
Abstract
Studies have recently suggested that blockade of 5-HT6 receptors (5-HT6R) improves memory processes. As episodic memory alteration is one of the first deficits observed during normal aging and in neurological and neuropsychiatric disorders (Alzheimer's disease, schizophrenia), the present study sought to characterize the effects of 5-HT6R blockade on spatial recognition memory, which can be considered as ‘episodic-like’ memory, in rodents. We quantified the effects of the selective 5-HT6R antagonist SB-271046 (10 mg/kg, i.p.), using the two-trial place recognition task in the Y-maze, on acquisition, consolidation, and retrieval of spatial recognition memory in young adult mice (6-week-old; intertrial intervals (ITIs) 30, 60, 120, 240, and 360 min) and on the consolidation of spatial recognition memory in aged mice (3-, 12-, 18-, and 21–month-old; ITI 60 and 240 min). SB-271046-treated young adult mice explored the new arm more after a 240-min (pre-acquisition) and 360-min (post-acquisition) ITI, whereas vehicle-treated animals failed to discriminate the new arm when the ITI exceeded 120 min (pre-acquisition) or 240 min (post-acquisition). Aged mice, which expressed spatial memory deficits, explored the new arm more after a 60-min ITI (21–month-old) and a 240-min ITI (18- and 21–month-old) when treated with SB-271046. Consequently, 5-HT6R blockade improves spatial recognition memory in adult mice and reverses age-related consolidation deficits of episodic-like memory. This study provides further support for the use of 5-HT6R antagonists in the treatment of episodic memory disorders related to aging as well as neurological disorders such as Alzheimer's disease and schizophrenia.
Similar content being viewed by others

INTRODUCTION
Extensive evidence suggests that 5-hydroxytryptamine (5-HT) plays an important modulatory role in learning and memory through action on multiple receptor subtypes (see for review, Hoyer et al, 2002) and interaction with multiple neurotransmitter systems (Buhot et al, 2000).
Among 5-HT receptors, 5-HT6 receptors (5-HT6R) are predominantly expressed in the rat and mouse central nervous systems, notably in the cerebral cortex, striatum, hippocampus, nucleus accumbens, and olfactory tubercles (Bibancos et al, 2007; Gérard et al, 1997; Monsma et al, 1993; Ruat et al, 1993; Svenningsson et al, 2007; Ward and Dorsa, 1996). Because of their distribution in limbic areas and the cerebral cortex, 5-HT6R are proposed to be involved in cognitive processes, novelty-seeking behavior as well as mood regulation (Ballaz et al, 2007; Barnes and Sharp, 1999; Svenningson et al, 2007; Wesolowska and Nikiforuk, 2007). Functional studies indicated that 5-HT6R exert an inhibitory effect on central cholinergic and glutamatergic neurotransmission and could be valuable targets in the treatment of cognitive disorders in which these neurotransmission systems are altered. Such a hypothesis is further supported by experimental studies showing that 5-HT6R antagonism promotes cognitive processes in the rat. In the Morris water maze paradigm, the selective 5-HT6R antagonists Ro 04-6790, SB-271046, SB-357134, and SB-399885 improved spatial reference memory retrieval in adult rats (Rogers and Hagan, 2001; Stean et al, 2002; Woolley et al, 2001) as well as the acquisition and retrieval of spatial reference memory in aged rats (Foley et al, 2004; Hirst et al, 2006). In the object discrimination task, Ro 04-6790 and SB-271046 increased the acquisition and consolidation of recognition memory in adult rats (Hirst et al, 2006; King et al, 2004), whereas BGC20-761, another 5-HT6R antagonist improved the consolidation of recognition memory in mature rats (Mitchell et al, 2006). Furthermore, Ro 04-6790, SB-271046, Ro4368554, and SB-399885 were all found to reverse the scopolamine-induced deficits in both the long-term memory passive avoidance task (Foley et al, 2004), the novel object discrimination task (recognition memory) (Lieben et al, 2005; Woolley et al, 2003) and the autoshaping learning task (Perez-Garcia and Meneses, 2005) in adult rats. Moreover, reversal of the delay-dependent deficits exerted by Ro 04-6790 and SB-271046 in the rat novel object discrimination task has been shown to be blocked by the NMDA receptor antagonist MK-801 (King et al, 2004). In vivo and in vitro microdialysis studies in the rat showed that 5-HT6R blockade elevates both extracellular acetylcholine and glutamate in the hippocampus and frontal cortex (Dawson et al, 2000, 2001; Marcos et al, 2006; Riemer et al, 2003; Shirazi-Southall et al, 2002; Sleight et al, 1998).
In the present study we investigated the effect of 5-HT6R blockade on spatial recognition memory in the mouse—the first study of its kind to our knowledge. The rationale for this study is threefold: (1) spatial recognition memory in the rodent can be likened to human episodic memory, which refers to the recollection of a unique past experience in terms of what happened, and where and when it happened (Tulving, 2001), and alteration of episodic memory is one of the first deficits observed in humans during normal aging and in neurological disorders such as Alzheimer's disease (Backman et al, 2001; Daselaar et al, 2003); (2) though the significance of 5-HT6R in the mouse is debated (Hirst et al, 2003), these receptors have been recently found in this species at high densities within various brain areas, including the cerebral cortex and hippocampus (Bibancos et al, 2007, Svenningsson et al, 2007) that are known to be pivotal in spatial recognition memory (Kesner et al, 1993; McDonald and White, 1993; Thinus-Blanc et al, 1996); and (3) pharmacological modulation of 5-HT6R in the mouse has been recently demonstrated to modify psychopharmacological performance (Svenningsson et al, 2007; Wesolowska and Nikiforuk, 2007). Thus, we first evaluated in young adult mice the effects of selective blockade of 5-HT6R by SB-271046 (Bromidge et al, 1999; Routledge et al, 2000) on acquisition, consolidation, and retrieval of spatial recognition memory in the two-trial place recognition task in the Y-maze (Dellu et al, 1992, 2000). Because (1) aging is associated with spatial learning impairment attributed to deficiency of information consolidation in the hippocampus (Barnes, 1979; Friedman et al, 2007; Gallagher et al, 1993, 2) memory consolidation has been associated with changes in 5-HT6R levels (Meneses et al, 2007), and (3) very few studies have evaluated the role of 5-HT6R in spatial memory in aged rodents (Foley et al, 2004; Hirst et al, 2006), we also investigated, in a second set of experiments, the effects of blockade of 5-HT6R on consolidation of spatial recognition memory in adult and aged mice.
MATERIALS AND METHODS
Animals
Animals were maintained in a controlled environment (22±2°C; 55±10% humidity) under a 12:12 h light/dark cycle (light on between 2000 and 0800 hours) with food and water available ad libitum. All experiments complied with European Directives and French law on animal experimentation (personal authorizations no. 14-05 for FD, no. 14-17 for MB, and no. 14–62 for PD).
Experiments were performed either on male NMRI mice (6-week old; 28.2±0.2 g) (experiment 1; n=390) from a local breeding facility (F1 from Centre d’Elevage René Janvier, Le Genest, France) or on 3-, 12-, 18-, and 21-month-old (sample sizes 68, 62, 49, and 42, respectively) female NMRI mice (experiment 2), purchased at the age of 3 weeks from the same supplier. Mice were housed in standard polycarbonate cages (42 × 29 × 15 cm) containing 15 (young adults) or 10 (aged) animals. In the aging studies, female mice were used because of their less-aggressive behavior toward other mice and the need to house several animals together for periods of up to 21 months. Moreover, Lamberty and Gower (1988) demonstrated that spatial learning performances between young adult male and female NMRI mice are similar. One week before Y-maze testing, mice were handled daily in their home cage by the experimenter. Aged animals with obvious health problems (tumors, cataracts) were excluded prior to starting the experiments.
