
Abstract
In all, 19 research subjects, with current histories of frequent cocaine use, were exposed to cocaine-related cues to elicit drug craving. We measured the change of occupancy of dopamine at D2-like receptors with positron emission tomography (PET) and inferred a change of intrasynaptic dopamine (endogenous dopamine release), based on the displacement of radiotracer [11C]raclopride. Receptor occupancy by dopamine increased significantly in putamen of participants who reported cue-elicited craving compared to those who did not. Further, the intensity of craving was positively correlated with the increase in dopamine receptor occupancy in the putamen. These results provide direct evidence that occupancy of dopamine receptors in human dorsal striatum increased in proportion to subjective craving, presumably because of increased release of intrasynaptic dopamine.
Similar content being viewed by others
INTRODUCTION
In individuals with drug abuse disorders, environmental stimuli associated with drug use can trigger craving, increasing the risk of relapse (O'Brien et al, 1990). The neurobiology of this complex phenomenon has been investigated over the past decade with brain imaging techniques, following experimental induction of cocaine craving in susceptible individuals. Thus far, activations or deactivations of the brain in response to presentation of cocaine-associated cues have been shown by positron emission tomography (PET) studies of glucose metabolism (Bonson et al, 2002; Grant et al, 1996) or blood flow (Kilts et al, 2001, 2004; Childress et al, 1999), as well as functional magnetic resonance imaging (fMRI) (Wexler et al, 2001; Risinger et al, 2005; Maas et al, 1998; Garavan et al, 2000; Breiter and Rosen, 1999). These results demonstrate that cocaine craving involves brain regions that are known to process cognitive information about memories and emotion. Some of these regions, such as the prefrontal cortex (Wexler et al, 2001; Risinger et al, 2005; Grant et al, 1996; Garavan et al, 2000; Bonson et al, 2002), anterior cingulate cortex (Childress et al, 1999; Garavan et al, 2000; Kilts et al, 2001, 2004; Maas et al, 1998; Risinger et al, 2005; Wexler et al, 2001), amygdala (Kilts et al, 2001; Grant et al, 1996; Childress et al, 1999; Breiter and Rosen, 1999; Bonson et al, 2002), and striatum (Risinger et al, 2005; Kilts et al, 2001, 2004; Garavan et al, 2000; Childress et al, 1999), are known to receive dopaminergic input (Alexander et al, 1990; Berke and Hyman, 2000; Dolan et al, 1995; Gerfen and Wilson, 1996), but none of these studies directly investigated the neurochemistry of craving.
Whereas both the rewarding effects of cocaine and the maintenance of addiction are known to involve dopaminergic neurotransmission (Amara and Sonders, 1998; Cannon and Bseikri, 2004; Fiorillo et al, 2003; Schultz, 2001; Volkow et al, 2004), previous studies of the effect of cocaine craving on dopaminergic functioning have not yielded conclusive results. Three previous studies found a correlation between cocaine craving and concentrations of the dopamine metabolite homovanillic acid (HVA) in either plasma or cerebrospinal fluid of cocaine abusers (Berger et al, 1996; Knoblich et al, 1992; Martin et al, 1989). A fourth study found higher levels of CSF HVA in recently abstinent cocaine-dependant patients as compared to controls, but no correlation between HVA levels and duration of abstinence (Roy et al, 2002). Administration of haloperidol, a dopamine receptor antagonist, reduced cue-induced cocaine craving in abstinent cocaine abusers in one study (Berger et al, 1996), but had a limited effect on subjective effects of cocaine in another (Sherer et al, 1989).
To further explore the relationship between cocaine craving and dopamine receptor (DAR) occupancy in cocaine abusers, we first tested whether cue-induced cocaine craving induced an increase in DAR occupancy, presumably by endogenous dopamine release. We then tested whether self-reports of cocaine craving correlate with the magnitude of DAR occupancy. In these tests, we used [11C]raclopride, a dopamine receptor antagonist that competes with endogenous dopamine for binding to dopamine D2 and D3 receptors, to measure the apparent D2/D3 binding potential as an indication of endogenous dopamine activity.
We tested the hypotheses that (1) cocaine abusers who report increased cocaine craving in response to cocaine-related cues (‘cravers’) have a greater DAR occupancy than abusers who do not report increased craving (‘non-cravers’), and (2) the change of DAR occupancy correlates with the intensity of craving for cocaine.
