
Abstract
Auditory processing and hearing-related pathologies are heavily influenced by steroid hormones in a variety of vertebrate species, including humans. The hormone estradiol has been recently shown to directly modulate the gain of central auditory neurons, in real time, by controlling the strength of inhibitory transmission via a nongenomic mechanism. The functional relevance of this modulation, however, remains unknown. Here we show that estradiol generated in the songbird homolog of the mammalian auditory association cortex, rapidly enhances the effectiveness of the neural coding of complex, learned acoustic signals in awake zebra finches. Specifically, estradiol increases mutual information rates, coding efficiency, and the neural discrimination of songs. These effects are mediated by estradiol's modulation of both the rate and temporal coding of auditory signals. Interference with the local action or production of estradiol in the auditory forebrain of freely behaving animals disrupts behavioral responses to songs, but not to other behaviorally relevant communication signals. Our findings directly show that estradiol is a key regulator of auditory function in the adult vertebrate brain.
Introduction
The perceptual processing of behaviorally relevant auditory signals relies on the adequate encoding and transformation of acoustic cues into discrete neural representations. Although these processes are constrained by structural properties of auditory circuits, hearing-driven responses are modulated by neurotransmitter and neuromodulatory systems, which converge to determine receptive field tuning properties (Chen and Jen, 2000; Wehr and Zador, 2003; Theunissen et al., 2004; Yan and Zhang, 2005; Zhang et al., 2005; Wutzler et al., 2008). Neurochemical signaling sets resolution during signal acquisition and determines the information from acoustic signals that is to be extracted, encoded, and ultimately interpreted by auditory circuits (Edeline et al., 1994; Edeline, 1995; Manunta and Edeline, 1997; Wehr and Zador, 2003; Wutzler et al., 2008). Although most research has explored how neurotransmitter and monoaminergic systems shape auditory coding, it has become clear that nontraditional neuromodulators contribute several operational rules of auditory networks.
One novel example of a nontraditional neuromodulator is the hormone 17β-estradiol [estradiol (E2)], which can rapidly and robustly modulate auditory physiology. For instance, auditory processing efficiency oscillates with E2 levels through the menstrual cycle (Walpurger et al., 2004), and women with Turner's syndrome, who are deficient in estrogens, exhibit hearing impairments and central auditory dysfunction (Güngör et al., 2000; Hederstierna et al., 2009a). In rodents and monkeys, E2 therapy recovers aspects of auditory processing degraded by overariectomies (Coleman et al., 1994; Golub et al., 2004), and estrogen receptor β-deficient mice exhibit severe progressive hearing loss that leads to early deafness (Simonoska et al., 2009). Although these findings implicated estradiol as a regulator of auditory function, only recently a causal link and a mechanistic understanding of this sensory–neuroendocrine interaction were demonstrated in songbirds (Remage-Healey et al., 2008, 2010; Tremere et al., 2009).
Songbirds offer an excellent model to study how auditory processing is shaped by neurosteroids, as they synthesize brain-generated estrogens abundantly and rely heavily on vocal communication for social behaviors (Gentner, 2004; Nowicki and Searcy, 2004; Forlano et al., 2006; Schlinger and London, 2006; Garcia-Segura, 2008; Remage-Healey et al., 2008). The songbird analog of the mammalian auditory association cortex [caudomedial nidopallium (NCM)] has generated special interest, as it appears to be central for auditory discrimination and memory formation (Bolhuis and Gahr, 2006; Gobes and Bolhuis, 2007; London and Clayton, 2008; Pinaud and Terleph, 2008). Importantly, large fractions of NCM neurons either produce or are sensitive to estradiol, indicating that neurosteroid-based signaling influences auditory circuit physiology (Shen et al., 1995; Bernard et al., 1999; Saldanha et al., 2000; Balthazart et al., 2003; Tremere et al., 2009). Estradiol levels in NCM are rapidly and locally regulated by experience in freely behaving animals (Remage-Healey et al., 2008), and this neurohormone instantaneously enhances the gain of hearing-driven responses by decreasing the strength of GABAergic transmission via a nongenomic mechanism (Tremere et al., 2009). Although these findings demonstrated that estradiol shapes auditory physiology, it is unclear what processing capabilities it offers auditory neurons.
Here we directly and quantitatively determined the functional relevance of estradiol's regulation of auditory processing. We found that brain-generated estradiol modulates auditory coding, and thereby improves the neural and behavioral discrimination of communication signals.
Materials and Methods
Animals
We used 100 adult zebra finches for both electrophysiological and behavioral studies (n = 74 males and 26 females; details below). As with our previous work (Tremere et al., 2009), given that no sex differences were detected in any parameter of our current studies, males and females were grouped for analysis. All procedures were approved by the University of Oklahoma Health Sciences Center Institutional Animal Care and Use Committee, and are in accordance with National Institutes of Health guidelines.
Neurophysiological recordings coupled to intracerebral pharmacology in awake restrained animals
Animals were fitted with a recording chamber and a metal head post under anesthesia and were repeatedly acclimated to restraint as we detailed previously (Pinaud et al., 2008b; Tremere et al., 2009, 2010). We used a multielectrode array consisting of eight tungsten microelectrodes (∼5 MΩ; 4 electrodes per hemisphere) that we developed and previously described in detail (Pinaud et al., 2008b; Tremere et al., 2009, 2010), where bilateral electrophysiological recordings were obtained from the NCM of awake, restrained birds. Two glass pipettes (tip internal diameter, ∼20 μm; Drummond Scientific) with solutions of interest were inserted into either hemisphere adjacent to the microelectrodes. Whereas one hemisphere received vehicle, the other was continuously infused with drugs that target the estrogen system (see below). Importantly, to determine whether drug effects were lateralized, vehicle-injected hemispheres were intentionally switched in subsets of animals. No significant differences were detected across hemispheres for all compounds tested. We therefore collapsed data for all hemispheres for each condition. Responsive sites and single-unit isolation were achieved by intermittent stimulation with bursts of white noise (Tremere et al., 2009). The single-unit signal-to-noise ratio was high. On average, the signal-to-noise ratio of action potentials obtained across all of our recordings approached 32 (peak-peak of action potential, normalized by the noise root mean square) (Lin and Liu, 2010).
Our auditory stimulation set consisted of four conspecific songs motifs (durations of 0.70, 0.67, 0.72, and 0.73 s). These songs were novel as they derived from three separately housed, nongenetically related colonies, and were chosen because they capture much of the natural variability in zebra finch songs, in the dimensions of speed rates and syllable complexity.
Stimuli were played in pseudo-random order through a speaker, via the control of a custom-written multitrial sequence program, at 70 dB sound pressure level amplitude. Following presentation of a “predrug” auditory stimulation set (25 trials/song), dimethylsulfoxide (DMSO; diluted in 50% saline), used as the control solution, was injected in one hemisphere and drug or hormone was injected into the contralateral hemisphere (Fig. 1). During these intracerebral pharmacological manipulations, birds were once again exposed to the auditory stimulation set (“drug” condition). Injections were performed with two hydraulic Narishige microinjectors and consisted of an initial loading dose (100 nl), followed by maintenance doses (10 nl), which occurred every 5–10 min throughout the duration of the stimulation session. We previously established that DMSO impacts neither electrophysiological responses nor the expression of activity-dependent molecular markers under our experimental conditions (Tremere et al., 2009). Injections in the experimental hemisphere included the following: E2 (Sigma; n = 10 birds) at 30 μg/ml, which is well within the physiological range in NCM (Remage-Healey et al., 2008, 2010); ICI 182,780 [7α,17β-[9-[(4,4,5,5,5-pentafluoropentyl)sulfinyl]nonyl]estra-1,3,5(10)-triene-3,17-diol (ICI); Sigma; n = 8 birds] at 100 μm, tamoxifen (TMX; Tocris Bioscience; n = 8 birds) at 100 μm, fadrozole (FAD; Sigma; n = 8 birds) at 100 μm, 1,4,6-androstatrien-3,17-dione (ATD; Steraloids; n = 8 birds) at 100 μm, 5β-dihydrotestosterone (5β-DHT; Steraloids; n = 4 birds) at 100 μm, and estrone (E1; Sigma; n = 4 birds) at 100 μm. All agents were diluted to their final concentration in DMSO. During the drug session, song stimulation commenced 5 min following infusions of E2, ICI, TMX, 5β-DHT, and E1, and 30 min after injections of FAD and ATD (Fig. 1). The latter schedule was used to ensure full inhibition of aromatase activity and suppression of local E2 levels (Wade et al., 1994; Remage-Healey et al., 2008, 2010; Tremere et al., 2009). At the end of the drug session, infusion of all solutions was interrupted and animals were kept in complete silence for 2 h, at which point birds were exposed to the stimulation set once again (Off + 2 h). Following an additional 2 h silence period, animals were exposed to the stimulus set one final time (Off + 4 h), and were immediately killed afterward (Fig. 1).

Flowchart of experimental configuration. After head posting and recording chamber implantation, all animals were subjected to several acclimation sessions (see Materials and Methods). On the day of the experiment, animals were exposed to two final acclimation sessions and were placed in groups based on the pharmacological manipulations to be carried out. A, For experiments with E2, ICI, TMX, 5β-DHT, and E1, animals were exposed to the stimulation set, during which song-driven neural responses were obtained bilaterally from NCM (predrug session). Subsequently, animals were unilaterally injected with one of the drugs above, and contralaterally infused with vehicle; maintenance doses were applied during a second presentation of the stimulus set (drug session). Drug infusion was then interrupted and animals were left in silence for 2 h, with the electrodes in place. Subsequently, birds were exposed to the stimulation set a third time (Off + 2 h) and were left in silence for an additional 2 h. Animals were then stimulated a final time (Off + 4 h) before brains were obtained for histological analysis. B, In studies with FAD and ATD, the experimental configuration was identical to the one described above, except that the drug session was only initiated 30 min after the initial drug infusion. This approach was taken to ensure that aromatase inhibition was efficient and maximal (Wade et al., 1994; Tremere et al., 2009) and that endogenous E2 levels in NCM decayed to minimal levels (Remage-Healey et al., 2008).
Confirmation of electrode position and drug diffusion
Following the recording session, electrolytic lesions were generated in both hemispheres of each animal (15 μA for 10 s), and brain sections were processed for cresyl violet histochemistry to confirm electrode placement (Pinaud et al., 2008b; Tremere et al., 2009). Importantly, we used two approaches to ensure that our drug diffusions were restricted to NCM. First, we previously measured drug diffusion from our injection pipettes and verified that our infusion regimens and volumes provide broad coverage (∼250 μm radius) that is highly restricted to NCM, even in the event that injections are misplaced by several hundred micrometers [NCM is a large area that spans ∼1.4 mm dorsoventral (DV) and ∼1.2 mm mediolateral (ML)] (Tremere et al., 2009). Second, we subjected our brain sections to fluorescence in situ hybridization directed at zenk mRNA. We previously showed that zenk expression in NCM is highly sensitive to manipulations directed at local actions of E2, which provided us with a convenient tool to probe the extent to which our pharmacological manipulations diffused within NCM tissue (Tremere et al., 2009). Finally, although the drug volumes used were very small, and diffusion was extensively controlled for in our studies, we emphasize that aromatase- and estrogen receptor-positive neurons are restricted to NCM in the caudomedial forebrain; thus, even in the unlikely event that any of our solutions (E2, estrogen receptor antagonists, and aromatase inhibitors) diffused away from NCM, these drugs would not impact the physiology of cells located in regions that surround NCM. Altogether, these assessments ensured that the diffusion of all solutions were restricted to, and selectively impacted, NCM neurons.
Pattern classification analysis
Song-driven electrophysiological responses were obtained from single units in NCM before (predrug), during (drug), and 2 and 4 h after interruption of drug infusions (Off + 2 h and Off + 4 h, respectively), as described previously. We used a decoding method based on a linear classifier to quantify the extent to which the responses of individual neurons could distinguish among the different songs tested. A linear decoder is conservative because it is biologically plausible and easy to implement neurally. Given that a linear decoder can reliably extract song identity from the firing patterns of NCM neurons (see Results), then many nonlinear decoding schemes should work as well. We emphasize that our goal is not to evaluate different decoding schemes but rather to quantify the effects of the actions of local E2 on the information carried by NCM neurons regarding song identity; the fact that a linear classifier is simple and performs well makes it ideal for this purpose.
