
Abstract
Vestibular ganglion neurons (VGNs) transmit information along parallel neuronal pathways whose signature distinction is variability in spike-timing; some fire at regular intervals while others fire at irregular intervals. The mechanisms driving timing differences are not fully understood but two opposing (but not mutually exclusive) hypotheses have emerged. In the first, regular-spiking is inversely correlated to the density of low-voltage-gated potassium currents (IKL). In the second, regular spiking is directly correlated to the density of hyperpolarization-activated cyclic nucleotide-sensitive currents (IH). Supporting the idea that variations in ion channel composition shape spike-timing, VGNs from the first postnatal week respond to synaptic-noise-like current injections with irregular-firing patterns if they have IKL and with more regular firing patterns if they do not. However, in vitro firing patterns are not as regular as those in vivo. Here we considered whether highly-regular spiking requires IH currents and whether this dependence emerges later in development after channel expression matures. We recorded from rat VGN somata of either sex aged postnatal day (P)9–P21. Counter to expectation, in vitro firing patterns were less diverse, more transient-spiking, and more irregular at older ages than at younger ages. Resting potentials hyperpolarized and resting conductance increased, consistent with developmental upregulation of IKL. Activation of IH (by increasing intracellular cAMP) increased spike rates but not spike-timing regularity. In a model, we found that activating IH counter-intuitively suppressed regularity by recruiting IKL. Developmental upregulation in IKL appears to overwhelm IH. These results counter previous hypotheses about how IH shapes vestibular afferent responses.
SIGNIFICANCE STATEMENT Vestibular sensory information is conveyed on parallel neuronal pathways with irregularly-firing neurons encoding information using a temporal code and regularly-firing neurons using a rate code. This is a striking example of spike-timing statistics influencing information coding. Previous studies from immature vestibular ganglion neurons (VGNs) identified hyperpolarization-activated mixed cationic currents (IH) as driving highly-regular spiking and proposed that this influence grows with the current during maturation. We found that IH becomes less influential, likely because maturing VGNs also acquire low-voltage-gated potassium currents (IKL), whose inhibitory influence opposes IH. Because efferent activity can partly close IKL, VGN firing patterns may become more receptive to extrinsic control. Spike-timing regularity likely relies on dynamic ion channel properties and complementary specializations in synaptic connectivity.
- cAMP
- IH
- IKL
- neuronal excitability
- spike-timing regularity
- vestibular ganglion
Introduction
The vestibular system sends information from the sensory periphery to the brainstem along parallel neuronal pathways whose signature distinction is a striking difference in spike-timing statistics. Spikes occur at regular intervals in one path and at irregular (noisy) intervals in the other (Baird et al., 1988; for review, see Goldberg, 2000). Spike-timing regularity correlates with other important features of the neuronal response, such as gain, phase, and sensitivities to different frequencies of head motion (for review, see Goldberg, 2000). It also appears to determine whether these neurons use a rate or temporal code to represent sensory information (Sadeghi et al., 2007). Given its central importance to vestibular function, the aim of our study is to understand the biophysical mechanisms that drive spike-timing regularity.
Pioneering studies considered many mechanisms to explain spike-timing regularity. These included the type and numbers of sensory hair cells contacted by individual neurons, differences in synaptic and/or dendritic morphology, and regional variations in ion channel composition (Smith and Goldberg, 1986; Baird et al., 1988; Fernández et al., 1988, 1990). Ultimately, these studies concluded that variations in ion channel composition are critical for driving differences in spike-timing regularity (Highstein and Politoff, 1978; Smith and Goldberg, 1986; Schessel et al., 1991), but the exact ion channel substrate remains under debate.
Electrophysiological and immunohistochemical data in rats and mice have shown expression for many groups of ion channels, both in the terminals and at the somata of vestibular afferents (Desmadryl et al., 1997; Chabbert et al., 2001; Risner and Holt, 2006; Iwasaki et al., 2008; Kalluri et al., 2010; Lysakowski et al., 2011; Almanza et al., 2012; Horwitz et al., 2014; Yoshimoto et al., 2015). For example, potassium channel expression varies by epithelial zone and could drive zonal variations in spike-timing regularity (Songer and Eatock, 2013; Meredith and Rennie, 2015). Indicative of heterogeneity in ion channel composition, vestibular ganglion neuron (VGN) somata respond with diverse firing patterns when injected with step currents. Somata with large low voltage-gated potassium currents (IKL) respond only at the onset of the current step (transient/phasic), whereas those with little IKL respond with sustained spike trains (sustained/tonic; Fig. 1). When driven with currents that mimic the random timing of synaptic input, transient-spiking VGNs respond with irregularly-timed spike trains, whereas sustained-spiking VGNs respond with more regular patterns (Kalluri et al., 2010). Although these results suggest that IKL can control spike-timing regularity, in vitro firing patterns do not have the degree of regularity seen in vivo.
Recent studies suggest that highly-regular spiking relies on the depolarizing currents provided by the hyperpolarization-activated cyclic nucleotide (HCN)-sensitive currents (i.e., IH currents conducted by HCN channels; Horwitz et al., 2014; Yoshimoto et al., 2015). This hypothesis is appealing because IH produces highly regular auto-rhythmic firing in other systems, famously in sinoatrial node myocytes allowing for pacemaking in the heart (Brown et al., 1979; DiFrancesco, 1981, 2010). Because IH expression in VGNs grows during development (Yoshimoto et al., 2015) and the current is sensitive to cyclic-nucleotide concentrations (for review, see DiFrancesco, 2010; for VGNs, see Almanza et al., 2012), its influence on spiking could be masked by experimental conditions.
To understand the ion channel substrate necessary for driving variations in spike-timing regularity, we performed in vitro whole-cell patch-clamp on isolated VGNs at ages when HCN channel expression has matured and in experimental conditions where the activation of the IH current is enhanced by increasing intracellular concentrations of cAMP. We report that enhancing IH increased spike rate, but failed to produce highly regular firing. Using a single compartment model of VGNs, we show that opposing interactions between IH and IKL currents can explain the lack of highly regular firing.
Materials and Methods
Preparations
Data were recorded from the superior portion of the vestibular ganglion in Long–Evans rats of either sex aged postnatal day (P)9–21 (P0, birth day). All animals were handled and housed in accordance with National Institutes of Health Guide for the Care and Use of Laboratory Animals. All animal procedures were approved by the University of Southern California Institutional Animal Care and Use Committee. Chemicals were obtained from Sigma-Aldrich unless otherwise specified. The temporal bones from the animals were dissected in chilled and oxygenated L-15 supplemented with 10 mm HEPES (Leibowitz medium). The superior part of the vestibular ganglia was detached from the distal and central nerve branches freeing it from the otic capsule. Bone fragments, debris, and any remaining connective tissue were removed from the surface of the ganglia. Ganglia from 2 to 4 litter-matched animals of either sex were pooled together. Ganglionic tissue was then incubated at 37°C in a solution of L-15 with 0.05% collagenase and 0.25% trypsin for 20–45 min, depending on age of the animal. The ganglia were then washed in fresh L-15 solution, dissociated by triturating through a series of polished Pasteur pipettes and allowed to settle onto poly-d-lysine-coated glass bottom culture dishes (MatTek). Culture dishes contained bicarbonate-buffered culture medium (minimal essential medium, Invitrogen), supplemented with 10 mm HEPES, 5% FBS, and 1% penicillin-streptomycin (Invitrogen). Cells were incubated for 16–24 h in 5% CO2/95% air at 37°C. Short-term incubation tends to remove supporting and satellite cells, debris from enzyme treatment, and promotes successful recordings as the animals entered the second postnatal week and beyond.
Electrophysiology
Cells were viewed at 400× using an inverted microscope (Zeiss, Axiovert 135 TV) fitted with Nomarski optics. A MultiClamp 700B amplifier, Digidata 1440 board, and pClamp 10.7 software (MDS; RRID:SCR_011323) were used to deliver, record, and amplify all signals. Custom stimuli were generated in MATLAB (MathWorks; RRID:SCR_001622) and delivered via the pClamp software. Recording pipettes were fabricated using filamented borosilicate glass. Pipettes were fire polished to yield an access resistance between 4 and 8 MΩ. Recording pipettes were coated with a silicone elastomer (Sylgard 184; Dow Corning) to reduce pipette capacitance.
The properties of ion channels were studied using whole-cell ruptured-patch or perforated-patch methods. The contents of the standard internal solution contained the following (in mm): 135 KCl, 3.5 MgCl2, 3 Na2ATP, 5 HEPES, 5 EGTA, 0.1 CaCl2, 0.1 Li-GTP, and titrated with 1 m KOH to a pH of 7.35 and an osmolarity of ∼300 mmol/kg. Both cAMP and dibutyryl (db)-cAMP were aliquoted and stored as a 10 mm stock at −20°C. Working concentrations were achieved by dilution into the intracellular solution on day of recording. Endogenous conditions were studied using the perforated patch-clamp method where the intracellular milieu is relatively unchanged. The contents of the perforated-patch internal solution contained the following (in mm): 75 K2SO4, 25 KCl, 5 MgCl2, 5 HEPES, 5 EGTA, 0.1 CaCl2, and titrated with 1 m KOH to a pH of 7.4. Amphotericin B (240/ml; Sigma-Aldrich) was dissolved in DMSO and added to the perforated-patch solution on day of recording. This allowed passage of small monovalent ions while preventing larger molecules from dialyzing.
