
Abstract
Endosomal membrane trafficking in dendritic spines is important for proper synaptic function and plasticity. However, little is known about the molecular identity and functional compartmentalization of the membrane trafficking machinery operating at the postsynaptic terminal. Here we report that the transport of AMPA-type glutamate receptors into synapses occurs in two discrete steps, and we identify the specific endosomal functions that control this process during long-term potentiation. We found that Rab11-dependent endosomes translocate AMPA receptors from the dendritic shaft into spines. Subsequently, an additional endosomal trafficking step, controlled by Rab8, drives receptor insertion into the synaptic membrane. Separate from this receptor delivery route, we show that Rab4 mediates a constitutive endosomal recycling within the spine. This Rab4-dependent cycling is critical for maintaining spine size but does not influence receptor transport. Therefore, our data reveal a highly compartmentalized endosomal network within the spine and identify the molecular components and functional organization of the membrane organelles that mediate AMPA receptor synaptic delivery during plasticity.
- Rab proteins
- endosomes
- synaptic plasticity
- dendritic spine
- hippocampus
- GluR1
- phosphorylation
Introduction
Intracellular membrane trafficking is an essential process in all eukaryotic cells, but it is particularly critical at synaptic terminals, in which a large number of specific ion channels, scaffolding molecules, and a variety of signal transduction modulators have to be precisely targeted to ensure proper synaptic function (McGee and Bredt, 2003; Sudhof, 2004; Kennedy and Ehlers, 2006). At excitatory synapses in the CNS, AMPA-type glutamate receptors (AMPARs) are responsible for most synaptic transmission. Nevertheless, these receptors are not permanent residents of the synapse. AMPARs can be inserted or removed from the postsynaptic membrane in an activity-dependent manner, leading to long-lasting changes in synaptic strength, such as long-term potentiation (LTP) and long-term depression (Barry and Ziff, 2002; Malinow and Malenka, 2002; Bredt and Nicoll, 2003; Esteban, 2003; Sheng and Hyoung Lee, 2003). These forms of synaptic plasticity are now widely accepted as cellular models of learning and memory processes (Bliss and Collingridge, 1993; Gruart et al., 2006; Pastalkova et al., 2006; Whitlock et al., 2006). In addition to this regulated trafficking, AMPARs cycle in and out of the synaptic membrane in a constitutive, activity-independent manner, which requires NSF (N-ethylmaleimide-sensitive factor) (Nishimune et al., 1998; Song et al., 1998; Luscher et al., 1999), Hsp90 (heat shock protein) (Gerges et al., 2004b), and NEEP21 (neuron-enriched endosomal protein 21 kDa) (Steiner et al., 2005) function.
This dynamic behavior suggests a complicated flow of AMPAR-bearing membrane organelles at the postsynaptic terminal. Indeed, we and others have recently demonstrated that endosomal trafficking is important for the synaptic delivery of AMPARs (Gerges et al., 2004a; Park et al., 2004). Nevertheless, the molecular identity of the intracellular organelles that are responsible for receptor transport at synapses is virtually unknown. In addition, it remains to be elucidated whether these organelles display specific compartmentalization at dendritic spines, the small protrusions in which most excitatory synapses are formed (Harris and Kater, 1994).
In this study, we examined the functional and molecular organization of the endosomal compartments that mediate the synaptic trafficking of AMPARs during LTP. To this end, we investigated the role of Rab proteins in this process, because this family of small GTPases are well known organizers of intracellular membrane sorting in eukaryotic cells (Pfeffer, 2001; Zerial and McBride, 2001). Using a combination of molecular biology, electrophysiology, and fluorescence imaging, we identified two distinct endosomal trafficking functions that separately handle (1) AMPAR translocation into the spine, mediated by Rab11, and (2) final delivery into the synaptic membrane, mediated by Rab8. These results constitute a functional and molecular dissection of the local endosomal network that operates at the postsynaptic terminal during plasticity.
Materials and Methods
Cloning and expression of recombinant proteins in hippocampal slices.