By applying the classifier to decode neural responses obtained before, during, and after drug application, we directly quantified how drug (E2, ICI, TMX, FAD, ATD, 5β-DHT, and E1) altered the information that NCM neurons carry about song structure and, consequently, how these pharmacological agents impact the effectiveness of the neuronal coding of different songs. Here we use the term “coding” to simply refer to the ability of auditory neurons to encode time-varying stimuli. Specifically, we recorded single-unit responses to song stimuli (25 trials for each of 4 songs), and spikes were grouped into 50 ms bins. These parameters (number of trials per song and bin size) were defined as optimal, based on initial studies aimed at determining the best possible performance of our classifier (data not shown). We next trained the decoder on binned discharge rates from each neuron from half of the predrug trials (training set) to generate a response template for our conditions of interest (before vs during and after drug treatment). Performance of the classifier was computed via a “test set,” which corresponds to the remaining half of the stimulation trials (control trials for cross-validation); the classifier assigned song identity based on the proximity between training and testing vectors, which was determined by calculating the Euclidean distance between test vectors and the templates for each song (Fig. 2). The precision of the classifier was quantified as the percentage of test trials in which the classifier correctly identified the song that produced the measured responses. To quantify how our pharmacological manipulations impacted the ability of NCM neurons to discriminate across the different stimuli, we trained our classifier with training sets obtained in the predrug sessions, as above, used test sets obtained during and after (2 and 4 h) drug treatment, and assessed our decoder's performance for each of these conditions.

Decoding approach based on a pattern classifier. Each recorded unit was driven by four songs (25 trials/song), in pseudo-random order, before, during, and after pharmacological treatment. A, The sonogram for a song stimulus is shown in the top panel, and a raster plot illustrating the discharge profile of a single-unit from NCM is shown in the bottom panel, along with its peristimulus time histogram. In the predrug session, data were split into a training set and a testing set, each including half of the stimulation trials per song. B, Spike counts from each trial were binned into 50 ms bins (a raw trace is shown for illustrative purposes and does not derive from the raster plot shown in A). The training set (n = 13 trials/song) was used to train the classifier, and the remaining half of the trials (n = 12/song) was used as testing vectors for cross-validation and calculation of classification accuracies. C, Activity vectors for each cell and song were plotted in hyperdimensional space to generate templates for each song (song 1 is illustrated). D, After generating templates for each of the four songs used, we tested the ability of the classifier to decode stimulus identity based on the neural responses obtained from the testing set activity vectors (or trials obtained during drug, Off + 2 h and Off + 4 h sessions). Illustrated is an attempt to decode the stimulus identity (Sx) that originated the orange activity vector. The classifier measures Euclidean distance (d) between “inquiry” vectors and each of the song templates, and assigns song identity based on the proximity between training and testing vectors. Classification accuracies are taken as the percentage of trials in which the classifier predicted the correct song based on the neural response.
To test whether pharmacological manipulations altered the structure of neural responses in the absence of changes in the neural discrimination by songs of NCM, we performed control experiments where we trained and tested the classifier with neurophysiological responses obtained from within the same experimental groups and pharmacological sessions for each song (e.g., trained the classifier with half of the trials obtained during E2 infusions and tested the decoder's performance with the remaining half of the E2 trials).
The ability with which NCM neurons discriminate across the different songs tested (neural discrimination) was investigated by evaluating the pattern of pairwise confusions obtained and measured by our classifier analysis. Pattern classification and confusion analyses were performed with custom-written Matlab routines.
“Indirect” calculation of mutual information rates
We have used an indirect method to calculate a lower bound on the total information as a conservative approach. We indirectly estimated mutual information (MI) I[X;Y] between stimuli X and neuronal responses Y of single NCM units to calculate the effectiveness of neural encoding. To this end, spike trains were binned into 50 ms bins and counted. Data were then divided into the following two segments: half of the trials were used as the training set and the remaining half as a testing set, in the same manner as described above. For each cell, we performed a logistic regression on the training set, which takes as input a vector of activity (spike counts/bin) and returns as output a joint probability distribution, P(y,x) = P(y|x)P(x), across both the four songs and different observed activity vectors. The distribution was normalized, and we used a Bayesian prior on the regression parameters to avoid overfitting (Bishop, 2006). The parameters of the regression were then saved (i.e., the weights, n × 4, where n is the number of bins), and the regression was applied to the test set without adjusting the weights. Thus, after training, each test trial response y is given a set of four probabilities by computing the logistic regression for each song. We next computed the entropy of the output distribution averaged across trials and subsequently averaged across cells. The entropy of the output distribution is defined as follows:
 or the sum over all four categories of P(c_i)log2(P(c_i)), where c is the class and i is an index over classes (i.e., with 4 classes, i is any value between 1 and 4). To measure the information that is associated with a specific song, we define the stimulus-specific information (SSI):
or the sum over all four categories of P(c_i)log2(P(c_i)), where c is the class and i is an index over classes (i.e., with 4 classes, i is any value between 1 and 4). To measure the information that is associated with a specific song, we define the stimulus-specific information (SSI):
 where H[X] = −
where H[X] = − Information units expressed in the Results reflect an entire trial for the set of four songs (i.e., values are bits/stimulus for a set of 4 stimuli where the maximum value attainable is 2).
Information units expressed in the Results reflect an entire trial for the set of four songs (i.e., values are bits/stimulus for a set of 4 stimuli where the maximum value attainable is 2).
As detailed above, we have chosen to compute information between the songs estimated from the neural activity (estimated via the logistic regression) and the actual songs, instead of directly computing the mutual information between the neural response and the songs. We have used this approach, commonly referred to as the “indirect method,” because information is rather sensitive to bias, especially when considering a limited dataset (i.e., limited number of stimulus-response samples obtainable in a real experiment). Thus, to be conservative, we have used the indirect method as it is a lower bound on the total information, is more adequate for computing information with a limited experimental dataset, has been previously used to quantify neural data, and offers other advantages over more classic methods of estimating information (Bialek et al., 1993; Rieke et al., 1997; Borst and Theunissen, 1999; Chacron et al., 2004; Chacron, 2006). Finally, we emphasize that it is not our goal to quantify the absolute information from our neural data, but rather to use a conservative metric to quantify how estradiol (or associated manipulations) modifies information handling by NCM neurons.
Spike normalization studies
To determine the extent to which E2- or drug-induced alterations in the discharge rate of NCM neurons account for changes in the effectiveness of the neural coding of songs, we carried spike normalization manipulations as follows.
Normalization of E2 treatment (which significantly increases discharge rates of NCM neurons).
We binned discharge rates (as detailed previously) for each stimulation trial before (pre-E2) and during (E2) infusions. Activity vectors obtained for each stimulation trial in the pre-E2 session were averaged to obtain a single activity vector per song, for each cell. Subsequently, activity vectors obtained for each stimulation trial during E2 infusions were subjected to spike extractions, which were performed at random spike timings across the spike train (for a schematic view of this manipulation, see Fig. 9A,B). Spike extractions for each activity vector (i.e., each trial during E2 infusions) were performed to precisely match the number of spikes present in the average activity vector obtained in the pre-E2 session for each individual neuron. In other words, the average spike rate used for the normalization procedure was calculated for the predrug session, for each neuron. We, therefore, did not assume an average spike rate for our neuronal population, but rather normalized the spike rate of each neuron to its own predrug state. This manipulation was also conducted in spike trains obtained in the Off + 2 h and Off + 4 h sessions. Subsequently, both intact and processed (spike-extracted) activity vectors were subjected to our pattern classification analyses, as detailed previously.
Normalization of ICI, TMX, FAD, and ATD treatments (which significantly decrease discharge rates of NCM neurons).
As above, activity vectors were obtained for each stimulation trial before (predrug) and during (drug) infusions. Average activity vectors were then obtained for each song and cell, by averaging individual activity vectors that derived from each stimulation trial. Next, activity vectors obtained for each stimulation trial during drug infusions (i.e., ICI, TMX, FAD, or ATD) were subjected to “spike insertions” to increase the number of spikes in a train, to match the average activity vectors obtained for each individual neuron in the predrug session (for a schematic view of this manipulation, see Fig. 9A,B). Spikes were inserted across the spike train, at random spike timings that followed a Poisson distribution (i.e., we used an exponential interspike interval distribution that is consistent with a Poisson spike count distribution). Importantly, this process was performed for spike trains obtained for each cell and song, for each of the drugs tested, such that activity vectors obtained during drug treatment would exhibit the same number of spikes contained in the predrug average activity vector obtained for each song (Fig. 9, compare A, B, spike n). As above, the average neuronal firing rate used to normalize responses was calculated for each neuron separately during the predrug session. We subsequently normalized the spike rate of each neuron to its own predrug state; thus, no assumptions were made regarding average spike rates for our population of neurons. Spike trains obtained during Off + 2 h and Off + 4 h sessions were also subjected to this manipulation. As above, intact and processed (spike-inserted) spike trains were subjected to pattern classification analyses.
Importantly, the aim of these spike normalization studies was to carry out an initial assessment on the extent to which the effects of E2 on the neuronal physiology of NCM are mediated by changes in the rate and/or temporal organization of song-driven spiking activity (rate vs temporal coding). We clarify that multiple and rather broad definitions exist for temporal coding (e.g., Theunissen and Miller, 1995; Rieke et al., 1997), but overall most reflect the precision of action potential timing. Given that a detailed study of the effects of E2 on the temporal response properties of NCM neurons falls beyond the scope of this work, we use here a definition of temporal coding to reflect drug-induced changes in classification accuracies and information rates that cannot be explained by global alterations in spike rates.
The spike normalization experiments detailed above were conducted using custom-written Matlab routines.
Song and call discrimination tasks
We examined whether the local synthesis and actions of E2 in the physiology of NCM impacts the auditory discrimination and, consequently, behavioral preference to auditory signals. To this end, we combined local in vivo pharmacological manipulations and standard two-choice discrimination tasks (Clayton, 1988; Gobes and Bolhuis, 2007; Holveck and Riebel, 2007; Remage-Healey et al., 2010). It has been well established that male zebra finches exhibit a robust behavioral preference toward the song of their tutors relative to a novel, previously unheard song (Riebel and Smallegange, 2003; Gobes and Bolhuis, 2007; Holveck and Riebel, 2007; Remage-Healey et al., 2010). This preference can be quantified by subjecting animals to a two-choice song task, which consists of placing birds in a cage subdivided into three connected chambers. Birds can freely fly across chambers. Either a tutor (TUT) song or a novel conspecific (CON) song is assigned to one of two speakers placed at the ends of each side chamber. One TUT and one CON song are played in alternation through each speaker. Songbirds have an innate preference to fly toward the chamber playing the TUT song. Importantly, the behavioral choice requires birds to process and discriminate between the two stimuli.