The series resistance ranged between 8 and 35 MΩ (higher values for perforated patch) and was compensated by 70 and 50%, respectively; giving final resistances between 2.5 and 17 MΩ. Uncompensated resistance was corrected off-line. Recordings were made at room temperature (25–27°C), unless otherwise stated, and in an external bath continuously perfused with fresh oxygenated L-15 media. Standard internal solution and perforated patch internal solution had junction potentials of +4.3 and +5.0 mV, respectively. In some experiments the bath was heated to between 35°C and 37°C with a heated platform and temperature controller (TC-344B; Warner Instruments). To maintain bath temperature with stability, room humidity was maintained using a bench-top humidifier. Junction potentials were computed with JPCalc (Barry, 1994) as implemented in pClamp 10.7 and not corrected for. Only recordings in which the cell had formed a gigaohm seal and maintained stable membrane potential were used. The stability of neurons was monitored by measuring resting potential, input resistance, firing patterns, and in some neurons the activation properties of IH, at regular intervals throughout the recording period.
Pharmacology
IH antagonists.
A stock solution of ZD7288 was stored at −20°C in 10 mm aliquots. An 80 μm working concentration was achieved by dilution in either L-15 for extracellular administration or in standard internal solution for intracellular administration. A 2 mm stock solution of CsCl in L-15 was prepared fresh on the day of recording for extracellular administration. Drugs were applied via a pressurized super-perfusion system (Warner Instruments). Measurements of IH were made at least 2–5 min after exposure to the drug.
Analysis
General.
All data were analyzed with pClamp 10 software (Clampfit; MDS Analytical Technologies). Input resistance (Rin) was calculated from voltage changes in response to a hyperpolarizing 10 or 20 pA step of current. The membrane time constant (τm) was acquired by fitting the resulting voltage response with a single exponential. Membrane capacitance (Cm) was computed as the ratio of membrane time constant and input resistance (τm/Rin).
Statistical analysis was done with Origin Pro (OriginLab; RRID:SCR_014212) and/or JMP Pro 13 (SAS Institute; RRID:SCR_014242). Statistical significance was estimated with Student's t test when variances were equal or with Welch's t test when variances were not equal. We used an α level of 0.05 for all statistical tests. One-way and two-way ANOVAs were applied followed by a post hoc Tukey's HSD analysis, as required, to examine the influence of cAMP concentration/condition on IH half activation, firing rate and coefficient of variation (CV). Statistics are reported as means ± SEM.
IH activation parameters.
We studied the voltage-dependent activation of IH using a voltage-clamp tail-current protocol (Fig. 2). The HCN channel that carries IH opens very slowly and the voltage-dependent activation of IH is sensitive to the length of hyperpolarization. We designed a protocol that allowed ample time for the channel to activate and provided an accurate measurement of the voltage-dependent activation. The challenge is that neurons can die when held at hyperpolarized voltages for extended periods of time. The protocol begins with a preconditioning voltage step, then a −100 mV “tail-step” for 300 ms and finally returns to −60 mV for 100 ms. The precondition voltage step is incremented by 5 mV to range between −135 and −40 mV. A 15 s delay was implemented between each step to allow enough time for HCN channels to close completely before starting the next voltage step. Several preconditioning lengths were tested (1.25, 1.7, and 2.5 s). Although the 2.5 s protocol provided the most depolarized measures of voltage-dependent activation and had the most complete IH activation, we found that 1.7 s provided comparable results without killing the neuron. The voltage-dependent activation of IH is characterized by plotting the currents flowing during the tail step (Itail) against the precondition voltage (Vpre). These data were well fit with a Boltzmann function (Eq. 1):
 where Imax is the maximum current measured during the tail, Vpre is the preconditioning/test voltage, V1/2 is the half-activation potential, k is the slope factor, and C is a constant. Measurements of Itail were taken 100 ms into the tail step to avoid contamination from other low-voltage-gated currents, particularly during the most positive voltage steps (−60 to −40 mV) and when IH was activated with cAMP/db-cAMP.
where Imax is the maximum current measured during the tail, Vpre is the preconditioning/test voltage, V1/2 is the half-activation potential, k is the slope factor, and C is a constant. Measurements of Itail were taken 100 ms into the tail step to avoid contamination from other low-voltage-gated currents, particularly during the most positive voltage steps (−60 to −40 mV) and when IH was activated with cAMP/db-cAMP.
The voltage-dependent time course for IH was estimated by fitting the current evoked at the −135 mV preconditioning step with a single exponential. All fits were obtained after a 60 ms delay from the start of the voltage step. Only fits with a correlation coefficient ≥0.985 were retained. Mean τ values from the single exponential fit at −135 mV were ∼327.9 ± 30.7 ms (n = 20), which are consistent with those reported by Almanza et al. (2012) in VGNs. We also tried fitting with the sum of two exponentials, which improved fits for nearly 80% of cells at −135 mV (Almanza et al., 2012; Liu et al., 2014), but this type of fit failed more often at depolarized potentials. To yield consistent fits as a function of voltage, we relied on the single exponential fits, which were uniformly good for all cells and voltages.
Pseudo-synaptic stimuli
To mimic natural synaptic input, we generated trains of simulated EPSCs (pseudo-EPSCs) with pseudorandom timing, as previously described by Kalluri et al. (2010). A stimulus set of 20 pseudo-EPSC sweeps was created and delivered to each VGN in current-clamp mode to drive spiking. Spikes were detected with the built-in spike thresholding algorithms of Clampfit 10.7
For each neuron the CV gives a measure of the spike-timing regularity. CV is computed as the SD of the interspike interval (ISI) divided by the mean ISI. Only CV values for neurons in which the SEM spike rate was <10% of the mean ISI were included. CV is inherently related to spike rate (Goldberg et al., 1990a). The normalized statistic CV* is often used to account for this relationship (Goldberg et al., 1990a). CV* is the CV at a standard spike rate and allows for a measure of regularity independent of spike rate. We did not use CV*, choosing instead to only compare CVs of neurons that were driven at the same mean spike rate.
Variations in spike rate were achieved by increasing or decreasing the mean amplitude of the pseudo-EPSCs. Although effective in changing spike rate, this is not a completely natural method of stimulation. A more natural stimulus would modulate both EPSC amplitude and EPSC rate. This stimulation method was chosen to maintain timing patterns (by maintaining a standard EPSC rate) to isolate the influence of ion channels from synaptic input.
Vestibular model
A single compartment conductance-based model was used to test the influence of IH and IKL on firing patterns. The model was implemented in MATLAB as a differential equation in which the net current across the neuronal membrane was taken as the sum of currents flowing through individual circuit elements connected in parallel (see Eq. 8).
 The currents were driven by the following five ionic conductances: sodium (gNa), low-voltage-gated potassium (gKL), high-voltage-gated potassium (gKH), hyperpolarization-activated mixed cationic (gH), and leak (gleak). This model is an extension of the single-compartment VGN model previously described by Hight and Kalluri (2016). Here we varied the conductance density, voltage-activation range, and kinetics of the IH current to mimic the influence of cAMP, as described by the in vitro characterizations in this study. In addition, we expanded on the previous description of gKL to include both a Kv1-like and Kv7-like components.
The currents were driven by the following five ionic conductances: sodium (gNa), low-voltage-gated potassium (gKL), high-voltage-gated potassium (gKH), hyperpolarization-activated mixed cationic (gH), and leak (gleak). This model is an extension of the single-compartment VGN model previously described by Hight and Kalluri (2016). Here we varied the conductance density, voltage-activation range, and kinetics of the IH current to mimic the influence of cAMP, as described by the in vitro characterizations in this study. In addition, we expanded on the previous description of gKL to include both a Kv1-like and Kv7-like components.
Hyperpolarization-activated cyclic nucleotide-gated mixed-cationic current.
Equations 3 through 5 governed the voltage-dependent kinetics of IH:


 Cell surface area (S) was fixed to yield a net capacitance of 10 pF. Conductance density (ḡh), reversal potential (EH), half-activation voltage V1/2, slope factor k, and voltage-dependent activation kinetics (τr) were based on experimentally derived values from this study. Half-activation voltage and activation kinetics were defined for two conditions; 100 μm cAMP and 500 μm dB-cAMP, which respectively represent minimum and maximum activation conditions. The parameter values for V1/2, k, a and e are summarized in Table 1. Conductance densities and other model parameters are summarized in Tables 2 and 3.
Cell surface area (S) was fixed to yield a net capacitance of 10 pF. Conductance density (ḡh), reversal potential (EH), half-activation voltage V1/2, slope factor k, and voltage-dependent activation kinetics (τr) were based on experimentally derived values from this study. Half-activation voltage and activation kinetics were defined for two conditions; 100 μm cAMP and 500 μm dB-cAMP, which respectively represent minimum and maximum activation conditions. The parameter values for V1/2, k, a and e are summarized in Table 1. Conductance densities and other model parameters are summarized in Tables 2 and 3.
Model Parameters for r∞ and τr selected from our experimental data
Model parameters
Additional model parameters
Low-voltage-gated K+ currents.
Equations 6 through 8 governed the voltage-dependent activation kinetics of IKL;
 IKL was driven in part by Kv1-like and Kv7-like conductances and γkv1 and γkv7 defined the fraction of total conductance contributed by each of the two components. The variables w4 and z represent activation and inactivation kinetics. The inactivation kinetics were adjusted to reflect the inactivation present in Kv1 channels and the lack thereof in Kv7 channels. The Kv1 component was modeled to activate nearly 10 times faster than the Kv7 component (compare Eqs. 7, 8):
IKL was driven in part by Kv1-like and Kv7-like conductances and γkv1 and γkv7 defined the fraction of total conductance contributed by each of the two components. The variables w4 and z represent activation and inactivation kinetics. The inactivation kinetics were adjusted to reflect the inactivation present in Kv1 channels and the lack thereof in Kv7 channels. The Kv1 component was modeled to activate nearly 10 times faster than the Kv7 component (compare Eqs. 7, 8):


Model implementation.
