
Abstract
In vertebrates, the receptor families deleted in colorectal cancer (DCC) and UNC5 mediate responses to the bifunctional guidance cue netrin-1. DCC mediates attraction, whereas a complex of DCC and UNC5 mediates repulsion. Thus, a primary determinant of the responsiveness of an axon to netrin-1 is the presence or absence of UNC5 family members on the cell surface. Currently, little is known about the role of receptor trafficking in regulating neuronal responses to netrin-1. We show that protein interacting with C-kinase 1 (PICK1) recruits activated protein kinase Cα (PKCα) to MycUNC5A at the plasma membrane, stimulating its endocytosis. We identify two PKCα phosphorylation sites at serines 408 and 587, as well as dileucine internalization motifs, which are required for this endocytosis. We find that PKCα-stimulated internalization of UNC5A alters the functional response of developing hippocampal axons to netrin-1, preventing UNC5A-mediated growth cone collapse and converting netrin-1-stimulated chemorepulsion to attraction. To address whether this conversion in axonal response occurs in neurons expressing endogenous levels of UNC5, we show that mouse cerebellar granule axons exhibit chemorepulsion in a netrin-1 gradient and that this chemorepulsion is converted to chemoattraction after PKCα activation. We demonstrate that this repulsion depends on UNC5A because Unc5a−/− axons are not repelled and show this conversion depends on PICK1 because PICK1−/− axons are not converted to chemoattraction after PKCα activation. Together, these data provide a potential mechanism to explain how developing neurons alter their responsiveness to netrin-1 at intermediate choice points as they navigate to their targets.
- trafficking
- PICK1
- UNC5
- netrin-1
- axon guidance
- PKC
Introduction
As developing neurons and axons navigate to targets, they must integrate signals from multiple guidance cues. Complicating this task is that guidance cues, like netrin-1 (NTN1), are bifunctional, acting as both attractants and repellants. Whether an axon is attracted or repelled depends on expression of two vertebrate families of netrin-1 receptors. The deleted in colorectal cancer (DCC) family, comprising DCC and neogenin (Keino-Masu et al., 1996), mediates attraction, whereas the UNC5 family, comprising UNC5A–D (Leonardo et al., 1997; Przyborski et al., 1998; Engelkamp, 2002), mediates repulsion in a complex with DCC (Hong et al., 1999). Thus, the primary difference between attraction and repulsion is the absence or presence of UNC5, but it remains unclear whether changes in cell surface levels of receptor determine sensitivity to netrin-1.
Several studies have shown that changes in second messenger signaling affect axonal responses to netrin-1. For example, attraction is converted to repulsion in Xenopus spinal neurons by inhibiting cAMP or protein kinase A (Ming et al., 1997) or by lowering intracellular calcium (Hong et al., 2000). Responses are also modulated by the ratio of cAMP/cGMP (Nishiyama et al., 2003) and by the activities of phosphatidylinositol-3-kinase (PI3-kinase), phospholipase C-γ (PLC-γ) (Ming et al., 1999), MAPK (mitogen-activated protein kinase) (Ming et al., 1999; Forcet et al., 2002; Campbell and Holt, 2003), and caspase-3 (Campbell and Holt, 2003). Although these studies describe alterations in responsiveness to netrin-1, little is known about the underlying mechanisms.
Recent evidence suggests that regulating the surface expression of receptors modulates responses to netrin-1. Increasing intracellular cAMP releases DCC to the cell surface (Bouchard et al., 2004) and increases axon extension in response to netrin-1, explaining, in part, the requirement for cAMP in netrin-1-mediated chemoattraction (Ming et al., 1997). Recently, we reported that protein interacting with C-kinase 1 (PICK1) recruits activated protein kinase Cα (PKCα) to UNC5A, resulting in specific loss of cell surface UNC5A (Williams et al., 2003b) and suggesting that PKCα modifies responses to netrin-1 by altering UNC5A surface expression. Together, these data support the hypothesis that second messenger signaling alters netrin-1 responsiveness by regulating cell surface receptor expression.
Here, we establish the mechanism underlying PKCα-stimulated loss of cell surface UNC5A and the functional consequences of this loss. Activated PKCα colocalizes with UNC5A at the surface, resulting in UNC5A internalization via endocytosis. Two PKCα phosphorylation sites in UNC5A are required as well as dileucine motifs, which likely serve as internalization signals. In addition, we demonstrate that PKCα-stimulated internalization of receptor reduces the responsiveness of UNC5A-expressing hippocampal axons to netrin-1, inhibiting growth cone collapse and converting chemorepulsion to attraction. To address whether this conversion occurs in neurons expressing endogenous UNC5A, we demonstrate that cerebellar granule axons are repelled by netrin-1 and this is converted to attraction after PKCα activation. This conversion requires UNC5A and PICK1 because cerebellar axons of Unc5a−/− neurons are not repelled by netrin-1, and cerebellar axons of PICK1−/− neurons are not converted to attraction after PKCα activation. Together, these data show that PICK1/PKCα-mediated endocytosis of UNC5A alters netrin-1-mediated responses.
Materials and Methods
Constructs and reagents.
The full-length rat MycUNC5A construct in the viral vector pSinRep5 (Invitrogen, Carlsbad, CA) was described previously (Williams et al., 2003b). Substitutions of codons for alanine were performed by PCR mutagenesis of MycUnc5A in the pSecTag (Invitrogen) vector using the QuickChange site-directed mutagenesis kit (Stratagene, La Jolla, CA). The MycUnc5A alanine substitution cDNAs, including a consensus Kozak and start site, were cloned into the viral vector pSinRep5 (Invitrogen) using the XbaI site. All PCR products and mutations were verified by sequencing. Phorbol 12-myristate 13-acetate (TPA) was obtained from Sigma (St. Louis, MO). Bisindolylmaleimide I (Bis) was obtained from Calbiochem (San Diego, CA). Sulfo-NHS-LC-biotin and immobilized NeutrAvidin agarose beads were purchased from Pierce (Rockford, IL). The chicken anti-Myc, anti-PKCα, anti-A2b antibodies were purchased from Aves Labs (Tigard, OR), BD Biosciences (San Jose, CA), or Alpha Diagnostics (San Antonio, TX), respectively. The anti-Myc (A-14), the anti-Myc (9E10), and anti-early endosomal antigen-1 (anti-EEA1) antibodies were a kind gift from Santa Cruz Biotechnology (Santa Cruz, CA). The goat anti-chicken FITC- and donkey anti-rabbit cyanine 3 (Cy3)-coupled secondary antibodies, as well as the Alexa543-conjugated phalloidin were purchased from Invitrogen.
Primary cell culture and infection.
Hippocampal neurons were harvested from embryonic day 19 (E19) rat tissue, dissociated with trypsin, triturated, and plated on poly-l-lysine treated glass coverslips (Assistent, Sondheim, Germany). Cells were plated in MEM with Earle's salts, 10% horse serum, 1 mm pyruvic acid, and 0.6% glucose at a density of 9.5 × 103 cells/cm2 for immunocytochemistry, collapse, and turning assays or 2.5 × 105 cells/cm2 for biotinylation experiments. After 2 h, media were replaced with Neurobasal A medium with B27 supplement, penicillin, streptomycin, and glutamine. Cells were incubated with pseudovirion-containing medium, encoding the receptor constructs as described, for 1 h and then returned to Neurobasal medium without virus for 24 h before experiments. Cerebellar granule cells were harvested from wild-type, Unc5a−/−, and PICK1−/− postnatal day 4 (P4) to P6 mice as described previously (Hatten et al., 1998). Briefly, cerebellums were treated with trypsin/DNase, triturated with fire-polished Pasteur pipettes, and isolated by Percoll gradient centrifugation. Granule cells were plated on poly-l-lysine treated glass coverslips (see above) at 2.0 × 104 cells/cm2 and incubated for 24–48 h in serum-free DMEM plus penicillin, streptomycin, glutamine, and 0.6% glucose and supplemented with B27 and N2 (Invitrogen).