Drug Administration
Mice received SB-271046 (5-chloro-N-(4-methoxy-3-piperazin-1-yl-phenyl)-3-methyl-2-benzothiophene sulfonamide hydrochloride), synthesized by Professor F Fabis and Dr M Paillet-Loilier (CERMN, Caen), at a dose of 10 mg/kg based on previous in vitro and in vivo studies in mice and rats (Foley et al, 2004; Hirst et al, 2003; Lindner et al, 2003; Rogers and Hagan, 2001; Svenningsson et al, 2007; Wesolowska and Nikiforuk, 2007; Woolley et al, 2001). The drug was dissolved in 9‰ NaCl as the vehicle and administered i.p. in a volume of 10 ml/kg. Vehicle-treated animals were used for comparison.
General Testing Procedure
Behavioral tests were conducted during the dark phase of the cycle from 0900 to 1800 hours. Thirty minutes before the beginning of the experiments, mice were placed in the experimentation room. The experimental protocol used to evaluate spatial recognition memory was based on Dellu et al (1992, 2000). In short, the Y-maze is made of Plexiglas with three arms, each measuring 34 cm long, 9 cm wide, and 13 cm high. The task consisted of two trials separated by different intertrial intervals (ITIs). During the first trial (training session; 5 min), the animal was placed in the starting arm, one of the two other arms being closed in a random choice. During the second trial (retrieval session; 2 min), animals had free access to the three arms and were allowed to explore the maze. During the two sessions, the time spent in the distal third of each arm was collected.
Experiment 1: Effects of 5-HT6R Blockade on Spatial Recognition Memory Performance in Young Adult Mice
To evaluate the effects of blockade of 5-HT6R on the three phases of the mnesic process, SB-271046 was administered 60 min before the training session (acquisition), immediately after the training session (consolidation) or 60 min before the retrieval session (retention). The 60-min delay was chosen based on the hypothesis that following i.p. administration, the peak concentration of SB-271046 in the blood would be obtained for a much shorter time, and clearance of the antagonist would be more rapid, than after p.o. administration (peak value at 180 min; Routledge et al, 2000). Under this scenario, administration of SB-271046 60 min before the first trial (acquisition) should have a maximal effect on the acquisition process while preserving the consolidation phase. Five ITIs (30, 60, 120, 240, and 360 min) were studied for the acquisition and consolidation phases, whereas only three ITIs (120, 240, and 360 min) could be studied for the retrieval phase because of the pharmacokinetic considerations previously described. For each phase studied and for each ITI, SB-271046-treated mice (n=14–15) were compared to saline-injected animals (n=14–15).
Experiment 2: Effects of 5-HT6R Blockade on Age-Related Spatial Recognition Memory Deficits in Aged Mice
Effects of the blockade of 5-HT6R on deficits in spatial recognition memory of aged mice were measured using the place recognition test. To take into account age-related locomotor, exploratory, visual, and motivational differences, irrespective of the pharmacological treatment, mice were tested within 4 weeks following their completion of the place recognition test for: (1) spontaneous locomotor activity (second week), (2) exploratory behavior (hole-board test; third week), and (3) visuospatial performance (cued learning test in the Morris water maze; fourth week). These delays were planned to rule out any interference with cognitive performance and any pharmacological influence.
Two-trial place recognition test in the Y-maze
On the basis of the results of experiment 1, the effects of blockade of the 5-HT6R on the consolidation phase of spatial recognition memory were evaluated at only two ITIs (60 and 240 min). The 5-HT6R antagonist was administered immediately after the training session. For each age (3, 12, 18, and 21 months) and for each ITI, SB-271046-treated animals (n=10–17) were compared to saline-treated mice (n=10–16).
Spontaneous locomotor activity test
Mice were tested using a photoelectronic actimeter (APELAB) (Boissier and Simon, 1965) that allowed us to automatically count the number of times the animals interrupt perpendicular light beams crossing the center of an individual box. Animals were tested for a total period of 30 min and the number of light-beam interruptions was collected every 5 min.
Hole-board test
Mice were tested in a hole-board test (APELEX) (Boissier and Simon, 1962; Makanjuola et al, 1977) that allowed us to automatically count the number of dips in holes made by the animal during exploration of the hole-board plate. Mice were tested for a total period of 5 min and the number of head dips was collected every 1 min.
Cued learning test in the Morris water maze
The apparatus was similar with the one described by Lelong et al (2001), and the procedure was based on Lamberty and Gower (1990, 1992). Each mouse from each age group performed three trials per day spaced by a 30 s ITI for 2 consecutive days. Escape latency to locate the platform (s), distance traveled (cm), and swim speed (cm/s) were monitored and recorded by means of a video computer-based storage system.
Statistical Analysis
Only those animals that clearly showed an exploratory behavior, ie that did not spend all their time in the start arm, were included in these analyses.
Repeated-measures analyses of variance (ANOVAs) were used for the comparison of the percentage of time spent in each arm (experiment 1: ‘ITI’ and ‘arm’ factors; experiment 2: ‘age’ and ‘arm’ factors) and for the discrimination index (New arm time−(starting arm time+open arm time)/2) (Mitchell et al, 2006) (experiment 1: ‘ITI’ and ‘treatment’ factors; experiment 2: ‘age’ and ‘treatment’ factors). For experiment 2, animals presenting stereotyped behaviors were excluded from the analysis (12 months, n=4; 18 months, n=2) (Table 1). In both experiments, when significant principal effects and interactions between factors were detected, a Student–Newman–Keuls (NK) multiple range test post hoc analysis was performed. When ANOVA detected differences among ITI, for the sake of clarity, only the differences in the percent time in the new arm between the longest ITI tested (360 min) and any other ITI were displayed. Student's t-test was used to compare the percent time in the new arm and the discrimination index to a random choice level of exploration (33.33% and 0 value, respectively) when appropriate.
In the spontaneous locomotor activity and hole-board tests, a repeated-measures ANOVA was used to evaluate the effect of age on the number of beam interruptions (locomotor activity) or head dips in holes (exploratory behavior). For cued learning in the Morris water maze test, a repeated-measures ANOVA was used to evaluate the effect of age on swim distances, escape latencies, and swim speeds. For these three tests also, a post hoc analysis was performed (NK multiple range test) in case of significant principal effects and interactions between factors.