The model was implemented in MATLAB 2016b running on a Windows 10 operating system. The differential equation (Eq. 2) for the membrane voltage, V(t), was numerically solved using a backward difference method. The associated code is accessible via the ModelDB repository at http://senselab.med.yale.edu/ModelDB/showModel.cshtml?model=244202.
Results
Whole-cell patch-clamp recordings were made by either ruptured-patch or perforated-patch methods in isolated vestibular ganglion neurons (VGN) from rats ranging in age between postnatal day (P)9 through P21 (n = 146). First, we describe the basic firing patterns of VGNs during the second and third postnatal weeks. Second, we define the sensitivity of the IH current in VGNs to modulation by cyclic adenosine monophosphate (cAMP), a well known modulator of HCN channels. Third, we describe the influence of IH on VGN firing patterns and spike-timing regularity. Finally, we explore the interaction between IH and IKL using a single compartment conductance-based model for VGN.
Step-evoked firing patterns
We classified neurons into two broad groups based on the firing patterns evoked by depolarizing current injections: transient-firing (83 of 146 neurons; Fig. 1A) and sustained-firing (63 of 146 neurons; Fig. 1B–D). These classifications are similar to those used by Kalluri et al. (2010) and Iwasaki et al. (2008). Transient-firing neurons displayed strong spike-train accommodation, firing a single spike at the onset of the current step. Beyond spike threshold, the firing pattern of most transient-spiking neurons was invariant to intensity. In a few rare cases, transient-firing neurons fired a second spike when injected with high amplitude currents (4/83 neurons; data not shown). Sustained-firing neurons displayed varying degrees of accommodation, ranging from cells that fired long trains of spikes (sustained-A firing patterns had spikes throughout the depolarizing step; Fig. 1D) to cells with intermediate degrees of accommodation (sustained-B/sustained-C firing patterns had shorter trains of spikes followed by voltage oscillations; Fig. 1C, B, respectively). Consistent with Kalluri et al. (2010), the degree of accommodation depended on intensity for the sustained-firing patterns. Figure 1E shows a sustained-firing pattern at a recording temperature between 35°C and 37°C.

Vestibular ganglion neurons produce different firing patterns in response to depolarizing current steps. A, Transient-firing neurons are characterized by a single action potential at the onset of suprathreshold depolarizing steps. B–D, Sustained-firing patterns are more variable. B, Sustained-C neurons have one or two spikes followed by prominent voltage oscillations. C, Sustained-B neurons show adaptation, producing several spikes at current onset and then cease firing (accommodate). D, Sustained-A neurons produce long spike trains throughout the current step. E, Sustained-A neuron at 35–37°C in 500 μm db-cAMP, which produces more spikes than in sustained-A neuron shown in D. The amplitude of the depolarizing current step that evokes spikes (current threshold) is shown in the label at the right of each trace and the dashed line represents −60 mV. Indicative of the presence of hyperpolarization-activated mixed cationic current (IH), most VGNs produced a voltage sag (arrow labeled Vsag) in response to large hyperpolarizing currents (−200 pA shown). Scale bars (top right trace) apply to all traces. For this figure, recordings were made using whole-cell ruptured-patch with 100 μm cAMP, except where the recording pipette contained 500 μm db-cAMP.
When injected with hyperpolarizing currents, most neurons (140/146) responded with a deep voltage hyperpolarization followed by a slow depolarization that eventually reached steady state (Fig. 1A–D). Such hyperpolarization-induced voltage “sags” indicate the presence of the HCN channel-mediated current, IH (Fig. 1C; Rennie and Streeter, 2006; Angelo and Margrie, 2011; Almanza et al., 2012). On average, voltage sags increased in amplitude with larger hyperpolarizing current injections and were first visible when the membrane potential fell below −80 mV.
Our goal was to understand how IH current influences spike-timing regularity. One way to do this is to compare responses before and after selectively blocking the current with channel-specific antagonists such as ZD7288 and CsCl. This approach was not successful here because both antagonists may affect more than just HCN channels. For example, CsCl is known to block other kinds of potassium channels including inward rectifying potassium channels (Bond et al., 1994; Takumi et al., 1995). Previous studies have also shown that ZD7288 can partially block sodium, T-type calcium, and/or potassium channels (Felix et al., 2003; Sánchez-Alonso et al., 2008; Wu et al., 2012). We suspected similar nonspecific effects in VGNs because action potential amplitudes decreased, and net conductance increased when either ZD7288 or CsCl blocked IH (5 of 8 cells; data not shown). This is a counter-intuitive result since blocking the channel should have decreased net conductance. Although not discussed, a similar effect in VGNs is evident in the recordings presented by Yoshimoto et al., 2015. We concluded that we could not rely on these channel antagonists to isolate the role of IH on firing patterns. As shown next, we chose to amplify IH by controlling the intracellular concentration of cAMP, which shifts the current's voltage activation range (Almanza et al., 2012).
Enhancing the activation of IH by controlling cAMP concentration
In voltage-clamp, hyperpolarization produced inward currents with a near instantaneous component (Iinst) and a slowly activating component that eventually reached steady state (Iss; Fig. 2A). Known IH antagonists, CsCl (Fig. 2A2) and ZD7288 (data not shown), blocked the slow current, suggesting that it was HCN mediated. The current's reversal potential (−32.7 ± 2.8 mV, n = 8) and voltage activation range and kinetics (discussed further below; Figs. 2, 3) are consistent with reported values for IH (Biel et al., 2009; Almanza et al., 2012; Yoshimoto et al., 2015). We measured the activation range of IH under several experimental conditions. First, we varied the concentration of simple cAMP in the intracellular pipette solution (at 10, 100, 200, and 500 μm). Second, we used perforated-patch methods (where the intracellular milieu is left intact) to estimate the endogenous concentration of cAMP. Third, we used 100 and 500 μm db-cAMP, a non-hydrolysable version of cAMP that resists degradation by phosphodiesterases. Fourth, we tested if the activation of IH is further enhanced by increasing recording temperature, as previously reported in the substantia nigra (Gambardella et al., 2012).

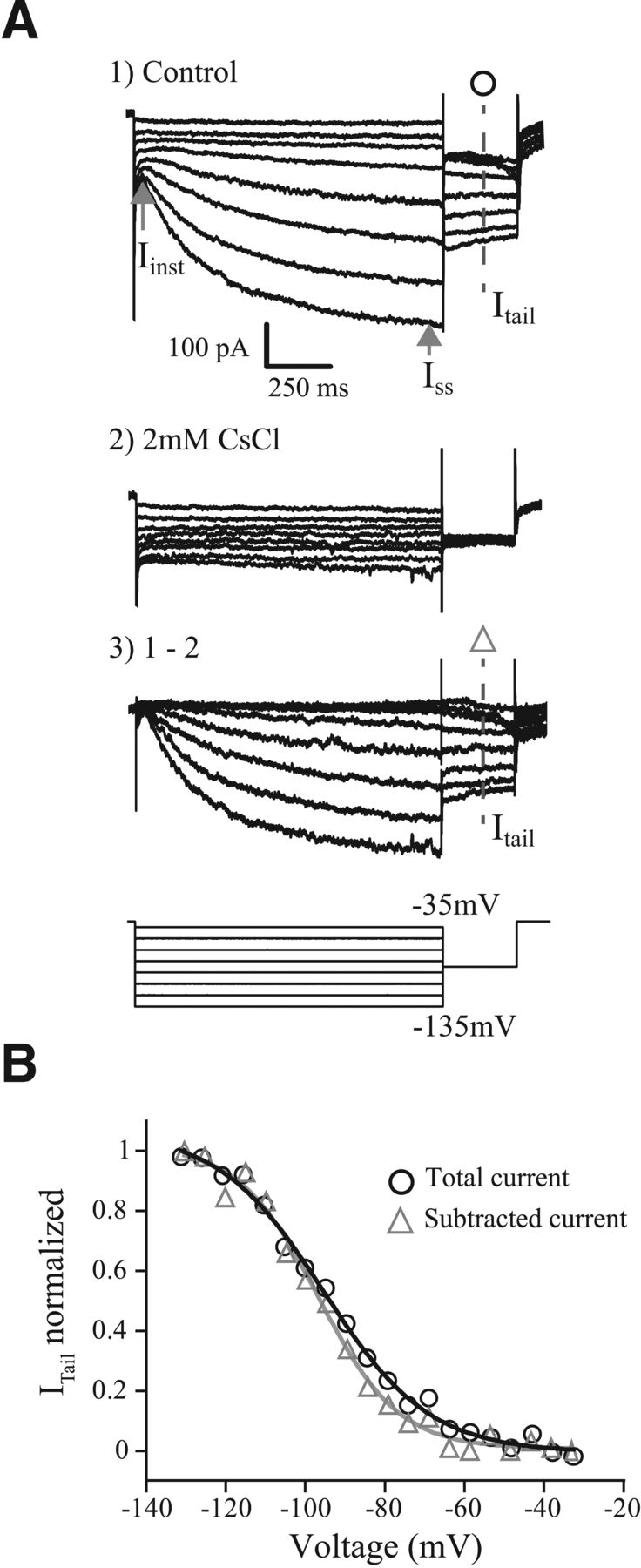
An electrophysiological and pharmacological method for characterizing the activation properties of IH. Current excited by a family of long duration (1.7 s) conditioning voltage steps ranging from −135 to −35 mV in 5 mV increments followed by a 300 ms tail step to −100 mV. For clarity, not every voltage-step response is shown. Current under control conditions (A1) and with CsCl (A2) and the Cs+-sensitive current obtained by subtraction 1–2 (A3). Gray arrows, The amplitude of the IH current was determined by taking the difference between the Iinst and the ISS. B, The voltage-dependent activation of IH is determined by plotting the magnitude of the tail current (Itail) as a function of the conditioning voltage. Curves were fit by a Boltzmann function and normalized by the maximum current. The fits generated from the tail of the control condition (○) and from the current isolated from the Cs+ induced block (Δ) are similar. All voltage-dependent characterization of IH in subsequent figures is derived from the control condition.