Transgenic mice.
PICK1−/− mice were generated and characterized as described previously (Gardner et al., 2005). To generate Unc5a−/− mice, 4.8 and 3.6 kb Unc5a genomic fragments were amplified from E14 embryonic stem (ES) cells using the Expand High Fidelity PCR system (Hoffmann-La Roche, Nutley, NJ) and cloned into the targeting vector. The targeting vector was electroporated into E14 ES cells. Homologous recombination events, detected by a 1.3 kb external probe, resulted in the deletion of exons 4–8. From 576 ES cell clones screened by Southern blot, 10 clones were identified. Genotyping of Unc5a mice was performed using the following primers to detect +/+ Unc5a (AGGGCCAAGAGTCAGCTGATCCAG and AGTCAAGCCACTGTCTGGTTAGCCTG) and β-galactosidase (TCGTCTGCTCATCCATGACC and GTCTCGTTGCTGCATAAACC). Lack of Unc5a mRNA was confirmed by reverse transcription-PCR and lack of UNC5A protein by immunohistochemistry.
Immunostaining.
Cells were treated and immunostained as described previously (Williams et al., 2003b), except cells were fixed with 4% paraformaldehyde (PFA) in PBS and antibodies included the following: MycUNC5A live-label: chicken anti-Myc (1 μg/ml); MycUNC5A fixed and permeabilized label: rabbit anti-Myc (A-14) (1 μg/ml), 0.6 μg/ml anti-PKCα, mouse anti-EEA1 (1:250), and anti-A2b (100 μg/ml). For endocytosis experiments, hippocampal cultures were live-labeled with chicken anti-Myc antibody for 15 min and then TPA treated (100 ng/ml) for times from 0 to 45 min in 5 min increments. Cells were then rinsed with ice-cold TBS followed by 4 min treatment with 0.5 m NaCl and 0.2 m acetic acid at 4°C to break remaining extracellular antibody–antigen interactions and then rinsed three times in ice-cold TBS and fixed in 4% PFA for immunostaining (Carroll et al., 1999).
For the quantitative immunofluorescence assay, all images were acquired with a Zeiss (Oberkochen, Germany) LSM 5 PASCAL confocal microscope at 63× magnification. The laser intensity (10%), exposure, and gain settings per channel were the same for each image, and each image was collected in the first pass to avoid error caused by photobleaching. Segments of individual axons and dendrites were excised from the images and color channels were split and converted to grayscale using Adobe Photoshop (Adobe Systems, San Jose, CA). Lower and upper thresholds were constrained (150–250 on a scale from 0 to 255) to exclude background and saturated pixels, and images were analyzed with NIH ImageJ (version 1.32j). Mean cell segment values (n ≥ 3 per cell) were calculated for each condition using ≥10 cells in hippocampal cultures from four different litters of E19 embryos.
Biotinylation assay.
Biotinylation assay was performed as described previously (Williams et al., 2003b). Briefly, 24 h after infection, hippocampal cells were treated as described and incubated at 37°C during treatments. Cells were washed twice with ice-cold HEPES-buffered Ringer's solution [HBR; (in mm): 10 HEPES, 150 NaCl, 7.2 KCl, and 2.25 CaCl2, pH 7.5]. The cells were then incubated in 1 mg/ml Sulfo-NHS-LC-biotin in HBR at 4°C for 20 min. Excess biotin was quenched by washing twice for 20 min each in HBR plus 100 mm glycine. Finally, cells were washed with cold Tris-buffered saline, lysed with immunoprecipitation buffer (50 mm Tris, 150 mm NaCl, 5 mm EDTA, 1% Triton X-100, and 10% glycerol, pH 7.5, containing aprotinin, leupeptin, and PMSF at 1 μg/ml each), and immunoprecipitated using immobilized NeutrAvidin agarose beads and immunoblotted using anti-Myc 9E10 (1 μg/ml).
Collapse assay.
One day in vitro (DIV) hippocampal neurons were infected with bicistronic virus expressing the MycUNC5A receptors, as indicated, and green fluorescent protein (GFP). At 2 DIV, cells were treated with PBS, netrin (300 ng/ml), or TPA (100 ng/ml), followed by netrin-1 (300 ng/ml) as indicated. Cells were then fixed, permeabilized, and immunostained for GFP. Cells were fixed with 4% PFA, and filamentous actin (F-actin) was stained with Alexa543-conjugated phalloidin. Cells were examined for growth cone collapse under Cy3 fluorescent light using a 63× objective on a Zeiss Axiovert 200. For quantitation, a single cell with a clearly defined axon expressing GFP was considered collapsed if it had no lamellipodia and two or fewer filopodia. Each experimental condition was repeated three or more times, and at least 50 random growth cones were examined per experiment. Significance was assessed by nonparametric Mann–Whitney test using GraphPad Software (San Diego, CA) InStat version 3.0b for Macintosh.
Axon turning assay.
Hippocampal and granule cells were assayed at 1–2 d in vitro. Microscopic gradients of netrin-1 were created as described previously (Zheng et al., 1996; Ming et al., 1997). Briefly, a stable gradient was created and maintained using a Picospritzer II (Parker Hannafin, Fairfield, NJ) ejecting 5 μg/ml netrin-1 in PBS at a pressure of 3 psi, frequency of 2Hz, and duration of 20 ms from a micropipette with a 1 μm opening. The micropipette was positioned 100 μm from the growth cone center at an angle of 45° with respect to the last 10 μm segment of the axon shaft. Cell images were recorded at times 0 and 30 min. For dual turning experiments, the pipette orientation was corrected after 30 min, and a second turn was recorded from 30 to 70 min. Only neurons that grew >10 μm in 30 min and 15 μm in 70 min were counted. In control experiments, PBS alone was ejected from the micropipette. Neurite extension and angle were measured using NIH ImageJ (version 1.32j). DCC function blocking was achieved by incubating with anti-DCC antibody (1 μg/ml in media) for 30 min before gradient onset. UNC5A-expressing neurons were identified by GFP reporter as described above. Cells were maintained at 37°C and atmospheric CO2 on a heated stage in buffer consisting of 135 mm NaCl, 6 mm glucose, 5.4 mm KCl, 0.45 mm KH2PO4, 0.65 mm NaH2PO4–H2O, 10 mm HEPES, pH 7.4, and Glutamax-1 and B27 supplement (Invitrogen) for up to 90 min of growth. Images were acquired by phase contrast using a 20× objective on a Zeiss Axiovert 200. Significance was assessed by nonparametric Mann–Whitney test.
Results
PKCα is recruited to UNC5A at the plasma membrane after activation
Recently, our laboratory showed that UNC5A is coimmunoprecipitated with PICK1 and PKCα. Moreover, we demonstrated that the association of PKCα with UNC5A depends on the activation of PKCα and the ability of UNC5A to bind PICK1 (Williams et al., 2003b). To determine whether PKCα is recruited to UNC5A at the plasma membrane where it could locally regulate UNC5A surface expression, we assessed the subcellular localization of the complex containing UNC5A and PKCα by confocal microscopy. At 1–2 DIV, we infected hippocampal neurons with a N-terminally Myc-tagged UNC5A construct (MycUNC5A) using the Sindbis virus expression system. Twenty-four hours later, the surface expression of MycUNC5A was examined by live-labeling using an anti-Myc antibody, followed by fixation, permeabilization, and labeling with an antibody directed against PKCα.
