
Abstract
The endocannabinoid-inactivating enzyme, fatty acid amide hydrolase (FAAH), and the transient receptor potential vanilloid type-1 (TRPV1) channel are new targets for the development of anxiolytic drugs. We studied the effect on anxiety-like behavior in the elevated plus maze of a dual FAAH/TRPV1 blocker, N-arachidonoyl-serotonin (AA-5-HT). In male C57BL/6J mice, acute intraperitoneal administration of AA-5-HT (0.1–2.5 mg/kg) increased both the time spent and the number of entries in the open arm, while being inactive at the highest dose tested (5 mg/kg). AA-5-HT was more potent than selective blockers of FAAH or TRPV1 (URB597 and SB366791, respectively). In male Swiss mice, AA-5-HT had to be administered chronically to observe an anxiolytic effect at an intermediate dose (2.5 mg/kg), the highest dose (5 mg/kg) being anxiogenic, and 1 mg/kg being ineffective. In both strains, the anxiolytic effects of AA-5-HT were paralleled by elevation of brain endocannabinoid levels and were reversed by per se inactive doses of the cannabinoid receptors of type-1 (CB1) receptor antagonist AM251, or the TRPV1 agonist, olvanil. Immunohistochemical localization of CB1 and TRPV1 receptors was observed in mouse prefrontal cortex, nucleus accumbens, amygdala, and hippocampus. Simultaneous ‘indirect’ activation of CB1 receptors following FAAH inhibition, and antagonism at TRPV1 receptors might represent a new therapeutic strategy against anxiety.
Similar content being viewed by others

INTRODUCTION
The G-protein-coupled cannabinoid receptors of type-1 (CB1) and the transient receptor potential vanilloid type-1 (TRPV1) channels act as molecular targets for some long-chain fatty acid amides, including anandamide (Devane et al, 1992; Zygmunt et al, 1999) and N-arachidonoyl-dopamine (see Huang et al, 2002) (Di Marzo et al, 2007; Starowicz et al, 2007a, for reviews). Anandamide homologues, like oleoyl- and linoleoyl-ethanolamide (Movahed et al, 2005), and lipoxygenase products, like 12-hydroperoxy-eicosatetraenoic acid (Hwang, 2000), activate TRPV1 but not CB1 receptors, whereas 2-arachidonoyl-glycerol (2-AG) (Mechoulam et al, 1995; Sugiura et al, 1995) is a selective endogenous cannabinoid receptor ligand with no activity at TRPV1 (De Petrocellis et al, 2000). A function has been established for brain presynaptic CB1 receptors as the targets of retrogradely acting endocannabinoids produced from postsynaptic neurons and inhibiting neurotransmitter release (Freund et al, 2003). The role in the brain of TRPV1, previously thought to only function in sensory neurons, is also emerging (see Kim et al, 2007; Starowicz et al, 2008, for reviews). Studies with selective CB1 and TRPV1 antagonists, and CB1−/− and TRPV1−/− mice suggest that endocannabinoids and endovanilloids tonically activate the two receptors and control several brain functions by exerting opposing neuromodulatory effects (Marinelli et al, 2005, 2007; Marsch et al, 2007; Starowicz et al, 2007b; Rubino et al, 2008).
Affective behaviors are among the functions that appear to be controlled by both CB1 and TRPV1 receptors. Depending on the dose, the animal species and its emotional background state, and the behavioral test used, CB1 agonists can both suppress and worsen anxiety-like behaviors in rodents (see Lafenetre et al, 2007; Viveros et al, 2007; Bisogno and Di Marzo, 2007, for reviews). Importantly, inhibitors of endocannabinoid degradation, and in particular blockers of the enzyme fatty acid amide hydrolase (FAAH), such as URB597 (Kathuria et al, 2003), which are capable to activate the targets of the endocannabinoids by prolonging the lifespan of the latter, exert anxiolytic effects (Kathuria et al, 2003; Patel and Hillard, 2006; Rubino et al, 2008; Moreira et al, 2008). However, Naidu et al (2007) could only find a weak anxiolytic-like effect of URB597 in the elevated plus maze (EPM) test of anxiety-like behavior under certain procedural conditions. Apart from differences in experimental protocols, these discrepancies might be due to the fact that, by inhibiting FAAH, compounds like URB597 also enhance the levels also of nonendocannabinoid FAAH substrates, such as the TRPV1 agonist, oleoylethanolamide. Likewise, the elevation of anandamide levels caused by FAAH inhibitors can also lead to the indirect activation of other proposed targets for this compound (Di Marzo et al, 2002; Oz, 2006), including TRPV1, which was recently suggested to participate in anxiety-like behaviors (Kasckow et al, 2004; Marsch et al, 2007). Indeed, Rubino et al (2008) found that direct injection of URB597 in the prefrontal cortex reduces anxiety-like behaviors in rats at low doses, and exacerbates them at high doses, the latter effect being blocked by a TRPV1 antagonist. The authors also showed that the prototypical TRPV1 agonist, capsaicin, causes anxiogenic effects. These findings are not surprising in view of increasing evidence supporting the function of TRPV1 receptors in stimulating glutamate release in various brain areas (Starowicz et al, 2008).
On the basis of this background we reasoned that compounds with the capability of blocking both FAAH and TRPV1 should produce more efficacious anxiolytic effects in rodents than selective FAAH inhibitors. One such dual FAAH/TRPV1 blocker is N-arachidonoyl-serotonin (AA-5-HT) (Bisogno et al, 1998; Maione et al, 2007), which exerts antihyperalgesic effects by inactivating both proteins (Maione et al, 2007; Ortar et al, 2007). As strain differences, possibly due to neuroanatomical, neurochemical, or genetic factors, exist in mice for their sensitivity to antidepressant or anxiolytic compounds (Crawley et al, 1997; Griebel et al, 2000; Lepicard et al, 2000; Leggio et al, 2008), we investigated here the potential anxiolytic effect of AA-5-HT (and, as a comparison, of diazepam) in both C57BL/6J and Swiss mice, using the EPM, a well-validated test to search for new anxiolytic agents (Pellow et al, 1985; File, 1992). Finally, to investigate the neuroanatomical relationships between CB1 and TRPV1 in brain areas controlling emotionality (prefrontal cortex, nucleus accumbens (Acb), amygdala, and hippocampus), we performed immunohistochemical studies on both Swiss and C57BL/6J mouse brain.
MATERIALS AND METHODS
Animals
Male Swiss and male C57BL/6J mice (obtained from Charles River, Italy) aged 7–8 weeks were used throughout the behavioral and pharmacological experiments. Two male adult Swiss mice, two male adult wild-type C57BL/6J mice (WT) and two male adult TRPV1-deficient mice (TRPV1-KO) with the C57BL/6J background obtained from Jackson ImmunoResearch Laboratories (Bar Harbor, ME, USA) were used for the immunohistochemical experiments. For at least 1 week prior each experiment, animals were housed five to a cage at a constant temperature of 21°C, and under a 12-h light/dark cycle (lights on between 0800 and 2000 hours), with food and water available ad libitum. Animals were randomly assigned to each treatment group and used only once in the behavioral experiments and then were killed at the end of behavioral procedures. All behavioral tests took place in an experimental room with the same light/dark cycle and the environmental conditions, such as humidity and temperature levels inside the room, similar to those of the housing facility. All experiments were carried out according to the European Community Council Directive 86/609/EEC and efforts were made to minimize animal suffering and to reduce the number of animals used. The rationale, design, and methods of this study have been approved by the Ethical Committee for Animal Research, University of Catania.