Effects of cAMP concentration and temperature on the activation of IH currents. A, IH currents activated by a family of voltage steps from −135 to −35 mV under several cAMP concentrations and temperature combinations. As in Figure 2, tail currents were used to construct current–voltage relationships. B, Normalized current–voltage relationships (means ± 95% confidence intervals) for IH are compared for the 10 μm cAMP and 500 μm db-cAMP conditions. The difference between the two conditions illustrates the potential range of IH activation. C, Box plots for each cAMP condition. V1/2 depolarizes dependent on cAMP concentration, db-cAMP conditions produce the most depolarized V1/2 values (shaded box plots). Increasing the concentration of db-cAMP from 100 to 500 μm did not further depolarize V1/2 and neither did increasing the recording temperature to 35°C. The endogenous concentration was obtained in perforated patch configuration. Mean slope value (k) for all the conditions was 8.3 ± 0.2 mV and not affected by cAMP concentration. D, A single exponential was used to fit to the currents produced by voltage steps from −135 to −65 mV. The time constant of the fits, τsingle, is plotted as function of voltage for 100 μm cAMP (blue; n = 17), 500 μm db-cAMP (red; n = 10), perforated patch (black; n = 4), and 500 μm db-cAMP at 35°C (green; n = 8). Solid lines show fits were produced using Equation 5 to show the relationship of τsingle values and voltage. Parameters for the fits at 100 μm cAMP, 500 μm cAMP, and 500 μm db-cAMP at 35°C are summarized in Table 1. E, For all conditions except those with db-cAMP, mean V1/2 hyperpolarized as a function of time. V1/2 ± SEM is plotted as a function of recording time for 10 μm cAMP (diamonds), 100 μm cAMP (cyan circles), 100 μm db-cAMP (gray downward triangle), 500 μm cAMP (squares), and 500 μm db-cAMP (red open squares).
Estimating the voltage-dependent activation of IH
To estimate the voltage-dependent activation properties of IH, we used a voltage-clamp tail protocol, which consists of 1.7-s-long preconditioning steps from −135 to −35 mV in 5 mV increments followed by a tail step to −100 mV (Fig. 2A, bottom). The long preconditioning step allowed IH to approach steady-state activation but was not so long as to kill cells because of prolonged hyperpolarization (see Materials and Methods). The tail step to −100 mV was within the typical activation range for HCN channels but hyperpolarized away from the activation range of most other channels known be expressed in VGNs (e.g., low-voltage-gated potassium channels). The currents measured during the tail step reflect the channels activated during the preconditioning steps but with a fixed driving force. The maximum current density (Imax/capacitance) was larger for sustained-firing neurons than for transient-firing neurons (11.8 ± 1.2 pA/pF vs 6.9 ± 0.6 pA/pF; t(25.8) = −3.6, p = 0.0013, Welch's unpaired t test), consistent with results reported by Yoshimoto et al. (2015). The normalized tail current (Itail/Itailmax) is plotted as a function of the preconditioning step to show the voltage-dependent activation of IH (Fig. 2B). The normalized activation curves generated from these whole-cell currents were similar to those extracted by pharmacologically isolating IH (Fig. 2B). These results led us to conclude that the tail current measurements were adequate for characterizing IH. All subsequent IH activation parameters are derived from the unblocked whole-cell currents.
Modulating the voltage-dependent activation range of IH
Examples of whole-cell currents recorded in response to the tail protocol from six cells, each at different intracellular cAMP concentrations and two obtained in 35°C are shown in Figure 3A. The activation curves shifted to more positive potentials as cAMP concentration increased; nearly 15 mV when two extreme concentrations were compared (10 and 500 μm db-cAMP; Fig. 3B). A similar shift has been reported for auditory afferents in response to cAMP analogues (Yi et al., 2010). We found a significant dependence of V1/2 on cAMP condition (F(6,44) = 11.67, p = 0.0001 based on one-way ANOVA; Fig. 3C). Based on similarity in V1/2, we estimate that the endogenous concentration of cAMP (the perforated-patch condition) is between 10 and 100 μm. This estimate is consistent with reports by Almanza et al. (2012). At 100 μm cAMP, the mean V1/2 and slope factor (k) are −94.32 ± 3.06 mV and 7.97 ± 1.31 mV (n = 14), respectively. This is also consistent with reported values for IH at 100 μm cAMP (−93 ± 2 and 9 ± 0.9 mV; Almanza et al., 2012). Interestingly, mean V1/2 values were not significantly different between the 100 and 500 μm db-cAMP conditions, suggesting that enzymatic degradation of cAMP is a critical parameter affecting cAMP concentration and IH activation.
Temperature did not further depolarize the activation range of IH. In the substantia nigra increasing recording temperature depolarizes the activation range of IH by as much as 15–20 mV (Gambardella et al., 2012). Here we found that recording in 500 μm db-cAMP with an elevated temperature (35°C) did not further depolarize the activation range of IH (Fig. 3C, compare gray box plots). These results suggest that the db-cAMP is at least as effective at shifting the activation range of IH as temperature. We did not test the influence of temperature alone on the activation range of IH, although such a temperature-dependent shift would offer a physiologically plausible mechanism for enhancing IH.
Modulating the activation time course
Next, we show that the time course of IH activation speeds up with increased concentrations of cAMP. We applied single exponential fits to the currents evoked by the family of voltage steps from −135 to −55 mV. A single exponential reliably fit our data between −135 and −55 mV. Double exponentials better fit the IH current at −135 mV (Almanza et al., 2012; Liu et al., 2014) but failed for the smaller hyperpolarizations as the size of IH current diminished. We relied on single exponential fits because they were robust in all cells and for wide range of voltage steps. In Figure 3D, the exponent τ is plotted as a function of voltage steps for recordings made under the four cAMP conditions (perforated-patch, black triangle; 100 μm cAMP, blue circle; 500 μm-dB cAMP, red square; 500 μm-dB cAMP @ 35°C, green triangles). The voltage-dependent trends for τ activation were fit with Equation 5 (see Materials and Methods) for each cAMP condition (Fig. 3D, solid lines). These same fits were used in the model described later. In each condition, the current activates fastest at extreme hyperpolarizations and slows as the voltage step approaches V1/2. As the voltage steps become more positive the activation kinetics speed up. Like the cAMP-dependent shift in voltage activation, the activation kinetics shift such that at any particular voltage (e.g., −90 mV) the 500 μm db-cAMP was the fastest. Similar cAMP driven acceleration in activation kinetics have been reported in various types of neurons including in chick nucleus laminaris, substantia nigra pars compacta and spiral ganglion (Yamada et al., 2005; Yi et al., 2010; Gambardella et al., 2012). Although increasing temperature did not further depolarize half activation voltage beyond that produced by 500 μm dB-cAMP (Fig. 3C), it did speed up activation kinetics (Fig. 3D, green symbols).
Time-dependent stability of IH modulation by cAMP
Although we found that the activation range of IH could be shifted by increasing cAMP concentration, the shifts were most robust and prolonged in the dB-cAMP conditions. Since we were characterizing the activation properties of IH and firing patterns in the same groups of cells, it was important for us to maintain stability for ∼30–40 min. To determine the time-stability of cAMP driven shift, we measured IH activation curves at regular time intervals throughout the recording session. Figure 3E shows mean V1/2 as a function of recording time and by cAMP condition. For all but the 500 μm dB-cAMP condition, the mean V1/2 was most depolarized during early recordings (0–10 min) and became progressively more hyperpolarized as neurons were held for longer periods (30–40 min). In Figure 3C, activation parameters of IH were taken from the earliest recording time possible. In db-cAMP, V1/2 values remained time stable, showing no significant difference between early (0–10 min) and late recordings (30–40 min; t(3.3) = −0.13, p = 0.907, paired t test). This is presumably because db-cAMP is a non-hydrolysable version of cAMP that can maintain elevated intracellular cAMP levels for hours (Campos-Toimil et al., 2008). We did not determine whether db-cAMP remains stable beyond 40 min. The ability to stably shift the activation range of IH was ultimately important for characterizing the influence of IH on firing patterns (presented in the next section). Most of our quantitative comparisons are between the 100 μm cAMP condition (which we estimate to be most like the endogenous condition) and the 500 μm db-cAMP condition (where we are confident that IH activation was the most robust and stable).