We used this behavioral paradigm to test how interference with the local actions of E2 in NCM neurons impact auditory discrimination, and consequently song preference, in adult male zebra finches (n = 25; 5/group). Birds were first prepared for in vivo pharmacological manipulations. To this end, animals were anesthetized and placed in a stereotaxic device. The outer layer of skull was opened over the caudomedial forebrain, and birds were implanted with guide cannulas bilaterally within NCM (anteroposterior: 0.5 mm; ML: 0.65 mm; DV: 1.2 mm), and with a metal head post. The head post and a plastic body tube allowed for the restraint of awake animals during bilateral microinjections both before the behavioral task and between the two test trials (see below). Animals were allowed to recover for a minimum of 2 d and were then individually subjected to five acclimation sessions (one every 1.5 h) in a walk-in sound booth where the behavioral chamber was located. Each acclimation session consisted of gently placing the bird in a body restraint tube, fixing it through the head post to a stereotaxic frame with an adapted clamp (located within the soundproof booth), and maintaining the animal in restraint for 15 min in the presence of the investigator. Following the restraint session, animals were then individually placed in the behavioral task cage. The following day, animals were subjected to two final acclimation sessions before being tested in the auditory discrimination task. Birds were tested twice to assess song preference before and during pharmacological treatment (predrug and drug sessions, respectively). For the predrug session, immediately before the onset of the task, animals were restrained and a “mock” infusion was simulated bilaterally, but no solutions were injected. Animals were then subjected to the song preference task, as detailed below. The interval between the predrug and drug sessions was 3 h. For the drug session, immediately before the onset of the task, animals were restrained and infused bilaterally with vehicle (controls), ICI, or TMX (0.5 μl at 100 μm, each). A subset of animals was bilaterally infused with either FAD or ATD (0.5 μl at 100 μm, each) 30 min before the beginning of the task. For both predrug and drug sessions, animals were individually placed in the behavioral cage, which was divided into three interconnected chambers; speakers connected to a computer were positioned outside each end of the cage. The behavior setup including cage, speakers, and computer were located in a walk-in soundproof booth with a one-way mirror. The task entailed two sessions separated by 30 min. Each session was initiated when birds were located in the center chamber. A single stimulus (TUT song; duration, 2.93 s) was played from one speaker, followed by a second stimulus (CON song; duration, 2.95 s), which was broadcast through the second speaker. The CON song was obtained from a separate colony to ensure that birds had not been previously exposed to this stimulus. These two stimuli were then played, one at a time, in alternation from each speaker for a total of 30 min, with the intersong interval set at 10 s. After completion of the first session, animals were removed from the behavioral cage, restrained, injected bilaterally a second time, and left in silence for 10 min. Birds were then returned to the behavioral cage and exposed to the same alternating stimuli in the second 30 min session, except that the songs allocated to each speaker were reversed. The length of time spent in each chamber, along with the specific times in which birds moved across chambers, was documented by two investigators that were blind to the treatment. Data were averaged across the observers, and a preference score was calculated. The preference score was obtained by dividing the time spent in the TUT song chamber by the combined time spent in both TUT and CON song chambers.
We exposed separate groups of animals to a call discrimination task, which is fundamentally similar to the song discrimination task detailed above, except that female and male calls are broadcast in alternation. It has been well documented that male zebra finches discriminate calls of both sexes, but display a significant behavioral preference toward female calls (Vicario et al., 2001; Gobes and Bolhuis, 2007). Importantly, the complexity and frequency range of calls and song elements are highly similar. Specifically, we used the classic “distance calls” (6 calls/sex; duration of male calls: 187 ± 14 ms [mean ± SE]; duration of female calls: 206 ± 18 ms). Birds were again assigned to vehicle, ICI, TMX, FAD, and ATD groups (n = 25; 5/group), and were subjected to the call discrimination task, following the procedures detailed above for song discrimination. Call/response ratios were obtained for each bird by dividing the total number of responses to female calls by the total number of responses to male calls. Data from animals belonging to the same experimental group were joined and averaged. As indicated previously, this behavioral task also requires birds to process and discriminate across the auditory stimuli, such that an appropriate behavioral response can be taken.
For both behavioral tasks, animals had to visit the chamber playing either stimuli at least twice, and were required to spend a minimum of 10% of the total test length at either chamber to meet motivation criteria (Gobes and Bolhuis, 2007; Holveck and Riebel, 2007; Remage-Healey et al., 2010). Importantly, none of the pharmacological treatments used in this study altered the responsiveness of the birds to the stimuli (i.e., did not alter the time spent in the central chamber during drug treatment or the animals' responsiveness to either stimuli). After behavioral data were collected, brains of all birds were histologically processed for verification of cannulae placement and injection locations.
Statistical analyses
To analyze the effects of pharmacological manipulations on the song-evoked neurophysiological responses, we used parametric statistics, as appropriate. However, we used nonparametric tests when normality assumptions were violated. Factorial ANOVAs included recordings from the same electrode before versus during and after drug (vehicle, E2, ICI, TMX, FAD, or ATD) as a repeated measure, and used data from each recording site as a sample. To avoid pseudo-replication effects, the degrees of freedom used when computing the probability for each F value were reduced to the number of stimuli and/or birds, as appropriate. We set statistical significance thresholds at p < 0.05.
For pattern classification analyses, we used a bootstrapping and resampling procedure to generate 10 sample subsets from the dataset (i.e., the set of neural responses obtained for each cell, for each of the conditions tested). This nonparametric approach was used to obtain a sampling distribution of classification accuracies, and consequently confidence intervals, for each cell before, during, and after pharmacological sessions. Data for each condition (predrug, drug, Off + 2 h, and Off + 4 h) were combined for all cells into separate groups. To test for significant effects across conditions, we fit a linear mixed-model regression to the data. Each comparison to be performed was numerically segregated (e.g., the predrug condition was coded with a numerical value of 1, and the drug condition with a numerical value of 2). To test for significant effects between any two groups (e.g., predrug vs drug or predrug vs Off + 4 h), we considered the slope of the regression β ≠ 0. To control for random effects, we also modeled random intercepts of different cells with the following regression model:
 where CR refers to the classification results, β is the slope of the regression, and CAT refers to the categories or groups (predrug, drug, Off + 2 h, or Off + 4 h). By running the regression, we obtained β, where increased β values reflected increased classification accuracies. To test whether the β value is significantly different from 0 (null hypothesis), we performed an independent one-sample t test with significance levels set at p < 0.05. In short, the R function provided us with the slope (i.e., β) and its SE. The t statistic was obtained by dividing the slope by its SE. Considering the degrees of freedom in our test, R automatically determined whether this t statistic corresponded to a significant p value. The purpose of using the linear regression with random effects was to ensure that the results were controlled such that any observed effects (e.g., the significant slope) were not due to individual cases in the sample, a situation in which a simple ANOVA would not be able to certify. Statistical comparisons for mutual information data across groups were also carried out against the null hypothesis of random association.
where CR refers to the classification results, β is the slope of the regression, and CAT refers to the categories or groups (predrug, drug, Off + 2 h, or Off + 4 h). By running the regression, we obtained β, where increased β values reflected increased classification accuracies. To test whether the β value is significantly different from 0 (null hypothesis), we performed an independent one-sample t test with significance levels set at p < 0.05. In short, the R function provided us with the slope (i.e., β) and its SE. The t statistic was obtained by dividing the slope by its SE. Considering the degrees of freedom in our test, R automatically determined whether this t statistic corresponded to a significant p value. The purpose of using the linear regression with random effects was to ensure that the results were controlled such that any observed effects (e.g., the significant slope) were not due to individual cases in the sample, a situation in which a simple ANOVA would not be able to certify. Statistical comparisons for mutual information data across groups were also carried out against the null hypothesis of random association.
For behavioral tests, repeated-measures two-way ANOVA was used to test for effects of session (before/after injection) and pharmacological treatment (vehicle vs ICI, TMX, FAD, or ATD) with Tukey post hoc analysis. Significance was set at p < 0.05.
Results
Rapid and persistent modulation of auditory-evoked responses by estradiol in the awake brain
We recently showed that brain-generated E2 rapidly and robustly modulates hearing-evoked activity of NCM neurons in the awake brain via a presynaptic, nongenomic mechanism that involves local suppression of inhibitory transmission (Tremere et al., 2009). It is unknown, however, whether E2 impacts the long-term physiology of NCM neurons. To test this possibility, we performed bilateral, multielectrode extracellular recordings in awake, restrained zebra finches before, during, and after (2 and 4 h) local, bilateral, and simultaneous intracerebral pharmacological manipulations, using a methodological approach that we developed and used previously (Pinaud et al., 2008b; Tremere et al., 2009, 2010). Auditory-evoked activity was obtained from single units in NCM during playbacks of a randomized series of conspecific songs.
Consistent with our previous findings, which have since been validated independently (Remage-Healey et al., 2010), we found that local production and action of E2 markedly modulates the physiology of NCM neurons (Fig. 3). Specifically, before local pharmacological interventions, the median (± SE) firing rate of NCM neurons was 28.8 ± 14 spikes/s. Infusions of physiological concentrations of E2 (30 μg/ml) significantly increased song-evoked discharge rates by 52%, to 43.7 ± 11 spikes/s (p = 0.02, factorial ANOVA) (Fig. 3B). Even though E2 concentrations in NCM return to baseline levels within 30 min after interruption of local infusions (Remage-Healey et al., 2010), we found that changes in NCM neuronal physiology persisted for several hours. Specifically, firing rates of NCM neurons 2 h after cessation of local E2 infusions remained significantly higher than preinfusion levels (Off + 2 h: 38.2 ± 9 spikes/s; p = 0.03, factorial ANOVA) (Fig. 3B). This effect subsided to control levels 4 h following termination of local E2 treatment (Off + 4 h: 30.1 ± 14 spikes/s; p = 0.09 vs pre-E2 levels, factorial ANOVA) (Fig. 3B). Contralateral infusions of vehicle did not affect song-evoked activity of NCM neurons. Likewise, their discharge rates were not different from preinfusion levels, 2 and 4 h following vehicle treatment.
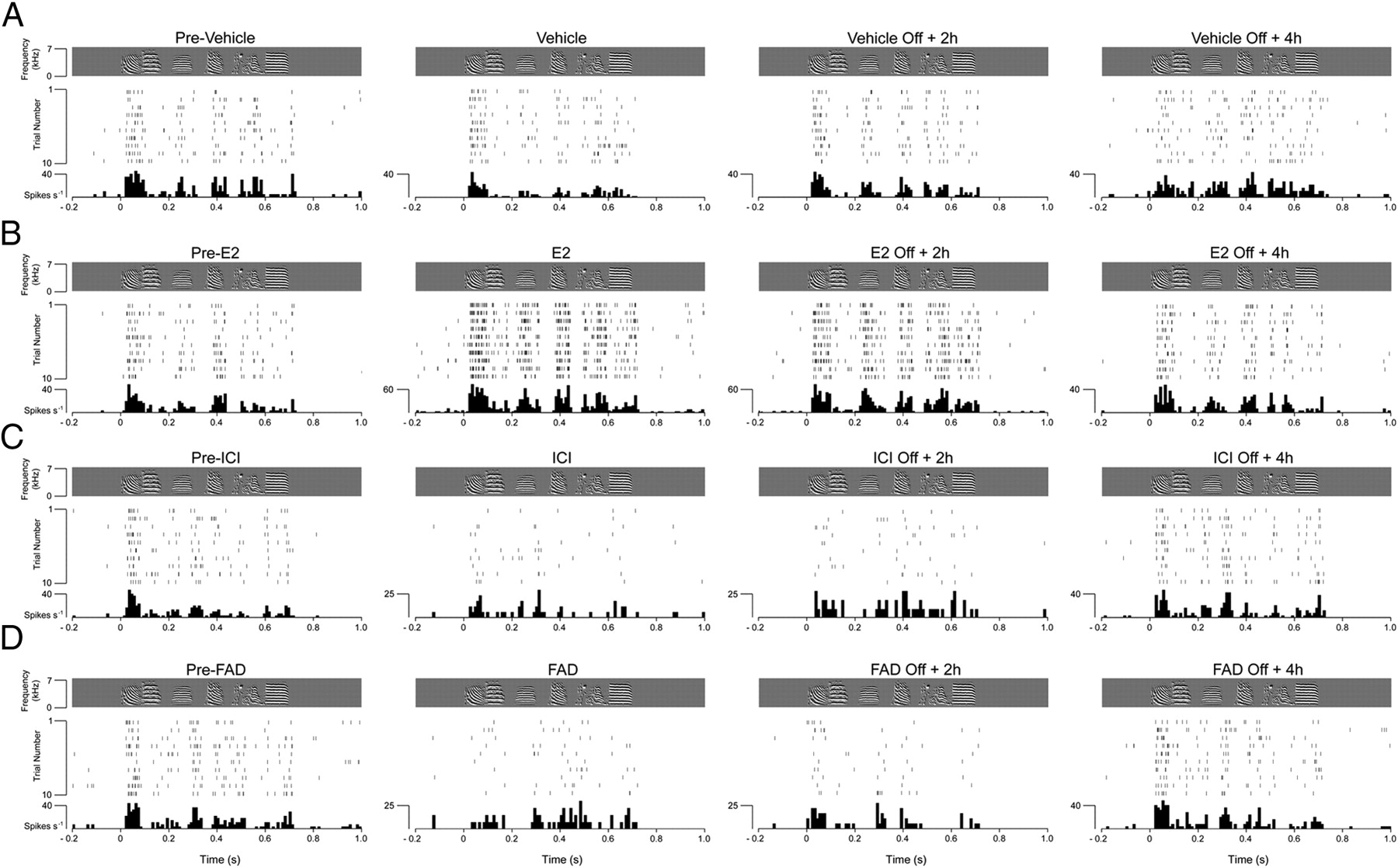
Estradiol rapidly and persistently enhances song-driven discharge rates of NCM neurons. A, Response profile of a representative NCM neuron before, during, and after (2 and 4 h) interruption of local vehicle infusion. Top (gray box), Spectrograms of the song stimulus. Middle, A raster plot for the spiking behavior of the same neuron during the first 10 renditions of the stimulus. Bottom, Peristimulus time histograms (PSTHs). Vehicle infusions did not affect the short- or long-term discharge rates of NCM neurons, as determined by quantitative and statistical analyses (see Results). B, Local infusions of physiological E2 levels rapidly and significantly increase song-evoked firing rates of NCM neurons. Shown is a spike raster plot and associated PSTH of a representative single unit in NCM, time aligned with the stimulus sonogram. E2-mediated effects on firing rates remain detectable 2 h, but not 4 h, following interruption of E2 infusions. C, Local blockade of estrogen receptors with ICI significantly decreased firing rates of NCM neurons. This effect could be detected 2 h after interruption of drug infusions. Shown are raster plots and PSTHs of a single NCM neuron, time aligned with the song stimulus, before, during, and after (2 and 4 h) local infusions of ICI, a high-affinity estrogen receptor antagonist. These effects were also detected when NCM neurons were treated with TMX, an estrogen receptor modulator that acts as an antagonist in NCM (data not shown, but detailed in Results). D, Local inhibition of aromatase with FAD, and suppression of the local production of estradiol in NCM, significantly decreased song-evoked discharge rates of NCM neurons. The effects of FAD on NCM discharge rates were detectable 2 h, but not 4 h, following interruption of drug infusions. Shown are the spectrograms of the stimulus (top panels), spike raster plots (middle panels), and PSTHs (bottom panels) of a single NCM neuron before, during, and after (2 and 4 h) stoppage of local FAD infusions. These effects were also observed when NCM neurons were treated with ATD, an additional aromatase inhibitor (data not shown, but detailed in Results). These findings indicate that locally produced estradiol exerts facilitatory effects in the song-evoked activity of NCM neurons.
