 4H, respectively) and dMegf8F1 mutants (Fig. 4C,G,H) compared with WT controls (Fig. 4A,G,H). Both increased active zone numbers and total PSD length in dMegf8HSC mutants were rescued to WT levels in the presynaptic rescue as seen in elav>dMegf8;dMegf8HSC (Fig. 4D,G,H), while the postsynaptic expression of dMegf8 in dMegf8HSC mutants, as seen in MHC>dMegf8;dMegf8HSC (Fig. 4E,G,H), did not show any changes in these parameters compared with dMegf8HSC mutants (Fig. 4B,G,H). Postsynaptic SSR morphology was severely compromised in dMegf8HSC (Fig. 4B,B′) and dMegf8F1 (Fig. 4C,C′) with thinner folds compared with WT controls (Fig. 4A,A′). Morphometric analysis of normalized SSR width (Fig. 4I) showed a significant reduction in dMegf8HSC mutants compared with WT controls (Fig. 4I). dMegf8F1 mutants also showed reduced normalized SSR width (Fig. 4I). Interestingly, reduction in the normalized SSR width was fully rescued when dMegf8 was expressed postsynaptically in dMegf8HSC mutants as seen in MHC>dMegf8;dMegf8HSC (Fig. 4I) but not by the presynaptic expression of dMefg8 as observed in elav>dMegf8; dMegf8 HSC (Fig. 4I). Together, the EM and morphometric analyses indicate that loss of dMegf8 leads to defective ultrastructural organization of both the presynaptic and postsynaptic areas at the NMJ and that dMegf8 functions both presynaptically and postsynaptically to organize the proper synaptic apparatus.
Synaptic transmission is reduced in dMegf8 mutants
As shown in the preceding sections, loss of dMegf8 results in reduced synaptic growth at the larval NMJ (Fig. 2) and causes ultrastructural abnormalities at the synapse (Fig. 4). We next examined the consequences of loss of dMegf8 on synaptic transmission at the NMJs, as well as whether presynaptic or postsynaptic expression of dMegf8 in dMegf8 mutants will restore the synaptic transmission in dMegf8 mutants. We performed electrophysiological analyses on muscle 6 of third-instar larval body walls of all relevant genotypes and recorded the EJPs in 0.5 mm [Ca2+]o at 0.2 Hz under identical conditions (Feeney et al., 1998; Shi et al., 2019). Representative EJP graphs from the control (wCS) (Fig. 5A), dMegf8HSC mutants (Fig. 5B), presynaptic dMegf8 expression in elav>dMegf8;dMegf8HSC mutants (Fig. 5C), and postsynaptic dMegf8 expression in MHC>dMegf8;dMegf8HSC mutants (Fig. 5D). dMegf8HSC mutants exhibited a reduction in EJP amplitude (Fig. 5B, quantified in Fig. 5E). The presynaptic expression of dMegf8 was able to fully rescue the EJP amplitude of dMegf8HSC mutants (Fig. 5C,E), whereas the postsynaptic dMegf8 expression did not rescue the EJP amplitude in dMegf8HSC mutants (Fig. 5D,E). Interestingly, the mEJP amplitudes did not show any significant differences in dMegf8HSC mutants, in elav>dMegf8;dMegf8HSC mutants and in MHC>dMegf8;dMegf8HSC mutants compared with wCS controls, suggesting that the synaptic vesicle contents were not altered by loss of dMegf8 (Fig. 5F). dMegf8HSC mutants and postsynaptic dMegf8 expression in MHC>dMegf8;dMegf8HSC mutants revealed severely decreased quantal contents compared with wCS controls, and the presynaptic dMegf8 expression in elav>dMegf8;dMegf8HSC mutants was similar to wCS controls (Fig. 5G). The paired pulse ratio (Fig. 5H) and the mEJP frequency (Fig. 5I) in dMegf8HSC mutants, presynaptic dMegf8 expression in elav>dMegf8;dMegf8HSC mutants and postsynaptic dMegf8 expression in MHC>dMegf8;dMegf8HSC mutants did not show any significant differences compared with wCS controls. Together, our data show that dMegf8 functions presynaptically for synaptic transmission at the NMJs.