Behavioral Tests
Elevated-plus-maze test
The apparatus consisted of two opposite open arms (30 × 5 cm) and two arms with walls (30 × 5 × 14 cm) that were attached to a central platform (5 × 5 cm) to form a cross. The maze was elevated 50 cm from the floor (Pellow et al, 1985). Illumination (∼300 lx) was provided at the center of the maze. After treatment, each animal was placed at the center of the maze with its nose in the direction of one of the closed arms, and observed for 5 min, according to the following parameters: number of entries in the open and closed arms, and time of permanence in each of them (ie the time spent by the animal in the open and closed arms). An entry was defined as all four paws having crossed the line between an arm and the central area. It is accepted that the anxiolytic effect of a drug treatment is illustrated by increased parameters in open arms (time and/or number of entries), although entries in closed arms and total entries reflect the motor component of the exploratory activity. From these values, both the percentage of time and of entries in open arms provided as the measures of anxiety-like behavior for each animal. On removal of each mouse, the maze floor was carefully wiped with a wet towel. The behavior of all animals was recorded on a tape using a video camera (Hitachi Videocam) and then scored in monitor display by two ‘blinded’ observers.
Drugs and Experimental Design
Diazepam, the CB1 receptor antagonist AM251 (N-(piperidin-1-yl)-5-(4-iodophenyl)-1-(2,4-dichlorophenyl)-4-methyl-1H-pyrazole-3-carboxamide), the TRPV1 receptor agonist olvanil ((N-vanillyl)-9-oleamide), the TRPV1 receptor antagonist SB366791 (N-(3-Methoxyphenyl)-4-chlorocinnamide), and the selective FAAH inhibitor URB597 (cyclohexylcarbamic acid 3′-carbamoyl-biphenyl-3-yl ester) were purchased from Sigma (St Louis, MO, USA). AA-5-HT was synthesized in Dr Di Marzo's laboratory, as previously described (Bisogno et al, 1998; De Petrocellis et al, 2000). All compounds were dissolved in saline containing 10% dimethyl sulfoxide and injected intraperitoneally (i.p.) in a volume of 10 ml/kg. Control animals i.p. received the same volume of vehicle-injected controls (VHC). Two different experiments were programmed and carried out.
In the first set of experiments, different groups of C57BL/6J mice (n=8–10) were treated acutely with AA-5-HT (0.1, 0.5, 1.0, 2.5, or 5 mg/kg), diazepam (1 mg/kg), URB597 (0.1, 0.5, or 1 mg/kg), or VHC. Animals received an i.p. injection of drug 30 min prior to the behavioral testing. To assess the involvement of CB1 or TRPV1 receptors in the behavioral effect of AA-5-HT, different groups of mice (n=8–10) were treated acutely with previously determined per se inactive doses of olvanil (0.1 mg/kg) or AM251 (1 mg/kg), alone or combined with AA-5-HT, at the dose of 0.1 mg/kg. Both a dose–effect relationship of SB366791 (0.1, 0.5, 1, or 2.5 mg/kg) and its synergistic effect with AA-5-HT (0.1 mg/kg) were also evaluated. When AA-5-HT was given in combination with olvanil, AM251, or SB366791, these compounds were injected 5 min before AA-5-HT, as described by Maione et al (2007).
In the second set of experiments, different groups of Swiss mice (n=10) were treated with AA-5-HT (1.0, 2.5, or 5 mg/kg), diazepam (1 mg/kg), or VHC, either acutely or subchronically (every other day for 7 days) to determine the dose–effect relationship of AA-5-HT in EPM test. As no effect was observed, except for an anxiogenic-like action accompanied by hypolocomotion at the highest dose, the experiments were repeated using a chronic, daily administration of AA-5-HT as described below. Different groups of Swiss mice (n=10) received one injection i.p. daily for 7 days of either AA-5-HT (1, 2.5, or 5 mg/kg per day), diazepam (1 mg/kg per day), URB597 (0.1, 0.5, or 1 mg/kg per day), or VHC. To study the role of CB1 or TRPV1 receptors in the effect of AA-5-HT, different groups of mice (n=10) were treated i.p. for 7 days with previously determined per se inactive doses of AM251 (1 mg/kg per day) or olvanil (0.1 mg/kg per day), alone or combined with AA-5-HT, at the dose of 2.5 mg/kg per day, which had produced anxiolytic-like effects in the dose–response experiment. Furthermore, different group of animals were treated with SB366791 (0.5, 1, or 2.5 mg/kg per day), alone, or combined with AA-5-HT (1 or 5 mg/kg per day), to assess both a dose–effect relationship of the compound and a synergistic effect with AA-5-HT in EPM test, respectively. When AA-5-HT was given in combination with olvanil, AM251 or SB366791, these compounds were injected 5 min before AA-5-HT as described by Maione et al (2007). On day 7, the last injection of AA-5-HT was made 30 min prior to the behavioral testing.
Diazepam was selected as the active control. The doses of the other compounds were selected based on the results of previous reports (De Petrocellis et al, 2000; Kasckow et al, 2004; Patel and Hillard, 2006; Consoli et al, 2007; Naidu et al, 2007; Moreira et al, 2008).
Measurement of Endocannabinoids
Each brain (N=4 for each experimental group, ∼400 mg wet tissue weight for C57BL/6J mice and ∼500 mg wet tissue weight for Swiss mice) was homogenized in five volumes of chloroform/methanol/Tris-HCl 50 mM (2 : 1 : 1) containing 50 pmol each of d8-anandamide and d5-2-AG. Deuterated standards were purchased from Cayman Chemicals (Ann Arbor, MI, USA). Homogenates were centrifuged at 13 000g for 16 min (4°C), the aqueous phase plus debris were collected and extracted again twice with one volume of chloroform. The organic phases from the three extractions were pooled and the organic solvents evaporated in a rotating evaporator. Lyophilized samples were then stored frozen at −80°C under nitrogen atmosphere until analysis. Extracts were resuspended in chloroform/methanol at a ratio of 99 : 1 by volume. The solutions were then purified by open-bed chromatography on silica as described in Maione et al (2006). Fractions eluted with chloroform/methanol at a ratio of 9 : 1 by volume (containing anandamide and 2-AG) were collected and the excess solvent was evaporated with a rotating evaporator, and aliquots analyzed by isotope dilution liquid chromatography/atmospheric pressure chemical ionization/mass spectrometry (LC-APCI-MS) carried out under conditions previously described (Maione et al, 2006) and allowing the separations of 2-AG and anandamide. MS detection was carried out in the selected ion-monitoring mode using m/z values of 356 and 348 (molecular ion +1 for deuterated and undeuterated anandamide), and 384.35 and 379.35 (molecular ion +1 for deuterated and undeuterated 2-AG). The area ratios between signals of deuterated and undeuterated compounds varied linearly with varying amounts of undeuterated compounds. Anandamide and 2-AG levels in unknown samples were therefore calculated on the basis of their area ratios with the internal deuterated standard signal areas.
Immunohistochemical Staining of CB1 and TRPV1 Receptors
Mice were deeply anaesthetized (pentobarbital, 60 mg/kg, i.p.) and perfused transcardially with saline followed by 4% paraformaldehyde in 0.1 M phosphate buffer (PB), pH 7.4. Each brain was removed from the skull, fixated for 2 h and then washed, and soaked for cryoprotection in 30% sucrose in PB at 4°C until it sank. Serial coronal frozen sections were cut through the brain at 12 μm-thick and collected on gelatine-coated slides (Mezel, Germany) in three alternate series: the first and second processed respectively for anti-TRPV1 and anti-CB1 immunohistochemistry using the ABC method, dehydrated, cleared in xylene, and coverslipped with DPX; the third series of sections was processed for CB1/TRPV1 double immunofluorescence, dried, and coverslipped with Vecthashield mounting medium (Vector Laboratories, Burlingame, CA).