RESULTS
Experiment 1: Effect of 5-HT6R Blockade on Spatial Recognition Memory Performance in Young Adult Mice
Effect of 5-HT6R blockade on acquisition
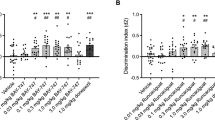
1. Percent time in arm exploration: Saline-treated mice. Analysis of the percent time spent in the three arms for saline-treated mice revealed a significant decrease in the time spent in the new arm as a function of ITI duration (‘ITI effect’ F4, 68=2.73; p<0.05/‘arm effect’ F2, 138=24.2; p<0.001/‘interaction ITI—arm’ F8, 136=5.24; p<0.001) (Figure 1a). Post hoc analysis confirmed this result and revealed significant differences between the 30- and 60-min ITIs when compared to 360-min ITI (p<0.01; NK). The percent time spent in the new arm differed from the percent time spent in the two other arms for the 30- and the 60-min ITIs (p<0.01; NK) and only differed from the percent time spent in the familiar arm for the 120-min ITI (p<0.05; NK). This result was confirmed by a percent time spent in the new arm above the random choice level (33.33%) for these ITI (p<0.05; t-test). For the 240- and 360-min ITIs, the mice had lost their discrimination capabilities (Figure 1a).

Influence of SB-271046 (10 mg/kg, i.p.) on acquisition of spatial recognition memory in the place recognition test in young adult mice. Data represent the mean (±SEM) percent time spent in the three arms during the retrieval session for saline-treated groups (n=14–15 per intertrial interval (ITI)) (a) and for SB-271046-treated groups (n=14–15 per ITI) (b). Following the training session, recall performances of mice were tested during the retrieval session after 30, 60, 120, 240, or 360 min. Mice were injected 60 min before the training session. Data represent the mean (±SEM) discrimination index for saline- and SB-271046-treated aged mice (c). *p<0.05, ***p<0.001 (360-min ITI vs other ITI); †p<0.05, ††p<0.01, †††p<0.001 (new arm vs start or familiar arm) two-way analysis of variance (ANOVA) followed by Newman–Keuls multiple range test. #p<0.05, ##p<0.01, ###p<0.001, Student's t-test vs random choice level of exploration (33.33% value for percent time and 0 value for discrimination index).
SB-271046-treated mice. In contrast, analysis of the percent time spent in the three arms for SB-271046-treated mice revealed a significant effect of arm exploration but not of ITI (‘arm effect’ F2, 138=35.9; p<0.001/‘interaction ITI—arm’ F8, 138=3.58; p<0.001) (Figure 1b). The percent time spent in the new arm differed from the percent time spent in both other arms for the 30-, 60-, and the 120-min ITIs (p<0.05; NK), and only differed from the percent time spent in the familiar arm for the 240-min ITI (p<0.05; NK). For the 360-min ITI, the mice did not explore preferentially any arm (Figure 1b). The percent time spent in the new arm was above the random choice level for the 30-, 60-, 120-, and 240-min ITIs (p<0.05; t-test) but not for the 360-min ITI (Figure 1b).
2. Discrimination index: Analysis of the discrimination index for the saline- and SB-271046-treated animals revealed a significant effect of ITI but not of treatment (‘ITI effect’ F4, 112=2.84; p<0.001) (Figure 1c). The discrimination indices for the SB-271046-treated mice differed from ‘absence of discrimination’ for longer ITI than for the saline-treated mice, up to 240 min for the former (p<0.05) compared with 60 min for the latter (p<0.01) (Figure 1c).
Effect of 5-HT6R blockade on consolidation
1. Percent time in arm exploration: Saline-treated mice. Analysis of the percent time spent in the three arms for saline-treated mice revealed a difference in exploration among the three arms whatever the ITI duration (‘Arm effect’ F2, 136=30.3; p<0.001). Nevertheless, the ‘ITI effect’ and the interaction between the two parameters were almost significant (F4, 68=2.04; p=0.09/F8, 136=30.3; p=0.08; respectively), as shown in Figure 2a; the figure also revealed a tendency to a decrease in the time spent in the new arm as a function of the duration of the ITI. The percent time spent in the new arm differed from the random choice level for all but the 360 min ITI (p<0.05; t-test) (Figure 2a).

Influence of SB-271046 (10 mg/kg, i.p.) on consolidation of spatial recognition memory in the place recognition test in young adult mice. Data represent the mean (±SEM) percent time spent in the three arms during the retrieval session for saline-treated groups (n=14–15 per intertrial interval (ITI)) (a) and for SB-271046-treated groups (n=14–15 per ITI) (b). Following the training session, mice were injected immediately and their recall performances were tested during the retrieval session after 30, 60, 120, 240, or 360 min. Data represent the mean (±SEM) discrimination index for saline- and SB-271046-treated aged mice (c). #p<0.05, ##p<0.01, ###p<0.0001, Student's t-test vs random choice level of exploration (33.33% value for percent time and 0 value for discrimination index).
SB-271046-treated mice. Analysis of the percent time spent in the three arms for SB-271046-treated mice revealed that animals discriminate the new arm at all the ITI tested (‘arm effect’ F2, 136=28.4; p<0.001) (Figure 2b). The percent time spent in the new arm was clearly above the random choice level for all the ITI (p<0.05; t-test).
2. Discrimination index: Analysis of the discrimination index for saline- and SB-271046-treated animals revealed a significant effect of ITI but not of treatment (ITI effect’ F4, 104=2.58; p<0.05) (Figure 2c). Saline-treated mice discriminated for ITI shorter than 360 min (p<0.05; t-test) though mice only tend to discriminate for the 120-min ITI (p=0.17; t-test), (Figure 2c). In contrast, the discrimination indices of SB-271046-treated mice differed from the ‘absence of discrimination’ for all ITIs tested (p<0.05; t-test) (Figure 2c).
Effect of 5-HT6R blockade on retrieval
1. Percent time in arm exploration: Saline-treated mice. Analysis of the percent time spent in the three arms for saline-treated mice revealed a significant effect of arm exploration but not of ITI (‘arm effect’ F2, 80=18.6; p<0.001/‘interaction effect—arm’ F4, 80=4.85; p<0.01) (Figure 3a). The percent time spent in the new arm differed from the time spent in the two other arms for both the 120- and the 240-min ITIs (p<0.01; NK), and only differed from the percent time spent in the start arm for the 360-min ITI (p<0.05; NK). The percent time spent in the new arm is higher than the random choice level for the 120- and 240-min ITIs only (p<0.01; t-test) (Figure 3a).

Influence of SB-271046 (10 mg/kg, i.p.) on retrieval of spatial recognition memory in the place recognition test in young adult mice. Data represent the mean(±SEM) percent time spent in the three arms during the retrieval session for saline-treated groups (n=15 per intertrial interval (ITI)) (a) and for SB-271046-treated groups (n=15 per ITI) (b). Following the training session, recall performances of mice were tested during the retrieval session after 120, 240, and 360 min. Mice were injected 60 min before the retrieval session. Data represent the mean (±SEM) discrimination index for saline- and SB-271046-treated aged mice (c). †p<0.05, ††p<0.01, †††p<0.001 (new arm vs start or familiar arm) two-way analysis of variance (ANOVA) followed by Newman–Keuls multiple range test. ##p<0.01, ###p<0.0001, Student's t-test vs random choice level of exploration (33.33% value for percent time and 0 value for discrimination index).
SB271046-treated mice. Similar results were observed for SB-271046-treated mice. (‘arm effect’ F2, 82=13.2; p<0.001/‘interaction effect—arm’ F4, 82=2.52; p<0.05) (Figure 3b). The percent time spent in the new arm differed from the percent time spent in the two other arms for both the 120- and the 240-min ITIs (p<0.01; NK) but not for the 360-min ITI. The percent time spent in the new arm differed from the random choice level only for the 120- and 240-min ITIs (p<0.01; t-test) (Figure 3b).