The green fluorescent protein (GFP)-tagged glutamate receptor subunit 1 (GluR1) construct and the truncated calcium/calmodulin-dependent kinase II (CaMKII) construct were made as described previously (Shi et al., 2001). Red fluorescence protein (RFP) is a tandem-dimer variant of DsRed (Discosoma red) (Campbell et al., 2002). All constructs were recloned in pSinRep5 for expression using Sindbis virus (Malinow et al., 1999) or in mammalian expression plasmids for biolistic delivery (Lo et al., 1994). Recombinant proteins were expressed in hippocampal CA1 pyramidal neurons from organotypic slice cultures (Gahwiler et al., 1997; Gerges et al., 2005), as indicated in each experiment. Slices were prepared from postnatal day 5–7 rats and cultured from 5 to 10 d. Expression of the recombinant proteins was for a minimum of 15 h, or for 36–48 h when expressing recombinant AMPAR subunits. All biosafety procedures and animal care protocols were approved by the University of Michigan Committee on Use and Care of Animals.
Biochemistry.
Hippocampal extracts were prepared from organotypic slice cultures in homogenization buffer containing 10 mm HEPES, 0.5 m NaCl, 10 mm NaF, 1 μm microcystin LR, 10 mm EDTA, 0.1 mm PMSF, 1% Triton X-100, and a protease inhibitor mixture (2 μg/ml) containing chemostatin, leupeptin, antipain, and pepstatin. Western blot analyses were performed with anti-GluR1 (Chemicon, Temecula, CA), anti-phospho-GluR1 [P-S831 and P-S845; Upstate (Charlottesville, VA) and Chemicon], and anti-GFP (Roche Diagnostics, Indianapolis, IN) antibodies. Western blots were developed with chemiluminescence (SuperSignal Kit; Pierce, Rockford, IL) and quantified by densitometric scanning under linear exposure conditions.
Confocal fluorescence and immunohistochemistry.
Fluorescence images were acquired with an Olympus Optical (Tokyo, Japan) FV500 confocal microscope using Fluoview software. Image analysis was performed as described previously (Gerges et al., 2005).
Electrophysiology.
Whole-cell recordings were obtained from infected or uninfected CA1 pyramidal neurons, using fluorescence and transmitted light illumination. The recording chamber was perfused with 119 mm NaCl, 2.5 mm KCl, 4 mm CaCl2, 4 mm MgCl2, 26 mm NaHCO3, 1 mm NaH2PO4, 11 mm glucose, 0.1 mm picrotoxin, 10 μm bicuculline, and 2 μm 2-chloroadenosine, at pH 7.4, gassed with 5% CO2/95% O2. Patch recording pipettes (3–6 MΩ) were filled with 115 mm cesium methanesulfonate, 20 mm CsCl, 10 mm HEPES, 2.5 mm MgCl2, 4 mm Na2ATP, 0.4 mm Na3GTP, 10 mm sodium phosphocreatine, and 0.6 mm EGTA at pH 7.25. Voltage-clamp whole-cell recordings were performed with multiclamp 700A amplifiers (Molecular Devices, Palo Alto, CA). Synaptic responses were evoked with two bipolar electrodes with single voltage pulses (200 μs, up to 20 V). The stimulating electrodes were placed over Schaffer collateral fibers between 300 and 500 μm from the recorded cells. Synaptic AMPA receptor-mediated responses were measured at −60 mV. LTP experiments were performed by pairing 0 mV postsynaptic depolarization with 3 Hz presynaptic stimulation (300 pulses). In these experiments, one of the stimulating electrodes was turned off during postsynaptic depolarization and therefore used as control (unpaired) pathway.
Statistical analysis.
All graphs represent average ± SEM values. Statistical differences were calculated according to nonparametric tests. Comparisons between multiple groups were performed with the Kruskal–Wallis ANOVA. When significant differences were observed, p values for pairwise comparisons were calculated according to two-tailed Mann–Whitney tests. Comparisons between cumulative distributions (see Fig. 2c) were performed according to two-sample Kolmogorov–Smirnov tests. p values are indicated in each figure. In addition, *p ≤ 0.05 and **p ≤ 0.01. n represents the number of cells in electrophysiological recordings and the number of spine–dendrite pairs in imaging experiments.