Single Immunohistochemistry
For single anti-TRPV1 and anti-CB1 immunohistochemistry, the sections were reacted for 10 min in 0.1% H2O2 to inactivate endogenous peroxidase activity and preincubated for 60 min at room temperature respectively in 10% of normal rabbit serum (NRS; Dako, Glostrup, Denmark) and of normal goat serum (NGS; Vector Laboratories) in 0.1 M Tris-buffered saline, pH 7.6 (TBS), containing 0.3% Triton X-100 and 0.05% sodium azide (Sigma). The sections were then incubated for 2 days at 4°C with NRS-diluted (1 : 200) N-terminal goat polyclonal anti-TRPV1 receptor (Santa Cruz Biotechnology, Santa Cruz, CA) or with NGS-diluted (1 : 1000) rabbit polyclonal anti-CB1 (Calbiochem, EMD Biosiences Inc., La Jolla, USA and Canada). After several rinses, the sections were incubated at room temperature for 2 h in biotinylated rabbit anti-goat or biotinylated goat anti-rabbit IgGs (Vector Laboratories) respectively for TRPV1 or CB1 immunohistochemistry followed by incubation for 1 h in the avidin-biotin complex (ABC Kit; Vectastain, Vector) diluted in TBS according to the supplier indications, and then in 0.05‰ 3-3′diaminobenzidine for 10 min (DAB Sigma Fast; Sigma-Aldrich).
Multiple Labeling with Immunofluorescence
After preincubation for 1 h in 10% normal donkey serum (NDS Jackson Immunoresearch Laboratories, West Grove, PA, USA) in PB, pH 7.4 containing 0.3% Triton X-100 and 0.05% sodium azide (Sigma), the sections were incubated for 2 days in a mixture of anti-CB1 receptor coupled to anti-TRPV1 receptor (diluted at a ratio of 1 : 100 in NDS respectively) at +4°C. TRPV1/CB1 double immunofluorescence was revealed by incubation for 4 h in a mixture of Alexa546 donkey anti-rabbit IgGs and Alexa488 donkey anti-goat IgGs (Molecular Probes Inc., Eugene, OR) diluted 1 : 50 in NDS. Controls of single and double immunoreactivity (ir) included (1) preabsorption of diluted antibodies with their respective immunizing peptides, except for CB1 receptor antibody whose specificity was confirmed in CB1-null mouse brain (see Cristino et al, 2006 for details) and (2) omission of either the primary antisera or the secondary antibodies. For double immnunofluorescence only, to make sure that there was no cross-reaction between the secondary and primary antibodies, incubation of the section with the first primary antibody and the second secondary antibody was performed. No immunostaining was detected in any of the controls. The sections processed for immunofluorescence were studied with an epifluorescence microscope equipped with the appropriate filters (Zeiss Axioscop); all the other material was investigated at the microscope under bright-field illumination. Images were acquired by using the digital camera JVC TK-1381 connected to the microscope and the image analysis software AxioVision Plus 4.0 for Windows (Carl Zeiss; Germany). Plates were generated adjusting the contrast and brightness of digital images (Corel Draw, 9.0).
Statistical Analysis
Behavioral data were analyzed using one-way ANOVA followed by post hoc Dunnett's test for multiple comparisons. Analytical data were analyzed using one-way ANOVA followed by post hoc Bonferroni's test. A p-value of 0.05 or less was considered as indicative of a significant difference.
RESULTS
Effects of AA-5-HT in C57BL/6J Mice
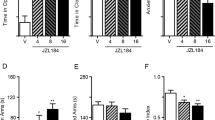
In C57BL/6J mice, both AA-5-HT and URB597 exhibited effects in the EPM already with acute i.p. treatment (Figure 1a). One-way ANOVA analysis revealed a main effect of drugs for percentage of time (F9,76=14.6; p<0.001) and entries (F9,76=10.13; p<0.001) in open arms compared to VHC-injected controls. Post hoc analysis revealed that already at the lowest dose tested (0.1 mg/kg), AA-5-HT significantly enhanced the percentage of time and entries in open arms (p<0.05). The effect of AA-5-HT (1 mg/kg) was similar to that induced by diazepam (1 mg/kg) (p<0.001), it decreased at 2.5 mg/kg and was not appreciable at the highest dose tested of 5 mg/kg, which induced a decrease of total arm entries, an index of reduced locomotor behavior (F9,76=4.5; p<0.001). Under these experimental conditions, URB597 exerted anxiolytic effects only at highest dose tested, as assessed by the significant increase of percentage of time (p<0.01) and entries in open arms (p<0.05). To test whether CB1 or TRPV1 receptors were involved in AA-5-HT modulation of anxiety behavior, we pretreated mice with AM251 or olvanil, a selective CB1 receptors antagonist and a selective TRPV1 agonist, respectively. Figure 1b shows that the anxiolytic effect of AA-5-HT (0.1 mg/kg) was not only abolished by a per se inactive dose of AM251 (1 mg/kg), but was also transformed into an anxiogenic effect, as a significant reduction of percentage of time in open arms was induced (p<0.05). An inactive per se dose of olvanil (De Petrocellis et al, 2000) (0.1 mg/kg) instead simply abolished the anxiolytic effect of AA-5-HT (0.1 mg/kg). The TRPV1 antagonist SB366791 significantly increased the percentage of time (p<0.01) and entries (p<0.05) into the open arm, thus showing an anxiolytic-like effect, only at the dose of 1 mg/kg. The highest dose tested of SB366791 (2.5 mg/kg) decreased the total arm entries, a index of impaired locomotor behavior (p<0.001). Co-administration of the highest inactive dose of SB366791 (0.5 mg/kg) with AA-5-HT (0.1 mg/kg) did not enhance the effect of the latter, but instead decreased the total arm entries, again possibly due to impaired locomotor behavior (p<0.001).

(a) Effects of N-arachidonoyl-serotonin (AA-5-HT) (0.1, 0.5, 1, 2.5, or 5 mg/kg), URB597 (0.1, 0.5, or 1 mg/kg), and diazepam (1 mg/kg) in C57BL/6J mice tested in the elevated plus maze (EPM) test. The test was performed 30 min after intraperitoneal (i.p.) administration of the drugs. Data are presented as means±SEM for the percentage of time spent in open arms (upper panel), the percentage of entries in open arms (middle panel), and the number of total arm entries (lower panel) from 8 to 10 mice. *Significantly different compared to vehicle-injected controls (VHCs) (p<0.05; Dunnett's test for multiple comparisons). **Significantly different compared to VHCs (p<0.01; Dunnett's test for multiple comparisons). ***Significantly different compared to VHCs (p<0.001; Dunnett's test for multiple comparisons). (b) Effects of olvanil (0.1 mg/kg), AM251 (1 mg/kg), or SB366791 (0.1, 0.5, or 1 mg/kg) alone or combined with AA-5-HT (0.1 mg/kg) in C57BL/6J mice tested in the EPM test. The test was performed 30 min after i.p. administration of the drugs. Olvanil (0.1 mg/kg), AM251 (1 mg/kg), or SB366791 (0.5 mg/kg) were injected i.p. 5 min before AA-5-HT (0.1 mg/kg). Data are presented as means±SEM for the percentage of time spent in open arms (upper panel), the percentage of entries in open arms (middle panel), and the number of total arm entries (lower panel) from 8 to 10 mice. *Significantly different compared to VHCs (p<0.05; Dunnett's test for multiple comparisons). **Significantly different compared to VHCs (p<0.01; Dunnett's test for multiple comparisons). ***Significantly different compared to VHCs (p<0.001; Dunnett's test for multiple comparisons).