METHODS
Participants and Design
In all, 19 individuals (16 men and three women; 16 African-Americans, two Caucasians, and one Hispanic) who met DSM-IV-TR criteria for stimulant abuse participated in the study. The age of the participants ranged from 33 to 44 years (mean=40.6 years). They reported snorting, smoking, and/or injecting cocaine at least twice a week for an average of 13 years (range: 3–26 years). All identified cocaine as their drug of choice, 18 of the 19 subjects had current or past histories of nicotine dependence, and all subjects had some history of other illicit substance abuse. The subjects were not seeking treatment for their addiction. Exclusionary criteria included lifetime history of any DSM-IV-TR axis I diagnoses (including dependence on any drug other than cocaine or nicotine), evidence of physical disease, history of head trauma, claustrophobia, or pregnancy. All participants received a detailed explanation of the procedures and provided written consent. Procedures were reviewed and approved by the Institutional Review Boards of the Johns Hopkins University and the National Institute on Drug Abuse.
Experimental Procedures
All participants tested positive for cocaine metabolites in urine at the screening interview, upon admission to an inpatient unit, or both, and reported regular recent cocaine use (at least twice per week). Participants resided on a research ward from at least 24 h before to at least 24 h after their participation in this study. During this time, participants had no access to illicit drugs, alcohol, or psychotropic medications. Participants were not allowed to use nicotine or caffeine for at least 6 h prior to the start of the brain imaging procedures on the study day.
When participants arrived at the PET center on the morning of the study, a venous catheter was placed in the antecubital vein for injection of the radioligand. Individuals were positioned on the scanner bed to allow a clear view of an overhead video monitor. [11C]raclopride (Ehrin et al, 1985), produced at the Johns Hopkins Hospital Biomedical Cyclotron facility, was injected at the beginning of each of two 90-min PET scans, which were separated by 2 h. For the first PET session, subjects received an average dose of 17.4±0.31 mCi (SEM), with an average mass of 1.19±0.13 μg, and an average specific activity of 6734±723 mCi/μmol. The administered dose for the second PET session averaged 17.6±0.26 mCi, with an average mass of 0.99±0.09 μg, and an average specific activity of 7948±861.23 mCi/μmol. The administered doses and specific activities of raclopride were not significantly different across the two sessions.
The first PET scan was obtained while the participant was exposed to neutral cues that consisted of a 10-min neutral cues videotape and a 45-min audiotape description of an outdoor experience involving natural beauty. The second PET scan was obtained while the participant was exposed to cocaine-related cues that consisted of a 10-min videotape of cocaine use and a 45-min audiotape of a variety of pleasurable experiences from cocaine use as described in interviews with cocaine abusers. Each video and audio tape was repeated continuously throughout the PET session, similar to prior studies of cerebral glucose metabolism during cocaine craving (Bonson et al, 2002; Grant et al, 1996). As in previously reported studies (Bonson et al, 2002; Grant et al, 1996), neutral cues were always shown in the first session to prevent associations of the study setting with the drug-related stimuli and to prevent any carry-over effects from the first session. By doing both sessions on the same day, participant drop-out was minimized.
Prior to the start of each session, and at 10-min intervals during the PET scanning, participants were asked six questions to assess their mental state. Three questions explicitly assessed craving (ie, ‘Do you have a craving or urge for cocaine?’, ‘Do you want cocaine?’, and ‘Do you need cocaine?’). Participants responded orally to these questions using a Likert-type scale ranging from 0 (no, not at all) to 10 (yes, extremely). The other three questions concerned mood and wakefulness (‘How good do you feel?’, ‘How ‘turned off’ do you feel?’, and ‘How awake are you?’), and were asked at the same time intervals. Responses to two of these questions (‘good’ and ‘awake’) were made by identifying which of five mannequins with different facial expressions best represented their internal state. Responses to the ‘turned-off’ questions used the Likert-type scale.
A session-appropriate evocative script was read aloud to participants following collection of baseline subjective measures, and was repeated midway through the scan. Each script was constructed to present a vivid description of the emotions and sensations associated with either nature or cocaine use, and was read by one of the investigators in an effusive manner as described previously (Bonson et al, 2002).
Thus, each session had the following schedule: (1) 10–15 min before PET scan: baseline subjective ratings collected; (2) 10 min before PET scan: session-appropriate script read, followed by start of audio and visual tapes; (3) 5 min after PET scan start: experimental subjective ratings collected; (4) 5–90 min after scan start: additional subjective ratings collected every 10 min; (5) 40–50 min after scan start: session-appropriate script re-read.