dMegf8 is required for proper synaptic transmission. A–D, Representative electrophysiological traces showing EJPs from (A) WT (wCS), (B) dMegf8HSC mutant, (C) presynaptic rescue (elav>dMegf8;dMegf8HSC), and (D) postsynaptic rescue (MHC>dMegf8;dMegf8HSC). E–I, Quantification of (E) EJP amplitude, (F) mEJP amplitude, (G) quantal contents, (H) paired pulse ratio, and (I) mEJP frequency in respective genotypes. E–I, Data are mean ± SEM (one-way ANOVA test with Tukey's multiple comparisons). E, F(4,58) = 11.53, +/+ versus dMegf8HSC: ****p < 0.0001, +/+ versus elav>dMegf8; dMegf8HSC: p = 0.9980, +/+ versus MHC>dMegf8; dMegf8HSC: **p = 0.0026. F, F(4,58) = 4.311, +/+ versus dMegf8HSC: p = 0.9658, +/+ versus elav>dMegf8; dMegf8HSC: p = 0.9936, +/+ versus MHC>dMegf8; dMegf8HSC: p = 0.0861. G, F(4,43) = 13.95, +/+ versus dMegf8HSC: ****p < 0.0001, +/+ versus elav>dMegf8; dMegf8HSC: p = 0.9503, +/+ versus MHC>dMegf8; dMegf8HSC: ****p < 0.0001. H, F(4,58) = 3.195, +/+ versus dMegf8HSC: p = 0.1143, +/+ versus elav>dMegf8; dMegf8HSC: p = 0.2908, +/+ versus MHC>dMegf8; dMegf8HSC: p = 0.0800. I, F(4,58) = 6.865, +/+ versus dMegf8HSC: p = 0.9998, +/+ versus elav>dMegf8; dMegf8HSC: p = 0.9088, +/+ versus MHC>dMegf8; dMegf8HSC: p = 0.2920.
Synaptic proteins regulating NMJ growth and structure show reduced expression in dMegf8 mutants
Since dMegf8 is a transmembrane protein and dMegf8 mutants display synaptic growth defects as well as deficits in synaptic ultrastructure and synaptic transmission, we next wanted to examine whether dMegf8 has any association or interactions with proteins that regulate synaptic growth and architecture. Synaptic transmembrane proteins are critical for trans-synaptic adhesion and signaling for proper synaptic organization and function (Sun and Xie, 2012; Banerjee et al., 2017; Banerjee and Riordan, 2018). Since most phenotypes of dMegf8HSC mutants were rescued presynaptically, dMegf8 likely functions in association with other presynaptic proteins to coordinate synaptic growth and organization. Two key proteins in the synaptic machinery that are well known in regulating synaptic growth, organization, and function are the trans-membrane proteins, Drosophila neurexin (Dnrx) (Li et al., 2007; Zeng et al., 2007) and the Type II receptor of the BMP signaling pathway wishful thinking (wit) (Aberle et al., 2002; McCabe et al., 2003; Banerjee et al., 2017; Banerjee and Riordan, 2018; Guangming et al., 2020). We next set out to investigate whether dMegf8 coordinates synaptic growth with Dnrx and/or Wit and might be part of a synaptic membrane protein complex. We first wanted to test whether the endogenous dMegf8 localization was affected in larval NMJ of dnrx and wit mutants. Similar to data presented in Figure 2, endogenous dMegf8 (green, Fig. 6A-D′′) localized to the NMJ synaptic terminals in WT (Fig. 6A,A′′) and absent in dMegf8HSC mutants (Fig. 6B,B′′). Compared with the WT dMegf8 localization (Fig. 6A,A′′), the dMegf8 localization in dnrx (Fig. 6C,C′′) and wit mutants (Fig. 6D,D′′) was diffuse with a significant decrease in the fluorescence intensity levels in both dnrx and wit mutants (quantified in Fig. 6J). Brp was used as a presynaptic marker (red, Fig. 6A′,A′′,B′,B′′,C′,C′′,D′,D′′). Next, we wanted to examine any alterations in the localization and/or fluorescence intensities of Dnrx and Wit in dMegf8HSC mutants by immunolocalization of Dnrx in WT, dMegf8HSC mutants, and dnrx mutants NMJs. We observed that Dnrx localization in dMegf8HSC mutants (Fig. 6F,F′) was also significantly reduced compared with WT controls (Fig. 6E,E′, quantified in Fig. 6K). We also tested for Wit localization; however, the endogenous Wit levels in the larval NMJs were undetectable using standard immunohistochemistry protocols with anti-Wit antibodies as reported previously (Nahm et al., 2013; Banerjee et al., 2017). Therefore, to study Wit localization, we followed a strategy of overexpressing Wit presynaptically using the elav-Gal4 driver in WT (elav-Gal4; UAS-wit, Fig. 6H,H′) and in dMegf8HSC mutant background (elav-Gal4/UAS-wit;dMegf8HSC, Fig. 6I,I′). The expression of Wit in elav-Gal4;UAS-wit;dMegf8 NMJs (red, Fig. 6I,I′) was dramatically reduced compared with Wit expression in elav-Gal4; UAS-wit (red, Fig. 6H,H′, quantified in Fig. 6L) in the WT background. The immunolocalization and levels of Dnrx and Wit were not significantly different between dMegf8HSC/dMegf8HSC homozygous mutants compared with dMegf8Δ8/dMegf8HSC trans-allelic mutants (data not shown). Together, these data demonstrate that dMegf8 is necessary for the proper localization and clustering of presynaptic Dnrx and Wit, and that these proteins may be interdependent for their proper assembly at the NMJ synaptic terminals.