Effects of AA-5-HT in Swiss Mice
In Swiss mice, either acute or subchronic (every other day for 7 days) treatment with AA-5-HT (1.0, 2.5, or 5 mg/kg, i.p.) did not exert any effect on anxiety-like behavior, except for an anxiogenic action at the highest dose, revealed by a reduction of percentage of time (F4,45=23.68; p<0.01) and entries in open arms (F4,45=18.64; p<0.01), with a significant reduction of total arm entries (F4,45=9.29; p<0.001) (Figure 2; data not shown). By contrast, diazepam (1 mg/kg) showed an anxiolytic-like effect under these conditions (p<0.001). For these reasons, we used a chronic daily treatment (7 days) in this mouse strain to evaluate the anxiety-like behavior of AA-5-HT in the EPM. For the dose–response relationship, one-way ANOVA analysis revealed a main effect of drugs for percentage of time (F7,72=8.5; p<0.0001) and entries (F7,72=9.2; p<0.0001) in open arms compared to VHC-injected controls. Post hoc analysis revealed that AA-5-HT exerted no effect at 1.0 mg/kg per day, an anxiolytic-like effect at 2.5 mg/kg per day in terms of a significant increase of percentage of time (p<0.01) and entries in open arms (p<0.01), slightly lower than that exerted by diazepam 1 mg/kg per day (p<0.001), and a reduction of percentage of time in open arms at 5 mg/kg per day (p<0.05). However, AA-5-HT (5 mg/kg per day) also induced a significant decrease of total arm entries (F7,72=5.5; p<0.001). URB597 (0.1–1 mg/kg per day) also exerted an anxiolytic effect, but only at the highest dose tested (p<0.05) (Figure 3a). Again, the anxiolytic effect of AA-5-HT (2.5 mg/kg per day) was transformed into an anxiogenic effect, in terms of significant decrease of percentage of time (p<0.01) and entries in open arms (p<0.05), by a per se inactive dose of AM251 (1 mg/kg per day), and was antagonized by olvanil (0.1 mg/kg per day). SB366791 (0.5 mg/kg per day), although inactive per se in this mouse strain augmented the effect of an inactive dose of AA-5-HT (1 mg/kg per day) by increasing significantly both the percentage of time and entries in open arms (p<0.001). On the other hand, SB366791 did not antagonize the anxiogenic-like effect of AA-5-HT (5 mg/kg per day), thus suggesting that this effect is not due to activation of TRPV1 (Figure 3b).

Effects of acute administration of N-arachidonoyl-serotonin (AA-5-HT) (1, 2.5, or 5 mg/kg) and diazepam (1 mg/kg) in Swiss mice tested in the elevated plus maze (EPM) test. The test was performed 30 m after the intraperitoneal (i.p.) administration of the drug. Data are presented as means±SEM for the percentage of time spent in open arms (upper panel), the percentage of entries in open arms (middle panel), and the number of total arm entries (lower panel) from 10 mice. **Significantly different compared to vehicle-injected controls (VHCs) (p<0.01; Dunnett's test for multiple comparisons). ***Significantly different compared to VHCs (p<0.001; Dunnett's test for multiple comparisons).

(a) Effects of repeated administration (7 days, daily) of N-arachidonoyl-serotonin (AA-5-HT) (1, 2.5, or 5 mg/kg), URB597 (0.1, 0.5, or 1 mg/kg), and diazepam (1 mg/kg) in Swiss mice tested in the elevated plus maze (EPM) test. On day 7, the test was performed 30 min after the intraperitoneal (i.p.) administration of the drugs. Data are presented as means±SEM for the percentage of time spent in open arms (upper panel), the percentage of entries in open arms (middle panel), and the number of total arm entries (lower panel) from 10 mice. *Significantly different compared to vehicle-injected controls (VHCs) (p<0.05; Dunnett's test for multiple comparisons). **Significantly different compared to VHCs (p<0.01; Dunnett's test for multiple comparisons). ***Significantly different compared to VHCs (p<0.001; Dunnett's test for multiple comparisons). (b) Effects of repeated treatment (7 days, daily) of olvanil (0.1 mg/kg per day), AM251 (1 mg/kg per day), or SB366791 (0.5, 1, or 2.5 mg/kg per day) alone or combined with AA-5-HT (1, 2.5, or 5 mg/kg per day) in Swiss mice tested in the EPM test. The test was performed 30 m after the i.p. administration of the drugs. Olvanil (0.1 mg/kg per day), AM251 (1 mg/kg per day), or SB366791 (0.5 mg/kg per day) were injected i.p. 5 min before AA-5-HT (1, 2.5, or 5 mg/kg per day). Data are presented as means±SEM for the percentage of time spent in open arms (upper panel), the percentage of entries in open arms (middle panel), and the number of total arm entries (lower panel) from 10 mice. *Significantly different compared to VHCs (p<0.05; Dunnett's test for multiple comparisons). **Significantly different compared to VHCs (p<0.01; Dunnett's test for multiple comparisons). ***Significantly different compared to VHCs (p<0.001; Dunnett's test for multiple comparisons).
Effect of AA-5-HT on Endocannabinoid Levels
Brains from mice submitted to either acute of chronic treatment with AA-5-HT, diazepam, or VHC were analyzed for their levels of anandamide and 2-AG. In C57BL/6J mice, acute administration of 0.1, 1.0, 2.5, but not 5.0, mg/kg of AA-5-HT caused elevation of both endocannabinoids (Figure 4a). The stimulatory effect of AA-5-HT on 2-AG levels is not unprecedented for FAAH inhibitors (Di Marzo and Maccarrone, 2008), and is unlikely due to inhibition of monoacylglycerol lipase (MAGL), the main enzyme deputed to 2-AG degradation, as the inhibitor, which exhibits an IC50 vs FAAH of 1–10 μM (Bisogno et al, 1998; Fowler et al, 2003), is inactive at inhibiting MAGL up to a 25 μM concentration (Bisogno et al, 2006), and is selective vs the other 2-AG hydrolyzing enzymes recently reported in the mouse brain (Blankman et al, 2007) (Professor B Cravatt, personal communication to V Di Marzo). In Swiss mice, chronic, but not acute, administration of 2.5 mg/kg, but not 1.0 or 5.0 mg/kg of AA-5-HT caused elevation of anandamide, but not 2-AG, levels (Figure 4b; data not shown). In C57BL/6J mice, acute administration of diazepam elevated both anandamide and 2-AG levels. In Swiss mice, acute or chronic administration of diazepam elevated only 2-AG or anandamide levels, respectively (Figure 4b; data not shown).

(a) Effect of acute intraperitoneal (i.p.) administration of N-arachidonoyl-serotonin (AA-5-HT) (0.1, 1, 2.5, or 5 mg/kg) or diazepam (1 mg/kg) in C57BL/6J mice on brain anandamide and 2-arachidonoyl-glycerol (2-AG) levels. (b) Effect of chronic (7 days, daily) i.p. administration of AA-5-HT (1, 2.5, or 5 mg/kg) or diazepam (1 mg/kg) in Swiss mice on brain anandamide and 2-AG levels. Data are means±SEM of N=4 different determinations. Means were compared by ANOVA followed by the Bonferroni's test. *P<0.05; **P<0.01; ***P<0.005.
Immunohistochemical Staining of TRPV1 Receptors in Brain Areas Involved in Affective Disorders
General considerations
A medium to large degree of colocalization between TRPV1 and CB1 receptors was found in all of the brain areas involved in affective disorders, and more strongly in the prefrontal cortex and the shell of nucleus Acb (Figure 5). Similar immunohistochemical TRPV1 staining was found in Swiss and C57BL/6J mouse strains (Figures 5 and 6). Finally, similar to previous results in the ABH TRPV1−/− mouse strain (see Cristino et al, 2006 for details), specificity of TRPV1-ir was confirmed by the absence of signals in the TRPV1−/− mice.