PET images were acquired on a GE Advance PET camera, which simultaneously acquired 35 slices, with an axial field of view of 15.2 cm. The scanning protocol consisted of 32 frames of 15 sec to 5 min each: four 15-sec frames, four 30-sec frames, three 1-min frames, two 2-min frames, five 4-min frames, and 12 5-min frames. Data were acquired in 3-D mode. The images were reconstructed using filtered back-projection with a ramp filter (image size 128 × 128, pixel size 2 × 2 mm2, inter-slice spacing 4.25 mm), resulting in a spatial resolution of 5.5 × 5.5 × 4.25 mm full-width-at-half-maximum at the center of the field of view. Data in the decay-corrected, reconstructed, dynamic images were expressed in units of concentration (μCi/ml).
In addition to the PET scan, each participant had a volumetric magnetic resonance imaging (MRI) scan for coregistration with the PET data to enhance anatomical definitions. The MRI data were acquired using a Spoiled GRASS (gradient-recalled acquisition in the steady state) (SPGR) sequence on a GE 1.5 T Signa Camera. To minimize movement during the MRI and PET scans, each participant was fitted with a thermoplastic facemask that was secured to the scanner bed during imaging.
Subject Grouping
In order to define craving operationally, we first calculated the mean of each participant's responses to the two questions ‘Do you want cocaine?’ and ‘Do you have a craving or urge for cocaine?’ over all nine post-baseline intervals for each session. We then took the average of these nine calculated means, obtaining a mean craving score for the responses during the cocaine cue videotape and another for the responses during the neutral videotape for each participant. The change in craving was calculated as the mean craving score during the craving videotape scan minus the mean craving score during the neutral videotape scan.
We did not include responses for the third craving question in our calculation of craving score (‘How much do you need cocaine?’) because so few of the research participants reported ‘needing’ cocaine during either scan. In addition, their mean responses to this item during the neutral (0.04±0.15) and cocaine cue (0.62±1.79) videotape scans were not significantly different (paired-sample t(18)=−0.151; p=0.15).
We calculated the craving score as a continuous variable for testing correlation with PET parameters. For the group analysis, a participant was classified as showing cue-induced craving (ie, being a ‘craver’) if he or she produced a larger mean craving score during the cocaine cues videotape scan than during the neutral cues videotape scan. Participants were classified as ‘non-cravers’ if they reported no cue-elicited increase in craving (ie, produced a difference score of 0). We also calculated each participant's peak craving as the highest mean response to the two questions about wanting or craving cocaine while he or she watched the cocaine cue videotape. We refer to this as the ‘peak’ craving reported by subjects. Both ‘mean’ and ‘peak’ craving scores were correlated with the results of DAR occupancy.
Image Analysis
The primary measure obtained with the PET measurements was binding potential (BP), which is the ratio of the density of available receptors to the dissociation constant (KD) of raclopride for dopamine D2 and D3 receptors (Mintun et al, 1984). The secondary measure was the percent change in BP from the neutral session to the cocaine session. The relative reduction in BP was taken as a change in DAR occupancy following cue exposure, whether due to an increase in dopamine release or a change in dopamine affinity, or both. It is still uncertain to what extent changes in BP reflect changes of dopamine concentration or affinity of D2/D3 receptors for dopamine (Gjedde et al, 2005). Here, we consider the term ‘occupancy’ as the most appropriate description. We determined BP during neutral and cocaine-cued PET sessions and calculated the change of BP and DAR occupancy. We made these estimates both in volumes-of-interest (VOIs) and in individual voxels of the image volume, and displayed the latter as parametric maps. We used two methods to generate parametric maps: a method using the multilinear reference tissue model with two parameters (MRTM2) (Ichise et al, 2003), and an approach using a linearized simplified reference tissue model, (LSRTM) (Zhou et al, 2003), as described below. Both sets of maps were used in statistical analyses using the program SPM2 (Wellcome Department of Imaging Neuroscience, London, UK).
Volumes-of-Interest and Derivation of Outcome Measures
Volumes of interest (VOIs) were defined for individual subjects on Spoiled Grass (SPGR) sequenced MRI volumes for the caudate nucleus and the putamen bilaterally, and for the cerebellum using interactive segmentation software. The software allowed users to select upper and lower MRI intensity thresholds to delineate the structure of interest such that manual drawing was required only to limit spatial boundaries of the structure when there was continuation of the within-threshold voxels to other structures. The software also allowed users to define regions-of-interest (ROIs) in any of three orthogonal planes, which facilitates choosing the entire structure of interest as a VOI. The ventral striatum, which was indistinguishable on SPGR MRI volumes, was separated from the caudate and putamen VOIs using published anatomical guidelines (Baumann et al, 1999). Briefly, MRI volumes were reoriented such that the mid-plane was vertical and the anterior-comissure-posterior commissure plane was horizontal. Bisectors of internal capsule were defined on each coronal slice as regression lines of voxels that were equidistant from outlines of putamen and caudate nucleus VOIs. Separately, the ventral boundaries of lateral ventricles were identified and fitted by a polynomial function on each side. The line (the ventral striatum separator) was defined such that it was perpendicular to the internal capsule bisectors and passed through the ventral corner of the lateral ventricle on each side. The so-defined ventral striatum separators were transferred back to the original MRI space. Of the voxels of putamen and caudate VOIs, those ventral to the plane made by the ventral striatum separators were classified as the ventral striatum. Dorsal putamen and caudate nucleus VOIs were divided into respective anterior and posterior subdivisions by the anterior commissural plane, which was perpendicular to the mid-plane and the anterior–posterior commissural plane (Martinez et al, 2003). The automated implementation of the guidelines was published previously (Oswald et al, 2005).