Interdependency of dMegf8, Dnrx, and Wit in their localization and stability. A–D′′, Confocal images in (A) +/+, (B) dMegf8HSC mutant, (C) dnrx mutant, and (D) wit mutant third instar larval NMJ labeled with anti-dMegf8 (green) and anti-Brp (red). E–G′, Confocal images in (E) +/+, (F) dnrx mutant, and (G) dMegf8HSC mutant third instar larval NMJ labeled with anti-Dnrx (green) and anti-Brp (red). H–I′, Confocal images of (H) elav>wit and (I) elav>wit; dMegf8HSC larval NMJ labeled with anti-Hrp (green) and anti-Wit (red). J, Quantification of dMegf8 fluorescence intensity/bouton area in WT (+/+), dMegf8HSC, dnrx, and wit mutants. Data are mean ± SEM; F(7,37) = 18.25, +/+ versus dMegf8HSC: ****p < 0.0001, +/+ versus dnrx−/−: ***p = 0.0005, +/+ versus wit−/−: ****p < 0.0001 (one-way ANOVA test with Tukey's multiple comparisons). K, Quantification of Dnrx fluorescence intensity/bouton area in WT (+/+), dnrx, and dMegf8HSC mutants. Data are mean ± SEM; F(2,21) = 46.54, +/+ versus dnrx−/−: ****p < 0.0001, +/+ versus dMegf8HSC: ****p < 0.0001 (one-way ANOVA test with Tukey's multiple comparisons). L, Quantification of Wit/Hrp fluorescence intensity ratio in elav>wit and elav>wit; dMegf8HSC mutants. Data are mean ± SEM; t(24) = 11.02, ****p < 0.0001 (unpaired Student's t test). Scale bars: A–D′′, 10 μm; E–G′, 5 μm; H–I′, 5 μm.
dMegf8 mutants show normal levels pMad and Trio, and dMegf8 shows genetic interactions with dnrx and wit
Since Wit localization was significantly reduced by loss of dMegf8, we next determined whether any downstream effectors of the BMP signaling pathway would also get impacted by loss of dMegf8. At the Drosophila NMJ, BMP signaling is essential for synaptic growth and homeostasis (Bayat et al., 2011). In retrograde BMP signaling pathway, the ligand glass bottom boat (Gbb) from postsynaptic muscles binds to presynaptic Type I receptors Thickveins (Tkv), Saxophone (Sax), and Type II receptor Wit leading to increased phosphorylation of BMP transcription factor, Mothers against dpp (Mad), and its subsequent accumulation in the nucleus (Aberle et al., 2002; Marques et al., 2002; McCabe et al., 2003; Dudu et al., 2006; Ball et al., 2010). pMad binds directly to Trio promoter and enhance the transcription of Trio. Trio activates other downstream effectors in the neuronal soma or at the synapses leading to alterations in the actin cytoskeleton and regulating synaptic growth (Awasaki et al., 2000; Ball et al., 2010). Immunolabeling of NMJs with anti-pMad (PS-1, red) and Hrp (green) in WT (Fig. 7A,A′) and dMegf8HSC mutants (Fig. 7B,B′) did not show any difference in fluorescence intensities of pMad (quantification in Fig. 7E). Similarly, pMad (SMAD, red) fluorescence levels in WT (Fig. 7C, quantified in Fig. 7F) and dMegf8HSC mutant VNC (Fig. 7D, quantified in Fig. 7F) also did not show any significant differences. Total levels of pMad (Smad) analyzed by immunoblots (Fig. 7G, quantified in Fig. 7I) from VNC of WT and dMegf8HSC mutants showed no significant difference while pMad levels in wit mutants showed severe reduction as reported previously (Fig. 7G,I) (Banerjee et al., 2017). Trio levels assessed by immunoblotting analysis of VNC also showed no significant difference between WT and dMegf8HSC mutants (Fig. 7H, quantified in Fig. 7J). These data indicate that loss of dMegf8 does not impact the levels of downstream BMP effectors, pMad and Trio.