Immunofluorescence showing the reciprocal distribution of transient receptor potential vanilloid type-1 (TRPV1) and cannabinoid receptors of type-1 (CB1) receptors in some brain areas participating in anxiety areas of Swiss mice. (a–a2) Prefrontal cortex (PrFCx) with intense single (a, a1) and double (a2) TRPV1 and CB1 receptors immunoexpression mostly restricted to layers II/III and V. Note the presynaptic CB1-ir as thin processes around the TRPV1-ir somata (a2). (b–c2) Nucleus accumbens. Some TRPV1-ir neuronal somata (b) receive CB1-ir nervous terminals (b1) in the accumbens core (AcbC) (b2), whereas a complete overlap between TRPV1-ir and CB1-ir exists at the soma level in the accumbens shell (AcbSh) subnucleus (c–c2). (d–e2) Amygdala. TRPV1-ir is present at the somatodendritic level in both putative pyramidal projection neurons and nonprojection neurons of the amygdalopyriform transition area (APir), posteromedial part of amygdalohippocampal area (AHiPM), and posteromedial cortical amygdaloid nucleus (PMCo) (d, e). Single CB1-ir was almost exclusively located in the PMCo area, being expressed as a very light immunopositive meshwork in the APir and AHiPM subnuclei (d1, e1). Fully overlapped TRPV1/CB1 coexpression was intense exclusively in the pyramidal neurons of PMCo subnucleus (d2, e2). Aca, anterior commissure. Scale bar: a–b2=50 μm; c–e2=100 μm.

Transient receptor potential vanilloid type-1 (TRPV1) single immunohistochemistry showing the distribution of TRPV1 in some brain areas of C57BL/6J mice controlling emotionality. (a, b). Prefrontal cortex (PrFCx). Single TRPV1-ir in wild-type C57BL/6J mice confirmed the layer-specific distribution and the almost no presence in the layer VI observed in the PrFCx of the Swiss strain (Figure 5) (a). No TRPV1-ir signal was obtained in the PrFCx from TRPV1-KO mice (b). (c, d). Nucleus accumbens (Acb). Single TRPV1-ir in wild-type C57BL/6J mice confirmed the receptor distribution in several neuronal cell bodies of the core (c, accumbens core (AcbC)) and shell (d, AcbSh) subnuclei observed in the Acb of the Swiss strain (Figure 5). (e, f). Amygdala. Somatodendritic single TRPV1-ir was prominent in many neurons located in the amygdalopyriform transition area (Apir), amygdalohippocampal area (AHiPM), and posteromedial cortical amygdaloid nucleus (PMCo) areas of the wild-type C57BL/6J mouse brain (e) whereas no immunostaining was observed in the same amygdaloid regions of TRPV1-KO brain (f). Aca, anterior commissure. Scale bar: a–b=50 μm; c–f=100 μm.
Prefrontal cortex
Both TRPV1 and CB1 receptor expression patterns were layer specific, and double immunofluorescence was mostly restricted to layers II/III and V (Figure 5a–a2). Both dotted and thin process-like elements of CB1-ir took up the neuropil of the labeled layers where they intermingled with TRPV1-ir somata, in particular in the layer V synapses of the prelimbic area of the prefrontal cortex. Single TRPV1-ir on TRPV1 littermate C57BL/6J mouse strains (Figure 6a) confirmed the immunofluorescence findings in the Swiss strain, whereas no TRPV1-ir signal was found in the prefrontal cortex of TRPV1-KO mice (Figure 6b).
Nucleus accumbens
Among the two anatomically and functionally distinct compartments of the nucleus Acb a lower degree of TRPV1/CB1-ir colocalization was found in the core (Figure 5b–b2; AcbC) where only a few CB1-ir terminals were apposed on TRPV1-ir soma compared to the complete overlapping of TRPV1/CB1-ir in the neuronal cell bodies of the medial shell (Figure 5c–c2; AcbSh). Single moderate CB1-ir fibers were also present in the medial shell. These data were confirmed by single TRPV1-ir observations in wild-type C57BL/6J mice (Figure 6c), whereas no TRPV1-ir was observed in the nucleus Acb of TRPV1 KO mice (Figure 6d).
Amygdala
The amygdala was densely labeled for the TRPV1 receptor. Somatodendritic immunostaining was observed in both putative pyramidal and projection neurons (with a single prominent dendrite or several large dendrites) and in the putative nonprojection neurons located in the amygdalopyriform transition area (APir), posteromedial part of amygdalohippocampal area (AHiPM), and posteromedial cortical amygdaloid nucleus (PMCo) (Figures 5d, e). Single CB1-ir was almost exclusively located in the PMCo area, being expressed as a very light immunopositive meshwork in the APir and AHiPM subnuclei (Figures 5d1, e1). Therefore TRPV1/CB1 coexpression was intense and fully overlapped in the pyramidal neurons of PMCo subnucleus only (Figures 5d2, e2). The immunofluorescence localization of TRPV1-ir distribution in the amygdaloid Apir, PMCo, and AHiPM subnuclei of Swiss strain was confirmed by single TRPV1-ir in wild-type C57BL/6J mice (Figure 6e), whereas no TRPV1-ir signal was found in the amygdala of TRPV1 KO mice (Figure 6f).
Hippocampal formation
In agreement with previous data (Cristino et al, 2006), complementary localization of CB1/TRPV1-ir was observed throughout the CA1-CA3 subfields of the Ammon's horn (data not shown). At the cellular level, TRPV1-ir was found on cell bodies and basal dendrites of many pyramidal neurons surrounded by a dense plexus of CB1-ir terminals.
DISCUSSION
We have provided evidence that cannabinoid CB1 receptors and vanilloid TRPV1 receptors can be simultaneously indirectly activated and antagonized, respectively, to produce anxiolytic-like actions in mice. Several findings support the role of the endocannabinoid system in the control of anxiety-like behaviors in rodents and of anxiety in humans. CB1 receptor agonists and antagonists usually exert anxiolytic- and anxiogenic-like effects, respectively, in rodents, although discrepant results might be observed under different experimental conditions (Lafenetre et al, 2007; Viveros et al, 2007; Bisogno and Di Marzo, 2007). In obese humans chronically treated with the CB1 receptor antagonist rimonabant (20 mg per day), anxiety is the cause of discontinuation in 1% of the rimonabant-treated vs 0.3% of the placebo-treated patients (Van Gaal et al, 2008). These findings suggest that, during stressful conditions leading to anxiety-like behaviors in mammals, endocannabinoids are produced and released in brain areas controlling emotional reactions (Marsicano et al, 2002; Hohmann and Suplita, 2006), to decrease fear and facilitate the adaptation to new environmental conditions by CB1 activation (Steiner et al, 2008; Lutz, 2007). Accordingly, inhibition of FAAH, which can prolong the lifespan of anandamide only where it is produced and degraded (Di Marzo et al, 2008), generally causes anxiolytic effects (Kathuria et al, 2003; Patel and Hillard, 2006; Rubino et al, 2008; Moreira et al, 2008). However, FAAH inhibition, by elevating the tissue levels of anandamide and oleoylethanolamide, might also indirectly activate TRPV1 receptors, as observed in animal models of pain, hyperdopaminergia, and neurodegenerative disorders (Maione et al, 2006; Tzavara et al, 2006; Lee et al, 2006; Morgese et al, 2007). As TRPV1 activation might cause effects opposite to those of CB1 activation in some CNS disorders (Morgese et al, 2007), including anxiety (Kasckow et al, 2004; Marsch et al, 2007), this phenomenon might reduce the efficacy of FAAH inhibitors (Naidu et al, 2007; Rubino et al, 2008). Moreover, it was shown that FAAH inhibition in the striatum, by elevating anandamide levels and TRPV1 activity, indirectly inhibits the biosynthesis of the other endocannabinoid, 2-AG (Maccarrone et al, 2008). Thus, FAAH inhibitors might at the same time elevate and inhibit CB1 receptor tone.