Conversely, blockade of estrogen receptors with ICI, a high-affinity estrogen receptor antagonist, or TMX, an estrogen-receptor modulator that acts as an antagonist in NCM, rapidly and significantly decreased song-evoked activity at these central auditory neurons. While the median discharge rate before drug infusion was 29.4 ± 11 spikes/s, ICI and TMX treatment rapidly and significantly decreased NCM neuronal activity to 18.7 ± 6 and 22.3 ± 8 spikes/s, respectively (all p < 0.05 vs predrug levels, factorial ANOVA). These effects also persisted for 2 h, but not 4 h, following drug infusions (Off ICI + 2 h: 20.6 ± 10 spikes/s; Off TMX + 2 h: 24.1 ± 10 spikes/s; Off ICI + 4 h: 30.6 ± 14 spikes/s; Off TMX + 4 h: 31.7 ± 12 spikes/s; p < 0.05 for Off drug + 2 h conditions, and p > 0.05 for Off drug + 4 h conditions, factorial ANOVA) (Fig. 3C).
We next investigated the extent to which these changes are mediated by E2 synthesized locally in NCM. To this end, we treated NCM with FAD or ATD, two specific aromatase inhibitors known to significantly decrease local E2 production in the songbird brain, including the NCM of awake, behaving animals (Wade et al., 1994; Remage-Healey et al., 2008). FAD and ATD significantly decreased NCM neuronal firing rates from preinfusion levels (pre-FAD: 32.4 ± 11 spikes/s; FAD: 12.8 ± 5 spikes/s; pre-ATD: 29.5 ± 14 spikes/s; ATD: 15.1 ± 7 spikes/s; all p < 0.05 vs predrug levels, factorial ANOVA). Notably, although E2 levels typically return to normal (predrug) levels within 30 min following aromatase inhibition in NCM (Remage-Healey et al., 2008), we found that the song-evoked physiology of NCM neurons was also altered in a persistent manner. Specifically, 2 h following interruption of FAD or ATD infusions, discharge rates of NCM neurons were reduced by 62 and 46% from predrug levels (Off FAD + 2 h: 12.2 ± 6 spikes/s; Off ATD + 2 h: 15.9 ± 9 spikes/s; all p < 0.05 vs predrug levels, factorial ANOVA). These effects were no longer detectable 4 h after interruption of drug infusions, when NCM neuronal firing rates were indistinguishable from the predrug state (Off FAD + 4 h: 34.1 ± 14 spikes/s; Off ATD + 4 h: 31.6 ± 11 spikes/s; all p > 0.08 vs predrug levels, factorial ANOVA) (Fig. 3D).
Together, these findings suggest that local production and action of E2 in NCM neurons rapidly enhance song-evoked activity in the awake brain. Moreover, these effects are long lasting and persist for at least 2 h following interruption of E2 infusions.
Quantifying neural coding in NCM neurons
While the results above and our previous findings (Tremere et al., 2009) clearly demonstrate that brain-generated estradiol can modulate, in real time, central auditory processing in the alert vertebrate brain, the functional relevance of such signaling remains unknown. What does estradiol offer NCM neurons in terms of auditory processing or computational capabilities? We hypothesized that a main functional consequence of the modulation of estradiol on the physiology of NCM is to enhance the neural coding of these central auditory neurons (i.e., to increase the ability of NCM neurons to encode time-varying stimuli, in our case, songs). To directly and quantitatively address this hypothesis, we implemented and used a decoding method based on a pattern classifier to quantify how much information NCM neurons carry about stimulus structure and, consequently, discriminate among the different songs tested before, during, and after local pharmacological manipulations directed at the estrogen network (Fig. 4).

Locally generated estradiol enhances auditory coding of NCM neurons. A, The ability of the classifier to decode song identity based on the neuronal responses of NCM neurons is plotted as a function of the experimental session [before, during, and after (2 and 4 h) pharmacological treatment]. Data for each pharmacological manipulation are grouped by color. Vehicle infusions in NCM (blue) do not alter classification accuracies over time. Local infusions of exogenous E2 (red) significantly enhance performance of the classifier, an effect that persists for at least 2 h following termination of hormone infusion. Impairment of local E2 signaling through either blockade of estrogen receptors with ICI (green) or local inhibition of aromatase with FAD (gray) significantly decreased auditory coding. These effects were also persistent (2 h), but returned to predrug levels 4 h following the interruption of drug infusions. B, The impact of the actions of local E2 on the effectiveness of the neuronal coding of songs of NCM was independently validated with estrogen receptor modulator TMX (green) and aromatase inhibitor ATD (gray). Note that local blockade of estrogen receptors or suppression of the local production of estradiol produces a significant and persistent decrease in the performance of the decoder. **p < 0.01; ***p < 0.001.
Our decoder performed with 81.3 ± 3.4% (mean ± SE) accuracy when trained with half of the stimulation trials recorded in the predrug session (Fig. 4A, blue line). Classification accuracies were not significantly affected by sustained vehicle infusions (drug session: 79.8 ± 2.9%; p = 0.76, Student's t test). Moreover, no changes in the performance of our decoder were detected when tested with neural responses obtained 2 h (Off + 2 h session) or 4 h (Off +4 h session) following the interruption of vehicle infusions (80.5 ± 3.1% and 76.9 ± 3.8%, respectively; p = 0.81 and p = 0.09, Student's t test, for Off + 2 h and Off + 4 h groups, respectively) (Fig. 4A).
To further assess whether local injections of vehicle influence the effectiveness of the neural coding of NCM neurons, we computed MI between the stimulus estimated from the neural activity (estimated via a logistic regression; see Materials and Methods) and the neural responses, a metric that quantifies how much information is transmitted on average about a stimulus. Information rates were not impacted by vehicle treatment in NCM neurons (Fig. 5A). Whereas MI approached 0.25 ± 0.05 bits (mean ± SE) before vehicle infusions, these rates were 0.24 ± 0.03 bits during vehicle infusions (p = 0.14, Student's t test). MI rates were also not significantly different from control (preinjection) levels when birds were stimulated 2 h (Off + 2 h group) or 4 h (Off + 4 h group) after interruption of vehicle infusions (0.24 ± 0.04 bits and 0.24 ± 0.03 bits; p = 0.09 and p = 0.21, Student's t test, for Off + 2 h and Off + 4 h groups, respectively) (Fig. 5A).

Locally generated estradiol increases the information transmitted about songs by NCM neurons. The MI between stimulus and neural responses is plotted as a function of the experimental session [before, during, and after (2 and 4 h) pharmacological treatment]. A–F, Each graph set shows mean MI data obtained under different pharmacological conditions that included vehicle (A), E2 (B), estrogen receptor antagonists ICI (C), and TMX (D), as well as inhibitors of aromatase activity FAD (E) and ATD (F). Note that information rates are not impacted by local vehicle infusions into NCM (A). Significant and persistent increases in MI rates are observed as a function of local E2 infusions (B). In contrast, these values are markedly decreased below predrug levels after blockade of estrogen receptors (C, D) or inhibition of the local production of E2 in NCM (E, F). Similar to estrogen receptor antagonism, local inhibition of aromatase yields effects on information rates that persist for several hours after interruption of drug treatment. *p < 0.05; **p < 0.01; ***p < 0.001.
Together, these data indicate that our classifier performs robustly and can reliably extract song identity from NCM neuronal responses. Furthermore, these control studies indicate that vehicle infusions do not drive short- or long-term changes in the ability of NCM neurons to carry information about, and discriminate among, the songs tested.
Increases in estradiol levels enhance auditory coding
We next tested whether local infusions of E2 affected auditory coding of NCM neurons. To this end, we recorded song-driven responses before, during, and after unilateral E2 infusions in NCM, and assessed the performance of our decoder, as detailed above (vehicle was infused contralaterally). We also examined the ability of NCM neurons to discriminate the different songs tested by generating confusion matrices that reveal the Euclidean distance between training and test vectors for each song, before, during, and after E2 treatment. While classification accuracies reached 79.3 ± 2.6% in the preinjection session, a significant increase in the decoder's performance to 88.6 ± 2.9% was detected during E2 treatment (p < 0.001, Student's t test) (Figs. 4A, red line, 6A,B). The E2-driven increase in classification accuracies directly reflected an enhancement in the neural discrimination of songs of NCM (i.e., decrease in values plotted off-diagonally in the confusion matrices for each cell) (Fig. 6A). This E2-mediated increase in coding effectiveness persisted for at least 2 h following the interruption of exogenous E2 infusions (Figs. 4A, 6A,B). Specifically, classification accuracy in the Off + 2 h condition (86.9 ± 3.6%) was significantly higher than in the pre-E2 condition (p = 0.002, Student's t test), but was not different from data obtained during E2 infusions (p = 0.38, Student's t test) (Figs. 4A, 6A,B). Finally, these coding changes largely subsided 4 h following E2 treatment (Off + 4 h: 80.5 ± 2.4%; p = 0.071 vs control levels, Student's t test) (Figs. 4A, 6A,B).