BMP downstream effectors in dMegf8 mutants and genetic interactions between dMegf8, dnrx, and wit. A–B′, Confocal images in (A) +/+ and (B) dMegf8HSC mutant third instar larval NMJ labeled with anti-PS-1 (red) and anti-Hrp (green). C, D, Confocal images of (C) +/+ and (D) dMegf8HSC mutant larval VNC labeled with anti-Smad (red). E, Quantification of PS-1 fluorescence intensity/bouton area in the specified genotypes in A, B. Data are mean ± SEM; t(34) = 0.1063, p = 0.9159 (unpaired Student's t test). F, Quantification of Smad fluorescence intensity/bouton area in the specified genotypes in C, D. Data are mean ± SEM; t(34) = 1.063, p = 0.2951 (unpaired Student's t test). G–J, Representative immunoblots showing total levels of Smad (G) and Trio (H), and corresponding quantification of ratio of band intensities of Smad (I) and Trio (J). I, Data are mean ± SEM; F(2,6) = 31.42, +/+ versus dMegf8HSC: p = 0.7653, +/+ versus wit−/−: **p= 0.0011 (one-way ANOVA with Tukey's multiple comparisons). J, Data are mean ± SEM; t(4) = 0.5412, p = 0.6171 (unpaired Student's t test). K–R, Confocal images in (K) dMegf8 heterozygote (dMegf8HSC+/−), (L) dMegf8, dnrx trans-heterozygote (dMegf8HSC+/−;dnrx+/−), (M) dnrx−/− mutant, (N) dMegf8, dnrx double mutant (dMegf8HSC−/−;dnrx−/−), (O) wit heterozygote (wit+/−), (P) dMegf8, wit trans-heterozygote (dMegf8HSC+/−;wit+/−), (Q) wit−/− mutant, and (R) dMegf8, wit double mutant (dMegf8HSC−/−;wit−/−) third instar larval NMJ labeled with anti-Hrp (green) and anti-Dlg (red). S, T, Quantification of total bouton numbers in indicated genotypes. S, Data are mean ± SEM; F(6,107) = 21.47, +/+ versus dMegf8+/−: p > 0.9999, +/+ versus dnrx+/−: p = 0.7589, +/+ versus dMegf8+/−;dnrx+/−: ****p < 0.0001, +/+ versus dnrx−/−: ****p < 0.0001, +/+ versus dMegf8−/−: ****p < 0.0001, +/+ versus dMegf8−/−; dnrx−/−: ****p < 0.0001, dMegf8+/−; dnrx+/− versus dnrx−/−: p = 0.5614, dnrx−/− versus dMegf8−/−: p = 0.9379, dnrx−/− versus dMegf8−/−; dnrx−/−: p = 0.8967 (one-way ANOVA test with Tukey's multiple comparisons). T, Data are mean ± SEM; F(7,120) = 21.32, +/+ versus dMegf8+/−: p = 0.9211, +/+ versus witA12/-: p = 0.9985, +/+ versus dMegf8+/−; witA12/-: ***p = 0.0009, +/+ versus dMegf8+/−; witB11/-: ***p = 0.0007, +/+ versus dMegf8−/−: ****p < 0.0001, +/+ versus witA12/B11: ****p < 0.0001, +/+ versus dMegf8−/−; witA12/B11: ****p < 0.0001, dMegf8−/− versus witA12/B11: p = 0.9612, dMegf8−/− versus dMegf8−/−; witA12/B11: p = 0.9994 (one-way ANOVA test with Tukey's multiple comparisons). Scale bars: A–H, 20 μm.