Results
Rab proteins involved in LTP
Endosomal trafficking has been proposed recently to mediate the synaptic delivery of AMPARs during LTP induction (Gerges et al., 2004a; Park et al., 2004). Nevertheless, endosomes are very heterogeneous and dynamic structures (Gruenberg, 2001; van Ijzendoorn, 2006). To test potential multiple routes for AMPAR synaptic delivery during plasticity, we evaluated the role of Rab4, Rab8, Rab11, and Rab5 in LTP. Among these proteins, Rab4, Rab8, and Rab11 are associated to exocytic (membrane delivery) routes (Huber et al., 1993; Sonnichsen et al., 2000), whereas Rab5 is well known to mediate endocytic trafficking (Bucci et al., 1992). Of these Rab proteins, only Rab8 appears to be involved in basal AMPAR synaptic transmission (Gerges et al., 2004a; Brown et al., 2005).
We performed pairing-induced LTP experiments in CA1 neurons from organotypic hippocampal slice cultures expressing dominant-negative mutants (dn) of Rab5, Rab4, Rab11, or Rab8 or uninfected (control) neurons (see Materials and Methods). Of note, these dominant-negative mutants have been abundantly used in previous studies to knockdown specific Rab-dependent membrane trafficking steps (for recent examples, see Emery et al., 2005; Hoepfner et al., 2005; Smith et al., 2007). In addition, this approach provides a more acute blockade of endogenous functions compared with standard RNA interference techniques (15 h in our experiments vs several days in typical RNA interference approaches). These experiments were performed blind, that is, the person performing the electrophysiological recordings and the analysis did not know which protein was being expressed.
As shown in Figure 1, a and b (black symbols), uninfected neurons displayed a significant potentiation of synaptic transmission (p = 0.003), relative to the unpaired pathway that did not receive LTP-inducing stimuli. In contrast, Rab8dn and Rab11dn (blue and red symbols, respectively) virtually abolished synaptic potentiation. Importantly, Rab4dn and Rab5dn (green and gray symbols, respectively) did not significantly alter LTP, indicating that the effects of Rab8dn and Rab11dn are not attributable to virus infection or nonspecific sequestration of regulatory proteins, such as GDP/GTP exchange factors or GDP dissociation inhibitors. Additionally, the stability of the control pathway, which did not receive LTP-inducing stimuli, was not altered by any of these Rabdn proteins (Fig. 1b). As mentioned previously, Rab8dn does depress basal synaptic transmission (Gerges et al., 2004a), but this basal depression does not change during the course of the experiment. The other Rabdn proteins tested here (Rab4dn, Rab11dn, and Rab5dn) do not alter basal synaptic transmission (Gerges et al., 2004a; Brown et al., 2005). Finally, none of these dominant-negative proteins altered passive membrane properties of the infected cell (input resistance and holding current) (Fig. 1c,d).

Rab8 and Rab11 are strictly required for LTP. a, Organotypic slice cultures were infected with viruses expressing Rab4dn (green), Rab11dn (red), Rab8dn (blue), or Rab5dn (gray). Whole-cell recordings were established from neurons expressing the Rab protein dominant-negative or uninfected cells. LTP was induced by pairing 3 Hz presynaptic stimulation with 0 mV postsynaptic depolarization (black arrow), as described in Materials and Methods. Inset, Sample trace of evoked AMPA receptor-mediated synaptic responses recorded at −60 mV before pairing (thin line) and 30 min after pairing (thick line) from control (uninfected) neurons or Rabdn-infected cells, as indicated. b, Normalized average potentiation of AMPAR-mediated responses collected between 30 and 35 min after LTP induction in paired (LTP) and unpaired (Control) pathways from neurons expressing different Rabdn constructs or uninfected, as indicated. Error bars represent SEM in all figures. n represents number of cells. c, d, Average values of input resistance (c) and holding current (d) from the LTP recordings shown in a and b.
The fact that both Rab8dn and Rab11dn fully block LTP suggests that these two Rab proteins participate in a common obligatory route for AMPAR synaptic delivery.