2. Discrimination index: Analysis of the discrimination index for saline- and SB-271046-treated animals revealed a significant effect of ITI but not of treatment (‘ITI effect’ F2, 50=4.45; p<0.05) (Figure 3c). The discrimination indices of both saline-treated and SB-271046-treated mice differed from the ‘absence of discrimination’ for the 120- and 240-min ITIs only (p<0.01; t-test) (Figure 3c).
Experiment 2: Effect of 5-HT6R Blockade on Age-Related Spatial Recognition Memory Deficits in Aged Mice
Two-trial place recognition test in the Y-maze
On the basis of the results of experiment 1, the effects of blockade of the 5-HT6R on the consolidation phase of spatial recognition memory were evaluated using 60-min and 240-min ITIs, intertrial intervals for which saline-treated young adult mice were able to discriminate the new arm and for which we could expect older animals to express memory deficits, especially for long ITI such as 240 min.
1. Percent time in arm exploration—60 min ITI: Saline-treated mice. Analysis of the percent time spent in the three arms for saline-treated mice for the 60-min ITI revealed a significant effect of arm exploration but not of age (‘arm effect’ F2, 88=9.76; p<0.01/‘interaction age—arm’ F6, 88=2.62; p<0.05) (Figure 4a). However, the percent time spent in the new arm differed from the time spent in the two other arms for the 3- and 18-month-old mice (p<0.05; NK). The percent time spent in the new arm differed from the random choice level for the 3- and 18-month-old mice (p<0.05; t-test) but not for the two other ages (Figure 4a). The 12-month-old mice did not show a preference for the new arm, probably due to an unusually long time spent in the start arm. In contrast to the 12 month-old, 21-month-old mice spent the same amount of time in the start arm as the 3- and 18-month-old mice but did not discriminate the new arm, more likely reflecting an age-related memory deficit (Figure 4a).

Effect of SB-271046 (10 mg/kg, i.p.) on consolidation of spatial recognition memory in the place recognition test in aged mice. Data represent the mean (±SEM) percent time spent in the three arms during the retrieval session for 3- (n=16), 12- (n=12), 18- (n=10), and 21- (n=10) month-old mice treated by saline (a) and for 3- (n=14), 12- (n=14), 18- (n=12), and 21- (n=10) month-old mice treated with SB-271046 (b). Following the training session, mice were injected immediately and their recall performances were tested in the retrieval session after a 60 min ITI. Data represent the mean (±SEM) discrimination index for saline- and SB-271046-treated aged mice (c). †p<0.05, ††p<0.01, (new arm vs start or familiar arm) two-way analysis of variance (ANOVA) followed by Newman–Keuls multiple range test. #p<0.05, ##p<0.01, ###p<0.0001, Student's t-test vs chance level of exploration (33.33% value for percent time and 0 value for discrimination index).
SB-271046-treated mice. For SB-271046-treated animals, analysis of the percent time spent in the three arms for the 60-min ITI revealed a significant effect of arm exploration, but found no age-related differences (‘arm effect’ F2, 92=22.3; p<0.001) (Figure 4b). In contrast to the saline-treated mice, the percent time spent in the new arm was above the random choice level for all tested ages (p<0.05; t-test) (Figure 4b).
2. Discrimination index—60 min ITI: These results were confirmed by the statistical analysis performed on the discrimination index for saline- and SB-271046-treated animals (‘age effect’ F3, 51=2.84; p<0.05/‘treatment effect’ F1, 17=4.16; p=0.06) (Figure 4c). In contrast to the SB-271046-treated mice that discriminate at all ages, saline-treated animals discriminate only for the 3- and 18-month-old ages (p<0.05; t-test) (Figure 4c).
3. Percent time in arm exploration—240 min ITI: Saline-treated mice. Analysis of the percent time spent in the three arms for saline-treated mice for the 240-min ITI revealed a significant effect of arm exploration but not of age (‘arm effect’ F2, 96=4.40; p<0.05/‘interaction age—arm’ F6, 96=3.84; p<0.01) (Figure 5a). Post hoc analysis revealed that only for the 3-month-old mice the percent time spent in the new arm was different from the percent time spent in any of the two other arms (p<0.01; NK) and above the random choice level (p<0.001; t-test). For the 240 min ITI, 12-month-old and older saline-treated mice did not spend more time in the new arm compared to the other arms, although we cannot rule out the possibility that the 12-month-old mice were able to discriminate the new arm but did not appear to do so because they had spent an unusually long time in the start arm (Figure 5a).

Effect of SB-271046 (10 mg/kg, i.p.) on consolidation of spatial recognition memory in the place recognition test in aged mice. Data represent the mean (±SEM) percent time spent in the three arms during the retrieval session for 3- (n=17), 12- (n=15), 18- (n=10), and 21- (n=10) month-old mice treated with saline (a) and for 3- (n=15), 12- (n=14), 18- (n=12), and 21- (n=11) month-old mice treated with SB-271046 (b). Following the training session, mice were injected immediately and their recall performance was tested in the retrieval session after 240 min. Data represent the mean (±SEM) discrimination index for saline- and SB-271046-treated aged mice (c). *p<0.05 (saline- vs SB-271046-treated mice), ††p<0.01, †††p<0.001 (new arm vs start or familiar arm) two-way analysis of variance (ANOVA) followed by Newman–Keuls multiple range test. ##p<0.01, ###p<0.0001, Student's t-test vs chance level of exploration (33.33% value for percent time and 0 value for discrimination index).
SB-271046-treated mice. Analysis of the percent time spent in the three arms for this duration of ITI for the SB-271046-treated animals revealed a significant effect of arm exploration but found no age-related differences (‘arm effect’ F2, 96=49.8; p<0.001) (Figure 5b). In contrast to saline-treated mice, the percent time spent in the new arm for SB-271046-treated animals differed from the random choice level for all tested ages (p<0.01; t-test) (Figure 5b).
4. Discrimination index—240 min ITI: Statistical analysis of the discrimination index for the saline- and SB-271046-treated animals confirmed these results, showing significant effects of treatment and age (‘treatment effect’ F1, 19=7.71; p<0.05/‘age effect’ F3, 57=3.70; p=0.05) (Figure 5c). Post hoc analysis showed significant differences between the saline- and SB-271046-treated groups for the 12- and 21-month-old mice (p<0.05; NK), whereas such differences failed to reach statistical significance for the 3- and 18-month-old mice. When compared with SB-271046-treated animals, the discrimination index for the saline-treated animals differed from the ‘absence of discrimination’ only for the 3-month-old mice (p<0.01; t-test) (Figure 5c).
Spontaneous locomotor activity test
Analysis of the locomotor activity of animals measured for 30 min revealed a difference between the various age groups (F3, 208=12.64; p<0.001), a result confirmed by a post hoc analysis (see Table 1 for details). When locomotor activity was measured for six consecutive 5-min periods, analysis of the number of beam interruptions revealed a difference in locomotor activity between the different age groups as well as a decrease in locomotor activity as a function of the duration of the test (‘age effect’ F3, 208=12.6; p<0.001/‘time effect’ F5, 1040=266; p<0.001/‘interaction age—time’ F15, 1040=3.80; p<0.001) (Table 1). Post hoc analysis indeed revealed differences between the various age groups for each 5-min interval of the test (p⩽0.01 for all 5-min periods).