Given the interdependency of dMegf8, Dnrx, and Wit localization as shown in Figure 6 and a common synaptic undergrowth phenotype seen in dMegf8, dnrx, and wit mutants (Fig. 2) (Aberle et al., 2002; Marques et al., 2002; Li et al., 2007; Banerjee et al., 2017), we were interested in determining whether dMegf8, dnrx, and wit displayed any genetic interactions to coordinate synaptic growth. Thus, for our genetic interaction studies, we used the synaptic undergrowth as our phenotypic readout and generated various genetic combinations of dMegf8 and dnrx as well as dMegf8 and wit (Fig. 7K–R). First, we examined the synaptic growth of dMegf8HSC+/− (Fig. 7K,S) and dnrx+/− (Fig. 7S) heterozygotes compared with WT and found no significant differences (Fig. 7S). Next, we analyzed the double heterozygous combination of dMegf8 HSC+/−; dnrx+/− (Fig. 7L,S) and found a significant decrease in bouton numbers compared with the single heterozygotes of dMegf8HSC+/− and dnrx+/− (Fig. 7S). We further compared the bouton numbers in single mutants of dMegf8 HSC−/− (Fig. 7S) and dnrx−/− (Fig. 7M,S) and found no significant differences between the two genotypes (Fig. 7S). Similarly, double mutants of dMegf8 HSC−/−; dnrx−/− (Fig. 7N,S) showed similar reduction in bouton growth compared with dMegf8 HSC−/− or dnrx−/− single mutants (Fig. 7S). These data suggest that dMegf8 and Dnrx loss affects synaptic growth similarly and that they may function together in regulating NMJ synaptic growth.
We next tested genetic interactions between dMegf8 and wit using a similar approach as presented above. NMJ bouton counts of dMegf8 HSC+/− and wit+/− (Fig. 7O,T) showed no differences compared with WT (Fig. 7T), while the double heterozygous combination of dMegf8 HSC+/−;wit+/− (Fig. 7P,T) displayed a significant reduction. Next, the single mutants of dMegf8HSC−/− and wit−/− (Fig. 7Q,T) showed undergrowth phenotypes similar to dMef8HSC−/−;wit−/− double mutants (Fig. 7R,T). These data indicate that dMegf8 and wit display genetic interactions and are involved in coordinating synaptic growth at NMJ. Together, our data show that dMegf8, dnrx, and wit show genetic interactions and function in synaptic growth and organization at the NMJ synapses without severely impacting the BMP signaling pathway.
dMegf8, Dnrx and Wit function in a biochemical complex
Given that the dMegf8 fluorescence intensity was reduced in dnrx and wit mutants and the localization of Dnrx and Wit was also affected in dMegf8 mutants, we wanted to examine the total levels of these proteins in the mutant backgrounds of one another (Fig. 8). In addition, since dMegf8 displayed genetic interactions with dnrx and wit, we also wanted to determine whether dMegf8, Dnrx, and Wit existed as an in vivo biochemical complex and potentially function together. Therefore, we performed immunoblots and coimmunoprecipitations to resolve the question whether they formed a molecular complex. Immunoblots were performed both from adult fly heads (Fig. 8A,B,D) and third instar larval musculature that contains NMJs (Fig. 8F,H), mostly because the expression levels of dMegf8 and Dnrx in the larval musculature were too low to be detected. All coimmunoprecipitation analyses were performed using fly head lysates because of large amount of protein needed for these experiments.

Biochemical interactions between dMegf8, Dnrx, and Wit. A–C, Representative immunoblots from adult head lysates showing total levels of dMegf8 in WT (+/+), dMegf8F1/Df (A) and in dnrx mutant (B), and quantification of the normalized ratio of protein band intensities (C). +/+ versus dMegf8F1/Df: t(4) = 6.815, **p = 0.0024 (unpaired Student's t test); +/+ versus dnrx−/−: t(6) = 0.1961, p = 0.8510 (unpaired Student's t test). D, E, Representative immunoblots from adult head lysates showing total levels of Dnrx in WT (+/+), dMegf8HSC, and dnrx mutants (as a negative control), and quantification of the normalized ratio of protein band intensities (E). +/+ versus dMegf8HSC: t(6) = 0.3921, p = 0.7085 (unpaired Student's t test). F, G, Representative immunoblots from larval musculature lysates showing levels of Wit in specified genotypes, and quantification of the normalized ratio of protein band intensities (G). F(2,11) = 45.91, +/+ versus dMegf8HSC: *p = 0.0107, +/+ versus elav>dMegf8: ***p = 0.0003 (one-way ANOVA test with Tukey's multiple comparisons). H, I, Representative immunoblots from larval musculature lysates showing levels of Wit in specified genotypes, and quantification of the normalized ratio of protein band intensities (I). F(2,12) = 37.98, +/+ versus elav>wit;dMegf8HSC: *p = 0.0396, +/+ versus elav>wit: ****p < 0.0001 (one-way ANOVA test with Tukey's multiple comparisons). Actin was used as the loading control in A, B, D, F, H. J, K, dMegf8 coimmunoprecipitated with anti-Dnrx (J) and anti-Wit (K) antibodies, respectively, and probed with anti-dMegf8. C, E, G, I, Data are mean ± SEM; *p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001; unpaired Student's t test (C,E) and one-way ANOVA test with Tukey's multiple comparisons (G,I).