Estradiol enhances the neural discrimination of songs. A–F, Two-dimensional confusion matrices and classifier's performance for three representative NCM neurons treated with E2 (A, B), ICI (C, D), and FAD (E, F). Top panels show confusion matrices measured from classifications of the neural data before, during, and after (2 and 4 h) pharmacological treatment, and provide an index of the ability of NCM neurons to discriminate among the different songs tested. Each matrix plots the Euclidean distance, coded in grayscale values, between actual (y-axis) and predicted (x-axis) songs. A perfect decoder (100% accuracy) would yield a diagonal confusion matrix, with no off-diagonal values (song misidentifications). Note that the pattern classifier reliably decodes song identity in the predrug condition. Bottom panels show classification accuracies for each representative neuron; dashed lines indicate data obtained for each song individually, and the solid black line shows data for all songs combined. A, B, Local infusions of E2 in NCM improve the neural discrimination of songs (A) and, consequently, significantly increase classification accuracies from preinfusion levels for all neurons (B). These changes persist for at least 2 h after interruption of E2 infusions. C, D, Local blockade of estrogen receptors with ICI leads to a significant and persistent disruption in the ability of NCM neurons to discriminate across songs (C), and in the performance of the decoder (D). These changes persist for at least 2 h after interruption of E2 infusions but return to predrug levels after 4 h. E, F, Suppression of the local production of estradiol with FAD drives a marked and lasting disruption in the structure of confusion matrices, indicating increased errors in song discrimination based on the neural responses (E). FAD significantly decreases the decoder's performance for all representative neurons (F).
To further assess how E2 impacts the ability of NCM neurons to encode stimulus information, we calculated MI rates before, during, and after (2 and 4 h) E2 infusions. We found that E2 significantly and rapidly increased MI rates of NCM neurons (pre-E2 session: 0.22 ± 0.03 bits; E2 session: 0.33 ± 0.06 bits; p < 0.001, Student's t test) (Fig. 5B). Consistent with our findings using the decoding method, increased MI rates were detectable 2 h, but not 4 h, following the interruption of E2 infusions (Off + 2 h: 0.34 ± 0.06 bits; Off + 4 h: 0.21 ± 0.04 bits; p = 0.004 and p = 0.085 from control levels, respectively, Student's t test) (Fig. 5B).
To test for the specificity of the effect of estradiol on auditory coding, we repeated the studies above, except that 5β-DHT or E1 were substituted for E2. 5β-DHT is a nonaromatizable androgen that is synthesized in the zebra finch brain, but is inactive at both androgen and estrogen receptors (Freking et al., 1998). Similar to aromatase, the synthetic enzyme of 5β-DHT (5β-reductase) is expressed at relatively high levels in the songbird forebrain, uses testosterone as a substrate, and has been recently used as a control in recent in vivo microdialysis behavioral pharmacological studies aimed at exploring the contributions of local (NCM) estradiol production in shaping auditory preferences in zebra finches (Remage-Healey et al., 2010). Thus, the use of 5β-DHT enabled us to both control the specificity of the effects of estradiol and to obtain the neurophysiological correlates of the in vivo behavioral pharmacology studies of Remage-Healey et al. (2010). Finally, similar to E2, E1 is synthesized from androstenedione by aromatase.
We found that infusions of 5β-DHT did not impact the performance of our decoder (pre-5β-DHT: 80.6 ± 7.2%; 5β-DHT session: 76.1 ± 5.6%; Off + 2 h: 73.9 ± 6.4%; Off + 4 h: 77.7 ± 5.9%; all p > 0.14 relative to predrug session, Student's t test) (Fig. 7A). Likewise, 5β-DHT did not affect MI rates of NCM neurons (pre-5β-DHT: 0.23 ± 0.03 bits; 5β-DHT session: 0.24 ± 0.02 bits; Off + 2 h: 0.26 ± 0.03 bits; Off + 4 h: 0.24 ± 0.02 bits; all p > 0.11 relative to predrug session, Student's t test) (Fig. 7B). Notably, infusions of E1 also failed to affect the classifier's performance (pre-E1: 78.2 ± 6.7%; E1 session: 81.6 ± 6.1%; Off + 2 h: 77.1 ± 7.3%; Off +4 h: 79.2 ± 6.3%; all p > 0.07 relative to predrug session, Student's t test) (Fig. 7C). Similarly, no changes in MI rates were detected as a result of E1 treatment (pre-E1: 0.25 ± 0.02 bits; E1 session: 0.27 ± 0.03 bits; Off + 2 h: 0.25 ± 0.02 bits; Off +4 h: 0.26 ± 0.02 bits; all p > 0.08 relative to predrug session, Student's t test) (Fig. 7D). These results indicate that auditory coding effectiveness and the neural discrimination of songs in NCM are not broadly impacted by steroidal molecules, but rather are selectively shaped by the actions of estradiol.

5β-DHT and E1 do not impact the effectiveness of the neural coding of, or the information transmitted about songs by, NCM neurons. A, The ability of the decoder to uncover song identity based on the neural responses in NCM is plotted as a function of the experimental session [before, during and after (2 and 4 h) 5β-DHT treatment]. No differences in the performance of the classifier were detected during or after 5β-DHT treatment. B, MI between neural responses and stimulus is plotted as a function of experimental session [before, during, and after (2 and 4 h) 5β-DHT infusions into NCM]. MI rates did not change as a result of the local treatment of NCM neurons with 5β-DHT. C, No changes in the performance of the classifier were detected during or after treatment of NCM neurons with E1. D, MI rates were also unaffected by E1 treatment, indicating that coding properties of NCM neurons are not broadly modulated by steroidal hormones, but rather are selectively shaped by estradiol.
Overall, the data detailed above indicate that one of the functional outcomes of estradiol's modulation of auditory processing in NCM is to increase the availability of information to these central auditory neurons, thereby enhancing auditory coding and the neural discrimination of songs.
Blockade of estradiol's local actions disrupts auditory coding
We next examined whether interference with the physiological actions of estradiol in NCM impacts the neural coding of songs. To this end, we infused ICI, a high-affinity estrogen receptor antagonist, in NCM. As with the studies above, we recorded song-evoked single-unit responses in NCM before, during, and after ICI infusions in awake animals (Fig. 1A). While unilateral infusions of vehicle did not affect the neural coding and discrimination of songs, as assessed by our decoder, contralateral application of ICI to NCM neurons significantly decreased the effectiveness of the neural coding of songs. Specifically, we found that classification accuracies significantly decreased from 81.4 ± 2.8% in the predrug session, to 46.9 ± 4.1% during ICI treatment, an effect that reflects a 42.4% drop in the performance of our decoder (p = 0.006, Student's t test) (Figs. 4A, green line, 6C,D). Blockade of estrogen receptors with ICI also led to a marked disruption in the structure of confusion matrices, indicating that the neural discrimination of songs was severely affected by this pharmacological manipulation (Fig. 6C, note off-diagonal structure). The effects of ICI lasted for at least 2 h (Off + 2 h: 51.4 ± 5.6%; p = 0.003 from predrug levels, Student's t test), but not 4 h after interruption of drug infusion (Off + 4 h: 80.3 ± 3.6%; p = 0.44 relative to predrug levels, Student's t test) (Figs. 4A, 6D). Consistent with these findings, the structure of confusion matrices remained disrupted 2 h, but returned to control profiles 4 h, following ICI treatment, suggesting that blockade of estrogen receptors exerts detrimental effects on the neural discrimination of songs by NCM (Fig. 6C).
We next quantified how much information is transmitted about songs by NCM neurons, by computing MI rates before, during, and after ICI treatment. MI rates significantly decreased as a result of ICI infusions (predrug: 0.25 ± 0.09 bits; ICI session: 0.09 ± 0.05 bits; p = 0.002, Student's t test) (Fig. 5C). The effects of ICI on MI rates persisted for 2 h, but not 4 h, following the end of drug infusions (Off + 2 h: 0.11 ± 0.05 bits; Off + 4 h: 0.26 ± 0.1 bits; p = 0.003 and p = 0.59 from control levels, respectively, Student's t test) (Fig. 5C).
To independently assess the effects of estrogen receptor blockade on the neural coding of songs of NCM, we repeated the experiments above, except that TMX, an estrogen receptor modulator that acts as an antagonist in NCM (Tremere et al., 2009), was locally infused. As was observed for ICI, local injections of TMX markedly decreased mean classification accuracy (predrug: 76.6 ± 4.3%; TMX session: 42.7 ± 5.3%; p = 0.008, Student's t test) (Fig. 4B, green line). Importantly, TMX-mediated effects also lasted for at least 2 h, but returned to control levels 4 h after cessation of drug infusions (Off + 2 h: 40.8 ± 5.6%; Off + 4 h: 79.3 ± 5.8%; p = 0.006 and p = 0.091 from control levels, respectively, Student's t test) (Fig. 4B). TMX also significantly impacted MI rates (predrug: 0.25 ± 0.03 bits; TMX session: 0.13 ± 0.02 bits; Off + 2 h: 0.14 ± 0.03 bits; p = 0.005 and p = 0.017, for drug and Off + 2 h sessions relative to predrug values, respectively, Student's t test). Predrug MI values, however, were detectable 4 h following stoppage of TMX infusion (Off + 4 h: 0.23 ± 0.02 bits; p = 0.55 from control levels, Student's t test) (Fig. 5D).
Overall, these findings show that local blockade of estrogen receptors, and therefore, interference with the physiological actions of E2 in a primary sensory area, largely and rapidly impairs the effectiveness of auditory coding, and the neural discrimination of behaviorally relevant communication signals in the adult brain.
Brain-generated estradiol enhances auditory coding and the neural discrimination of songs
The findings detailed above suggest that estradiol locally regulates the auditory coding of songs in NCM. However, the source of estradiol required for the modulation of NCM (locally produced vs circulating) remains unclear. NCM neurons express high levels of the estrogen-synthetic enzyme aromatase (Shen et al., 1995; Metzdorf et al., 1999; Saldanha et al., 2000; Pinaud et al., 2006). Estradiol levels in the NCM of behaving animals increase as a result of auditory experience (Remage-Healey et al., 2008), and we showed that locally produced estradiol enhances the gain of NCM neurons in alert animals (Tremere et al., 2009), a finding that has been recently validated independently in anesthetized birds (Remage-Healey et al., 2010). It is, therefore, plausible that locally produced E2 shapes the effectiveness of the neural coding of songs in NCM.
To directly address this possibility, we unilaterally pretreated the NCM of awake animals (30 min before stimulus onset) with a loading dose of FAD, a selective aromatase inhibitor known to significantly suppress E2 synthesis in songbird tissue, including the NCM of behaving animals (Wade et al., 1994; Remage-Healey et al., 2008, 2010). We subsequently recorded song-driven responses during maintenance doses of FAD (see Materials and Methods, Fig. 1B); vehicle was infused contralaterally. In essence, this manipulation unilaterally suppresses the local production of estradiol in NCM, while maintaining the contralateral and gonadal production of steroids intact.
FAD significantly decreased the performance of our classifier from 78.5 ± 4.6% in the predrug session to 37.5 ± 4.4% during FAD infusions (p < 0.001, Student's t test) (Figs. 4A, gray line, 6E,F). This effect corresponded to an average decrease of 52.2% in the decoder's performance, which concomitantly occurred with a disruption in the neural discrimination of songs by NCM (Fig. 6E). The effects of FAD on the decoder's performance, and the ability of NCM neurons to discriminate across songs, persisted for at least 2 h (Off + 2 h: 48.6 ± 5.1%; p = 0.0014 and p = 0.128 relative to pre-FAD and FAD sessions, respectively, Student's t test), but not 4 h after interruption of FAD infusions (Off + 4 h: 81.1 ± 4.8%; p = 0.091 from control levels, Student's t test) (Figs. 4A, gray line, 6F).
To further assess the effects of FAD in the neural coding of songs, we computed MI before, during, and after drug treatment. We found that MI rates decreased by 41% as a consequence of FAD infusions in NCM (predrug: 0.27 ± 0.06 bits; FAD session: 0.09 ± 0.05 bits; p = 0.004, Student's t test) (Fig. 5E). Interestingly, although E2 levels can be rapidly reinstated in NCM after blockade of aromatase activity (Remage-Healey et al., 2008), we found that FAD effects in NCM's neuronal coding were detectable 2 h, but not 4 h, after interruption of drug infusions (Off + 2 h: 0.1 ± 0.02 bits; Off + 4 h: 0.28 ± 0.05; p < 0.001 and p = 0.55 from predrug levels, respectively, Student's t test).