Before stimulation, we find that PKCα staining appears in a punctate pattern evenly distributed within the growth cones of developing axons, whereas the live-labeled MycUNC5A appears punctate across the cell surface (Fig. 1A). Activation of PKC using TPA leads to recruitment of PKCα to MycUNC5A, as evidenced by overlapping staining at the plasma membrane of the growth cone (Fig. 1B). The labeling for both MycUNC5A and PKCα appears more smooth and homogeneous after TPA stimulation, and the growth cones have a simpler structure with fewer lamellipodia and filopodia (Fig. 1B). This is likely because of receptor clustering on the cell surface after recruitment of activated PKCα to the MycUNC5A/PICK1 complex. This type of clustering has been reported for epidermal growth factor receptors before endocytosis when they move quickly from lipid rafts to bulk plasma membrane where they can be internalized via clathrin-coated vesicles (Sorkin and Waters, 1993; Alexander, 1998; Carpenter, 2000). Interestingly, it was reported recently that netrin-1 signaling emanates from lipid rafts (Guirland et al., 2004), supporting the idea that the change in UNC5A staining and growth cone morphology after PKCα activation may be caused by migration of UNC5A from lipid rafts to other membrane microdomains before removal of UNC5A from the cell surface. As a control, we also expressed MycUNC5AΔ3C, a construct missing the three C-terminal amino acids that mediates the interaction of MycUNC5A with PICK1 and PKCα (Williams et al., 2003a). Accordingly, expression of MycUNC5AΔ3 essentially eliminates the smooth, homogeneous pattern of overlapping MycUNC5A/PKCα immunostaining seen for wild-type receptor after TPA stimulation (Fig. 1C). The role of the α isoform of PKC in the PICK1-dependent loss of UNC5A surface expression is consistent with previous findings demonstrating that PICK1 binds specifically to this isoform (Staudinger et al., 1997). Together with our previous data showing that UNC5A/PICK1 and activated PKCα form a complex, these results support a model in which PICK1 mediates recruitment of PKCα to UNC5A at the plasma membrane.

Activated PKCα is recruited to UNC5A at the plasma membrane of hippocampal growth cones. A, Live-labeled MycUNC5A receptors (green) do not colocalize with endogenous PKCα (red). B, Activated PKCα and MycUNC5A are colocalized at the cell surface after a 15 min TPA (100 ng/ml) treatment. C, MycUNC5AΔ3 does not colocalize with PKCα after TPA treatment. The MycUNC5AΔ3C construct encodes a C-terminal deletion of three amino acids that eliminates the PICK1 binding site, preventing PKCα recruitment to UNC5A. The dashed lines represent the outline of the growth cones as visualized in phase contrast. Scale bar, 5 μm.
PKCα-dependent endocytosis of UNC5A
At least two potential mechanisms, ectodomain shedding or receptor endocytosis, exist for removing receptors from the cell surface after PKCα activation. With shedding, the extracellular domain of the receptor is cleaved and released from the cell surface by proteolytic enzymes. Indeed, PKCα phosphorylates and activates proteolytic enzymes such as tumor necrosis factor-α converting enzyme, leading to the release of extracellular domains of receptors such as the growth hormone receptor, interleukin-15 receptor α, and c-Kit from the cell surface (Baumann, 2002; Baumann and Frank, 2002; Wang et al., 2002; Budagian et al., 2004; Cruz et al., 2004). To address whether PKCα activation leads to UNC5A ectodomain shedding, we examined the media for UNC5A extracellular domain after PKCα activation.
MycUNC5A-infected hippocampal neurons were incubated with PBS as a control or TPA to activate PKCα. The media were removed for TCA precipitation to assay for receptor shedding, and the cultures were biotinylated to confirm cell surface expression of the receptor (Fig. 2A). To biotinylate cell surface receptors, the infected neurons were placed on ice to stop cellular trafficking, and surface proteins were labeled with biotin. Labeling was then quenched and biotinylated proteins were precipitated from cell lysates using streptavidin–agarose beads. Both the biotinylated proteins and TCA media precipitates were subsequently separated by SDS-PAGE and Western blotted using anti-Myc antibody. As expected (Williams et al., 2003b), PKCα activation leads to a loss of UNC5A from the cell surface, as evidenced by lower levels of surface, biotinylated receptor in TPA treated cultures (Fig. 2A). Immunoblots of the TCA-precipitated proteins from the media reveal that, although some UNC5A ectodomain is shed, there is no increase in UNC5A ectodomain after TPA treatment (Fig. 2A). It should be noted that detection of any shed protein in the TCA-precipitated protein blots required long exposures (>2 h), indicating that very low levels of MycUNC5A ectodomain are released into the media. In addition, attempts to immunoprecipitate and immunoblot media from TPA-treated hippocampal cultures failed to show an increase in shed MycUNC5A (data not shown). These data indicate that receptor ectodomain shedding does not contribute to the PKCα-mediated loss of UNC5A from the cell surface.

UNC5A is internalized, and not shed, after PKCα activation. A, Hippocampal neurons expressing MycUNC5A were TPA treated (100 ng/ml) as indicated to reduce cell surface expression of MycUNC5A. Biotinylation assays demonstrate the loss of MycUNC5A from the cell surface without a concomitant increase in UNC5A ectodomain in the TCA-precipitated cell-culture media (compare top blot with middle blot). Total cellular MycUNC5A was evaluated to control for infection efficiency. B–D, MycUNC5A receptors colocalize with EEA-1 after brief TPA treatment. B, Surface MycUNC5A receptors were live-labeled and treated with TPA to stimulate internalization. Antibody was subsequently stripped from noninternalized MycUNC5A by mild acid treatment. C, Brief (5 min) TPA treatment induces MycUNC5A colocalization with EEA-1. D, After longer (40 min) TPA treatment, there is diminished colocalization of MycUNC5A with EEA-1 likely caused by continued trafficking of MycUNC5A. Scale bar, 10 μm.
PKCα activation has also been shown to remove cell surface receptors through endocytosis. For example, endocytotic trafficking of AMPA receptors (AMPARs) during synaptic plasticity is thought to involve a PICK1/PKCα-dependent mechanism (for review, see Malenka, 2003). We used confocal microscopy to investigate whether UNC5A is internalized and colocalized with the endocytotic marker EEA-1 after PKCα activation. MycUNC5A-expressing hippocampal neurons were live-labeled with an antibody directed against the Myc tag, followed by TPA treatment for increasing periods of time. Before fixation, cells were acid-stripped (Carroll et al., 1999) to remove antibody bound to cell surface receptors; under these conditions, antibodies bound to internalized receptors are protected. Cells were then fixed and coimmunostained for UNC5A and the early endosomal marker EEA-1. At time 0, we detect little internalized or surface-expressed UNC5A (Fig. 2B), demonstrating that acid stripping efficiently removes antibody bound to the receptor at the cell surface. After 5 min of TPA treatment, internalized UNC5A begins to colocalize with EEA-1, indicating its presence in the early endosome (Fig. 2C). This colocalization diminishes over the time course of our experiment as the receptor moves to other endosomal compartments (Fig. 2D). Together with the data presented in Figure 1, these results support a mechanism whereby PKCα activation leads to removal of UNC5A receptor from the cell surface via an endocytotic mechanism.