Effects of cAMP on firing patterns
Step-evoked firing patterns with increased cAMP
To understand how cAMP driven enhancement of IH influenced firing patterns, we compared spikes evoked by ∼100–120 pA step currents in 100 μm cAMP (n = 12) and 500 μm db-cAMP (n = 10). We also recorded at 35°C with 500 μm cAMP (n = 8) to test whether increasing temperature together with elevated cAMP concentrations had an additional influence on spike timing. Figure 4, A1 and A2, shows spike traces from example sustained-spiking neurons in the three recording conditions. For each neuron, we quantified the first spike latency (tp) and the slope of the afterhyperpolarization trajectory (AHP slope; Fig. 4B1,B2, filled symbols are for the examples in A). We applied two-way ANOVAs to test the influence of the three cAMP conditions and firing-pattern category on each of the two spike features. Both, first spike latency and AHP slope were significantly dependent on cAMP condition (F(2,29) = 4.47, p = 0.02; F(2,30) = 5.10, p = 0.01, respectively) and AHP slope also depended on firing-pattern (F(1,30) = 7.58, p = 0.01). Post hoc analysis by Tukey HSD indicated that first spike latency tended to be shorter and AHP slopes were steeper in the two 500 μm db-cAMP conditions than in the 100 μm cAMP condition (p = 0.030 and p = 0.086, respectively, for first spike latency; p = 0.026 and p = 0.031, respectively, for AHP slope). Neither feature was significantly different between the 500 μm db-cAMP and 500 μm db-cAMP at 35°C conditions (p = 0.96 for first spike latency; p = 0.99 for AHP slope). These results suggest that more IH current was available in elevated cAMP to speed up afterhyperpolarization trajectories and spike timing, but that raising temperature had no additional influence on these two spike features (Fig. 4B,D).

The effects of a db-cAMP and temperature on step-evoked firing patterns. A1, Example responses to a depolarizing current step for sustained-firing VGNs; one recording in 100 μm cAMP (black) and the second in 500 μm db-cAMP at 25°C (red) conditions. A2, Example responses to a depolarizing current step for sustained-firing VGNs; one recording in 500 μm db-cAMP at 25°C (red) condition and the second in 500 μm db-cAMP at 35°C (cyan) condition. VGN firing patterns in A were obtained in response to similar current step (∼100–120 pA). First spike latency (tp) and the slope of the afterhyperpolarization (AHP) trajectory were obtained as shown in A1. AHP slope was obtained by taking the slope of a line fit to the region illustrated in A. Dashed gray line shows −60 mV. B1, B2, Data for time to peak (ms) and AHP slope (mV/ms) for transient (□) and sustained (Δ) is shown for all three conditions 100 μm cAMP (black n = 11) and 500 μm db-cAMP at both 25°C (red; n = 10) and 35°C (cyan; n = 8). Solid symbols are data for the example VGN shown in A.
Noise (pseudo-EPSC) driven firing patterns with increased cAMP
Next, we examined whether the enhanced activation of IH yielded regular spiking. Although spontaneous spiking in vivo is driven by quantal release of neurotransmitter from hair-cells, dissociated VGNs require current injection to induce spiking. To mimic the random timing of synaptic input, we injected “pseudo-synaptic” currents into VGN somata (n = 47; for additional detail see Materials and Methods; Kalluri et al., 2010). The size of the unitary EPSC was varied to drive neurons at different rates (Fig. 5A,B). Moreover, the average EPSC size needed to drive neurons to similar rates was larger in transient-firing neurons than sustained-firing neurons (Fig. 5C). Based on the steeper AHPs in step-evoked firing patterns, we hypothesized that firing rates should be higher in 500 μm db-cAMP where the enhanced activation of IH contributed to the depolarization phase of spike generation. Recordings with 500 μm db-cAMP (n = 11) produced a mean firing rate of 30.4 ± 2.9 spikes/s, significantly higher than the 18.5 ± 1.0 spikes/s in the 10/100 μm condition (p < 0.001, Tukey HSD). However, when we looked at each condition by firing type we saw that this increase in mean firing rate was mainly driven by sustained-B-firing neurons (n = 5), which produced firing rates as high as 70 spikes/s in db-cAMP (Fig. 5C, black triangles). Again, increasing the temperature from 25°C to 35°C did not further increase mean firing rates beyond what was achieved with db-cAMP alone (32.6 ± 3.2 spikes/s, n = 8 vs 30.4 ± 2.9 spikes/s, n = 11).

Enhancing IH activation by increasing cAMP concentration does not increase spike-timing regularity. A, Spike trains in response to pseudo-synaptic current injection. Spike rate increased as the amplitude of unitary pseudo EPSC increased. B, ISI histograms constructed from spike trains in response to frozen variations of pseudo-synaptic currents. Mean spike interval (ISI) decreased as unitary EPSC amplitude increased compare black and gray histograms. C, Firing rate (spikes/s) as a function of mean EPSC amplitude (pA) in different cAMP concentrations. Mean firing rate was impacted by cAMP condition and mean EPSC amplitude. D, CV as a function of mean ISI (ms). Symbols represent cAMP conditions: 10 μm cAMP (n = 4; X), 100 μm cAMP (n = 12; gray ●), 200 μm cAMP (n = 5; orange rectangle), 500 μm cAMP (n = 6; black ○), 500 μm db-cAMP (n = 12; blue ▿), and 500 μm db-cAMP at 35°C (n = 8; red ▶). Fourty-seven cells contributed to this plot, and an individual cell could have contributed multiple points.
Next, we quantified the regularity of spike timing with the CV of the ISI histograms (Fig. 5B). Smaller CV values correspond to narrower histograms and more regularly-timed spike intervals. Figure 5D plots CV as a function of mean ISI measured for all cAMP conditions for (n = 47), of which eight were in 500 μm db-cAMP between 35°C and 37°C. Recall (from Materials and Methods) that we drove each cell to spike at different rates to allow us to compare CV across neurons at similar spike rates. Consequently, there are >47 points on the plot because each cell contributes more than one point.
We conducted a slope regression analysis to test the influence of ISI and cAMP condition on CV. Spike-train regularity was significantly dependent on ISI (F(1,130) = 7.015, p = 0.0092) but not on cAMP condition (F(4,130) = 0.32, p = 0.8624). In other words, this analysis suggests that once we account for the effect that increasing cAMP has on ISI (recall from Fig. 5C that cAMP condition does affect spike rate or the inverse of ISI), cAMP has no further influence on regularity. For example, even in 500 μm db-cAMP, were the activation range of IH is most depolarized and ISIs were as short as 10 ms, firing patterns were still very irregular (with CV values > 0.45; Fig. 5D, blue triangles). This is counter to the findings in neonatal calyx terminals (Horwitz et al., 2014), where increasing cAMP or blocking IH reduced CV by an order of magnitude (ranging from CV = 0.8–0.08).
Increasing the temperature from 25°C to 35°C in 500 μm db-cAMP did not significantly change mean firing rates (30.4 ± 2.9 spikes/s, n = 11 vs 32.6 ± 3.2 spikes/s, n = 8; p = 0.56). There is significant overlap between the red triangles and blue triangles in Figure 5D, except for a few spike trains with CV < 0.5 (which we discuss in the next section). These results suggest that regular spike trains do not emerge even with faster channel kinetics at higher temperature.
Overall, firing patterns were more irregular here than observed by Kalluri et al. (2010), who recorded from neonatal VGNs without explicitly controlling the activation of IH. Thus, although activation of IH via cAMP was sufficient to increase firing rates, it did not produce highly regular firing.
Developmental changes in firing patterns indicate acquisition of IKL
An explanation for why VGNs failed to produce highly regular firing can be seen by examining the firing pattern of each cell in response to step currents. In Figure 6A, we recolored the symbols in Figure 5D according to step-evoked firing pattern. Triangles indicate recordings made between 35°C and 37°C. Transient-firing neurons (Fig. 6A) had the highest CV values (blue symbols), sustained-B firing neurons had intermediate CV values (gray symbols), and sustained-A firing neurons had the lowest CV values (red symbols). One sustained-A spiking neuron encountered in 500 μm db-cAMP at 35°C produced the most regular spike train in this study and was similar to that seen for sustained-A neurons by Kalluri et al. (2010) (Fig. 6A, red triangle with CV ∼0.2). This suggests that the firing pattern is the dominant predictor for spike-train regularity.

Age-dependent changes in firing pattern and passive membrane properties. A, The influence of firing-type on spike-timing regularity. Same data as in Figure 5D but separated by step-evoked firing pattern; transient-firing (blue; n = 29), sustained-B (gray; n = 14), and sustained-A (red; n = 4). Data obtained in 35°C are distinguished by the symbol ▶ (n = 8). Shaded regions represent the CV range reported by Kalluri et al. (2010) before the first 2 postnatal weeks. B–D, Data from the present study from more mature neurons (open symbols; n = 130) are pooled with that from the younger cells reported by Kalluri et al. (2010) (solid symbols; n = 192). B, Examples of VGN firing types in response to a 750 ms long current step. Bar plots showing the proportion of cells that fall into each firing-pattern group from the first, second, and third postnatal weeks. C, Resting potential becomes more negative with age for both firing types. During early postnatal days, the resting potential for sustained-firing cells (red triangles) were more depolarized than for transient-firing cells (blue squares). As indicated by the convergence of the two trend lines, this difference diminished at older ages. Trend lines are exponential fits to the pooled dataset with 95% confidence intervals (shaded regions) for transient (blue) and sustained (red) firing neurons D, Input resistance (Rin) as a function of membrane capacitance (Cm), for 56 sustained and 71 transient neurons. On average, Cm is larger for older neurons, regardless of firing pattern. Rin becomes smaller for older sustained-spiking neurons. Together, A–D follow the idea that sustained-spiking cells are acquiring additional potassium currents as they mature.