VOIs were transferred from MRI to PET spaces as follows: First, individuals' MRI volumes were coregistered to mean PET volumes (averages from 10 to 90 min frames) of neutral and cue sessions, respectively, using information theory (Collignon et al, 1995) as implemented in SPM2 (http://www.fil.ion.ucl.ac.uk/spm). The transformation parameters were applied to VOI masks to transfer VOIs to PET spaces. Subsequently, VOIs were applied to individual frames to obtain time-activity curves (TACs) of regions. Frames acquired between 3 and 90 min were coregistered to the 20 min frame using SPM2 image coregistration routines to correct for between-frame head motion. Transformation parameters from the 3 min frame were applied to frames taken before 3 min.
BP values of striatal regions were obtained by the Logan graphical analysis (Logan et al, 1990, 1996), using the cerebellum as the reference region. Changes in DAR occupancy were calculated as 1 minus the ratio of the cue to neutral session BP and then expressed as a percentage.
Statistical Analysis
BP and change in DAR occupancy (calculated as the percent change in BP) were extracted from the VOIs described above, for each subject. Means and SD were calculated for BP and change in DAR occupancy for each striatal VOI for both craving and non-craving groups. As the cerebellum VOI was the reference region, it was not included in the statistical analysis. To test for between-groups differences, we used a robust linear regression model, where group status (ie, craver vs non-craver) was encoded using dummy variables, and set as the independent variable. The PET variables (BP and change in DAR occupancy) were set as the dependent variables, and age was input as a covariate.
The change in DAR occupancy and craving scores for each individual participant were incorporated into another linear regression to test for correlations between the two variables. All statistical analyses were completed using Stata 8 (StataCorpLP, College Station, Texas).
Statistical Parametric Mapping (Using MRTM2)
We constructed parametric maps of BP of [11C]raclopride using MRTM2 and the cerebellum as the reference region (Ichise et al, 2003). The parametric maps were coregistered to standard brain space (Talairach and Tournoux, 1988), in two steps. First, individual subjects' MRIs were spatially normalized to a standard T1 volume of SPM2. The spatial normalization parameters were used to transfer BP volumes to the standard space. The spatially normalized BP volumes were averaged across all subjects to generate a standard BP volume template. Then, BP volumes were spatially normalized to the standard BP volume (Meyer et al, 1999) and smoothed with a Gaussian kernel of 10 mm, full-width-at half-maximum. Volumes of change in DAR occupancy were constructed in the same manner as in the VOI analysis (percent change in BP). We performed a voxel-wise comparison of change in DAR occupancy in craving and noncraving subjects using Statistical Parametric Mapping software (Friston et al, 1991) (SPM2) within a level of p<0.005, uncorrected, and the minimal cluster size at 21 voxels (0.1 ml). A voxel-wise simple regression was used to identify voxels where change in DAR occupancy correlated with changes in average craving score between the neutral and cocaine cues sessions. The same criteria used for voxel-wise t-test were applied for these analyses.
We set the minimum number of voxels for clusters at 21 voxels for the following reasons: first, SPM2 yields p-values for the statistical significance regarding volumes (ie, extent thresholds). In our setting (10 mm filter, the search volume=28.7 ml), a cluster of 22 voxels had a p-value of 0.047 for the p value determined by SPM2. Second, SPM2 also provides the volume of the ‘resolution element’ (resel), a statistical volume which represents the appropriate sampling element for a given data set of PET images (Worsley et al, 1992), which was 20.3 in our setting.