UNC5A phosphorylation is required for PKCα-mediated internalization
Given that PKCα activity stimulates UNC5A internalization, it is likely that UNC5A is directly phosphorylated by PKCα. Because the intracellular domain (ICD) of UNC5A is phosphorylated by PKCα in vitro (Williams et al., 2003b), we analyzed the sequence using PhosphoBase for potential PKCα phosphorylation sites. This revealed five consensus PKCα phosphorylation motifs within the ICD of UNC5A, supporting a model in which activated PKCα, recruited to the receptor complex by PICK1, directly phosphorylates UNC5A. To test this, we generated an UNC5A receptor containing alanine substitutions at all five consensus PKCα sites [MycUNC5A(Δ5PKC)] and using two different assays determined whether these mutations affected internalization in response to PKCα activation.
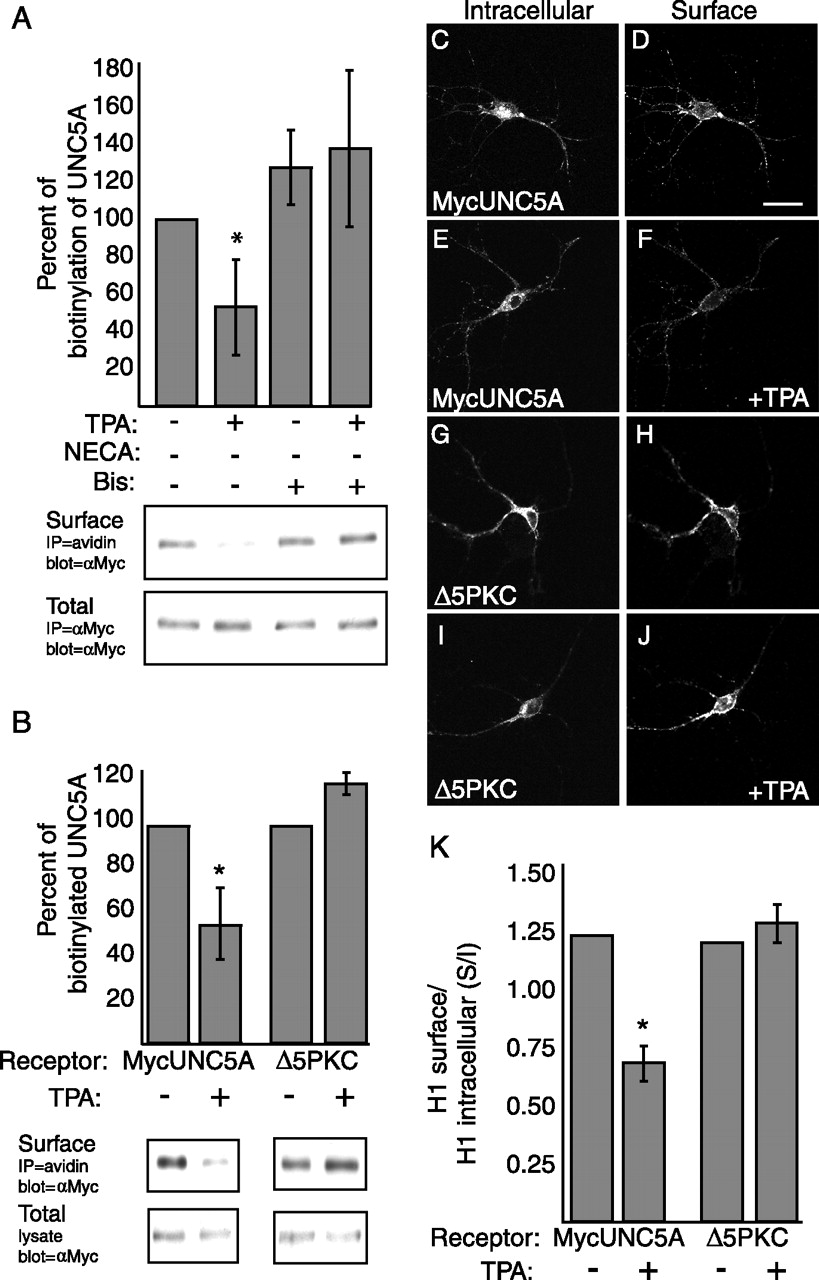
Cell surface biotinylation assays were performed and quantitated. To control for variation in expression levels between conditions, biotinylated MycUNC5A was normalized to 100% by comparing with total MycUNC5A expression. To verify the specific role of PKCα in UNC5A endocytosis, we treated the cultures with the PKC inhibitor, bisindolylmaleimide I, before analyzing the loss of wild-type UNC5A from the cell surface after PKC activation (Fig. 3A). We find that pretreating hippocampal cultures with Bis blocks the TPA effect on internalization, whereas Bis treatment alone has no effect. These data verify that PKCα plays a specific role in UNC5A internalization. This is further verified by analysis of the MycUNC5A(Δ5PKC) receptor construct. Biotinylation analysis reveals that MycUNC5A(Δ5PKC) receptors are biotinylated on the cell surface, indicating that they are expressed and trafficked appropriately to the cell surface, but fail to internalize in response to PKCα activation (Fig. 3B). In contrast, the surface expression of wild-type MycUNC5A decreases by ∼50% (Fig. 3B). We verified this result using an immunostaining assay to quantitate receptor internalization. Similar to the biotinylation assay, we controlled for variations in construct expression by determining the ratio of surface to intracellular (S/I) MycUNC5A staining (Fig. 3C–K). In this assay, cell surface receptors were live-labeled using a chicken anti-Myc antibody. Cultures were then fixed, permeabilized, and labeled with a rabbit anti-Myc antibody to identify intracellular receptors. The S/I ratios of TPA-treated cultures were compared with ratios observed in identically infected, untreated control neurons. Again, we observe that cell surface expression of MycUNC5A (Δ5PKC) does not decrease in response to the activation of PKCα, whereas expression of wild-type MycUNC5A is significantly reduced (Fig. 3K) (p < 0.001). These data indicate that phosphorylation of one or more sites by PKCα within the ICD of UNC5A are necessary for PKCα-mediated internalization.

Alanine substitution of all five consensus PKCα phosphorylation sites (Δ5PKC) in UNC5A results in a receptor that fails to internalize in response to PKCα activation. A, Twenty-four hours after infection with MycUNC5A, hippocampal neurons were incubated in the presence or absence of PBS (control) or TPA (100 ng/ml) as indicated to induce PKCα activation and receptor internalization. Bis (100 nm) treatments were performed for 2 h before treatment with PBS or TPA. The graph represents the mean ± SD of at least three separate experiments. For quantitation of the immunoblots, biotinylated receptor was normalized to total receptor expression from the same cultures to control for any variation in expression levels between conditions. The normalized results for each condition were then converted to a percentage of the untreated control condition, which was considered 100%. Graphs represent the mean ± SD of at least three experiments. B, Quantified biotinylation experiments on cells expressing MycUNC5A or MycUNC5A(Δ5PKC), in the presence and absence of TPA stimulation, as indicated. C–J, Quantitative immunofluorescence assays on hippocampal neurons show that TPA treatment results in internalization of MycUNC5A but not MycUNC5AΔ5PKC (compare F, J). Cells were infected with Sindbis viruses and TPA treated (100 ng/ml) as indicated to promote internalization. Intensity of live-labeled UNC5A receptors (C, E, G, I) was compared with the intracellular UNC5A receptor pool (D, F, H, J). The results were quantified by determining the total fluorescence intensity for both the surface-labeled and intracellular-labeled MycUNC5A over the same defined area and graphed as a ratio of S/I (K). The data represent the mean ± SEM S/I ratio. n ≥ 10 cells per condition; *p < 0.001, t test. Scale bar, 20 μm.