The above described firing-pattern-dependent stratification in CV is qualitatively consistent with that previously reported in younger VGNs (Kalluri et al., 2010) but is quantitatively very different. The overall range of CV values is much more compressed within the irregular/high CV region at the older ages in this study than at younger ages (shaded regions indicate the range of CVs reported by Kalluri et al., 2010). One striking difference between the two studies is that here we saw fewer cells that responded to current injections with sustained-A spike patterns, which appears to be a developmentally driven change. Figure 6B illustrates that the proportion of VGNs that fire with sustained-firing patterns diminishes during the first 3 postnatal weeks. During the first postnatal week, equal numbers of cells had either transient-firing or sustained-firing patterns (Fig. 6B); among the sustained-firing group ∼12% had a fully sustained-A firing pattern. These proportions are very different by the third postnatal week when nearly 70% of cells had transient-firing patterns with very few cells having sustained-A firing patterns (∼5%).
A shift toward the more phasic sustained-B-spiking and transient-spiking patterns is consistent with an increase in the expression of low-voltage-gated potassium currents (Iwasaki et al., 2008; Kalluri et al., 2010; Yoshimoto et al., 2015). Also, consistent with this suggestion, we found that there was an age-dependent hyperpolarization in resting potential (Vrest) and decrease in input resistance (Rin) for all cell types. In Figure 6, C and D, we pooled data from the present study (n = 146) with those from Kalluri et al. (2010) (n = 192) to illustrate the age-dependent changes in these passive membrane properties. Resting potentials of sustained-spiking (red) and transient-spiking (blue) VGNs were different during early postnatal days but begin to converge with that of transient firing neurons by the third postnatal week (Fig. 6C). Membrane capacitance (Cm) was typically larger in the older dataset (Fig. 6D) with transient-firing neurons being larger (40.9 ± 1.4 pF, n = 77) than sustained-firing neurons (31.4 ± 1.7 pF, n = 60; t(132.3) = 4.45 p = 0.0001, unpaired t test). When compared by cell size, Rin values were smaller in transient-firing neurons than sustained-firing neurons at younger ages (Fig. 6D, red triangles, blue squares,), but the differences are less striking at older ages. These data suggest that the ion channel properties of VGNs are becoming less heterogeneous with development. This developmental shift in firing patterns likely accounts for why we did not see highly regular firing, despite enhancing the activation of IH.
Modeling the influence of IH activation in the presence and absence of IKL
Our initial prediction was that by recording from older ages and increasing the activation of IH we would have seen increased sustained-A spiking in response to current steps and more regular spiking in response to pseudo-synaptic stimulation. However, on average, we saw the opposite result in that VGN responses shifted toward transient/irregular spiking. Next, we will show through model simulations that this result can be accounted for by considering the interaction between IKL and IH, both of which are present in nearly all VGNs at older ages.
We used a single-compartment conductance-based vestibular ganglion model in which we could control the conductance density of IKL and the activation of IH (see Materials and Methods; Hight and Kalluri, 2016). Sustained-A and sustained-B firing patterns were simulated (Fig. 7A1,B1) by setting gKL to 0 and 0.65 (mS/cm2), respectively. The equations describing gKL are the same as those described by Hight and Kalluri (2016) for currents carried through an equal combination of Kv1 and Kv7 type ion channels. Sodium channel density (gNa) was set to 20 mS/cm2 for all simulations. This value was large enough to avoid spike train adaptation because of sodium channel inactivation (Hight and Kalluri, 2016). A family of 1500 ms step depolarizing currents was used to elicit spike trains for each condition. Figure 7, A1 and B1, shows responses at current threshold. The conductance density for the HCN channel, gH, was set from 0.07 through 0.91 mS/cm2. This range is between 0.5 and 7 times the mean value we measured in VGNs (0.13 mS/cm2). Here we show the result for the largest value because conductance density for IH is reported to be larger in calyx terminals than in somata (Horwitz et al., 2014). We repeated simulations with two IH conditions that represented the current's most hyperpolarized and depolarized voltage-dependent activation (minimum and maximum activation, representative of the 100 μm cAMP and 500 μm db-cAMP conditions, respectively). The parameters describing the voltage activation range and kinetics for these conditions were based on the data shown in Figure 3 and are represented by Equations 3–5 (see Materials and Methods; parameters are summarized in Table 1).

The influence of shifting the activation range of IH explored in a conductance-based model. Model results from sustained-A and -B type VGNs in response to a 1500 ms depolarizing current step (A3, B3, bottom). Two IH conditions are shown in each panel; minimum activation (black) and maximum activation (dashed red). A, B, The sustained-A type model was produced by setting gNa to 20 mS/cm2 and gKL to 0. The sustained-B type model was produced by setting gNa to 20mS/cm2 and gKL to 0.65 mS/cm2 composed equally of Kv1 and Kv7. A1, B1, Voltage-clamp responses for the sustained-A and -B models under model conditions for IH current (minimum activation, where V1/2 = −96 mV and maximum activation, where V1/2 = −84 mV. Black triangle (▴), The max activation produced spontaneous firing at rest. B, Sustained-B neurons exhibit accommodation as voltage oscillations are present in all IH conditions after ∼60 ms following the onset of the stimulus. A2, B2, IKL current (pA/cm2) as a function of time for A and the shaded region in B. A2, No IKL current is seen because gKL is set to 0 for the sustained-A type VGN. B2, The IKL current at rest and during the AHP grows larger as IH is activated. Hatched region, To estimate the net charge (Q/cm2) carried by IKL we took the sum of the area under each minimum, which is proportional to the shaded region. A3, B3, IH current (pA/cm2) as a function of time for A and the shaded region in B. A3–B3, As expected the IH current is larger at rest and throughout the stimulus for max activation cases than for the minimum-activation case. Note that because of the slow kinetics of the HCN channel, the current of IH switches from inward (−) to outward (+) during the upswing of the spike.
Activation of IH excites faster spiking in the absence of IKL
Figure 7A1–A3 shows the firing patterns and flow of currents through gH and gKL for the sustained-A simulation. Only the beginning of each spike train is shown to emphasize the details of spike shape and timing. Figure 7, A2 and A3, shows the flow of IH and IKL currents into and out of the cell (negative and positive currents, respectively). When IH was minimally activated, very little IH current flows into the cell before current injection begins (i.e., the resting phase; Fig. 7A3). During current injection, the membrane potential depolarizes and the model produces spikes throughout the current step. Between each spike (as the cell emerges from the afterhyperpolarization), a little IH current flows into the cell when the membrane potential is more negative than the reversal potential for IH (−40 mV). When the membrane potential crosses the reversal potential, IH current flows out of the cell. When the activation range of IH is shifted in the maximum activation condition (dashed trace), there is significant IH flowing into the cell during rest and between spikes. As a result of this extra depolarizing current, the model begins to spike spontaneously (Fig. 7A1, black triangles). The flow of inward and outward currents through the IH conductance is also larger during the current injection resulting in shorter mean intervals between spikes (11.97 ± 0.12 vs 12.32 ± 0.01 ms) and more evoked spikes during the step (139 vs 122 spikes). Also note that the spikes are much shorter in the maximum activation condition because the slow inactivation of IH means that the channel remains open during the action potential, adding another large repolarizing potassium current past its reversal potential. Note that because of the spontaneous-spiking, the increase in spike rate during the current step is not immediately visible but, becomes more obvious by ∼60 ms after stimulus onset (black arrow). Overall, the excitatory influence of IH is consistent with the pace-making role of this current in cardiac cells (DiFrancesco, 2010) and with previous suggestions for vestibular afferents (Horwitz et al., 2014; Yoshimoto et al., 2015).
Activation of IH dampens spiking in the presence of IKL
The influence of IH on firing patterns is different in the presence of IKL. In Figure 7B1–B3, we show a simulated sustained-B firing-pattern by setting gKL to 0.65 mS/cm2. Again, the flow of IKL and IH currents during the simulation are shown under the firing pattern (Fig. 7B2,B3). As expected for a model containing IKL there is some outward current flowing during the resting phase (Fig. 7B2). This translates to a more hyperpolarized resting potential for the sustained-B firing pattern (Fig. 7B) than for the sustained-A firing pattern (Fig. 7A). During current injection, the spike trains exhibited accommodation as the IKL current prevented the simulation from reaching threshold, as evidenced by the loss of spikes and emergence of voltage oscillations in the later portions of the current step. In the maximum IH activation condition, there was much more inward IH current in the resting condition and a resulting depolarization in resting potential. However, this depolarization shifts the membrane potential further into the activation range for gKL thereby also increasing the amount of outward flowing IKL. The average outward flowing IKL is larger in the maximum activation condition than in the minimum activation condition. This illustrates that activation of IH recruited more IKL both at rest and during spiking (Fig. 7B2). This recruitment results in spike trains accommodating faster in the maximum-activation condition than in the minimum-activation condition (Fig. 7B1). Thus, in the presence of IKL, IH has a net dampening effect on spiking.