Next, we independently verified these findings. To this end, we pretreated NCM with ATD, a specific inhibitor of aromatase activity (Wade et al., 1994) (Fig. 1B). We previously showed that suppression of the local production of estradiol with ATD markedly decreased the gain of hearing-driven responses in the NCM of awake songbirds (Tremere et al., 2009). Blockade of local estradiol production in NCM with ATD reproduced the effects of FAD. In particular, ATD treatment significantly decreased classification accuracy (predrug: 81.3 ± 4.6%; ATD session: 34.5 ± 4.4%; p < 0.001, Student's t test) (Fig. 4B, gray line). Similar to our FAD findings, TMX also drove changes in the neuronal coding of NCM that were detectable for at least 2 h, but not 4 h, after interruption of the pharmacological treatment (Off + 2 h: 39.1 ± 5.1%; Off + 4 h: 71.2 ± 4.8%; p = 0.003 and p = 0.64, from control levels, respectively, Student's t test) (Fig. 4B).
To further quantify the effects of ATD on the neural coding of songs, we also computed MI rates from our neural data. Whereas predrug rates averaged 0.26 ± 0.03 bits, treatment with ATD significantly decreased these values to 0.08 ± 0.02 bits (p = 0.006, Student's t test) (Fig. 5F). ATD-mediated effects on MI rates persisted for 2 h, but returned to predrug levels 4 h, after interruption of drug infusions (Off + 2 h: 0.09 ± 0.02 bits; Off + 4 h: 0.23 ± 0.03 bits; p = 0.008 and p = 0.39 from control levels, respectively, Student's t test) (Fig. 5F).
Together, these findings conclusively demonstrate that estradiol produced by a central auditory area promotes a rapid and robust enhancement of auditory coding in the adult, alert vertebrate brain. Furthermore, our observations indicate that a brain-generated estrogen enhances the ability of auditory neurons to discriminate across natural, communication-based sensory signals. Finally, our data suggest that the effects of locally generated estradiol on the neural coding of songs in NCM are long lasting, and persist for at least 2 h.
Estradiol directly affects the ability of NCM neurons to discriminate songs
The data shown above suggest that brain-generated estradiol regulates the neural discrimination of songs in NCM. Importantly, this conclusion is partly derived from findings indicating that the decoder's performance is significantly decreased when it is trained with predrug trials and tested with trials obtained during, or after, blockade of estrogen receptors or inhibition of aromatase (e.g., ICI or ICI Off + 2 h). An alternative interpretation for these findings is that, even though our pharmacological manipulations may alter the response properties of NCM neurons (thus leading to changes in classification performance), such changes may occur in the absence of actual alterations in the neural discrimination of songs. To directly address this issue and quantify the extent to which estradiol influences the ability of NCM neurons to discriminate songs, we computed the decoder's performance when training and testing it with neural data obtained within the same drug condition and pharmacological sessions. More specifically, we trained the classifier with half of the trials associated with each drug session and tested it with the remaining half of the trials of that session (e.g., trained the classifier with half of the E2 Off + 2 h trials, and tested it with the remaining half of the trials of this very session).
We found that our decoder performed with 80.1 ± 3.1% accuracy when trained with half of the trials recorded during vehicle infusions and tested with the remaining trials of the same session (Fig. 8A). When trained and tested with data obtained 2 h (Off + 2 h) or 4 h (Off + 4 h) after stoppage of vehicle infusion, classification accuracies were 79.4 ± 3.0% and 78.9 ± 2.8%, respectively. Performance of the decoder in each of these conditions was not significantly different from the predrug states (all p > 0.08, Student's t test).

NCM-derived estradiol directly enhances the neuronal discrimination of songs by NCM. The ability of the decoder to identify stimulus identity based on the responses of NCM neurons was probed when the classifier was both trained and tested with trials obtained within each session of interest (e.g., trained with half of the trials within the E2 session, and tested with the remaining trials of that very same session; see Materials and Methods, and Results). This study directly test whether drug-induced alterations in the responses of NCM neurons can occur in the absence of changes in their neural ability to discriminate songs, as assessed by the performance of our decoder. In each graph set, the decoder's performance (y-axis) is plotted as a function of the recording session (predrug, drug, Off + 2 h, or Off + 4 h; x-axis), for each pharmacological intervention of interest. A, We found that vehicle infusions did not influence the performance of the classifier in the short or long term (up to 4 h). B, In contrast, E2 treatment significantly increased classification accuracies. This effect was long lasting and persisted for at least 2 h after interruption of exogenous infusions of hormone. C, D, Blockade of estrogen receptors with ICI (C) or TMX (D) markedly suppressed the performance of the decoder. This effect persisted for 2 h, but not 4 h, after cessation of drug infusions. E, F, Inhibition of aromatase in NCM via local infusions of FAD (E) or ATD (F) markedly decreased the ability of NCM neurons to discriminate songs, as assessed by the classifier's performance. Similar to the findings obtained with estrogen receptor blockade, inhibition of aromatase drove long-lasting changes, which persisted for at least 2 h, in the neuronal discrimination of songs of NCM. G, H, Infusions of 5β-DHT (G) or E1 (H) into NCM did not affect the short- or long-term performance of the decoder, indicating that estradiol selectively affected the neural discrimination of songs in NCM neurons. *p < 0.05; **p < 0.01; ***p < 0.001.
Interestingly, increases in E2 levels by exogenous infusions of this hormone in NCM led to a significant enhancement in classification accuracies. When trained and tested with neural data obtained during E2 infusions, our decoder performed at 91.4 ± 2.8% accuracy (p = 0.006 relative to predrug levels) (Fig. 8B). This effect persisted for 2 h, but not 4 h, after E2 treatment (Off + 2 h: 88.6 ± 3.2%; Off + 4 h: 80.1 ± 3.1%; p = 0.03 and p = 0.08 relative to preinfusion levels, respectively, Student's t test).
Local blockade of estrogen receptors not only altered NCM neuronal responses, but also directly affected how well NCM neurons discriminate songs. When trained and tested with data obtained during the same experimental session, we observed marked changes in the performance of our decoder. Classification accuracies were significantly decreased during ICI infusions; this effect also persisted for 2 h, but not 4 h, after drug treatment (ICI session: 41.6 ± 3.9%; Off + 2 h: 44.2 ± 4.2%; Off + 4 h: 76.4 ± 3.3%; p = 0.003, p = 0.005 and p = 0.09 relative to pre-ICI levels, respectively, Student's t test) (Fig. 8C). Similar findings were obtained when estrogen receptors were blocked with TMX (TMX session: 44.6 ± 4.8%; Off + 2 h: 43.9 ± 3.9%; Off + 4 h: 78.2 ± 3.3%; p = 0.02, p = 0.009 and p = 0.07 relative to pre-TMX levels, respectively, Student's t test) (Fig. 8D).
We next tested whether suppression of the local production of estradiol, via inhibition of aromatase, regulates the neural discrimination of songs by NCM neurons. To this end, we trained and tested our decoder with neural responses obtained during FAD treatment. We also tested the extent to which these effects are long lasting by training and testing the classifier with data obtained 2 and 4 h after interruption of drug infusion. We found that aromatase inhibition with FAD significantly impairs the ability of NCM neurons to discriminate songs, as revealed by marked decreases in the performance of our decoder. Specifically during FAD infusions, our decoder performed with 35.5 ± 4.4% accuracy (p < 0.001 relative to pre-FAD levels, Student's t test) (Fig. 8E). This effect persisted for 2 h, but not 4 h, after interruption of FAD infusions (Off + 2 h: 41.3 ± 4.1%; Off + 4 h: 79.1 ± 2.8%; p = 0.006 and p = 0.11 relative to pre-FAD levels, respectively, Student's t test). Independent validation of these findings was obtained with ATD, which also significantly decreased the performance of our classifier (ATD session: 38.5 ± 4.1%; Off + 2 h: 34.9 ± 3.6%; Off + 4 h: 78.4 ± 2.9%; p = 0.007, p = 0.004 and p = 0.09 relative to pre-ATD levels, respectively, Student's t test) (Fig. 8F).
Finally, we reassessed the specificity of the effects of E2 on the neural discrimination of songs. To this end, we trained and tested our classifier with neural responses obtained during or after either 5β-DHT or E1 treatment. We found that neither the androgen nor the estrogen affected the decoder's performance (5β-DHT session: 79.3 ± 5.6%; 5β-DHT Off + 2 h: 78.7 ± 5.5%; 5β-DHT Off + 4 h: 79.8 ± 6.3%; all p > 0.07 relative to pre-5β-DHT levels; E1 session: 77.4 ± 5.8%; E1 Off + 2 h: 79.1 ± 5.5%; E1 Off + 4 h: 76.8 ± 5.1%; all p > 0.08 relative to pre-E1 levels, Student's t test) (Fig. 8G,H).
In summary, the findings above indicate that E2, and more specifically E2 produced in NCM, directly regulates the neural discrimination of songs by NCM neurons. Our results also show that these effects are long lasting and persist for at least 2 h after interruption of pharmacological treatment.
Estradiol regulates the neural discrimination of songs by shaping both rate and temporal coding
We next set out to determine how brain-generated estradiol enhances the neural coding of songs. Given that local estradiol production increases the discharge rates of NCM neurons, it is plausible that the effects of E2 on spike rates of NCM neurons (rate code) directly determine a neuron's coding and discrimination ability. If this interpretation is correct, elimination of the effects of estradiol on neuronal spike rates should eliminate the changes in coding driven by this neurohormone. To directly test this possibility, we obtained song-driven spike trains during E2 infusions (as well as Off + 2 h and Off + 4 h), normalized these spike rates to pre-E2 infusion levels by randomly extracting spikes across the train, and then reassessed the performance of our decoder when tested with these processed spike trains (Fig. 9A,B). Importantly, this approach enabled us to segregate effects of estradiol on rate versus temporal coding of songs.

Estradiol-dependent effects in neural coding cannot be fully explained by increases in firing rates. A, B, Illustration of spike extraction and insertion manipulations. A, Schematic representation of song-evoked spike trains obtained from an NCM neuron during infusions of vehicle (blue), estradiol (red), estrogen receptor antagonists ICI or TMX (green), and aromatase inhibitors FAD or ATD (gray). Illustrated are the typical effects of these agents: vehicle does not affect firing rates. E2 robustly increases discharge rates of NCM neurons. In contrast, blockade of estrogen receptors or inhibition of aromatase activity decrease NCM neuronal firing rates. For illustrative purposes, we suggest that the number of action potentials in the spike train increases from 10 to 20 during E2 treatment and decreases from 10 to 6 during ICI/TMX or FAD/ATD treatment (see n values on the right side of spike trains). B, We normalized the firing rates of NCM neurons during pharmacological treatment by obtaining neurophysiological responses and computationally processing individual spike trains to match the number of spikes in the drug session with that in the predrug session (see Materials and Methods). The diagram illustrates the two types of manipulations implemented. For E2 infusions (red), which increase NCM neuronal spike rates, we randomly eliminated spikes from the train (asterisks) to match the spike numbers of the average predrug session. For ICI/TMX or FAD/ATD infusions (green and gray, respectively), which decrease NCM neuronal spike rates, we inserted spikes at random spike times in the train (asterisks) to match the spike numbers of the average predrug session (see Materials and Methods). Note the n values on the right side of spike trains, and compare them against the n values in A. This spike normalization process eliminates the effects of pharmacological agents on NCM neuronal firing rates. These manipulations were also performed in the same manner for Off + 2 h and Off + 4 h sessions (not illustrated). C–H, The performance of the classifier in decoding the identity of the song stimulus based on the neuronal responses of NCM neurons is plotted as a function of the experimental session [before, during, and after (2 and 4 h) pharmacological treatment]. Data obtained with intact (not normalized) and processed (normalized) spike trains are shown for each drug group in solid and dashed lines, respectively. Note that although the normalization process (and consequently elimination of firing rate effects) diminishes the effects of all pharmacological agents on the decoding performance, this manipulation does not eliminate drug effects on NCM's neuronal coding of songs. These findings indicate that E2 affects both rate and temporal coding of auditory signals in NCM neurons. *p < 0.05; **p < 0.01; ***p < 0.001.