Serines 408 and 587 of UNC5A are required for PKCα-mediated internalization
To determine the specific phosphorylation site(s) necessary for PKCα-mediated internalization of UNC5A, we generated individual alanine substitutions at each of the five consensus PKCα sites. These mutants were tested for cell surface expression in response to PKCα activation using both surface biotinylation and immunostaining assays.
Immunostaining hippocampal neurons reveals that all receptors with single alanine substitutions are expressed at the cell surface (Fig. 4A), verifying that alanine substitutions do not disrupt processing and expression of the receptors. Figure 4A shows that the S/I ratio of hippocampal neurons expressing alanine substitutions at serines 408 and 587 (S408A and S587A) does not decrease in response to the activation of PKCα, compared with the wild-type MycUNC5A control, which exhibits a significant decrease (p < 0.001) in S/I in response to PKCα activation. Three other receptors containing alanine substitutions, S445A, S461A, and T646A, behave like wild-type receptors. Biotinylation confirms the results for the S408A and S587A receptors obtained using the immunostaining analysis. The S408A and S587A receptors fail to internalize in response to PKCα activation, compared with the wild-type MycUNC5A control, which exhibits a significant decrease in cell surface expression in response to PKCα activation (Fig. 4B) (p < 0.001). Because alanine substitution at either S408 or S587 prevents endocytosis, our findings suggest these sites act cooperatively to mediate PKCα-dependent internalization of UNC5A. This is consistent with a hierarchical order of phosphorylation in which phosphorylation of one site is permissive for phosphorylation of subsequent site(s). Such a mechanism is known to regulate the levels of Cdc25A protein degradation (Donzelli et al., 2004) and endocytosis of the δ-opioid receptor (Kouhen et al., 2000).

PKCα-mediated internalization of UNC5A is dependent on serines 408 and 587. Hippocampal neurons were infected with either wild-type MycUNC5A or MycUNC5A containing alanine substitutions at the indicated serine or threonine residues and then TPA treated (100 ng/ml) as indicated to induce receptor internalization. A, Results of quantitative immunofluorescence assays, comparing changes in the S/I ratios in response to PKCα activation. The data represent the mean ± SEM S/I ratio from ≥10 cells per condition and are graphed as total surface fluorescence intensity over total internal fluorescence intensity for a fixed area. B, Biotinylation results confirm that MycUNC5A S408A and S587A do not internalize in response to TPA treatment. Graph represents the mean ± SD, where n ≥ 3. *p < 0.001, t test.
Netrin-1-induced growth cone collapse is dependent on UNC5A surface expression
Our data show that cell surface levels of UNC5A are regulated by PKCα, raising the possibility that the response of a growth cone may depend on the activation state of PKCα. To address how PKCα-mediated changes in UNC5A surface expression affect netrin-1 receptor activity, we examined receptor-mediated growth cone collapse. We assessed growth cone collapse by infecting primary hippocampal neurons at 1 DIV with bicistronic virus expressing the MycUNC5A receptors, as indicated, and GFP (Fig. 5A–C). At 2 DIV, cells were treated with PBS, NTN1, or TPA followed by netrin-1 as indicated. Cells were then fixed, permeabilized, and immunostained for GFP, and randomly chosen growth cones were scored for collapse. A growth cone was considered collapsed if it had no lamellipodia and two or fewer filopodia. We previously reported that netrin-1 induced growth cone collapse of hippocampal neurons, which express only low levels of UNC5s endogenously, requires UNC5A overexpression. Pretreating these MycUNC5-expressing neurons with TPA to stimulate PKCα activation before exposure to netrin-1 downregulates UNC5A surface expression and results in reduced growth cone collapse (Fig. 5C) (Williams et al., 2003b). Using this assay, we tested hippocampal neurons expressing the UNC5A alanine substitution receptors for growth cone responsiveness to netrin-1 (Fig. 5).

Evaluation of the netrin-1 responsiveness of alanine-substituted UNC5A receptors. Hippocampal neurons (2 DIV), expressing the indicated MycUNC5A receptor constructs, were evaluated for growth cone collapse in the presence of NTN1. Growth cones were treated for 45 min with PBS (A), NTN1 (300 ng/ml; B), or TPA (100 ng/ml; C) followed by NTN1 (300 ng/ml) as indicated and then evaluated for growth cone collapse. The results are reported as the percentage of growth cones collapsed in which ≥50 growth cones were counted from at least three separate experiments. D, Examples of representative MycUNC5A-expressing growth cones before and after netrin-1 treatment. A growth cone was considered collapsed if F-actin staining contained no lamellipodia and two or fewer filopodia. E, Growth cone collapse in uninfected neurons was measured as a control for the treatment conditions. *p < 0.001, one-way ANOVA. Error bars represent SEM.
Neurons expressing the alanine-substituted receptors exhibit a level of collapse similar to wild-type expressing neurons in the absence and presence of netrin-1 (Fig. 5A,B, respectively). This behavior shows that alanine-substituted receptors are functionally responsive to netrin-1. Growth cones of hippocampal neurons expressing S445A, S461A, and T646A receptors exhibit growth cone collapse in the presence of netrin-1 and like wild-type expressing neurons, fail to collapse in the presence of netrin-1 when it follows TPA treatment (Fig. 5B,C) (p < 0.001). Given that we have been unable to detect any effect of netrin-1 on UNC5A expression (our unpublished observation), these results are consistent with our data showing these receptors are internalized as a consequence of PKCα activation. In contrast, growth cones of neurons expressing UNC5A(Δ5PKC), S408A, or S587A receptors collapse in the presence of netrin-1, even after PKCα activation (Fig. 5C), consistent with our data that these receptors fail to internalize after PKCα activation (Figs. 2, 3). TPA treatment, itself, has no affect on the behavior of growth cones because growth cones of uninfected hippocampal neurons incubated with TPA before netrin-1 treatment do not collapse (Fig. 5E).
These data highlight the importance of UNC5A endocytosis in regulating netrin-1-mediated responses and strongly suggest that PKCα is directly involved in this internalization. In contrast to growth cone response of neurons expressing wild-type UNC5A (Fig. 5), neurons expressing receptors that fail to internalize after PKCα activation [UNC5A(Δ5PKC), S408A, or S587A] (Figs. 3, 4) continue to exhibit netrin-1-mediated growth cone collapse even after TPA treatment (Fig. 5). Given that growth cone collapse is often used as an indicator of chemorepulsive response, these data suggest that endocytosis of UNC5A, which reduces the percentage of growth cones that collapse, will also convert netrin-1-mediated chemorepulsion to chemoattraction. It is interesting to note that growth cones expressing receptors that fail to internalize continue to exhibit netrin-1-induced collapse, indicating that endocytosis, per se, is not required for UNC5A-mediated responses. This is in contrast to semaphorin 3A (Sema3A)-mediated growth cone collapse, which requires neuropilin-1 (NP-1) internalization (Castellani et al., 2004), and recent findings that receptor tyrosine kinases signal from signaling endosomes after their internalization (for review, see Howe and Mobley, 2005). Together, these data demonstrate that PKCα-stimulated endocytosis of UNC5A efficiently alters the response of a growth cone to netrin-1 and extend our findings that serines 408 and 587 of UNC5A are necessary for PKCα-mediated endocytosis of the receptor.