Hole-board test
Analysis of the number of dips in holes for a total period of 5 min revealed a significant difference between the different age groups (F3, 207=7.57; p<0.001). Post hoc analysis confirmed this and indicated that if there was basically no difference in activity between age groups for the three last 1-min periods, older mice dipped less their head in the holes during the first two 1-min periods (see Table 2 for details).
Cued learning test in the Morris water maze
Analysis showed a reduction in the swim distance between the 2 days of testing without age-related differences (‘day of testing effect’ F1, 204=162; p<0.001) and significant age- and day-related differences in escape latency (‘age effect’ F3, 204=9.98; p<0.001/‘day of testing effect’ F1, 204=176; p<0.001) (Table 3). Analysis of swim speed revealed only an effect of age (‘age effect’ F3, 204=13.4; p<0.001/‘interaction age—day of testing’ F3, 204=3.45; p<0.05) (Table 3). Post hoc analysis revealed that the swim speeds of the 12-, 18-, and 21-month-old mice were significantly lower than the swim speed of the 3-month-old animals (see Table 3 for details).
DISCUSSION
In the present work, we demonstrate, for the first time to our knowledge, that the selective blockade of 5-HT6R, as obtained through the use of SB-271046, improves the acquisition and consolidation of spatial recognition memory in young adult mice and counteracts age-related deficits in consolidation of spatial recognition memory.
Discrimination performance toward the new arm by young adult mice decreased as a function of the increase in ITIs between the training and retrieval sessions in the two-trial place recognition task. Indeed, vehicle-treated mice failed to discriminate the new arm when the ITI exceeded 120 or 240 min for animals treated in the pre- or post-acquisition period, respectively. In contrast, SB-271046-treated mice explored the new arm more than the two other arms for ITIs 240 and 360 min long (pre- and post-acquisition treatments, respectively), a result in favor of an increase in the mnesic trace of spatial information. As SB-271046 failed to modify the retrieval performance when administered prior to the retrieval session, such an effect is specific to acquisition and consolidation. On the basis of pharmacokinetic considerations (Routledge et al, 2000), we speculate that SB-271046 administered 60 min before the first trial (acquisition) should interfere to a lesser extent with the consolidation processes than when injected just after the first session, and should thus mainly affect the acquisition phase. This hypothesis seems to be supported by the fact that the beneficial effect of 5-HT6R blockade seems more robust on consolidation (Figure 2b) than on acquisition because this latter effect shows a tendency to decrease with time (Figure 1b). Therefore, we suggest that 5-HT6R antagonists modulate the post-acquisition stage rather than the acquisition stage of learning and memory processes. Given the robust and long-lasting positive effect of blockade of 5-HT6R on the consolidation phase, it would be worthwhile to evaluate if this beneficial effect is still present for ITI longer than 360 min. Our results are consistent with previous studies performed in the rat demonstrating that 5-HT6R blockade has positive effects on acquisition and consolidation, but not on retrieval, of nonspatial mnesic information (King et al, 2004). The effects of 5-HT6R blockade on acquisition, consolidation, and retrieval seem however to depend on the type of memory under study (ie reference and recognition memory). Indeed, 5-HT6R blockade was without effect on the acquisition of spatial reference memory (Hirst et al, 2006; Lindner et al, 2003; Rogers and Hagan, 2001; Stean et al, 2002, Woolley et al, 2001), whereas it increases acquisition of nonspatial (object recognition test; King et al, 2004) and spatial (place recognition test; present work) recognition memory. 5-HT6R blockade improves memory consolidation in a nonspatial recognition task (King et al, 2004) and an autoshaping Pavlovian/instrumental learning task (Meneses, 2001; Perez-Garcia and Meneses, 2005). 5-HT6R blockade had no effect on the retrieval of nonspatial (object recognition test, King et al, 2004) and spatial (place recognition test, present work) recognition memory, whereas retrieval of spatial reference memory has been found to be either unchanged (Lindner et al, 2003) or improved in similar conditions (Rogers and Hagan, 2001; Woolley et al, 2001).
Considering the beneficial effect of blockade of 5-HT6R on the consolidation process of new information in spatial recognition memory and the fact that such a process is impaired during aging (Barnes, 1979; Gallagher et al, 1993; Wilson et al, 2005), we have also investigated the influence of 5-HT6R antagonism on the consolidation of spatial recognition memory in young adult, adult, and aged mice. Overall, our results strongly suggest that 5-HT6R blockade reverses age-related deficits in the consolidation of spatial recognition memory, which began at 21 months of age for the 60-min ITI and at 18 months for the 240-min ITI (Figures 4 and 5). As the antagonist was administered during the consolidation period, we conclude that 5-HT6R blockade improves and normalizes the consolidation process of spatial information in aged mice. On the basis of the discrimination indices (Figures 4c and 5c), there was a substantial effect of SB-271046 on the 12-month-old mice, apparently by countering the absence of discrimination that was observed in 12-month-old control (saline-treated) mice (see below). However, by comparing directly the percent time spent in the new arm with reference to a random exploration (random choice level of 33.33%) for the 60-min ITI (Figures 4a and b), one can conclude there is a rather small effect of SB-271046 on both 12- and 18-month-old mice. Though no other work has evaluated the effect of 5-HT6R blockade on the consolidation of spatial recognition memory in aged animals, our results are consistent with previous studies in the rat that demonstrate an improvement of task acquisition and recall in a spatial learning paradigm in 20- and 22-month-old rats following acute or chronic 5-HT6R antagonism (Foley et al, 2004; Hirst et al, 2006). These age-related deficits, consistent with previous studies on spatial memory (Bach et al, 1999; Barnes, 1979; Gallagher and Pelleymounter, 1988, Gower and Lamberty, 1993; Granger et al, 1996), are attributable to dysfunctions in memory processes on the basis that impaired locomotor activity in the two-trial place recognition test in the Y-maze could not be mistaken for a memory deficit because the test is based on the choice between a novel place and familiar places (Dellu et al, 1992, 2000) and the percentage of time spent in exploration is used as the measure. Indeed, in our hands, exploratory behavior was found to be only slightly reduced in the hole-board test (Table 2), and such an effect can probably be linked to the decrease in spontaneous locomotor activity (Table 1). Moreover, analyses of swim speed, escape latency, and distance traveled observed in the cued learning Morris water maze test are consistent with an absence of visual and motivational deficits (Table 3). As already shown (Ammassari-Teule et al, 1994; Dellu et al, 1992; Lamirault et al, 2003), the total exploration time during the second trial of the recognition memory test was found to be similar for the 3-, 12-, 18-, and 21-month-old mice, suggesting that exploratory behavior in aged mice did not decrease with age (online Supplementary Figure S1). Consequently, the slight age-related decrease in spontaneous activity observed in our control tests as well as by others (Lamberty et al, 1989, 1990; Francia et al, 2006) is assumed to have little or no consequence in the evaluation of memory performance as it is not linked to a reduction in exploratory activity during the memory test itself. Control tests were also important for the interpretation of the unexpected lack of preference for the new arm exhibited by 12-month-old, saline-treated, mice (Figures 4 and 5). This result, probably due to the unusually long time mice spent in the start arm, could be linked to either an increased anxiety or a reduced motivation to explore the maze as well as to the implication of intra- or extra-maze cues. None of these hypotheses could however find support as (1) the motivational indices of the cued learning test (Table 3) as well as the times spent in the start and open familiar arms during the first session were similar in all animal groups (online Supplementary Figure 2), (2) olfactory traces were reduced and homogenized between each passage in the Y-maze as recommended (Dellu et al, 1992), and (3) parallel experiments revealed that the random allocation of the start arm did not modify this specific behavior of the animals (data not shown). Moreover, the absence of preference for the new arm is not linked to a difference in locomotor activity because 12-month-old mice showed no significant difference in performance compared to younger animals (0–5 min interval, a time equivalent to the 2 min duration of the second trial in the Y-maze; Table 1).