Notably, classification accuracies were not impacted by normalization of spike rates in the vehicle condition (Fig. 9C). This manipulation, however, decreased the enhancement of classification accuracies seen during estradiol treatment from 88.6 ± 2.9% to 81.6 ± 2.2% (Fig. 9D). Remarkably, these normalized spike trains, obtained during E2 treatment, still yielded classification accuracies that were significantly higher than vehicle and pre-E2 levels (78.9 ± 3.4% and 76.9 ± 2.6%, respectively; all p < 0.05, Student's t test). Classification accuracies were also significantly higher than pre-E2 levels in normalized spike trains obtained 2 h (Off + 2 h: 82.3 ± 2.0%; p = 0.02, Student's t test), but not 4 h, after E2 treatment (Off + 4 h: 78.4 ± 2.4%; p = 0.19, Student's t test) (Fig. 9D). Even though the increase in classification accuracies mediated by E2 in normalized spike trains was not as robust as that detected in intact spike trains, these findings clearly demonstrate that increases in spike rate cannot fully account for the E2-mediated changes in the neural coding of songs.
Normalization of firing rates during blockade of estrogen receptors with ICI and TMX (addition of spikes with random spike timings; see Materials and Methods) (Fig. 9A,B) revealed marked changes in classification accuracies. Specifically, normalized trains obtained during ICI and TMX infusions yielded classification accuracies that were significantly lower than predrug levels (ICI: 62.5 ± 3.8%; TMX: 62.3 ± 4.4%; all p < 0.05 relative to predrug levels, Student's t test) (Fig. 9E,F). These effects in the neural coding of normalized spike trains were still detectable 2 h (ICI Off + 2 h: 63.4 ± 4.3%; TMX Off + 2 h: 65.1 ± 4.0%; all p < 0.05 relative to predrug levels, Student's t test), but not 4 h, after interruption of pharmacological treatment (ICI Off + 4 h: 78.6 ± 3.2%; TMX Off + 4 h: 73.2 ± 3.3%; all p > 0.08 relative to predrug levels, Student's t test) (Fig. 9E,F). The findings above show that the general pattern of changes in the effectiveness of auditory coding by NCM neurons, resulting from blockade of estrogen receptors, is conserved for intact versus normalized spike trains. Notably, by eliminating drug effects on firing rates through normalization, the magnitude of coding changes observed with either antagonist was reduced, albeit still statistically significant.
We next assessed whether the enhancement in the neural coding of songs elicited by locally generated estradiol is mediated by increases in NCM neuronal discharge rates. To this end, we normalized spike rates of NCM neurons during FAD and ATD treatment to predrug levels and assessed the classifier's performance when tested with these processed spike trains (Fig. 9A,B). Normalization of discharge rates during aromatase inhibition with FAD and ATD did not abolish the robust decrease in classification accuracies induced by both agents. To the contrary, normalized spike trains collected during FAD and ATD infusions revealed classification accuracies that were markedly lower than those observed during the predrug session (FAD: 58.6 ± 4.1%; ATD: 61.7 ± 4.8%; all p < 0.05 relative to predrug levels, Student's t test) (Fig. 9G,H). Such drug-induced effects on normalized spike trains were also detectable at 2 h (FAD Off + 2 h: 56.7 ± 4.4%; ATD Off + 2 h: 58.5 ± 5.1%; all p < 0.05 relative to predrug levels, Student's t test), but not 4 h, after stoppage of drug infusions (FAD Off + 4 h: 74.2 ± 4.6%; ATD Off + 4 h: 75.2 ± 3.6%; all p > 0.12 relative to predrug levels, Student's t test) (Fig. 9G,H).
Together, these findings directly demonstrate that locally generated estradiol enhances the neuronal coding of songs of NCM, in part by increasing neuronal discharge rates. However, given that normalization of spike rates does not abolish the effects of estradiol on the neural discrimination of songs, we postulate that this neurohormone may also shape NCM's temporal coding of songs.
Brain-generated estradiol selectively and robustly enhances the behavioral discrimination songs
Our findings provide strong evidence that estradiol produced in NCM shapes the effectiveness of auditory coding to enhance the neural discrimination of behaviorally relevant acoustic signals. It is unclear, however, whether this E2-driven enhancement of the neural discrimination of songs translates into improved behavioral performance in auditory-based tasks. To directly address this question, we combined bilateral intracerebral pharmacological manipulations in awake animals and standard auditory discrimination tasks developed for songbirds (Clayton, 1988; Riebel et al., 2002; Riebel and Smallegange, 2003; Gobes and Bolhuis, 2007; Remage-Healey et al., 2010). It is well established that adult zebra finches exhibit marked behavioral preference for the song of their tutor relative to a novel, previously unheard conspecific song (Riebel et al., 2002; Riebel and Smallegange, 2003; Bolhuis and Gahr, 2006; Gobes and Bolhuis, 2007; Holveck and Riebel, 2007). Moreover, the strength of this behavioral preference is strongly correlated when comparing operant- and preference-based tests (Holveck and Riebel, 2007). We, therefore, used a two-choice preference test to determine whether discrimination of TUT versus a novel CON song is impacted by estradiol's local modulation in NCM. This test requires birds to encode, process, and discriminate two songs (TUT vs CON), and take an appropriate behavioral output (fly to the chamber playing the TUT song) (see Materials and Methods).
Before intracerebral pharmacological manipulations, birds assigned to all groups exhibited marked behavioral preference toward the TUT song (predrug session: 76.9 ± 2.9%, mean ± SE). Moreover, song preference in animals assigned to the control (vehicle) group did not significantly differ from birds assigned to each experimental group (ICI, TMX, FAD, or ATD; all p > 0.11, ANOVA).
Bilateral blockade of estrogen receptors with ICI largely disrupted TUT song preference. Specifically, although a marked preference for TUT song was observed in birds bilaterally injected with vehicle (77.7 ± 3.1%), this selectivity was largely abolished in animals bilaterally infused with ICI (53.9 ± 2.6%; p < 0.001, ANOVA) (Fig. 10A).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Brain-generated estradiol impacts the auditory processing in NCM to shape behavioral preference to songs, but not calls, in freely behaving animals. A, Bilateral infusions of vehicle into the NCM do not affect the strong behavioral preference of adult birds to tutor over conspecific song. However, bilateral blockade of estrogen receptors with ICI or TMX, or suppression of the local production of E2 with FAD or ATD, largely abolish song preference. B, Bilateral infusions of vehicle, estrogen receptor antagonists ICI and TMX, and aromatase inhibitors FAD and ATD into NCM do not change call preference in freely behaving animals. Horizontal gray lines indicate no preference. *p < 0.05; **p < 0.01; ***p < 0.001.
To independently test the effects of local blockade of estrogen receptors on the behavioral discrimination of songs, we bilaterally injected TMX into NCM, in a separate group of animals. As was observed with ICI, we found that TMX infusions severely disrupted TUT song preference in freely behaving animals, presumably by interfering with the animal's ability to discriminate across songs (TMX: 59.2 ± 4.7%; p = 0.037, ANOVA) (Fig. 10A).
We next assessed whether these behavioral effects were mediated by locally produced estradiol. To this end, we bilaterally pretreated the NCM of a group of birds with FAD and exposed animals to the auditory discrimination task (see Materials and Methods). Similar to observations made with ICI and TMX, we found that suppression of the local production of estradiol in NCM with FAD eliminated TUT song preference (52.1 ± 3.2%; p < 0.001 vs vehicle-injected controls, ANOVA) (Fig. 10A). To independently examine the effects of locally produced E2 in the behavioral discrimination of songs, we bilaterally pretreated the NCM of a separate group of birds with the aromatase inhibitor ATD. As was observed with FAD, ATD treatment largely abolished TUT song preference in behaving animals (50.1 ± 3.7%; p = 0.009 from vehicle-injected controls, ANOVA) (Fig. 10A).
The findings described above suggest that estradiol produced locally in NCM is necessary for the behavioral expression of song preference in an auditory discrimination task. It is possible, however, that interference with the estradiol network in NCM (with ICI, TMX, FAD, and ATD) essentially disrupted auditory processing regardless of the behavioral relevance of the auditory stimuli used. To explore this possibility, we subjected a separate group of male zebra finches to a call discrimination task that is fundamentally similar to the song preference task detailed above, except that long calls from different sexes are presented in place of TUT and CON songs (Gobes and Bolhuis, 2007) (see Materials and Methods). Importantly, long calls and song syllables of zebra finches are highly similar acoustically both in complexity and frequency range. Although male zebra finches are capable of discriminating calls of both sexes, they exhibit a marked behavioral preference to female calls (Vicario et al., 2001; Gobes and Bolhuis, 2007).
We, therefore, tested whether interference with actions of estradiol in NCM also impacts call discrimination. Before local pharmacological manipulations, animals assigned to all groups exhibited robust and statistically identical preference for female calls (call/response ratio = 1.88 ± 0.3, mean ± SE; a ratio of 1 indicates no preference, and ratios >1 indicate preference for female over male calls). Remarkably, bilateral blockade of estrogen receptors with ICI and bilateral local suppression of estradiol synthesis with FAD both failed to affect call discrimination (Fig. 10B). Vehicle-, ICI-, and FAD-treated animals equally preferred female calls (call/response ratios were 1.83 ± 0.2, 1.71 ± 0.3, and 1.67 ± 0.3, respectively; p = 0.14 and p = 0.46 for ICI and FAD groups, relative to controls, respectively, ANOVA).
To independently assess these findings, we bilaterally treated two separate groups of animals with TMX and ATD. Consistent with the findings obtained with ICI and FAD, blockade of estrogen receptors or acute suppression of the local estradiol levels with TMX and ATD, respectively, did not impact the call/response ratios (ratios were 1.76 ± 0.2, 1.65 ± 0.2, and 1.69 ± 0.3 for vehicle-, TMX-, and ATD-treated animals, respectively; p = 0.26 and p = 0.61 for TMX and ATD groups, relative to controls, respectively, ANOVA) (Fig. 10B). These findings indicate that the auditory discrimination of complex sounds is not affected by interference with the actions of estradiol in NCM. Remarkably, locally generated estradiol appears to selectively impact the processing of songs in NCM.
In summary, our findings demonstrate that brain-generated estradiol increases the availability of information in central auditory neurons and, as a result, strongly facilitates the neural discrimination of learned, behaviorally relevant communication signals. These effects of estradiol appear to directly and selectively shape the discrimination and preference of songs at the behavioral level. Importantly, given that, at present, it is impossible to carry out neurophysiological recordings in freely behaving animals performing an auditory discrimination task during bilateral, intracerebral pharmacological manipulations, we are not able to conclusively determine that estradiol-mediated changes in NCM's neural coding of songs causally shape auditory discrimination at the behavioral level. However, our data are highly suggestive that the estradiol-dependent enhancement of the neural discrimination of songs translates into an improved behavioral performance of animals in the song discrimination task detailed above.
Discussion
This study showed that brain-generated estradiol enhances the informational content and resolution of auditory signals encoded by central auditory neurons, thereby rapidly improving the neural discrimination of songs. The positive impact of E2 in the NCM's neuronal performance, via mechanisms that affect both rate and temporal coding, selectively shapes behavioral preferences to song in auditory discrimination tasks. Our observations build upon recent findings that we and others have reported, indicating that brain-generated estradiol is a key regulator of central auditory function in the adult songbird brain (Maney et al., 2006; Remage-Healey et al., 2008, 2010; Svec et al., 2009; Tremere et al., 2009; Vyas et al., 2009; Sanford et al., 2010). More specifically, local action and production of estradiol in NCM directly modulates sensory responses and auditory-based behaviors, on a time-scale that is compatible with rapidly acting neuromodulators. For example, Remage-Healey et al. (2008) reported rapid increases in estradiol levels in NCM as a result of auditory experience in freely behaving animals. We recently demonstrated that local E2 infusions instantaneously increased hearing-driven discharge rates in the NCM of awake zebra finches. Consistent with this view, blockade of estrogen receptors and aromatase inhibition in NCM suppressed hearing-driven neuronal firing rates (Tremere et al., 2009), indicating that that brain-generated estradiol regulates, in real time, the gain of auditory responses in the adult alert brain. Finally, we also recently showed that estradiol facilitates auditory-driven responses in NCM by selectively suppressing local inhibitory transmission via a nongenomic, presynaptic mechanism (Tremere et al., 2009). Here we quantitatively determined a functional relevance of estradiol's modulation of auditory processing. Our findings revealed that the information that NCM neurons carry about stimulus structure can be rapidly and selectively modulated by locally generated estradiol. Functionally, such modulation translates into an enhanced ability of neurons in NCM to discriminate across different stimuli.