Dileucine motifs are required for PKCα-mediated endocytosis of UNC5A
We have identified two consensus sites for PKCα-mediated phosphorylation that are necessary for UNC5A endocytosis (Figs. 4, 5), but it is unlikely that their phosphorylation is sufficient for internalization. Endocytosis of a receptor is a multistep process requiring association with proteins that interact with endocytotic machinery. For example, several receptor systems are internalized in a PKC- and clathrin-dependent manner (Ferrari et al., 1999; Hipkin et al., 2000; Xiang et al., 2001; Reinhardt et al., 2002; Srinivasan et al., 2002), with dileucine motifs serving as recognition sites for adaptor proteins that mediate the formation of clathrin-coated pits before endocytosis (for review, see Bonifacino and Traub, 2003). In most cases, the motifs that are required for endocytosis are located within close proximity of the transmembrane domain of the receptor. In UNC5A, serine 408 is within close proximity of the transmembrane domain as are two dileucine motifs at amino acids 419/420 and 455/456 that might serve as internalization motifs. These findings indicate there may be a requirement for dileucine motifs as well as direct PKCα phosphorylation of the receptor for endocytosis. To test this, we introduced alanine substitutions at leucines 419, 420 and 455, 456 (4L/A) within the MycUNC5A construct and tested this receptor for its ability to internalize after PKCα activation. Using both immunostaining (Fig. 6A–I) and biotinylation (Fig. 6J) assays to determine cell surface expression, we find that the MycUNC5A(4L/A) mutant fails to internalize in response to PKCα activation, indicating that dileucine motifs are necessary for PKCα-mediated endocytosis of UNC5A. In the growth cone collapse assay, neurons expressing MycUNC5A(4L/A) collapse in response to netrin-1, and pretreatment with TPA fails to reverse this activity (Fig. 6K), providing further evidence that one or both dileucine motifs are necessary for PKCα-dependent endocytosis of UNC5A. Interestingly, Bin-Amphiphysin-Rvs (BAR) domains, similar to the one found in PICK1, have been identified recently as modules involved in trafficking proteins between membrane domains, and in amphiphysin-2, the BAR domain is required to recruit and polymerize clathrin (Peter et al., 2004). This suggests that PICK1 could serve dual roles both by recruiting PKCα to UNC5A at the plasma membrane and by mediating interactions with the endocytotic machinery.

Mutation of two dileucine motifs in UNC5A results in a receptor that fails to internalize in response to PKCα activation. Hippocampal neurons were infected with Sindbis viruses encoding wild-type MycUNC5A receptor or a MycUNC5A receptor containing alanine substitutions at leucines 419, 420 and 455, 456 (4L/A). Neurons were TPA treated (100 ng/ml) as indicated, and both biotinylation and immunostaining assays assessed changes in receptor surface expression. A–I, Quantitative immunofluorescence assays. Intensity of live-labeled UNC5A receptors (B, D, F, H) was compared with intracellular UNC5A receptors (A, C, E, G). TPA treatment results in MycUNC5A internalization but no change in cell surface levels of MycUNC5A-4L/A (compare D, H). Scale bar, 20 μm. I, The results were quantified by determining the total fluorescence intensity for both the surface-labeled and intracellular-labeled MycUNC5A over the same defined area and graphed as a ratio of surface-to-intracellular fluorescence. The data represent the mean ± SEM S/I ratio. n ≥ 10 cells per condition; *p < 0.001, t test. J, Biotinylation data from MycUNC5A-4L/A-expressing neurons reveal no significant change in receptor surface expression after TPA treatment (100 ng/ml) compared with wild type. Representative immunoblots from cells treated as indicated are shown. K, Collapse assays demonstrate that PKCα activation does not attenuate netrin-1-induced (300 ng/ml) collapse in UNC5A-4L/A-expressing growth cones compared with wild-type MycUNC5A-expressing controls. *p < 0.001, t test. Error bars represent SEM.
UNC5A-mediated repulsion is converted to attraction by changes in UNC5A surface expression
Netrin-1 functions as a bidirectional guidance cue capable of attracting and repelling neurons and their growth cones. When the netrin-1 receptor DCC is expressed in the absence of UNC5A, the cell is attracted to netrin-1 (Keino-Masu et al., 1996; Fazeli et al., 1997). In contrast, a cell is repelled by netrin-1 when UNC5s form a netrin-1-dependent complex with DCC (Hong et al., 1999). To examine whether selective removal of UNC5A from the cell surface converts repulsion to attraction, we monitored the behavior of growth cones in a liquid gradient of netrin-1 in the presence and absence of bath application of TPA to stimulate endocytosis of UNC5A.
Similar to netrin-1-mediated axon guidance in Xenopus spinal neurons (Ming et al., 1997), growth cones of uninfected rat hippocampal neurons exhibit positive turning angles, indicating attraction to a microscopic gradient of netrin-1 (Fig. 7A, D–F). This attraction requires DCC as a DCC function-blocking antibody abolishes the response (Fig. 7 B,D–F). In addition, repulsion in a netrin-1 gradient requires expression of UNC5A as growth cones expressing wild-type MycUNC5A exhibit negative turning angles in a netrin-1 gradient (Fig. 7C–F) (p < 0.01). Adding TPA to the media during continued netrin-1 treatment reverses this UNC5A-dependent repulsion, converting it to attraction (Fig. 7G–I) (p < 0.05). At the 70 min time point, axons have redirected toward the netrin-1 source, indicating that internalization of UNC5A by PKCα activation reverses the functional response of axons to netrin-1. The effect of PKCα activation is specific to UNC5A-expressing neurons, because netrin-1-mediated chemoattraction of uninfected neurons is not altered by TPA addition (Fig. 7D,E). Together with the results of our growth cone collapse assays, in which PKCα-mediated internalization of UNC5A reverses growth cone responses to netrin-1 (Fig. 5), these results indicate that axon repulsion is converted to attraction in a netrin-1 gradient by selectively removing UNC5A from the cell surface. The activation of PKCα leads to the internalization of UNC5A (Fig. 2), whereas DCC remains on the cell surface to mediate attraction toward the netrin-1 source (Fig. 7E,G) (Williams et al., 2003b). Interestingly, a recent report demonstrated that treatment of growth cones with 150 ng/ml netrin-1 causes rapid desensitization and endocytosis of DCC (Piper et al., 2005), but this apparently does not occur at the very low netrin-1 concentrations present at the leading edge of the soluble netrin-1 gradient. This concentration is calculated to be ∼5 ng/ml of netrin-1 based on a similar method delivering a soluble gradient of dibutyryl cAMP (Lohof et al., 1992). In support, we observe that netrin-1-dependent attractive turning is continuous in a netrin-1 gradient for up to 75 min in uninfected hippocampal neurons (our unpublished observation), making it unlikely that endocytosis of DCC is playing a role in these assays. Together, these data support the hypothesis that PKCα-mediated internalization of UNC5A is an important mechanism for efficiently removing receptors from the cell surface, thus regulating the functional response of a growth cone to netrin-1.