Rab8 and Rab11 mediate separate anatomical steps in the synaptic delivery of AMPA receptors
Our results shown above suggest that AMPARs depend on both Rab8- and Rab11-driven endosomal populations to reach their synaptic targets. To test whether these Rab proteins are involved in different morphological steps during the activity-dependent delivery of AMPARs, we monitored the regulated trafficking of GFP-tagged GluR1 into dendritic spines while blocking specific Rab-dependent endosomal transport using different dominant-negative mutants.
We used biolistic gene-gun delivery to coexpress GFP–GluR1 with constitutively active CaMKII (tCaMKII) for 36 h in hippocampal slices (see Materials and Methods). These conditions mimic LTP and drive the transport of GluR1 into spines (Piccini and Malinow, 2002) and synapses (Hayashi et al., 2000). GFP–GluR1 and tCaMKII were cotransfected with RFP-tagged Rab8dn, Rab11dn, Rab4dn, or cytosolic RFP, as control. To assess local GFP–GluR1 delivery to CA1 spines, we quantified GFP accumulation at spine heads and adjacent dendritic shafts using confocal microscopy and line plots of fluorescence intensity (Gerges et al., 2005). We then compared the average spine/dendrite ratios of GFP–GluR1 for the different conditions. The image analysis was performed blind with respect to the RFP-tagged protein being expressed. As shown in Figure 2, Rab8dn and Rab11dn produced opposite effects on GluR1 spine/dendrite distribution. Coexpression of Rab11dn significantly reduced the amount of GFP–GluR1 reaching the spine head (Fig. 2b, red column, c, left shift of spine/dendrite ratios). In contrast, Rab8dn led to a marked accumulation (almost doubling) of GFP–GluR1 at spines compared with the adjacent dendritic shaft (Fig. 2b, blue column, c, right shift of spine/dendrite ratios). This result is remarkable, considering that Rab8dn blocks LTP expression (Fig. 1a,b) and depresses basal synaptic transmission (Gerges et al., 2004a). Rab4dn did not significantly alter the regulated delivery of GFP–GluR1 into spines compared with cytosolic RFP expression.

Rab protein requirement for the regulated trafficking of GluR1 into spines. a, Representative confocal images of a spine and the adjacent dendritic shaft from neurons transfected with GFP–GluR1 and tCaMKII plus RFP (top) or Rabdn–RFP constructs (bottom 3 panels), as indicated. GFP fluorescence signal (green) represents GluR1 partition between spine and dendrite. Scale bars, 1 μm. b, Average spine/dendrite ratio of GFP fluorescence intensities across spine–dendrite pairs from neurons transfected with GFP–GluR1 and tCaMKII plus RFP or Rabdn–RFP constructs, as indicated. GFP–GluR1 accumulation was calculated as described in Materials and Methods. n represents the number of spine–dendrite pairs from six different neurons per condition. c, Cumulative probability distributions of spine/dendrite ratios from the same data plotted in b.
These results reveal that, although both Rab8 and Rab11 are required for LTP, they mediate spatially separated trafficking steps. Specifically, these data suggest that GluR1 is sequentially transported through Rab11-driven endosomes for its regulated translocation from dendrites into spines and then through Rab8-driven compartments for its functional insertion into synaptic membranes. Of note, the differing phenotypes of Rab4dn, Rab8dn, and Rab11dn confirm the suitability of these dominant-negative proteins to interfere with specific endosomal functions.
Rab4-dependent membrane trafficking is critical for spine size maintenance
It has been reported recently that membrane trafficking from recycling endosomes is required for the maintenance of spines and their activity-dependent growth (Park et al., 2006). Given our results on the spatial separation of Rab8- and Rab11-dependent trafficking, we decided to evaluate the role of these Rab proteins in the maintenance of spine size. For these experiments, we coexpressed RFP-tagged Rab dominant-negative mutants with enhanced GFP, as a marker for spine volume. Image acquisition and analysis was performed blind.