The influence of IH depends on IKL density and channel composition
In Figure 7, we showed single examples in which IH could both excite and dampen spiking, depending on the presence of IKL. In further simulations we varied the size of gKL from 0 to 0.8 mS/cm2 (sustained-A through transient) while keeping all other conductance densities fixed to explore whether the effect on spiking depends on the relative densities of the two conductances. The composition of gKL was comprised of one of the following: entirely Kv1 (Fig. 8, green), entirely Kv7 (blue), or equal proportions of the two conductances (magenta). We elicited spike trains using a fixed-length (1500 ms), fixed amplitude (55 pA), suprathreshold current step. Simulations were repeated in both the maximum (solid symbols) and minimum (open symbols) activation conditions for IH to examine the influence of activating IH. Figure 8A plots the number of spikes evoked by the current step as function of gKL when comprised entirely by Kv1 type channels. As gKL increases, the number of spikes evoked by the current step decreases. Beyond a threshold value, there is a dramatic transition, after which spike number drops precipitously with increasing gKL. There are effectively two regimes to the response. First, there is a tonic regime where many spikes are evoked during the current injection and firing patterns are largely sustained-A. Second, there is a phasic regime where firing patterns are sustained-B and transient.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
IKL is the dominant influence on spike number and spike timing. Model VGN with gKL values between 0.0 and 0.8 mS/cm2, Kv1 to Kv7 ratios for IKL of 1.0 Kv1(green, ● or ○), 50% Kv1 and Kv7 (magenta, ■ or □), or 1.0 Kv7 (blue, ▴ or Δ) were driven with a 1500 ms depolarizing current step (55 pA). Each model VGN had either minimum (open symbols) and maximum IH (solid symbols) IH activation. Trend curves through the data are shown as solid or dashed lines, respectively. A, Only spikes during the current step were used to compute spike number. Spike number as a function of gKL (1.0 Kv1) for min and max IH activation. Spike number falls as a gKL increases and drops sharply once a large enough gKL is reached and because of rapid adaption of the spike train (data not shown). Max activation of IH provides a modest boost in spike number over min activation but requires a smaller gKL value to illicit adaptation. B, Spike number as a function of gKL for three model VGNs with IKL ratios of 1.0 Kv1, Kv1/Kv7, and 1.0 Kv7. For all three model VGNs, spike number falls as gKL increases and drops sharply once a large enough gKL is reached and adaption occurs. This gKL value is smallest in 1.0 Kv7 and largest in 1.0 Kv1 model VGNs. Max activation of IH condition requires a smaller gKL value to elicit accommodation for both the 1.0 Kv1 and 1.0Kv7 model VGNs. C, Spike number as a function of net charge carried by IKL (Q IKL, pC/cm2). Activation of IH provides a modest increase in spike number regardless of net charge by IKL or composition of IKL. D, CV as a function of net charge carried by IKL for model VGNs that produced at least 10 spikes. CV increased as net charge by IKL increased. CV values overall were highly regular (CV < 0.05) because of the lack of irregularity imposed by the step stimulus.
Because gKL can be comprised of different channel subtypes, we also examined the influence of channel composition by repeating the above simulations with 100% Kv7 or equal proportions of Kv1 and Kv7 (Fig. 8B). Recall that the Kv1 and Kv7 channel models have the same voltage-gated activation range but differ in that Kv1 activates quickly and experiences inactivation whereas the Kv7 activates slowly but does not inactivate (Eqs. 6,7). Regardless of channel composition, spike numbers decreased with increasing gKL. However, the amount of gKL needed to elicit the transition between the tonic and phasic regimes was much smaller when gKL included Kv7 components. Also, spike number was more sensitive to increases in Kv7 than to increases in Kv1 (seen as a steeper slope in the spike number vs gKL curve for the Kv7 and mixed conditions). These differences in the effective potency of Kv7 comes from the differences in the net steady-state current produced by the two models; because Kv7 does not inactivate, it produces more steady-state current that the Kv1 model, which experiences significant inactivation. Differences in the time course of activation also impacts firing pattern. Because Kv1 activates much more quickly than Kv7, the transition between the tonic and phasic regimes is much more abrupt for the Kv1 case. In contrast, for both the Kv7 and mixed Kv7/Kv1 conditions, the spike rate (proportional to total number of spikes) slows downs substantially before the transition to the phasic regime.
When IH is maximally activated, the influence of gKL is enhanced, and the transition between the tonic and phasic regimes occurs at smaller gKL values than in the minimum IH condition (Fig. 8A,B, compare dashed lines to dotted lines within each gKL condition). The influence of shifting the activation of IH is very different in the regimes before and after the transition. In the tonic regime, the maximum activation condition produces more spikes than the minimum activation condition (i.e., the excitatory effect from the previous figure). However, as gKL increases, the relative gain in spike numbers provided by IH diminishes. After the transition, the maximum activation condition produces the opposite effect in that for any fixed value of gKL, there is a decrease in the number of spikes evoked (i.e., the type of dampening seen in Fig. 7B). Together, these results show that the outward flowing potassium currents carried by gKL counteract the excitatory influence of IH. Moreover, more gKL is recruited when the activation range of IH is depolarized toward the activation range of gKL.
To examine the influence of IH in isolation from the complexities of gKL recruitment and kinetics, we plotted the spike number and the regularity of the spike train as a function of QKL, the net subthreshold charge delivered by gKL (Fig. 8C,D, respectively). We computed QKL as the area under the lower envelop of the IKL versus time curve (Fig. 7B2, hatched area). Note that this is not the same as the net charge delivered by gKL during the entire spike train, but rather an estimate of the current during the subthreshold portions of the spike-train. As expected, spike number decreases as QKL increases. Now, however spike number falls along a single curve rather than three parametric curves for each combination of Kv1/Kv7. In this plot, we see that the influence of IH alone (when isolated from its ability to recruit IKL) is excitatory in that it produces more spikes. This is true in both the tonic and phasic regimes (compare open symbols to closed symbols).
To examine the influence of IH on spike-timing regularity in the simulations, we computed the CV values for the step-evoked spike trains that produced >10 spikes (Fig. 8D); meaning that regularity was only computed within the tonic regime. We make three main observations pertaining to CV. First, all CV values were highly regular (CV 0.005–0.05). We attributed this to the stimulus type, compared with the randomly arriving EPSC trains, steps of current impose relatively little irregularity onto the sustained-A model. Second, overall CV increased with QKL. Because the transition from the tonic to phasic regime was most abrupt for the Kv1 dominated gKL (Fig. 8B), the change in regularity was also most abrupt for this channel composition (Fig. 8D, compare green symbols to blue + magenta). Third, activation of IH produced only modest changes in CV between 2 and 8 pC/cm2 (Fig. 8D, compare solid and dashed trend lines). Spike-timing regularity is much more sensitive to variations in low-voltage-gated potassium currents than to IH. In other words, highly-regular spiking could not be produced by simply increasing IH, but only occurred with small QKL. Interestingly, CV depends on QKL non-monotonically, with a small upturn toward higher CV values at the lowest QKL. In this regime, the step-evoked spike trains are non-adapting with spikes arriving with clock-like precision. Because CV is the ratio of the SD to the mean of spike intervals, the decrease in mean intervals combined with invariance in the SD produces the increase in CV. Similarly, maximizing IH within this regime produces faster spiking but also produces higher CVs.
Discussion
A developmental convergence in VGN firing patterns
Here we considered whether highly-regular spiking relies on HCN channels, known to be present in both VGN and calyx terminals (Almanza et al., 2012; Meredith et al., 2012; Horwitz et al., 2014; Yoshimoto et al., 2015). We recorded firing patterns from VGNs after expression for HCN channels is developmentally upregulated (Almanza et al., 2012; Yoshimoto et al., 2015) and we enhanced the activation of HCN-mediated currents (referred to as IH in the text) by increasing intracellular concentrations of cyclic-AMP (Accili et al., 2002; Almanza et al., 2012; Meredith et al., 2012). With HCN-mediated currents so enhanced, we found faster subthreshold depolarizations and faster firing rates, but did not find more regular spiking. A concomitant developmental increase in low-voltage-gated potassium currents (referred to as IKL in the text) likely countered the influence of HCN-mediated currents to prevent highly-regular spiking.
Ample evidence supports the idea that low-voltage-gated potassium currents grow with development. Patch-clamp recordings in VGNs show that the currents are larger in the second postnatal week than in the first (Kalluri et al., 2010). Consistent with this, we found resting potentials hyperpolarized and input resistance decreased with age. Overall, VGN responses became less diverse, converging toward more phasic and irregular firing patterns.
Developmental upregulation in the Kv7 group of low-voltage-gated potassium channels (also known as KCNQ channels) is likely driving this change. Immunohistochemistry and pharmacology showing expression for and regional upregulation of Kv7 channels support this conjecture (Rennie and Streeter, 2006; Pérez et al., 2009; Kalluri et al., 2010; Meredith and Rennie, 2015). Immunolabeling for Kv7 channels is initially confined to neurons innervating the central/striolar zones of the vestibular sensory epithelia and eventually spreads to the peripheral/extrastiolar zones (Hurley et al., 2006). As a consequence, calyx terminals in the peripheral zones have little low-voltage-gated potassium currents during early postnatal days (Songer and Eatock, 2013) but have significant increase in these currents mediated by the second to third postnatal weeks (in gerbils, Meredith and Rennie, 2015; in mice, Ramirez et al., 2018; based on sensitivity to Kv7-specific antagonists, linopiridine or XE991). By P21, nearly all VGNs in mice immunolabel for Kv7 (Rocha-Sanchez et al., 2007). Consistent with this, conversion from transient to sustained spiking can require blocking of both Kv1- and Kv7-mediated low-voltage-gated potassium currents (Kalluri et al., 2010). Together, these results suggest that most vestibular afferents eventually acquire low-voltage-gated potassium currents, driven, in part, by a developmental upregulation in Kv7 channels.