Turning of hippocampal growth cones in response to a netrin-1 gradient, UNC5A overexpression, and TPA treatment. A, Images of wild-type growth cone at onset (left) and end (right) of a 30 min exposure to a soluble NTN1 gradient (see Materials and Methods for gradient conditions). B, Neuron cultures were incubated with DCC function-blocking antibody 30 min before gradient onset. C, Neurons overexpressing UNC5A at the cell surface are repelled from the netrin-1 gradient. Scale bar, 10 μm. D, Traces of individual growth cone trajectories at 30 min exposure in PBS [control (CTRL)], NTN1, NTN1 with anti-DCC pretreatment, MycUNC5A overexpression in PBS, MycUNC5A overexpression in NTN1, NTN1 with TPA pretreatment. Overexpressed cell surface receptors, antibody preincubations, or TPA pretreatments are indicated at the bottom and gradient condition at the top. E, The average turning angle in degrees (top) and average net neurite extension in micrometers (bottom) for all neurons tested. Overexpressed cell surface receptors, antibody preincubations, or TPA-pretreatments are indicated at the bottom and gradient condition at the top. The asterisk indicates statistically significant difference from control; *p < 0.01, Mann–Whitney test. Scale bar, 10 μm. F, The distribution of all turning angles, plotted at the end of 30 min exposure to netrin-1 gradient. The percentage value refers to the percentage of growth cones with turning angles less than or equal to a given angular value. Positive turning angles correspond to attraction toward the netrin-1 point source. G, Hippocampal growth cone expressing UNC5A on the cell surface at the onset of a netrin-1 gradient and after 30 min (turn 1). After 30 min, the position of netrin-1-containing pipette (5 μg/ml) with respect to the growth cone was reset to time 0 parameters, TPA was added to the bath (100 ng/ml) to induce UNC5A endocytosis, and growth cone was imaged again at 60 min (turn 2). H, Traces of individual growth cone trajectories at 60 min. The large arrows indicate repositioning of the netrin-1 point source at 30 min. The small arrows indicate point along trajectory at which time TPA was added to the bath. I, The average turning angle in degrees (left) and the net neurite extension in micrometers (right). Negative turning angles indicate repulsion, whereas positive turning angles indicate attraction to the netrin-1 point source. **p < 0.05, Mann–Whitney test. Error bars represent SEM.
UNC5A-dependent chemorepulsion of cerebellar granule cells in a netrin-1 gradient is converted to attraction in a PICK1/PKCα-dependent manner
Thus far, we have used overexpression to study UNC5A trafficking. To investigate whether repulsion is converted to attraction by PKCα activation in neurons expressing endogenous levels of UNC5, we used primary cultures of postnatal, murine, cerebellar granule cells (P4–P6). It has been shown that netrin-1 repels both the parallel fibers and granule cells that grow out of the external germinal layer of postnatal cerebellar explants (Alcantara et al., 2000). Consistent with these data, we find that wild-type cerebellar granule cell axons exhibit chemorepulsion in a netrin-1 gradient (Fig. 8A,E,F), suggesting an UNC5-dependent axon response. To examine whether PKCα activation reverses this response, cultured granule cells were treated with TPA before exposure to the netrin-1 gradient (Fig. 8A,E,F). As observed in UNC5A-expressing rat hippocampal neurons (Fig. 7G–I), activation of PKCα in native granule cells converts chemorepulsion to chemoattraction in a netrin-1 gradient (Fig. 8B,E,F) (p < 0.01). Together, these data suggest that granule cells use a PICK1/PKCα-dependent mechanism to remove UNC5A from the cell surface, which, in turn, alters responses to netrin-1.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Granule cells from P6 cerebellum are repelled by netrin-1 stable gradients. A–D, Representative images of granule cells at time 0 and 30 min after exposure to NTN1 gradient. Wild-type granule cells are repelled from the NTN1 point source (A), and pretreatment with 100 ng/ml TPA for 30 min before NTN1 gradient onset converts the turning response to attraction (B). Granule cells cultured from Unc5a −/− animals are attracted toward the NTN1 point source (C). In contrast, granule cells cultured from PICK1−/− mice are repelled by the NTN1 gradient and this response is not converted to attraction after TPA treatment (D). E, Composite traces from all granule cells subjected to 30 min NTN1 gradients with genotype, TPA pretreatment, and scale bars as indicated. F, The average turning angle in degrees (top) and average net neurite extension in micrometers (bottom) for all granule cells assayed. G, The distribution of granule cell turning angles. Positive turning angles indicate attraction to the netrin-1 point source, and negative turning angles indicate netrin-1 repulsion. *p < 0.01 Mann–Whitney test. Error bars represent SEM. CTRL, Control.
Studies have shown that Unc5a–d are expressed in the rodent cerebellum (Leonardo et al., 1997; Alcantara et al., 2000; Zhong et al., 2004). Of these, only Unc5a–c are known to be expressed in mouse granule cells (Leonardo et al., 1997; Alcantara et al., 2000), whereas Unc5d has been shown to be expressed in Purkinje cells in rats (Zhong et al., 2004). To confirm the requirement for Unc5a in netrin-1-mediated chemorepulsion of cerebellar granule cell axons, we analyzed axonal turning responses of neurons derived from Unc5a-null mice (Unc5a−/−) (Fig. 8C,E,F). Unlike the repulsive turning observed when wild-type cerebellar granule cells are assayed, Unc5a−/− axons are attracted to netrin-1, demonstrating a requirement for UNC5A in netrin-1-mediated chemorepulsion. This result also suggests that the loss of UNC5A surface expression alone via a PICK/PKCα-dependent mechanism is sufficient to convert netrin-1-mediated responses. Although these data show a requirement for UNC5A in netrin-1 dependent chemorepulsion, a role for UNC5B or UNC5C in this process has not been ruled out. To confirm the requirement for PICK1 in mediating this directional conversion, we also examined whether granule cell axons from PICK1 null (PICK1−/−) mice alter their response to netrin-1 in the axon turning assay (Fig. 8D–F) (Gardner et al., 2005). Like wild-type cerebellar granule cell axons, PICK1−/− granule cell axons exhibit chemorepulsion in a soluble netrin-1 gradient. Unlike wild-type cerebellar granule cell axons, however, activation of PKCα fails to convert their response to chemoattraction (Fig. 8D–F), and instead, these neurons continue to be repelled. Together, these data definitively demonstrate that PICK1 is required for the TPA-stimulated conversion of UNC5A-mediated repulsion to attraction in a netrin-1 gradient.
Discussion
Here, we demonstrate that PKCα activation leads to UNC5A endocytosis, resulting in altered migratory responses to netrin-1. Investigation into the mechanisms underlying UNC5A endocytosis reveals that internalization depends on two PKCα phosphorylation sites and dileucine motifs, which likely serve as internalization signals. Our functional assays demonstrate that the response of a developing axon to netrin-1 is dependent on the presence or absence of UNC5A. We show that UNC5A-mediated chemorepulsion is converted to chemoattraction by PKCα activation, leading to the removal of UNC5A, but not DCC (Williams et al., 2003b), from the cell surface. Additionally, we demonstrate a requirement for PICK1 by showing that axons of PICK1-null neurons continue to exhibit chemorepulsion after PKCα activation. Together, these studies show that regulated trafficking of guidance receptors provides a mechanism to modulate the response of a migrating growth cone to netrin-1.
A role for second messenger signaling in modulating neuronal responses to netrin-1
There is a wealth of data describing how activation of second messenger pathways alters neuronal responses to netrin-1. For example, Xenopus spinal neurons are attracted in a netrin-1 gradient by increases in cAMP levels, PKA activity (Ming et al., 1997), and intracellular calcium (Hong et al., 2000). This attraction can be blocked by inhibiting numerous signaling pathways such as PI3-kinase, PLC-γ (Ming et al., 1999), MAPK (Ming et al., 1999; Forcet et al., 2002; Campbell and Holt, 2003), and caspase-3 (Campbell and Holt, 2003). Moreover, modulating the ratio of cAMP/cGMP alters responses to netrin-1, with higher ratios favoring attraction and lower ratios favoring repulsion (Nishiyama et al., 2003). Thus, many signal transduction pathways modulate the way that a neuron reacts in a netrin-1 gradient, but the cellular events that must be initiated to alter the trajectory of a growth cone are not well understood.