Spatial memory formation involves several brain areas, including the hippocampus, rhinal cortices, and neocortical regions such as the parietal cortex (Holscher, 2003; Thinus-Blanc et al, 1998). Indeed, studies suggest that the hippocampus (1) is dedicated to the creation and use of spatial or cognitive maps and is therefore critical for the encoding and expression of relationships between items of memory (Eichenbaum, 1999; Kesner et al, 1991) and (2) is important for short-term consolidation of spatial information and supervises consolidation of spatial memory in the entorhinal cortex before some transfer occurs to the neocortex for permanent storage (Cho and Kesner, 1996; Steckler et al, 1998). The hippocampus, as well as brain regions of the medial temporal lobe, is essential for declarative memory and notably episodic memory (Squire and Zola, 1996). Human episodic memory refers to the recollection of a unique past experience in terms of what happened, and where and when it happened (Tulving, 2001). Interestingly, recent studies suggest that nonhuman mammals have the ability to build higher-order memory for unique events that incorporate information about what, where, and when, lending strong support to the idea that animals are endowed with episodic-like memory (Dere et al, 2005a, 2005b). In the place recognition task used in the present study, spatial recognition information could be considered as episodic-like information. Young adult mice remembered the arms visited during the training session (what and where) after a delay of 240 min (when), and consequently preferentially explored the new arm during the retrieval session. On the basis of the high density of 5-HT6R in the brain regions implicated in spatial memory processes (Bibancos et al, 2007, Gérard et al, 1997; Ruat et al, 1993; Ward and Dorsa, 1996) and the results previously discussed that strongly suggest that the blockade of 5-HT6R improves the acquisition and consolidation of episodic-like memory in young-adult mice and counteracts age-related deficits in such a memory process, 5-HT6R antagonists are proposed as a therapeutic alternative for the treatment of episodic memory deficits which are among the first signs of cognitive decline related to aging and neurological disorders such as Alzheimer's disease (Friedman et al, 2007; Small et al, 2003). The mechanisms by which 5-HT6R antagonists can counteract age-related deficits in memory processes are yet unknown. One can however hypothesize a withdrawal of the inhibition exerted by serotonin onto the cholinergic system, notably in the hippocampus (Foley et al, 2004; Lieben et al, 2005; Perez-Garcia and Meneses, 2005; Riemer et al, 2003; Shirazi-Southall et al, 2002)—this latter being largely altered during aging (Buhot et al, 2000; Decker and McGaugh, 1991; McEntee and Crook, 1992)—and/or an interaction with the hippocampal glutamatergic system, leading to an increase in synaptic plasticity (eg long-term potentiation; Bliss and Collingridge, 1993; Dawson et al, 2000, 2001; Foster, 1999; Lynch et al, 2006; Morris, 2006), which is altered during aging (see for review Lynch et al, 2006). Other concepts have been recently proposed (see Perez-Garcia and Meneses, 2008).
In summary, we have demonstrated that 5-HT6R blockade improves spatial recognition memory in young adult mice and reverses age-dependent deficits in a place recognition task. Our results suggest that 5-HT6R could be important in the consolidation of the episodic-like information of episodic-like memory. Thus, 5-HT6R antagonists could be of great therapeutic value in the treatment of cognitive disorders where deficits in episodic memory are a major feature, including aging, Alzheimer's disease, and schizophrenia. Interestingly, 5-HT6R antagonists such as LY-483518, SB-742457, and PRX-07034 have already entered clinical trials for the treatment of cognitive impairment associated with Alzheimer's disease and schizophrenia, and other antagonists are under active development (Holenz et al, 2006; www.epixmed.com).
References
Ammassari-Teule M, Fagioli S, Rossi-Arnaud C (1994). Radial maze performance and open-field behaviours in aged C57BL/6 mice: further evidence for preserved cognitive abilities during senescence. Physiol Behav 55: 341–345.
Bach ME, Barad M, Son H, Zhuo M, Lu YF, Shih R et al (1999). Age-related defects in spatial memory are correlated with defects in the late phase of hippocampal long-term potentiation in vitro and are attenuated by drugs that enhance the cAMP signaling pathway. Proc Natl Acad Sci USA 96: 5280–5285.
Backman L, Small BJ, Fratiglioni L (2001). Stability of the preclinical episodic memory deficit in Alzheimer's disease. Brain 124: 96–102.
Ballaz SJ, Akil H, Watson SJ (2007). Analysis of 5-HT6 and 5-HT7 receptor gene expression in rats showing differences in novelty-seeking behavior. Neuroscience 147: 428–438.
Barnes CA (1979). Memory deficits associated with senescence: a neurophysiological and behavioral study in the rat. J Comp Physiol Psychol 93: 74–104.
Barnes NM, Sharp T (1999). A review of central 5-HT receptors and their function. Neuropharmacology 38: 1083–1152.
Bibancos T, Jardim DL, Aneas I, Chiavegatto S (2007). Social isolation and expression of serotonergic neurotransmission-related genes in several brain areas of male mice. Genes Brain Behav 6: 529–539.
Bliss TV, Collingridge GL (1993). A synaptic model of memory: long-term potentiation in the hippocampus. Nature 361: 31–39.
Boissier JR, Simon P (1962). [The exploration reaction in the mouse. Preliminary note.]. Therapie 17: 1225–1232.
Boissier JR, Simon P (1965). [Action of caffeine on the spontaneous motility of the mouse]. Arch Int Pharmacodyn Ther 158: 212–221.
Bromidge SM, Brown AM, Clarke SE, Dodgson K, Gager T, Grassam HL et al (1999). 5-Chloro-N-(4-methoxy-3-piperazin-1-yl- phenyl)-3-methyl-2-benzothiophenesulfon- amide (SB-271046): a potent, selective, and orally bioavailable 5-HT6 receptor antagonist. J Med Chem 42: 202–205.
Buhot MC, Martin S, Segu L (2000). Role of serotonin in memory impairment. Ann Med 32: 210–221. Review.
Cho YH, Kesner RP (1996). Involvement of entorhinal cortex or parietal cortex in long-term spatial discrimination memory in rats: retrograde amnesia. Behav Neurosci 110: 436–442.
Daselaar SM, Veltman DJ, Rombouts SA, Raaijmakers JG, Jonker C (2003). Neuroanatomical correlates of episodic encoding and retrieval in young and elderly subjects. Brain 126: 43–56.
Dawson LA, Nguyen HQ, Li P (2001). The 5-HT(6) receptor antagonist SB-271046 selectively enhances excitatory neurotransmission in the rat frontal cortex and hippocampus. Neuropsychopharmacology 25: 662–668.
Dawson LA, Nguyen HQ, Li P (2000). In vivo effects of the 5-HT(6) antagonist SB-271046 on striatal and frontal cortex extracellular concentrations of noradrenaline, dopamine, 5-HT, glutamate and aspartate. Br J Pharmacol 130: 23–26.