For these reasons, we hypothesized that FAAH inhibitors with intrinsic TRPV1 antagonist activity, like AA-5-HT (Maione et al, 2007), might be more efficacious against anxiety than selective FAAH inhibitors like URB597. AA-5-HT was shown previously to elevate brain endocannabinoid levels without affecting serotonin levels when administered i.p. up to a concentration of 5 mg/kg, and therefore was deemed suitable to be tested on anxiety-like behaviors (de Lago et al, 2005). We report here that, in C57BL/6J mice submitted to the EPM test, AA-5-HT is more potent that URB597 at reducing anxiety-like behavior, and that its effect is counteracted not only by a CB1 antagonist, but also by the selective TRPV1 agonist, olvanil, at doses inactive per se. Importantly, doses of olvanil higher than 1 mg/kg were previously found to cause anxiety-like behavior in rats in the EPM (Kasckow et al, 2004). The selective TRPV1 antagonist, SB366791, was also found here to be less efficacious than AA-5-HT at exerting an anxiolytic-like action in C57BL/6J mice. When the EPM test was carried out in Swiss mice, AA-5-HT was less potent and efficacious than in C57BL/6J mice and had to be administered daily for a week to observe effects on anxiety-like behaviors. However, also in this case, the anxiolytic-like effect of AA-5-HT was counteracted not only by a CB1 antagonist, but also by olvanil. In agreement with the role of TRPV1 receptor blockade in the antianxiety effects of AA-5-HT are also the findings with per se inactive doses of SB366791, which either significantly augmented, or occluded, the effects of doses of AA-5-HT that were either inactive or anxiolytic per se, ie chronic 1 mg/kg in Swiss mice and acute 0.1 mg/kg in C57BL/6J mice, respectively. Unlike C57BL/6J mice, in Swiss mice SB366791 failed to produce anxiolytic effects per se, hinting at a possible lower efficacy of TRPV1 antagonism against anxiety-like behaviors in this mouse strain.
In agreement with the role of both FAAH and TRPV1 blockade in the effects of AA-5-HT on anxiety-like behaviors, we found here that treatment of mice with anxiolytic doses of this compound was always accompanied by elevation of brain anandamide levels. In the case of acute treatments in C57BL/6J mice, we observed also an elevation of 2-AG levels, which might explain in part why, in this strain, AA-5-HT was more potent and efficacious after acute treatments than in Swiss mice. However, AA-5-HT appeared to be more effective also at elevating brain anandamide levels in this strain compared to Swiss mice.
An intriguing finding of our experiments in the EPM is that the CB1 receptor antagonist AM251, at a dose inactive per se, not only abolished the anxiolytic action of AA-5-HT, but it also transformed it into an anxiogenic-like effect. This suggests that, when CB1 receptors are blocked, the dose of AA-5-HT used (0.1 mg/kg), while still elevating anandamide levels, might not be sufficient any longer to counteract TRPV1 receptors, possibly because these are otherwise tonically inhibited in part by CB1 receptors. In fact, there is evidence that CB1 receptor activation counteracts TRPV1 activation, particularly when the two receptors are coexpressed in the same cell (Ahluwalia et al, 2003; Hermann et al, 2003).
Indeed, we did observe here that TRPV1 receptors are often coexpressed with CB1 receptors in those brain areas that are known to control affective behaviors, such as: (1) the II/III and V layers of the PrFC, which receive projections from the prelimbic area; (2) the shell of nucleus Acb, which receives projections from the prefrontal cortex and other subcortical limbic brain regions; (3) the amygdaloid formations, of which the PMCo subnucleus shows the highest level of TRPV1/CB1 colocalization; and, (4) the CA1-CA3 regions of the hippocampus. These anatomical findings suggest that if compounds that are capable of both antagonizing TRPV1 and indirectly activating CB1 (by inhibiting FAAH) are delivered to these brain nuclei, they can potentially affect anxiety-like responses by modulating simultaneously the activity of both receptor types.
We observed that the highest dose of AA-5-HT was either ineffective, in the case of acute treatments in C57BL/6J mice, or produced anxiogenic effects, in the case of the chronic treatment of Swiss mice. A similar anxiogenic action of a high dose of URB597, administered in the prefrontal cortex, was recently suggested to be due to indirect activation of TRPV1 receptors as it was blocked by a TRPV1 antagonist (Rubino et al, 2008). In the case of AA-5-HT, however, the anxiogenic effects of the highest dose was not significantly modified by SB366791 at a dose inactive per se but still capable to augment the effect of a low dose of AA-5-HT. It is possible that high chronic doses of this compound inhibit motor behavior, as evidenced by its inhibition of total arm entries in Swiss mice. The lack of anxiolytic-like effects of the high dose of AA-5-HT in both strains, instead, might be due to the fact that this dose caused no effects on brain anandamide levels, thus suggesting that TRPV1 antagonism alone might not be sufficient to reduce anxiety. However, this might be true in Swiss mice, where selective blockade of TRPV1 with SB366791 was always insufficient to reduce anxiety-like behaviors, but not in C57BL/6J mice, in which SB366791 was efficacious at an intermediate dose. We have no clear-cut explanation as to why the acute or chronic administration of the 5 mg/kg dose of AA-5-HT was ineffective at elevating brain anandamide levels. In the only previous study in which this was tested in mice, we found no effect of 5 mg/kg AA-5-HT on anandamide levels after 3 days of subchronic (every other day) i.p. administration, although subchronic administration for 7 days elevated both anandamide and 2-AG brain levels (van der Stelt et al, 2006). It is possible that, like their actions on anxiety-like behaviors (present study and Moreira et al, 2008; Naidu et al, 2007), the effects of FAAH inhibitors on brain endocannabinoid levels are species-, strain-, treatment-, and dose specific.
A recent study showed that the anxiolytic-like effect of diazepam in the EPM test could be attenuated by the CB1 antagonist AM251, whereas a sub-effective dose of diazepam significantly augmented the effects of a sub-effective dose of URB597 (Naderi et al, 2008). Our present finding of elevated brain endocannabinoid levels following either acute or chronic treatment with diazepam might explain these intriguing previous data. In fact, it is possible that diazepam exerts its anxiolytic action in part by elevating endocannabinoid levels, thereby indirectly activating CB1 receptors. To confirm this hypothesis a biochemical mechanism for this phenomenon will have to be found, bearing in mind the GABA-enhancing and neuron hyperpolarizing properties of benzodiazepines. In fact, endocannabinoids are usually biosynthesized and released following depolarization of postsynaptic neurons or their stimulation with metabotropic receptor agonists, or both (Freund et al, 2003), and no evidence exists to date for GABA-induced endocannabinoid formation.
In conclusion, we have confirmed here the opposing roles of CB1 and TRPV1 receptors in the control of anxiety-like behaviors in mice, and have described for the first time the anxiolytic-like effects of AA-5-HT in the EPM test. We have also shown that CB1 and TRPV1 receptors exhibit a similar expression pattern in the mouse brain areas involved in fear and anxiety, thus providing anatomical support to the present pharmacological and biochemical experiments, suggesting that AA-5-HT owes its anxiolytic-like action to its property of dual indirect CB1 ‘enhancer’ and TRPV1 blocker. Further studies are now required to investigate the effect of AA-5-HT in other animal models of anxiety, and to evaluate its toxicological profile in animals and its safety in humans, to substantiate the potential therapeutic use of this or other dual FAAH/TRPV1 inhibitors in affective disorders.
References
Ahluwalia J, Urban L, Bevan S, Nagy I (2003). Anandamide regulates neuropeptide release from capsaicin-sensitive primary sensory neurons by activating both the cannabinoid 1 receptor and the vanilloid receptor 1 in vitro. Eur J Neurosci 17: 2611–2618.
Bisogno T, Cascio MG, Saha B, Mahadevan A, Urbani P, Minassi A et al (2006). Development of the first potent and specific inhibitors of endocannabinoid biosynthesis. Biochim Biophys Acta 1761: 205–212.