One appealing hypothesis is that changes in second messenger activity regulate cell surface levels of netrin-1 receptor. These changes in receptor levels subsequently alter receptor-linked signaling events that direct attractive or repulsive responses to a guidance cue. We show that PKCα activation converts netrin-1-mediated chemorepulsion to chemoattraction by removing repellent UNC5A receptors from the cell surface. We hypothesize that this may be a general mechanism and that multiple intracellular signaling pathways may feedback to regulate the expression of receptors at the cell surface. For example, the requirement for PLC-γ activity in netrin-1-dependent chemoattraction of Xenopus spinal neurons may be caused, at least in part, by its activation of PKCα and the subsequent endocytosis of UNC5. This model may also explain how high intracellular cAMP levels favor attraction of Xenopus spinal neurons in a netrin-1 gradient (Ming et al., 1997), because it was recently shown that high levels of cAMP release DCC from intracellular pools, leading to increased cell surface expression (Bouchard et al., 2004). Together, these data suggest that some signaling pathways regulate the way neurons respond to netrin-1 by modulating cell surface expression of its receptors. More importantly, they provide a framework for thinking about previous studies on the effects of second messenger signaling on neuronal responses to guidance cues.
The potential role for G-protein-coupled receptor signaling in regulating netrin-1 activity
Although there is a growing understanding of how these signaling pathways alter growth cone responses to guidance cues, it remains unclear how these pathways are regulated in the organism during development. Seven transmembrane, G-protein-coupled receptors (GPCRs) are emerging as key modulators of second messenger pathways controlling the response of axons to a variety of guidance cues. For example, both glutamate and the chemokine stromal cell-derived factor-1 signal through GPCRs to modulate cAMP and Rho activity, leading to alterations in the repulsive response of growth cones to Slits and Semaphorins (Chalasani et al., 2003; Kreibich et al., 2004). For netrin-1, potential GPCR candidates are the adenosine receptors, which are known to regulate multiple signaling pathways including PKC and cAMP (Feoktistov and Biaggioni, 1998). The adenosine receptor family member A2bR has been shown to bind netrin-1 and play a role in netrin-1-mediated cAMP production and commissural axon outgrowth (Corset et al., 2000). Moreover, it has been shown that, in a single trajectory, retinal growth cones modify their response to netrin-1 from chemoattraction to chemorepulsion with a corresponding reduction in the levels A2bR, DCC, and cAMP (Shewan et al., 2002). An adenosine receptor agonist prevents this switch, suggesting that the activity of A2bR modifies the response of these axons, potentially by regulating surface levels of receptors (Shewan et al., 2002).
The potential role of receptor recycling in regulating netrin-1-mediated activity
Although our data indicate that endocytosis of UNC5A converts netrin-1-mediated chemorepulsion to chemoattraction, it does not address whether UNC5A is delivered to the cell surface to convert the response back again. One possible mechanism is that endocytosed UNC5 receptors, rather than being degraded, are recycled to the cell surface via recycling endosomes. Such a mechanism occurs for the AMPARs during long-term potentiation (LTP). AMPARs are endocytosed and sorted into recycling endosomes, which rapidly supply these receptors to the postsynaptic membrane during LTP (Park et al., 2004). Like UNC5A, the endocytotic trafficking of AMPARs involves PICK1/PKCα-dependent receptor phosphorylation (for review, see Malenka, 2003). Our data show that UNC5A undergoes a similar regulated endocytosis after PKCα activation, colocalizing with EEA-1. It is possible that UNC5A may share additional aspects of the endocytotic mechanism with AMPARs. Although additional studies are required to determine the fate of endocytosed UNC5 receptors, the similarities between AMPAR and UNC5 trafficking suggest that endocytosed UNC5A receptors enter the recycling endosome pathway. In this way, UNC5A receptors may be stored intracellularly, allowing them to be quickly mobilized to the cell surface to mediate chemorepulsion.
A general mechanism for regulating axon guidance cue responses
Accumulating evidence supports the hypothesis that altering levels of axon guidance receptors at the cell surface is a general mechanism for regulating the response of axons to their cues. For example, regulating cell surface levels of the Robo receptor controls the response of growth cones to the repulsive guidance cue Slit. Although the mechanism is still unclear, the data show that the sorting protein Comm regulates Robo surface expression (Keleman et al., 2002, 2005; Myat et al., 2002). Because Slit is expressed at the Drosophila midline during development, the loss of surface Robo expression allows axons, which would otherwise be repelled by Slit, to cross the midline. After crossing the midline, Robo expression is upregulated to prevent axons from recrossing. Thus, in the case of Robo, regulated changes in surface levels alter the response of a developing axon to Slit.
There is also evidence that endocytosis may be required for directly mediating, as well as altering, responses to other axon guidance cues. Recent findings point to a mechanism by which the cell adhesion molecule L1 is involved in mediating Sema3A activity through an interaction and coendocytosis with its receptor NP-1. Sema3A binding of NP-1 induces internalization of the NP-1/L1 complex through an L1- and clathrin-dependent mechanism (Kamiguchi and Lemmon, 2000; Castellani et al., 2004), and this cointernalization is required for Sema3A-induced endocytosis and chemorepulsion (Castellani et al., 2004). Similarly, endocytosis of the ephrin/Eph complex initiates ephrin-mediated growth cone collapse, switching activity of the complex from cell adhesion to repulsion. Ephrin-mediated growth cone collapse is dependent on Eph-mediated activation of Vav2 (Cowan et al., 2005) and ephexin1 (Shamah et al., 2001; Sahin et al., 2005), with Vav2 involved in Rac-dependent endocytosis of the ephrin–Eph complex (Cowan et al., 2005) and ephexin1 involved in RhoA-dependent changes in F-actin assembly and contractility (Shamah et al., 2001; Sahin et al., 2005). It has been suggested that guidance cue receptor internalization may play a mechanical role in axon guidance by removing cell membrane causing localized growth cone collapse, which in turn leads to repulsion (Castellani et al., 2004), but currently, it is unclear whether there is a causal relationship between receptor endocytosis and repulsive signaling. Although a mechanical mechanism differs from the instructive role that receptor trafficking plays in netrin-1- and Slit-mediated guidance, together these examples highlight the importance of regulating cell surface levels of guidance receptors on the growth cone.
In sum, our data suggest that altering the levels of axon guidance receptors on the cell surface is one mechanism by which developing neurons adjust responsiveness to guidance cues. Although it is possible that activated PKCα can alter the direction of growth cone migration by changing intracellular signaling mechanisms downstream of netrin-1, here we delineate a mechanism by which it directly participates in converting repulsion to attraction. In our model, PKCα, after activation, is recruited, through its adaptor PICK1, to UNC5A receptors on the growth cone surface where it phosphorylates UNC5A at two sites. This leads to the internalization of UNC5A and the conversion of netrin-1-mediated repulsion to attraction. Thus, the data reveal a mechanism that allows navigating growth cones to rapidly adjust their sensitivity to guidance cues as they traverse complex terrain.
Footnotes
- Received August 16, 2005.
- Revision received February 2, 2006.
- Accepted February 3, 2006.
-
This work was supported by National Institutes of Health Grants NS39572-01 (L.H.) and AG22903-01 (J.L.B.). We thank David Feldheim for Sindbis virus reagents, Marc Tessier-Lavigne and Xiaowei Lu for Unc5a−/− mice, and Yishi Jin for use of her confocal microscope.
-
*J.L.B. and W.L.M. contributed equally to this work.
-
J. L. Bartoe's present address: Department of Biology, Providence College, Providence, RI 02918.
-
J. Xia's present address: Department of Biochemistry, Hong Kong University of Science and Technology, Clear Water Bay, Kowloon Hong Kong.
- Correspondence should be addressed to Lindsay Hinck at the above address. Email: hinck{at}biology.ucsc.edu