Decker MW, McGaugh JL (1991). The role of interactions between the cholinergic system and other neuromodulatory systems in learning and memory. Synapse 7: 151–168.
Dellu F, Contarino A, Simon H, Koob GF, Gold LH (2000). Genetic differences in response to novelty and spatial memory using a two-trial recognition task in mice. Neurobiol Learn Mem 73: 31–48.
Dellu F, Mayo W, Cherkaoui J, Le Moal M, Simon H (1992). A two-trial memory task with automated recording: study in young and aged rats. Brain Res 588: 132–139.
Dere E, Huston JP, De Souza Silva MA (2005a). Episodic-like memory in mice: simultaneous assessment of object, place and temporal order memory. Brain Res Brain Res Protoc 16: 10–19.
Dere E, Huston JP, De Souza Silva MA (2005b). Integrated memory for objects, places, and temporal order: evidence for episodic-like memory in mice. Neurobiol Learn Mem 84: 214–221.
Eichenbaum H (1999). The hippocampus and mechanisms of declarative memory. Behav Brain Res 103: 123–133.
Foley AG, Murphy KJ, Hirst WD, Gallagher HC, Hagan JJ, Upton N et al (2004). The 5-HT(6) receptor antagonist SB-271046 reverses scopolamine-disrupted consolidation of a passive avoidance task and ameliorates spatial task deficits in aged rats. Neuropsychopharmacology 29: 93–100.
Foster TC (1999). Involvement of hippocampal synaptic plasticity in age-related memory decline. Brain Res Brain Res Rev 30: 236–249.
Francia N, Cirulli F, Chiarotti F, Antonelli A, Aloe L, Alleva E (2006). Spatial memory deficits in middle-aged mice correlate with lower exploratory activity and a subordinate status: role of hippocampal neurotrophins. Eur J Neurosci 23: 711–728.
Friedman D, Nessler D, Johnson Jr R (2007). Memory encoding and retrieval in the aging brain. Clin EEG Neurosci 38: 2–7.
Gallagher M, Burwell R, Burchinal M (1993). Severity of spatial learning impairment in aging: development of a learning index for performance in the Morris water maze. Behav Neurosci 107: 618–626.
Gallagher M, Pelleymounter MA (1988). Spatial learning deficits in old rats: a model for memory decline in the aged. Neurobiol Aging 9: 549–556.
Gérard C, Martres MP, Lefevre K, Miquel MC, Verge D, Lanfumey L et al (1997). Immuno-localization of serotonin 5-HT6 receptor-like material in the rat central nervous system. Brain Res 746: 207–219.
Gower AJ, Lamberty Y (1993). The aged mouse as a model of cognitive decline with special emphasis on studies in NMRI mice. Behav Brain Res 57: 163–173.
Granger R, Deadwyler S, Davis M, Moskovitz B, Kessler M, Rogers G et al (1996). Facilitation of glutamate receptors reverses an age-associated memory impairment in rats. Synapse 22: 332–337.
Hirst WD, Abrahamsen B, Blaney FE, Calver AR, Aloj L, Price GW et al (2003). Differences in the central nervous system distribution and pharmacology of the mouse 5-hydroxytryptamine-6 receptor compared with rat and human receptors investigated by radioligand binding, site-directed mutagenesis, and molecular modeling. Mol Pharmacol 64: 1295–1308.
Hirst WD, Stean TO, Rogers DC, Sunter D, Pugh P, Moss SF et al (2006). SB-399885 is a potent, selective 5-HT6 receptor antagonist with cognitive enhancing properties in aged rat water maze and novel object recognition models. Eur J Pharmacol 553: 109–119.
Holenz J, Pauwels PJ, Diaz JL, Merce R, Codony X, Buschmann H (2006). Medicinal chemistry strategies to 5-HT(6) receptor ligands as potential cognitive enhancers and antiobesity agents. Drug Discov Today 11: 283–299.
Holscher C (2003). Time, space and hippocampal functions. Rev Neurosci 14: 253–284.
Hoyer D, Hannon JP, Martin GR (2002). Molecular, pharmacological and functional diversity of 5-HT receptors. Pharmacol Biochem Behav 71: 533–554.
Kesner RP, Bolland BL, Dakis M (1993). Memory for spatial locations, motor responses, and objects: triple dissociation among the hippocampus, caudate nucleus, and extrastriate visual cortex. Exp Brain Res 93: 462–470.
Kesner RP, Farnsworth G, Kametani H (1991). Role of parietal cortex and hippocampus in representing spatial information. Cereb Cortex 1: 367–373.
King MV, Sleight AJ, Woolley ML, Topham IA, Marsden CA, Fone KC (2004). 5-HT6 receptor antagonists reverse delay-dependent deficits in novel object discrimination by enhancing consolidation—an effect sensitive to NMDA receptor antagonism. Neuropharmacology 47: 195–204.
Lamberty Y, Gower AJ (1988). Investigation into sex-related differences in locomotor activity, place learning and passive avoidance responding in NMRI mice. Physiol Behav 4: 787–790.
Lamberty Y, Gower AJ (1990). Age-related changes in spontaneous behavior and learning in NMRI mice from maturity to middle age. Physiol Behav 47: 1137–1144.
Lamberty Y, Gower AJ (1992). Age-related changes in spontaneous behavior and learning in NMRI mice from middle to old age. Physiol Behav 51: 81–88.
Lamirault L, Guillou C, Thal C, Simon H (2003). (−)-9-Dehydrogalanthaminium bromide, a new cholinesterase inhibitor, enhances place and object recognition memory in young and old rats. Neurobiol Learn Mem 80: 113–122.
Lelong V, Dauphin F, Boulouard M (2001). RS 67333 and D-cycloserine accelerate learning acquisition in the rat. Neuropharmacology 41: 517–522.
Lieben CK, Blokland A, Sik A, Sung E, van Nieuwenhuizen P, Schreiber R (2005). The selective 5-HT6 receptor antagonist Ro4368554 restores memory performance in cholinergic and serotonergic models of memory deficiency in the rat. Neuropsychopharmacology 30: 2169–2179.
Lindner MD, Hodges Jr DB, Hogan JB, Orie AF, Corsa JA, Barten DM et al (2003). An assessment of the effects of serotonin 6 (5-HT6) receptor antagonists in rodent models of learning. J Pharmacol Exp Ther 307: 682–691.
Lynch G, Rex CS, Gall CM (2006). Synaptic plasticity in early aging. Ageing Res Rev 5: 255–280.
McDonald RJ, White NM (1993). A triple dissociation of memory systems: hippocampus, amygdala, and dorsal striatum. Behav Neurosci 107: 3–22.
McEntee WJ, Crook TH (1992). Cholinergic function in the aged brain: implications for treatment of memory impairments associated with aging. Behav Pharmacol 3: 327–336.
Makanjuola RO, Hill G, Maben I, Dow RC, Ashcroft GW (1977). An automated method for studying exploratory and stereotyped behaviour in rats. Psychopharmacology (Berl) 52: 271–277.
Marcos B, Gil-Bea FJ, Hirst WD, Garcia-Alloza M, Ramirez MJ (2006). Lack of localization of 5-HT6 receptors on cholinergic neurons: implication of multiple neurotransmitter systems in 5-HT6 receptor-mediated acetylcholine release. Eur J Neurosci 24: 1299–1306.