Bisogno T, Di Marzo V (2007). Short- and long-term plasticity of the endocannabinoid system in neuropsychiatric and neurological disorders. Pharmacol Res 56: 428–442.
Bisogno T, Melck D, De Petrocellis L, Bobrov MYu, Gretskaya NM, Bezuglov VV et al (1998). Arachidonoylserotonin and other novel inhibitors of fatty acid amide hydrolase. Biochem Biophys Res Commun 248: 515–522.
Blankman JL, Simon GM, Cravatt BF (2007). A comprehensive profile of brain enzymes that hydrolyze the endocannabinoid 2-arachidonoylglycerol. Chem Biol 14: 1347–1356.
Consoli D, Leggio GM, Mazzola C, Micale V, Drago F (2007). Behavioral effects of the beta3 adrenoceptor agonist SR58611A: is it the putative prototype of a new class of antidepressant/anxiolytic drugs? Eur J Pharmacol 573: 139–147.
Crawley JN, Belknap JK, Collins A, Crabbe JC, Frankel W, Henderson N et al (1997). Behavioral phenotypes of inbred mouse strains: implications and recommendations for molecular studies. Psychopharmacology 132: 107–124.
Cristino L, De Petrocellis L, Pryce G, Baker D, Guglielmotti V, Di Marzo V (2006). Immunohistochemical localization of cannabinoid type 1 and vanilloid transient receptor potential vanilloid type 1 receptors in the mouse brain. Neuroscience 139: 1405–1415.
de Lago E, Petrosino S, Valenti M, Morera E, Ortega-Gutierrez S, Fernandez-Ruiz J et al (2005). Effect of repeated systemic administration of selective inhibitors of endocannabinoid inactivation on rat brain endocannabinoid levels. Biochem Pharmacol 70: 446–452.
De Petrocellis L, Bisogno T, Davis JB, Pertwee RG, Di Marzo V (2000). Overlap between the ligand recognition properties of the anandamide transporter and the VR1 vanilloid receptor: inhibitors of anandamide uptake with negligible capsaicin-like activity. FEBS Lett 483: 52–56.
Devane WA, Hanus L, Breuer A, Pertwee RG, Stevenson LA, Griffin G et al (1992). Isolation and structure of a brain constituent that binds to the cannabinoid receptor. Science 258: 1946–1949.
Di Marzo V, Bisogno T, De Petrocellis L (2007). Endocannabinoids and related compounds: walking back and forth between plant natural products and animal physiology. Chem Biol 14: 741–756.
Di Marzo V, Capasso R, Matias I, Aviello G, Petrosino S, Borrelli F et al (2008). The role of endocannabinoids in the regulation of gastric emptying: alterations in mice fed a high-fat diet. Br J Pharmacol 153: 1272–1280.
Di Marzo V, De Petrocellis L, Fezza F, Ligresti A, Bisogno T (2002). Anandamide receptors. Prostaglandins Leukot Essent Fatty Acids 66: 377–391.
Di Marzo V, Maccarrone M (2008). FAAH and anandamide: is 2-AG really the odd one out? Trends Pharmacol Sci 29: 229–233.
File SE (1992). Usefulness of animal models with newer anxiolytics. Clin Neuropharmacol 15: 525A–526A.
Fowler CJ, Tiger G, López-Rodríguez ML, Viso A, Ortega-Gutiérrez S, Ramos JA (2003). Inhibition of fatty acid amidohydrolase, the enzyme responsible for the metabolism of the endocannabinoid anandamide, by analogues of arachidonoyl-serotonin. J Enzyme Inhib Med Chem 18: 225–231.
Freund TF, Katona I, Piomelli D (2003). Role of endogenous cannabinoids in synaptic signaling. Physiol Rev 83: 1017–1066.
Griebel G, Belzung C, Perrault G, Sanger DJ (2000). Differences in anxiety-related behaviours and in sensitivity to diazepam in inbred and outbred strains of mice. Psychopharmacology 148: 164–170.
Hermann H, De Petrocellis L, Bisogno T, Schiano Moriello A, Lutz B, Di Marzo V (2003). Dual effect of cannabinoid CB1 receptor stimulation on a vanilloid VR1 receptor-mediated response. Cell Mol Life Sci 60: 607–616.
Hohmann AG, Suplita II RL (2006). Endocannabinoid mechanisms of pain modulation. AAPS J 8: E693–E708.
Huang SM, Bisogno T, Trevisani M, Al-Hayani A, De Petrocellis L, Fezza F et al (2002). An endogenous capsaicin-like substance with high potency at recombinant and native vanilloid VR1 receptors. Proc Natl Acad Sci USA 99: 8400–8405.
Hwang D (2000). Fatty acids and immune responses: a new perspective in searching for clues to mechanism. Annu Rev Nutr 20: 431–456.
Kasckow JW, Mulchahey JJ, Geracioti Jr TD (2004). Effects of the vanilloid agonist olvanil and antagonist capsazepine on rat behaviors. Prog Neuropsychopharmacol Biol Psychiatry 28: 291–295.
Kathuria S, Gaetani S, Fegley D, Valiño F, Duranti A, Tontini A et al (2003). Modulation of anxiety through blockade of anandamide hydrolysis. Nat Med 9: 76–81.
Kim SR, Lee da Y, Chung ES, Oh UT, Kim SU, Jin BK (2007). Roles of transient receptor potential vanilloid subtype 1 and cannabinoid type 1 receptors in the brain: neuroprotection versus neurotoxicity. Mol Neurobiol 35: 245–254.
Lafenetre P, Chaouloff F, Marsicano G (2007). The endocannabinoid system in the processing of anxiety and fear and how CB1 receptors may modulate fear extinction. Pharmacol Res 56: 367–381.
Lee J, Di Marzo V, Brotchie JM (2006). A role for vanilloid receptor 1 (TRPV1) and endocannabinnoid signalling in the regulation of spontaneous and L-DOPA induced locomotion in normal and reserpine-treated rats. Neuropharmacology 51: 557–565.
Leggio GM, Micale V, Drago F (2008). Increased sensitivity to antidepressants of D(3) dopamine receptor-deficient mice in the forced swim test (FST). Eur Neuropsychopharmacol 18: 271–277.
Lepicard EM, Joubert C, Hagneau I, Perez-Diaz F, Chapouthier G (2000). Differences in anxiety-related behavior and response to diazepam in BALB/cByJ and C57BL/6J strains of mice. Pharmacol Biochem Behav 67: 739–748.
Lutz B (2007). The endocannabinoid system and extinction learning. Mol Neurobiol 36: 92–101.
Maccarrone M, Rossi S, Bari M, De Chiara V, Fezza F, Musella A et al (2008). Anandamide inhibits metabolism and physiological actions of 2-arachidonoylglycerol in the striatum. Nat Neurosci 11: 152–159.
Maione S, Bisogno T, de Novellis V, Palazzo E, Cristino L, Valenti M et al (2006). Elevation of endocannabinoid levels in the ventrolateral periaqueductal grey through inhibition of fatty acid amide hydrolase affects descending nociceptive pathways via both cannabinoid receptor type 1 and transient receptor potential vanilloid type-1 receptors. J Pharmacol Exp Ther 316: 969–982.
Maione S, De Petrocellis L, de Novellis V, Moriello AS, Petrosino S, Palazzo E et al (2007). Analgesic actions of N-arachidonoyl-serotonin, a fatty acid amide hydrolase inhibitor with antagonistic activity at vanilloid TRPV1 receptors. Br J Pharmacol 150: 766–781.
Marinelli S, Di Marzo V, Florenzano F, Fezza F, Viscomi MT, van der Stelt M et al (2007). N-arachidonoyl-dopamine tunes synaptic transmission onto dopaminergic neurons by activating both cannabinoid and vanilloid receptors. Neuropsychopharmacology 32: 298–308.