First, we examined the total levels of dMegf8 in the dMegf8 point mutation allele isolated from the ethyl methane sulfonate screen, dMefg8F1 (Fig. 8A, quantified in Fig. 8C). dMegf8 levels were reduced by ∼25% in dMefg8F1 mutants compared with WT control (Fig. 8A, quantified in Fig. 8C). Immunoblotting analyses showed that dMegf8 levels (Fig. 8B, quantified in Fig. 8C) were not significantly affected in dnrx mutants (Fig. 8B, quantified in Fig. 8C). Likewise, Dnrx levels (Fig. 8D, quantified in Fig. 8E) levels were not significantly altered in dMegf8 mutants compared with the WT in the fly head lysate preparations, indicating that in the adult head lysates the protein levels of dMegf8 and Dnrx were not affected in each other's mutants. We next tested the levels of Wit in WT and loss- and gain-of dMegf8 backgrounds using the larval musculature (Fig. 8F,H). The total levels of Wit were significantly decreased in dMegf8HSC mutants compared with WT (Fig. 8F, quantified in Fig. 8G). Wit levels were significantly elevated in presynaptic dMegf8 overexpression (Fig. 8F, quantified in Fig. 8G). Also, consistent with the finding that Wit fluorescence levels were reduced at NMJ (Fig. 6), total levels of Wit from lysates of Wit overexpression in dMegf8 mutant background (elav-Gal4;UAS-wit;dMegf8−/−) were also significantly decreased when compared with Wit overexpression in the WT background (Fig. 8H, quantified in Fig. 8I), indicating that dMegf8 loss led to a significant reduction in total Wit levels. Actin was used as loading control for all immunoblots (Fig. 8A,B,D,F,H).
For coimmunoprecipitation analyses, because of the extremely low level of dMegf8 expression, no association with dMegf8 and Dnrx/Wit was observed with coimmunoprecipitations using antibodies to endogenous proteins at normal levels. To overcome this, we used adult head lysates from presynaptic dMegf8 overexpression to test whether Dnrx and Wit would coprecipitate dMegf8. As shown in Figure 8J, immunoblots of dMegf8 show IP with Dnrx antibodies coprecipitated dMegf8 from dMegf8 overexpression lysates, which was absent from dnrx mutant IPs confirming the association between dMegf8 and Dnrx. Coimmunoprecipitation with anti-Wit monoclonal antibodies also detected dMegf8 from dMegf8 overexpression lysates, which was absent in the Protein-A bead controls (Fig. 8K). IP using Dnrx and Wit antibodies efficiently precipitated Dnrx and Wit, respectively (data not shown) (Banerjee et al., 2017). Together, these results demonstrate that dMegf8, Dnrx, and Wit exist in vivo as a molecular complex and may function together in synaptic organization and function.
Discussion
Many neurodevelopmental and psychiatric disorders have been associated with disturbances in synaptic organization and function (Nanou and Catterall, 2018; Batool et al., 2019; Parenti et al., 2020). Since CS subjects show intellectual disabilities (Taravath and Tonsgard, 1993; Twigg et al., 2012; Giacopuzzi et al., 2017), it has remained unknown whether human MEGF8 will have a role in synaptic function. Our characterization of dMegf8 in the neuromuscular synapses provides the first evidence that Megf8 proteins have synaptic functions. Our findings reveal that dMegf8 mutants have multiple abnormalities in synaptic development, ultrastructural organization, and physiological functions, and that dMegf8 forms a biochemical complex with two well-characterized synaptic proteins Dnrx and Wit. Our studies provide insights into possible human MEGF8 functions and lay the groundwork for further characterization of the mechanisms underlying the intellectual disabilities associated with the CS.