We also tested whether the E2 effects on the neuronal physiology of NCM are persistent. Acute and sequential manipulations in the actions of E2 led to persistent changes in neural coding that were apparent for at least 2 h. Importantly, these sustained changes appear to remain beyond an extended “washout” period for the pharmacological effects. The most conservative explanation for our observed results is that degradation of each pharmacological agent used lasted for at least 2 h. Although this possibility can only be conclusively probed with in vivo drug degradation studies, we believe this is an unlikely scenario. For instance, previous studies showed that decreases in the E2 levels in NCM induced by FAD can be recovered within 30 min after FAD treatment (Remage-Healey et al., 2008, 2010). The FAD concentrations used were identical to these previous studies, and, therefore, the kinetics of drug washout/degradation should be nearly indistinguishable. Thus, an alternative possibility is that the enduring changes in the physiology of NCM detected here may be implemented via an experience-dependent engagement of E2-regulated cellular processes that sustain altered neuronal coding properties. One intriguing idea is that E2 promotes synaptic plasticity in NCM circuitry via nongenomic mechanisms. In the rodent hippocampus and hypothalamus, E2 modulates synaptic transmission. For instance, the amplitude of non-NMDA receptor EPSCs is markedly increased by E2 treatment (Wong and Moss, 1992; Gu and Moss, 1996; Woolley, 2007). Furthermore, NMDA-mediated synaptic events and the magnitude of long-term potentiation are rapidly and positively enhanced by E2 (Foy et al., 1999; Foy, 2001; Woolley, 2007). Both NMDA and non-NMDA glutamatergic receptors are expressed in NCM (Wada et al., 2004). Thus, if E2 nongenomically modulates excitatory synaptic physiology in NCM through processes that are akin to those reported for the hippocampus, such mechanisms could provide a synaptic basis for the effects detected here. Estradiol is unlikely to modulate non-NMDA synaptic events in NCM, as we previously showed that CNQX-sensitive spontaneous EPSCs are not impacted by E2 in NCM slices (Tremere et al., 2009). However, it remains to be determined whether E2 affects synaptically evoked NMDA receptor currents in NCM neurons and, if so, whether such a mechanism is necessary for our observed effects. Alternatively, E2-dependent modulation of inhibitory synaptic transmission may provide a mechanistic basis for these sustained effects. We showed that E2 strongly suppresses GABAergic transmission in NCM via a presynaptic, nongenomic mechanism (Tremere et al., 2009), and that GABAA-mediated inhibition is central to the encoding and temporal organization of song-driven responses in NCM (Pinaud et al., 2008b). Whether or not the effects of E2 on inhibitory transmission in NCM causally account for the persistent changes in neural coding is not known at present, and will be a main focus of our future studies.
A final possibility that may contribute to the sustained effects of E2 in the physiology of NCM could involve the engagement of E2-dependent gene expression. In fact, auditory experience both elevates E2 levels (Remage-Healey et al., 2008) and drives complex gene regulatory programs in NCM (Mello et al., 1992; Velho et al., 2005; Huesmann and Clayton, 2006; Pinaud et al., 2008a; Dong et al., 2009). Importantly, we recently showed that local production of estradiol in NCM is both necessary and sufficient to induce the expression of several plasticity-associated genes thought to be required for sensory learning (Tremere et al., 2009). Given that most of the genes previously studied by us, and now known to be regulated by E2, act as transcription factors for genes involved in neurotransmitter release (e.g., synapsins I and II), it is plausible that E2 may drive long-term changes in the molecular machinery involved in synaptic transmission. Alternatively, based on our previous findings that E2 shapes inhibitory transmission (Tremere et al., 2009), it is possible that the long-term “disappearance” of E2-mediated effects (i.e., 4 h after pharmacological challenge) may be mediated by a homeostatic mechanism involving genomic cascades. For instance, inhibitory transmission may be upregulated after 4 h of E2 treatment, which could account for the decreased neurophysiological responses and ability of NCM neurons to discriminate songs. Although future studies will be required to test these possibilities, it is conceivable that these gene expression cascades may trigger and/or sustain E2-mediated functional retuning of NCM neurons, a position that is consistent with earlier reports showing that the selectivity of hearing-driven gene expression responses in NCM can be modulated by circulating E2 levels (Maney et al., 2006), and that manipulations in plasma or local E2 levels markedly shape gene expression in NCM (Tremere et al., 2009; Sanford et al., 2010). Thus, the recently uncovered close interdependence of estradiol levels and plasticity-associated gene expression in NCM highlights protein synthesis-dependent events as possible mechanisms for sustained changes in neural coding of NCM neurons triggered by E2. In short, future studies will determine whether E2 mediates long-term changes in the physiology of NCM by affecting nongenomic and/or genomic mechanisms (McEwen, 2002; Woolley, 2007; Tremere et al., 2009).
Our results also showed that the actions of estradiol in NCM are key to shaping behavioral responses to song, putatively by influencing neural computations associated with auditory discrimination. These observations are congruent with behavioral pharmacology findings from other groups. For example, increased plasma E2 levels accentuate behavioral responses to songs (Svec et al., 2009), particularly those with complex acoustic features (Vyas et al., 2008, 2009). Similarly, inhibition of aromatase in NCM disturbs the capacity of male zebra finches to respond behaviorally to song stimuli (Remage-Healey et al., 2010). We found that acute interference with local estrogenic modulation in NCM dramatically disrupts the behavioral expression of song preference. Remarkably, these observations cannot be accounted for by a general disruption in auditory processing (i.e., the ability to process sounds) but rather reflect a selective impairment in the perceptual processing of learned songs. These data raise the intriguing possibility that NCM neurons in the zebra finch may be tuned to the temporal sequencing of syllables within a song, a process influenced by brain-generated E2, but not these individual elements per se, as in the case of calls. This possibility is supported by findings suggesting that NCM neurons may integrate temporal information of both songs and calls (Terleph et al., 2007), and that E2 modulates temporal and rate response properties of NCM neurons, as revealed by our spike-normalization studies. A caveat to our study, however, is that our normalization method does not account for temporal modulation in firing rates, which still consists of a form of coarse temporal coding, but one that occurs as a result of modulations within the stimulus, as opposed to specific changes in the temporal patterns of spike trains. Nonetheless, our findings suggest that E2 putatively influences both the rate and the temporal organization of song-evoked responses. At any rate, NCM neurons exhibit selectivity to conspecific songs (Mello et al., 1992; Chew et al., 1996; Velho et al., 2005; Pinaud and Terleph, 2008). Given that our manipulations disrupted song, but not call discrimination, our findings raise the intriguing possibility that estradiol may shape the neuronal selectivity of NCM, as suggested by earlier gene expression findings (Maney et al., 2006) and, consequently, guide the behavioral preference to songs. One possible neural substrate required for these selective effects is the noradrenergic system. Noradrenergic projections densely innervate aromatase-positive neurons in NCM and are required for estradiol-dependent auditory behaviors in songbirds (Appeltants et al., 2004; Vyas et al., 2008), suggesting that catecholaminergic and estrogenic circuits may closely interact.
Numerous studies have previously suggested an interaction between estradiol levels and hearing function. For instance, auditory processing was shown to be more efficient in women than men, hearing thresholds oscillate with estradiol levels during the menstrual cycle in women (Jerger and Hall, 1980; Davis and Ahroon, 1982; Walpurger et al., 2004), and Turner's syndrome patients experience an array of hearing-associated pathologies (Hultcrantz et al., 1994; Güngör et al., 2000). Rodents and nonhuman primates subjected to ovariectomies experienced large distortions in auditory brainstem responses that were recovered by estrogen replacement therapy (Coleman et al., 1994; Golub et al., 2004), and ERβ-deficient mice were shown to exhibit severe progressive hearing loss that led to early deafness (Simonoska et al., 2009). Collectively, these findings are consistent with the notion that estradiol's modulation of auditory processing may generalize to mammals. We underscore here, however, that these studies do not causally link estradiol regulation and hearing function, do not segregate whether the source of hormone is peripheral or central, and, importantly, do not separate if this interaction occurs in the cochlea and/or in brain structures. In fact, research findings suggest that aromatase and estrogen receptors are expressed in the cochlea of a variety of vertebrate species (Forlano et al., 2005; Meltser et al., 2008; Noirot et al., 2009; Maruska and Fernald, 2010). If future research confirms this sensory–neuroendocrine interaction in the mammalian auditory cortex, it will be important to consider translational implications of these findings, some of which we highlight here. First, local blockade of estrogen receptors and suppression of E2 synthesis significantly impairs auditory processing in songbirds. It is, therefore, plausible that hearing dysfunction related to estrogen insufficiency, a well documented phenomenon in humans and animal models, may be a process that occurs entirely (or partially) in the brain, as opposed to the prevalent view that altered processing stems from peripheral organ problems (Elkind-Hirsch et al., 1992, 1994; Coleman et al., 1994; Barnes et al., 2006; Hultcrantz et al., 2006; Hederstierna et al., 2009a,b, 2010). This possibility highlights the importance of seriously considering brain-based mechanisms of hearing dysfunction. Second, an array of stressors and age-related processes are known to markedly impact estradiol levels in humans and could potentially influence central auditory function. Third, our findings may have implications for circumstances that drive hormonal shifts in humans, including those that result from genetic disorders (e.g., Turner syndrome), menopause, hormone supplements such as contraceptives, hormone replacement therapy, and antiestrogen chemotherapies, most of which impact auditory function and have direct consequences for human health issues (Hultcrantz et al., 2006; Hederstierna et al., 2010).
Finally, it is important to strongly emphasize that the potential implications outlined above are conjectural, at best, based on data available to date, and that any possible causal interactions between neurosteroidal regulation and auditory processing in the mammalian brain are currently highly tenuous. Our intention is to simply stress how the recent findings obtained in songbirds have opened, and may continue to open, new avenues for the exploration of how hormones modulate neuronal physiology, and may bring a new perspective on the mechanistic bases and operational rules of sensory systems more broadly.
Footnotes
- Received August 19, 2010.
- Revision received December 16, 2010.
- Accepted January 3, 2011.
-
This work was supported by grants from the National Institutes of Health (Grant R01-DC-010181), the National Science Foundation (Grant 1064684), the Schmitt Foundation, and the Reynolds Foundation (to R.P.). We thank Dr. Gregory DeAngelis for help with pattern classification analyses; and Drs. David Knill, Alex Pouget, Gregory DeAngelis, and Ting Qian for insightful discussions. We also thank Dr. Jin Jeong for help with histological analyses; Victor Vuong for help with the behavioral tasks; and Shaista Hussain for efforts in the earlier stages of this work. Finally, we thank the two anonymous reviewers who provided highly constructive recommendations on how to improve our manuscript. R.P. designed the research; L.A.T. and R.P. performed the research; L.A.T. and R.P. analyzed the data; R.P. wrote the paper.
- Correspondence should be addressed to Dr. Raphael Pinaud, Department of Physiology and Reynolds Oklahoma Center on Aging, 975 N.E. 10th Street, BRC-West, Room 1309B, University of Oklahoma Health Sciences Center, Oklahoma City, OK 73104. raphael-pinaud{at}ouhsc.edu
- Copyright © 2011 the authors 0270-6474/11/313271-19$15.00/0