As shown in Figure 3 and in marked contrast with our results on GluR1 distribution, Rab4dn produced the most drastic reduction in spine size. Rab8dn and Rab11dn led to more moderate spine shrinkage. Rab5dn did not produce any significant change in spine size. These results indicate that multiple endosomal functions participate in basal membrane trafficking at the spine, Rab4-dependent processes being the most critical for the maintenance of spine size. These data also suggest that the synaptic delivery of AMPARs and the recycling of bulk membrane within the spine are mediated by different endosomal populations, because Rab4dn did not significantly alter LTP or GluR1 distribution despite its drastic effect on spine size. Finally, none of these Rabdn proteins affected spine length (Fig. 3c).
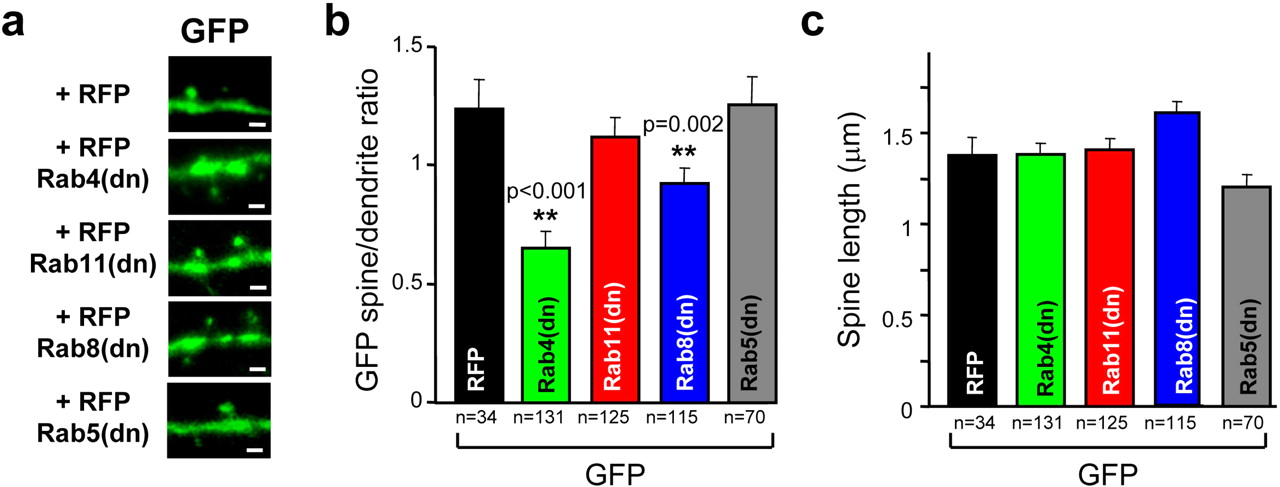
Rab dominant-negative effect on spine size and length. a, Representative confocal images of spines and adjacent dendritic shafts from neurons transfected with GFP and RFP (top) or with GFP and RFP–Rabdn constructs (bottom 4 panels), as indicated. GFP fluorescence signal (green) represents total GFP distribution to indicate volume of the spine and adjacent dendritic shaft. Scale bars, 1 μm. b, Average spine/dendrite ratio of GFP fluorescence was calculated and analyzed as described for Figure 2. c, Average spine length was calculated as the distance between the peak of GFP fluorescence intensity in the spine and in the adjacent dendritic shaft from neurons expressing GFP and different RFP-tagged constructs, as indicated. n represents number of spine–dendrite pairs from five to seven neurons per condition.
Blockade of endosomal trafficking does not affect basal AMPA receptor phosphorylation
It has been shown previously that GluR1 phosphorylation at Ser845 by protein kinase A is required for LTP (Esteban et al., 2003). In addition, GluR1 is phosphorylated at Ser831 during LTP induction and CaMKII activation (Barria et al., 1997a,b; Mammen et al., 1997). Therefore, we decided to test whether blockade of Rab protein function could alter AMPAR phosphorylation, potentially altering the competency of the receptor to undergo synaptic delivery.