Megf8 in synaptic development and function
Megf8 has been linked to various developmental abnormalities, including congenital heart defects as well as neurodevelopmental and axon guidance defects with early lethality associated with the murine homozygous null allele (Engelhard et al., 2013; Kong et al., 2020; Wang et al., 2020). These phenotypes have pointed to defects in Hedgehog and BMP signaling pathways (Twigg et al., 2012; Engelhard et al., 2013; Lloyd et al., 2018; Kong et al., 2020; Wang et al., 2020). The dMegf8HSC mutants reported here produced viable and fertile adults with motor coordination deficits (Fig. 1), unlike the dMegf8 null reported by Lloyd et al. (2018), showing a late larval lethality. Although both alleles were generated using CRISPR/Cas9 methodology, it remains to be seen whether genetic background or a second site mutation caused dMegf8Δ8 lethality (Lloyd et al., 2018). Our studies provide evidence that dMegf8 plays an important role during Drosophila larval NMJ development as dMegf8 localizes in synaptic terminals both presynaptic and postsynaptically, and loss of dMegf8 leads to synaptic undergrowth, while overexpression of dMegf8 in neurons causes synaptic overgrowth pointing to a presynaptic dMegf8 requirement. The synaptic undergrowth in dMegf8 mutants is fully rescued by presynaptic and not by the postsynaptic expression of dMegf8, further demonstrating that dMegf8 functions primarily in the presynaptic compartment.
While we did not see any disruption in the apposition of the presynaptic active zone protein, BRP, with the postsynaptic GluRIIA, the number of BRP-positive puncta/bouton area was significantly increased in dMegf8 mutants, which was consistent with the ultrastructural analyses revealing increased active zones in dMegf8 mutant synapses (see below). These findings are suggestive of a loss of dMegf8 impacting the proper organization of synaptic active zones. dMegf8 mutants displayed diffuse Dlg distribution throughout the boutons unlike the peripheral rim of the boutons where Dlg normally localizes. Presynaptic expression of dMegf8 in dMegf8HSC mutants was able to restore Dlg localization better than the postsynaptic expression of dMegf8. Dlg has been previously shown to function both presynaptic and postsynaptically and loss of Dlg affects the SSR at the boutons (Budnik et al., 1996). The synaptic structural abnormalities are often accompanied by corresponding functional abnormalities. We observed significant reduction in the EJP amplitude and the quantal content in dMegf8 mutants, which was restored by dMegf8 presynaptic and not by the postsynaptic expression. dMegf8 loss also affected the ultrastructural organization of the synapses with specific defects in the presynaptic elements as well as the postsynaptic specializations. Interestingly, restoration of dMegf8 expression either presynaptically or postsynaptically revealed that most functions of dMegf8 are presynaptic, except the SSR specialization, which was rescued by dMegf8 postsynaptic expression, suggesting that dMegf8 also has postsynaptic functions. Our studies are consistent with many of the synaptic phenotypes previously observed in mutations associated with genes involved in trans-synaptic functions (e.g., dnrx, dnlg1, dnlg2, and wit) (Aberle et al., 2002; McCabe et al., 2003; Li et al., 2007; Banovic et al., 2010; Chen et al., 2012; Banerjee et al., 2017), indicating that dMegf8 may be involved or functions in close association with these proteins. Our data thus provide insights that vertebrate Megf8 may also play an essential role in synaptic function as human MEGF8 has been linked to neurodevelopmental and psychiatric disorders. As our studies highlight the synaptic role of dMegf8 at the NMJs, it is possible that dMegf8 also functions in proper axonal growth, guidance, maturation, and fasciculation during neuronal development. As reported previously, CS patients present multiple developmental anomalies in addition to the intellectual disabilities (Engelhard et al., 2013), it is plausible that human MEGF8 might be involved in a plethora of functions in the developing nervous system ranging from neuronal survival, axon growth, guidance, maturation, fasciculation, target innervation, and synapse formation. Future studies on dMegf8 will explore its functions in some of these neural development processes.
dMegf8 and BMP signaling at the NMJ
The dMegf8 primary structure and its synaptic localization and the NMJ synaptic defects observed in dMegf8 mutants suggested that it could potentially interact with other known synaptic proteins. Our immunohistochemical analyses showed that the absence of dMegf8 or Dnrx leads to their diffuse synaptic localization as well as protein instability in each other's mutant backgrounds (refer Fig. 6). However, the immunoblot analysis of dMegf8 or Dnrx did not show any variation in overall levels in each other's mutant backgrounds (Fig. 8), suggesting that, despite the total protein levels remaining unchanged, dMegf8 and Dnrx fail to properly localize and cluster at the NMJ terminals and remain diffuse indicating that dMegf8 and Dnrx are mutually required for their proper synaptic localization. This is also reflected by the reduced levels of Wit in dMegf8 mutants, indicating that loss of dMegf8 affects Wit localization and/or its stability at the NMJ. Together, these data underscore the important role that dMegf8 plays in the proper localization of other synaptic proteins and suggest interdependency of dMegf8, Dnrx, and Wit for their proper subcellular localization and/or stability at the NMJ.