Meneses A (2001). Role of 5-HT6 receptors in memory formation. Drug News Perspect 14: 396–400.
Meneses A, Manuel-Apolinar L, Castillo C, Castillo E (2007). Memory consolidation and amnesia modify 5-HT6 receptors expression in rat brain: an autoradiographic study. Behav Brain Res 178: 53–61.
Mitchell ES, Hoplight BJ, Lear SP, Neumaier JF (2006). BGC20-761, a novel tryptamine analog, enhances memory consolidation and reverses scopolamine-induced memory deficit in social and visuospatial memory tasks through a 5-HT6 receptor-mediated mechanism. Neuropharmacology 50: 412–420.
Monsma Jr FJ, Shen Y, Ward RP, Hamblin MW, Sibley DR (1993). Cloning and expression of a novel serotonin receptor with high affinity for tricyclic psychotropic drugs. Mol Pharmacol 43: 320–327.
Morris RG (2006). Elements of a neurobiological theory of hippocampal function: the role of synaptic plasticity, synaptic tagging and schemas. Eur J Neurosci 23: 2829–2846.
Perez-Garcia G, Meneses A (2005). Oral administration of the 5-HT6 receptor antagonists SB-357134 and SB-399885 improves memory formation in an autoshaping learning task. Pharmacol Biochem Behav 81: 673–682.
Perez-Garcia G, Meneses A (2008). Memory formation, amnesia, improved memory and reversed amnesia: 5-HT role. Behav Brain Res 2008; e-pub ahead of print 21 January 2008.
Riemer C, Borroni E, Levet-Trafit B, Martin JR, Poli S, Porter RH et al (2003). Influence of the 5-HT6 receptor on acetylcholine release in the cortex: pharmacological characterization of 4-(2-bromo-6-pyrrolidin-1-ylpyridine-4-sulfonyl)phenylamine, a potent and selective 5-HT6 receptor antagonist. J Med Chem 46: 1273–1276.
Rogers DC, Hagan JJ (2001). 5-HT6 receptor antagonists enhance retention of a water maze task in the rat. Psychopharmacology (Berl) 158: 114–119.
Routledge C, Bromidge SM, Moss SF, Price GW, Hirst W, Newman H et al (2000). Characterization of SB-271046: a potent, selective and orally active 5-HT(6) receptor antagonist. Br J Pharmacol 130: 1606–1612.
Ruat M, Traiffort E, Arrang JM, Tardivel-Lacombe J, Diaz J, Leurs R et al (1993). A novel rat serotonin (5-HT6) receptor: molecular cloning, localization and stimulation of cAMP accumulation. Biochem Biophys Res Commun 193: 268–276.
Shirazi-Southall S, Rodriguez DE, Nomikos GG (2002). Effects of typical and atypical antipsychotics and receptor selective compounds on acetylcholine efflux in the hippocampus of the rat. Neuropsychopharmacology 26: 583–594.
Sleight AJ, Boess FG, Bos M, Bourson A (1998). The putative 5-ht6 receptor: localization and function. Ann NY Acad Sci 861: 91–96.
Small BJ, Mobly JL, Laukka EJ, Jones S, Backman L (2003). Cognitive deficits in preclinical Alzheimer's disease. Acta Neurol Scand Suppl 179: 29–33.
Squire LR, Zola SM (1996). Structure and function of declarative and nondeclarative memory systems. Proc Natl Acad Sci USA 93: 13515–13522.
Stean TO, Hirst WD, Thomas DR, Price GW, Rogers D, Riley G et al (2002). Pharmacological profile of SB-357134: a potent, selective, brain penetrant, and orally active 5-HT(6) receptor antagonist. Pharmacol Biochem Behav 71: 645–654.
Steckler T, Drinkenburg WH, Sahgal A, Aggleton JP (1998). Recognition memory in rats—II. Neuroanatomical substrates. Prog Neurobiol 54: 313–332.
Svenningsson P, Tzavara ET, Qi H, Carruthers R, Witkin JM, Nomikos GG et al (2007). Biochemical and behavioural evidence for antidepressant-like effects of 5-HT6 receptor stimulation. J Neurosci 27: 4201–4209.
Thinus-Blanc C, Save E, Poucet B (1998). The associative parietal cortex and spatial processing in rodents. C R Acad Sci III 321: 179–183.
Thinus-Blanc C, Save E, Poucet B, Foreman N (1996). Effects of parietal cortex lesions on spatial problem solving in the rat. Behav Brain Res 81: 115–121.
Tulving E (2001). Episodic memory and common sense: how far apart? Philos Trans R Soc Lond B Biol Sci 356: 1505–1515.
Ward RP, Dorsa DM (1996). Colocalization of serotonin receptor subtypes 5-HT2A, 5-HT2C, and 5-HT6 with neuropeptides in rat striatum. J Comp Neurol 370: 405–414.
Wesolowska A, Nikiforuk A (2007). Effects of the brain-penetrant and selective 5-HT6 receptor antagonist SB-399885 in animal models of anxiety and depression. Neuropharmacology 52: 1274–1283.
Wilson IA, Ikonen S, Gallagher M, Eichenbaum H, Tanila H (2005). Age-associated alterations of hippocampal place cells are subregion specific. J Neurosci 25: 6877–6886.
Woolley ML, Bentley JC, Sleight AJ, Marsden CA, Fone KC (2001). A role for 5-ht6 receptors in retention of spatial learning in the Morris water maze. Neuropharmacology 41: 210–219.
Woolley ML, Marsden CA, Sleight AJ, Fone KC (2003). Reversal of a cholinergic-induced deficit in a rodent model of recognition memory by the selective 5-HT6 receptor antagonist, Ro 04-6790. Psychopharmacology 170: 358–367.
Acknowledgements
We thank Professor Frederic Fabis and Dr Magalie Paillet-Loilier (CERMN, Caen) for synthesis of the 5-HT6R antagonist SB-271046. We also thank Christiane Cliquet and Claudine Fauchon for technical assistance and gratefully acknowledge the helpful suggestions of Professor Raymond Chichery and the final language revision of Dr Daryl S Henderson (Squirrel Scribe). Virginie Da Silva Costa is supported by funding from the French Ministère de l’Education Nationale, de l’Enseignement Supérieur et de la Recherche.
Author information
Authors and Affiliations
Corresponding author
Additional information
DICLOSURE/CONFLICTS OF INTERESTS
The authors report no conflict of interest.
Supplementary Information accompanies the paper on the Neuropsychopharmacology website (http://www.nature.com/npp)
Rights and permissions
About this article
Cite this article
Da Silva Costa, V., Duchatelle, P., Boulouard, M. et al. Selective 5-HT6 Receptor Blockade Improves Spatial Recognition Memory and Reverses Age-Related Deficits in Spatial Recognition Memory in the Mouse. Neuropsychopharmacol 34, 488–500 (2009). https://doi.org/10.1038/npp.2008.94
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/npp.2008.94
Keywords
This article is cited by
-
5-HT6 receptor blockade differentially affects scopolamine-induced deficits of working memory, recognition memory and aversive learning in mice
Psychopharmacology (2012)
-
The role of serotonin in the NMDA receptor antagonist models of psychosis and cognitive impairment
Psychopharmacology (2011)
-
Correlations among central serotonergic parameters and age-related emotional and cognitive changes assessed through the elevated T-maze and the Morris water maze
AGE (2010)