To examine the role of endosomal trafficking in basal AMPAR phosphorylation, we prepared homogenates of hippocampal slice cultures after overnight expression of Rab4dn, Rab11dn, Rab8dn, or Rab5dn. Uninfected slices were used as control. Total and phosphorylated GluR1 levels were then assayed by Western blot analysis using specific antibodies. As shown in Figure 4, none of these Rabdns exhibited any significant alterations in the phosphorylation levels of GluR1 Ser831 or Ser845. Therefore, the blockade of LTP by Rab8dn and Rab11dn cannot be explained by an alteration in GluR1 phosphorylation.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Dominant-negative Rab proteins do not alter basal levels of GluR1 phosphorylation. a, Western blot representative example of the effect of Rabdn expression on GluR1 phosphorylation at Ser845 and Ser831. Protein extracts were prepared from microdissected CA1 regions expressing the indicated Rabdn protein fused to GFP. b, c, Quantification of GluR1 phospho-serine 845 (P-S845; b) or phospho-serine 831 (P-S831; c) relative to total GluR1. Values are normalized to the ratio of phospho/total GluR1 from uninfected slices processed in parallel. n represents number of independent experiments.
Discussion
The results presented in this work constitute the first molecular and functional dissection of the local endosomal network that operates at postsynaptic terminals during synaptic plasticity in the hippocampus. Perhaps one of the most striking findings of this study is that a relay of distinct Rab-dependent endosomal functions is required for the transport of AMPARs within the micrometer-sized confines of the postsynaptic terminal. This conclusion is based on electrophysiological data monitoring functional endogenous receptors at synapses and fluorescence imaging experiments visualizing the partition of receptors between dendritic and spine compartments.
We found that the delivery of new AMPARs into synapses requires two distinct steps. First, Rab11-driven endosomes mediate the translocation of AMPARs into spines. Accordingly, blockade of Rab11 function prevents LTP and impairs AMPAR accumulation within the spine. However, Rab11-dependent trafficking is not sufficient to complete the delivery of AMPARs into the synaptic membrane. Rab8 function is also required. Interestingly, Rab8-dependent AMPAR trafficking appears to operate entirely within the spine. In fact, Rab8dn does not impair, and it actually enhances, AMPAR localization at spines. Therefore, we propose that AMPAR cargo driven by Rab11 endosomes from the dendritic shaft is handed over to a separate, Rab8-controlled compartment within the spine, which mediates the final insertion of AMPARs into the synaptic membrane. GluR1 translocation into spines is an activity- and CaMKII-dependent process (Shi et al., 1999; Piccini and Malinow, 2002; Kopec et al., 2006). Therefore, the involvement of two different endosomal populations for AMPAR synaptic delivery may have developed to allow a separate control of activity-dependent (dendrite-to-spine) and constitutive (intraspine) trafficking of AMPARs.
In addition, we found that there is a remarkable segregation of endosomal trafficking within the spine under basal conditions. Specifically, our results suggest that Rab4-driven endosomes are the main mediators of continuous membrane recycling at spines. Thus, blockade of Rab4 function leads to a drastic reduction in spine size. Interestingly, this membrane circuit does not seem to be used by AMPARs under basal conditions, because Rab4dn does not alter AMPAR synaptic transmission (Gerges et al., 2004a). We now report that Rab8-dependent trafficking, which does control AMPAR constitutive cycling (Gerges et al., 2004a), also contributes to the maintenance of spine size, although to a lesser extent than Rab4-driven membrane recycling. Of note, Rab11 appears to have a minor contribution to spine maintenance under basal conditions.
In conclusion, this work has allowed us to lay out a functional map of the membrane sorting network that controls AMPAR transport at postsynaptic terminals. In addition to providing new information about the machinery that mediates AMPAR trafficking, our results shed light into basic cell biological processes responsible for sustaining synaptic function and driving the physiological changes associated with synaptic plasticity.
Footnotes
- Received September 17, 2007.
- Revision received October 11, 2007.
- Accepted October 12, 2007.
-
This work was supported by National Institute of Mental Health Grants MH070417 (J.A.E.) and F31-MH070205 (T.C.B.). We thank Ronald Holz, Jeffrey Martens, Maria Soengas, and members of the Esteban laboratory for critical reading of this manuscript.
- Correspondence should be addressed to José A. Esteban, Department of Pharmacology, University of Michigan Medical School, 1150 West Medical Center Drive, Ann Arbor, MI 48109-0632. estebanj{at}umich.edu
- Copyright © 2007 Society for Neuroscience 0270-6474/07/2713311-05$15.00/0