Previously, Dnrx was shown to be essential for proper synaptic growth (Li et al., 2007; Südhof, 2008), and regulation of the BMP signaling pathway in coordination with BMP Type II receptor, Wit (Banerjee et al., 2017; Banerjee and Riordan, 2018). Interestingly, the murine Megf8 has also been shown to function as a modifier of BMP4 signaling in the trigeminal ganglion neurons (Engelhard et al., 2013). These observations suggest that dMegf8 along with other proteins is involved in BMP signaling. This is further strengthened by our genetic analysis, which revealed that dMegf8 displays genetic interactions dnrx and wit, and that trans-heterozygous combinations of dMegf8HSC+/−;dnrx+/− and dMegf8HSC+/−;wit+/− displayed a significant reduction in bouton growth compared with the single heterozygotes, supporting the possibility that these genes function together in a dose-dependent manner. Furthermore, double mutants of dMegf8HSC−/−;dnrx−/− and dMegf8HSC−/−,wit−/− showed no significant differences in the bouton counts compared with their single mutants. These findings strongly support that dMegf8, dnrx, and wit function cooperatively to coordinate synaptic growth. Interestingly, however, there was no significant difference in pMad localization either in the VNC or at the NMJ or Trio levels in dMegf8 mutants compared with WT (Fig. 7). One of the possibilities might be that dMegf8 works with Dnrx and Wit in a large complex structurally to regulate synaptic development, but not participate directly in signaling functions as part of the BMP pathway (Fig. 9). Another possibility is that, although the proper localization and stability of Wit requires dMegf8, other BMP receptors, such as Tkv and Sax, could still function effectively to activate downstream signaling of the BMP pathway. Thus, dMegf8 may function presynaptically to coordinate BMP signaling to ensure normal synaptic bouton growth and also postsynaptically to organize the SSR and other postsynaptic structures, which are both necessary for proper synaptic function. A more detailed analyses of some of these questions will be addressed as part of our future investigations.

Schematic model of dMegf8 function at the NMJ synapse. A schematic model depicting dMegf8 and other known presynaptic and postsynaptic proteins at the NMJ. Based on our immunohistochemical, ultrastructural, electrophysiological, biochemical, and genetic rescue analyses, we propose that dMegf8 functions presynaptically to coordinate BMP signaling for the synaptic bouton growth and postsynaptically to organize the SSR and other synaptic structures that are necessary for proper synaptic function. While dMegf8 interactions with Dnrx and Wit are established, uncovering potential molecular interactions between dMegf8 and Dnlg1, Tkv, and Sax, or other synaptic proteins would shed further light on the functions of dMegf8 at the synapse.
dMegf8 molecular complex
Previous studies on Megf8 have not reported any biochemical interactions with other proteins that would link Megf8 to neuronal functions. Our biochemical analyses of dMegf8 showed that dMegf8, Dnrx, and Wit exist in a large biochemical complex. While the overall levels of dMegf8 and Dnrx did not seem to change in each other's mutant backgrounds using adult head lysates, the levels of these proteins at the NMJ containing musculature are too low to detect any changes in their levels. Interestingly, the total levels of Wit using the musculature lysates were affected by the presence or absence of dMegf8, as presynaptic expression of dMegf8 increased Wit levels at the NMJ. As the endogenous dMegf8 levels were extremely low for the IP analysis, we used dMegf8 overexpression for IP, which revealed that dMegf8, Dnrx, and Wit form a large protein complex. How these proteins may interact with each other and what the stoichiometry of these interactions is remain to be fully investigated. Given the complex domain structure and large size of dMegf8, it is likely that dMegf8 interacts with a host of other presynaptic and postsynaptic proteins. It will be of significant interest to know what these proteins are and how they function in a macromolecular complex at the NMJ or other synapses. Further elucidation of the dMegf8 genetic and molecular functions and the signaling complexes that dMegf8 engages in will uncover the relevant functions of the human MEGF8 that is associated with deficits seen in the Carpenter syndrome or other psychiatric disorders.
Footnotes
This work was supported by the Zachry Foundation for Neuroscience Research and the University of Texas Health Science Center at San Antonio to M.A.B. We thank all members of the S.B. and M.A.B. laboratories for helpful discussions; and the Electron Microscopy Core Facility at UT Health for assistance.
The authors declare no competing financial interests.
- Correspondence should be addressed to Swati Banerjee at banerjees{at}uthscsa.edu or Manzoor A. Bhat at bhatm{at}uthscsa.edu





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}