Comparison to previous studies
Iwasaki et al. (2008) described a different developmental trajectory in which a larger fraction of VGNs had tonic/sustained firing patterns as function of development. They interpret this as reflecting an age-dependent increase in HCN channel expression (Yoshimoto et al., 2015) coupled with a relatively age-invariant dependence for Kv1 expression (Iwasaki et al., 2012). This trajectory is satisfying because it describes a pattern of ion channel expression that parallels the in vivo increase in regular-spiking (Curthoys, 1979). However, unlike the present study and those of Soto and colleagues (Limón et al., 2005; Pérez et al., 2009; Almanza et al., 2012), Yoshimoto et al. (2015) recorded from small cells whose size, resting potentials and input resistance did not change with age. Their cells also lacked Kv7-mediated currents because they could convert firing patterns from transient to sustained-firing by only blocking Kv1 channels (Iwasaki et al., 2012). In contrast, we saw a wide range of cells sizes, age-dependent hyperpolarization in resting potential and, in a previous study, showed that it is necessary to block both Kv1 and Kv7 channels to convert firing patterns (Kalluri et al., 2010). Whether or not methodological variations, including differences in enzyme composition and maturational sensitivity to enzymes leads to sampling different subpopulation and/or alters ion channel properties remains to be resolved (Quandt, 1987; Matteson and Carmeliet, 1988; Armstrong and Roberts, 1998).
Horwitz et al. (2014) showed impressive control over spike-timing regularity by blocking or enhancing HCN-mediated currents in the calyx terminals of immature dimorphic afferents. This is likely because immature calyces do not yet have their full complement of potassium currents. Our modeling suggests that the absence of low-voltage-gated potassium currents in immature calyces created the ideal condition for HCN-mediated currents to influence spike-timing regularity. Indeed, Songer and Eatock (2013) found sustained spiking and highly regular spontaneous spiking in early postnatal extrastriolar calyxes, indicating that these neurons lacked low-voltage-gated potassium currents. In contrast, striolar calyces have these currents and respond to injected currents with transient/phasic spiking and spontaneously spike with irregular timing. We also considered whether HCN-mediated currents could exert a stronger influence on spike timing at calyx terminals where the current's densities are larger than they are at somata (Horwitz et al., 2014). Our modeling argues against this line of reasoning because even with seven times the HCN channel density found in the somata, the size of low-voltage-gated potassium currents was still the dominant factor controlling spike timing. These results suggest that differences in the size of HCN-mediated currents between terminals and somata do not account for the prevalence of phasic and irregular firing in the present study.
Ion channel mechanisms controlling spike-timing
HCN-channels are permeable to both sodium and potassium ions, which gives them a reversal potential ∼−40 mV (which is near spike threshold in VGN; DiFrancesco, 1981; present study). At subthreshold potentials, current flows into the cell through HCN channels. This steady inward current depolarizes the membrane potential toward spike threshold driving highly-regular auto-rhythmic firing (DiFrancesco, 2010; Horwitz et al., 2014). This follows our observations in 500 μm db-cAMP, where we found steeper afterhyperpolarization potentials and faster spike rates that are akin to those seen in vivo (Fig. 4). This role of HCN-mediated currents in shaping the afterhyperpolarization potential is similar in spirit (if not in the details) to the phenomenological “afterhyperpolarization current” model for explaining spike-timing regularity in vestibular neurons (Smith and Goldberg, 1986). Because HCN channels inactivate slowly, the resulting steady inward current can raise resting potential and increase net conductance, simultaneously sharpening excitatory postsynaptic potentials and speeding action potential timing (in the auditory nerve, Yi et al., 2010; in the vestibular nerve, Meredith et al., 2012).
HCN-mediated currents influence spiking very differently when they are found with low-voltage-gated potassium currents. As the inward flowing HCN-mediated current depolarizes a neuron, the membrane potential comes closer into the activation range of low-voltage-gated potassium channels. When these potassium channels open, the net outward current works against the HCN-mediated inward current to pull the membrane potential away from spike threshold, making it harder for the neuron to reach spike threshold. How quickly spiking is inhibited depends on the type of low-voltage-gated potassium channel. Fast-activating currents from Kv1 channels quickly tamp-down spiking, whereas the slower activating currents mediated by Kv7 channels often allow several spikes to trigger before the outward current is large enough to squelch spiking (Fig. 7).
The interactions between multiple ion channels determine neuronal excitability, which is a general feature of neuronal biophysics. The role of any particular channel is greatly influenced by the complement of other channels found within neurons' membrane. In early development, HCN channels in vestibular afferents can be excitatory and drive auto-rhythmic spiking, but, their role flips with the maturational upregulation of potassium channels. This interaction between HCN-mediated and low-voltage-gated potassium currents is similar to that described in the auditory brainstem (Bal and Oertel, 2000; Rothman and Manis, 2003; McGinley and Oertel, 2006; Mathews et al., 2010; Cao and Oertel, 2011; Khurana et al., 2012), where HCN-mediated currents enhance the inhibitory influence of low-voltage-gated potassium currents. These results suggest that although HCN-mediated currents grow during maturation, they counter-intuitively suppress VGN excitability by recruiting larger low-voltage-gated potassium currents. This drives maturing VGNs in vitro toward phasic and irregular spike-timing statistics.
If low-voltage-gated potassium currents are ubiquitously upregulated in vestibular afferents, then existing models accounting for spiking-timing regularity need revision. Our results argue against the hypothesis that developmental upregulation in HCN channel expression is responsible for developmental increases in regular-spiking in vivo. The ion channel complement controlling spike-timing is becoming more complex during maturation. Added complexity comes with added flexibility. In maturing VGN, low-voltage-gated potassium currents are driven through by both Kv1 and Kv7 channels. The Kv7-mediated currents can be modulated by cholinergic efferent neurons acting on muscarinic acetylcholine receptors. Muscarinic acetylcholine receptors are a G-protein-coupled receptors that trigger secondary-messenger cascades that ultimately close Kv7 channels (Selyanko et al., 1992; Selyanko and Brown, 1993; for review, see Brown, 2010; for review, see Greene and Hoshi, 2017).
.Closing Kv7 channels in VGNs (either directly by blocking the channels or indirectly using the mAChR agonist oxotremorine-M) decreases membrane conductance, converts in vitro firing patterns from high-threshold transient-firing to low-threshold sustained-firing (Pérez et al., 2009) and increases the regularity of spontaneous firing in extra-striolar calyces (Ramirez et al., 2018). Consistent with this, turtle vestibular efferent neurons close Kv7 channels to excite a slow increase of in vivo afferent spike rate (Holt et al., 2017). Similarly, blocking Kv7 channels suppresses in vivo afferent response to transient head motions in mice (likely encoded by irregular afferents; Lee et al., 2017). This suggests that maturing vestibular afferents acquire ion channel properties than can be dynamically controlled, with some neurons possibly acquiring the flexibility to move between the regular- and irregular-firing categories, perhaps based on the tone established by descending efferent projections. This is interesting for neurons outside the vestibular system that also require control over their spike timing and suggests one way in which this level of dynamic control can be achieved.
Our results also suggest that temperature differences (e.g., room temperature vs body temperature) do not significantly account for the differences in spike-timing regularity between in vitro and in vivo experiments. We previously found that increasing recording temperature speeds up ion channel kinetics and spike rates (present study; Kalluri et al., 2010). Similarly, spike rates are nearly six times faster at body temperature than room temperature in auditory nerve (Wu et al., 2016). Surprisingly, here we found that in vitro spike-timing for VGNs is neither faster nor more regular at body temperature than at room temperature when cAMP levels are already elevated (CV > 0.5 in both cases). At first blush, this seems surprising because increasing body temperature speeds up ion channel kinetics (shown here for HCN channels in VGNs) and is known to depolarize the voltage activation range of HCN channels (Gambardella et al., 2012). However, as our experiments show the combination of elevated cAMP concentrations and elevated temperature are not enough to unveil the highly regular spiking found in vivo (CV < 0.1; Baird et al., 1988). This supports our primary conclusion that highly regular-spiking in vestibular ganglion neurons is limited by the presence of low-voltage-gated potassium channels and not by the size, speed, and activation state of HCN channels.
Ultimately, the ion channel mechanisms we describe in vitro represent only a piece of the puzzle contributing to in vivo responses. Vestibular afferents have diverse dendritic morphologies, connect to different types and numbers of hair cells in different epithelial zones, and express a variety of ion channels (many were not considered here). In addition to ion channels, models have shown that the size and rate of synaptic events also shapes spike-timing statistics. For example, turtle vestibular neurons having small compact terminal arbors with bouton synapses on few hair cells might produce large and temporally brief synaptic events whereas neurons with extended arbors containing hundreds of bouton-like terminals on many spatially dispersed hair cells may produce small but rapidly arriving events (Goldberg and Holt, 2013; Holmes et al., 2017). A sustained-A model can produce remarkably regular firing patterns when stimulated by a steady depolarization, such as that provided by a step of current (Fig. 8) or by the convergence of small but rapidly arriving synaptic events (Hight and Kalluri, 2016). Consistent with this, regular-firing dimorphic afferents in the peripheral zones of canal afferents appear to have more bouton terminals than those in the central zones (Goldberg et al., 1990b). A similar stimulus with small events arriving at high-rate applied to the transient-spiking model cannot even produce a spike train (Hight and Kalluri, 2016). In the future, we will need to consider whether ion channel composition (including efferent modulation) and terminal morphology/connectivity are complimentary specializations, tuned together over development to produce regular and irregular firing patterns.
Footnotes
- Received July 13, 2018.
- Revision received January 14, 2019.
- Accepted January 19, 2019.
This work was supported by NIH NIDCD Grants R03 DC012652, DC012652S, and R01 DC015512, and the Department of Otolaryngology at the University of Southern California. We thank Maya Monges-Hernandez for technical assistance, and Alexander Markowitz and Ruth Anne Eatock for commenting on previous versions of the paper.
The authors declare no competing financial interests.
- Correspondence should be addressed to Radha Kalluri at radha{at}usc.edu
- Copyright © 2019 the authors