Marinelli S, Pascucci T, Bernardi G, Puglisi-Allegra S, Mercuri NB (2005). Activation of TRPV1 in the VTA excites dopaminergic neurons and increases chemical- and noxious-induced dopamine release in the nucleus accumbens. Neuropsychopharmacology 30: 864–870.
Marsch R, Foeller E, Rammes G, Bunck M, Kössl M, Holsboer F et al (2007). Reduced anxiety, conditioned fear, and hippocampal long-term potentiation in transient receptor potential vanilloid type 1 receptor-deficient mice. J Neurosci 27: 832–839.
Marsicano G, Wotjak CT, Azad SC, Bisogno T, Rammes G, Cascio MG et al (2002). The endogenous cannabinoid system controls extinction of aversive memories. Nature 418: 530–534.
Mechoulam R, Ben-Shabat S, Hanus L, Ligumsky M, Kaminski NE, Schatz AR et al (1995). Identification of an endogenous 2-monoglyceride, present in canine gut, that binds to cannabinoid receptors. Biochem Pharmacol 50: 83–90.
Moreira FA, Kaiser N, Monory K, Lutz B (2008). Reduced anxiety-like behaviour induced by genetic and pharmacological inhibition of the endocannabinoid-degrading enzyme fatty acid amide hydrolase (FAAH) is mediated by CB1 receptors. Neuropharmacology 54: 141–150.
Morgese MG, Cassano T, Cuomo V, Giuffrida A (2007). Anti-dyskinetic effects of cannabinoids in a rat model of Parkinson's disease: role of CB(1) and TRPV1 receptors. Exp Neurol 208: 110–119.
Movahed P, Jonsson BA, Birnir B, Wingstrand JA, Jorgensen TD, Ermund A et al (2005). Endogenous unsaturated C18 N-acylethanolamines are vanilloid receptor (TRPV1) agonists. J Biol Chem 280: 38496–38504.
Naderi N, Haghparast A, Saber-Tehrani A, Rezaii N, Alizadeh AM, Khani A et al (2008). Interaction between cannabinoid compounds and diazepam on anxiety-like behaviour of mice. Pharmacol Biochem Behav 89: 64–75.
Naidu PS, Varvel SA, Ahn K, Cravatt BF, Martin BR, Lichtman AH (2007). Evaluation of fatty acid amide hydrolase inhibition in murine models of emotionality. Psychopharmacology 192: 61–70.
Ortar G, Cascio MG, De Petrocellis L, Morera E, Rossi F, Schiano-Moriello A et al (2007). New N-arachidonoylserotonin analogues with potential ‘dual’ mechanism of action against pain. J Med Chem 50: 6554–6569.
Oz M (2006). Receptor-independent actions of cannabinoids on cell membranes: Focus on endocannabinoids. Pharmacol Ther 111: 114–144.
Patel S, Hillard CJ (2006). Pharmacological evaluation of cannabinoid receptor ligands in a mouse model of anxiety: further evidence for an anxiolytic role for endogenous cannabinoid signaling. J Pharmacol Exp Ther 318: 304–311.
Pellow S, Chopin P, File SE (1985). Are the anxiogenic effects of yohimbine mediated by its action at benzodiazepine receptors? Neurosci Lett 55: 5–9.
Rubino T, Realini N, Castiglioni C, Guidali C, Viganó D, Marras E et al (2008). Role in Anxiety Behavior of the Endocannabinoid System in the Prefrontal Cortex. Cereb Cortex 18: 1292–1301.
Starowicz K, Cristino L, Di Marzo V (2008). TRPV1 receptors in the central nervous system: potential for previously unforeseen therapeutic applications. Curr Pharm Des 14: 42–54.
Starowicz K, Maione S, Cristino L, Palazzo E, Marabese I, Rossi F et al (2007b). Tonic endovanilloid facilitation of glutamate release in brainstem descending antinociceptive pathways. J Neurosci 27: 13739–13749.
Starowicz K, Nigam S, Di Marzo V (2007a). Biochemistry and pharmacology of endovanilloids. Pharmacol Ther 114: 13–33.
Steiner MA, Wanisch K, Monory K, Marsicano G, Borroni E, Bächli H et al (2008). Impaired cannabinoid receptor type 1 signaling interferes with stress-coping behavior in mice. Pharmacogenomics J 8: 196–208.
Sugiura T, Kondo S, Sukagawa A, Nakane S, Shinoda A, Itoh K et al (1995). 2-Arachidonoylglycerol: a possible endogenous cannabinoid receptor ligand in brain. Biochem Biophys Res Commun 215: 89–97.
Tzavara ET, Li DL, Moutsimilli L, Bisogno T, Di Marzo V, Phebus LA et al (2006). Endocannabinoids activate transient receptor potential vanilloid 1 receptors to reduce hyperdopaminergia-related hyperactivity: therapeutic implications. Biol Psychiatry 59: 508–515.
van der Stelt M, Mazzola C, Esposito G, Matias I, Petrosino S, De Filippis D et al (2006). Endocannabinoids and beta-amyloid-induced neurotoxicity in vivo: effect of pharmacological elevation of endocannabinoid levels. Cell Mol Life Sci 63: 1410–1424.
Van Gaal L, Pi-Sunyer X, Després JP, McCarthy C, Scheen A (2008). Efficacy and safety of rimonabant for improvement of multiple cardiometabolic risk factors in overweight/obese patients: pooled 1-year data from the Rimonabant in Obesity (RIO) program. Diabetes Care 31 (Suppl 2): S229–S240.
Viveros MP, Marco EM, Llorente R, López-Gallardo M (2007). Endocannabinoid system and synaptic plasticity: implications for emotional responses. Neural Plast 2007: 52908.
Zygmunt PM, Petersson J, Andersson DA, Chuang H, Sørgård M, Di Marzo V et al (1999). Vanilloid receptors on sensory nerves mediate the vasodilator action of anandamide. Nature 400: 452–457.
Acknowledgements
These experiments were supported in part by the PhD International School Program in Neuropharmacology, University of Catania Medical School. We thank Dr C Wotjak, Max Planck Institute of Psychiatry, Munich, Germany, for the gift of perfused TRPV1 knockout mouse brains. Technical assistance from Mr Davide Castelluccio is also acknowledged.
Author information
Authors and Affiliations
Corresponding author
Additional information
Disclosures/Conflicts of interest
The authors declare that they have no conflicting interests.
Rights and permissions
About this article
Cite this article
Micale, V., Cristino, L., Tamburella, A. et al. Anxiolytic Effects in Mice of a Dual Blocker of Fatty Acid Amide Hydrolase and Transient Receptor Potential Vanilloid Type-1 Channels. Neuropsychopharmacol 34, 593–606 (2009). https://doi.org/10.1038/npp.2008.98
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/npp.2008.98
Keywords
This article is cited by
-
Potential application of endocannabinoid system agents in neuropsychiatric and neurodegenerative diseases—focusing on FAAH/MAGL inhibitors
Acta Pharmacologica Sinica (2020)
-
Cannabinoids, TRPV and nitric oxide: the three ring circus of neuronal excitability
Brain Structure and Function (2020)
-
The potential value of capsaicin in modulating cognitive functions in a rat model of streptozotocin-induced Alzheimer’s disease
The Egyptian Journal of Neurology, Psychiatry and Neurosurgery (2019)
-
Opposing roles of dorsomedial hypothalamic CB1 and TRPV1 receptors in anandamide signaling during the panic-like response elicited in mice by Brazilian rainbow Boidae snakes
Psychopharmacology (2019)
-
Interaction between interleukin-1β and type-1 cannabinoid receptor is involved in anxiety-like behavior in experimental autoimmune encephalomyelitis
Journal of Neuroinflammation (2016)