Statistical Parametric Mapping (Using LSRTM)
We also performed the SPM2 analyses with parametric maps for BP and DAR occupancy generated by LSRTM-based approach (Zhou et al, 2003). To create the parametric maps, an automated frame-to-frame image registration was first applied to dynamic images (Lin et al, 1996). Then, parametric images of BP were generated with LSRTM-based linear regression with spatial constraint approach. These images were then spatially normalized, using SPM2, to standard space (Talairach and Tournoux, 1988). Lastly the same voxel-wise statistical analyses applied to the first set of parametric maps were applied to the LSRTM-based maps (Friston et al, 1991). Table 3 shows the peak voxels and associated statistics for the correlation of DAR occupancy significantly correlated with the mean craving scores. Thus, we were able to confirm our results with two different parametric modeling methods.
Choice of Models for Parametric Mapping
We employed two methods of parametric mapping, MRTM2 and LSRTM. The conventional ‘simplified reference tissue method’ (SRTM) yields regional BP values that essentially are identical to values obtained with compartmental deconvolution of tissue uptake functions from arterial input functions (Lammertsma and Hume, 1996). SRTM2, for the estimation of two model parameters, uses a common value for the fractional brain-to-blood clearance constant of the reference tissue (k2R) for all tissue regions (Wu and Carson, 2002). The multiple regression version of the tissue reference method, MRTM2 (Ichise et al, 2002), uses multiple linear regression as its deconvolution operation. Direct comparison of MRTM2 with SRTM and SRTM2 in simulated PET studies concluded that MRTM2 is as accurate as SRTM2, which in turn is superior to SRTM (Ichise et al, 2003), and computationally more efficient as well. The ‘linearized simplified reference tissue method’, LSRTM, uses ridge regression with spatial constraints rather than deconvolution to reduce noise-induced bias in the case of less than optimal signal-to-noise ratios (Zhou et al, 2003) and hence serves to replicate results when optimal signal-to-noise ratios are undetermined and could be relevant to the method used (Buchert et al, 2004).
RESULTS
Craving Response
In all, 11 of 19 (58%) participants reported an increase in craving for cocaine during exposure to the cocaine-related cues (‘cravers’). The means of their responses to the two craving questions during the cocaine cues videotape PET scans ranged from 0.33 to 6.17, with a mean of 2.47 (SD=2.03). The craving score for this group was significantly greater than zero, as tested by one-tailed t-test (t (10)=4.03; p=0.002). The mean craving score of cocaine abusers who did not show cue-induced increase in craving was 0.01 (SD=0.02), and this was not significantly greater than zero (t(7)=1.0; p=0.351). For descriptive purposes, we also recorded the peak mean of each individual's maximal responses to the questions about craving and wanting cocaine as an index of the maximal craving reported by each participant. The peak responses ranged from 0.5 to 8, with an average of 3.95, and were recorded at an average time of 25 min after the start of the scan. This peak craving was used as a secondary test of correlation with PET parameters based on the judgment that the average ratings of craving over the entire scan interval represented a more reliable measure of subjective drug craving.
Responses to the questions about wakefulness and mood were not statistically different (p>0.05) between the two groups or the two PET sessions (Table 1). Cravers and noncravers did not differ significantly in demographics nor degree or duration of cocaine use (see Table 2 for summary demographics). Cravers reported using cocaine for an average of 13.2±7.5 years (SD), which was not statistically different from the noncraver average of 12.5±5.34 years. Of the 11 cravers, 10 reported regular cigarette use, eight reported regular alcohol use, nine reported heroin use, seven reported marijuana use, and one subject reported also using tranquilizers. All of the eight noncravers reported smoking cigarettes regularly, all eight reported regular alcohol use, seven reported heroin use, six reported marijuana use, and two subjects reported using additional drugs, including tranquilizers and hallucinogens.
VOI Analysis
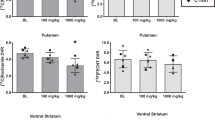
Cravers had a significantly greater change in DAR occupancy change in the right anterior putamen (4.3±1.0% SEM) than noncravers (0.9±1.0% SEM; p<0.05). This change was regionally specific and lateralized, as no significant changes were observed in other striatal areas. Furthermore, the change in DAR occupancy in left anterior putamen of cravers correlated positively both with time-averaged craving (r=0.64, p=0.033) and peak craving (r=0.75, p=0.008). The significance levels were determined by the linear regression methods described above in the methods.
SPM Analysis with MRTM2
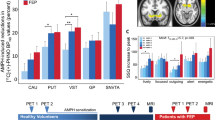
Using MRTM2 maps, we found that cravers had significantly greater change in DAR occupancy than noncravers in a cluster in the right anterior putamen (peak Talairach coordinates (28, 0, 4), peak t=7.6, volume=33 voxels, p=0.005, uncorrected) (Figure 2a, Table 3). As with the VOI method, the change of DAR occupancy correlated positively with mean craving score among cravers in the left anterior/posterior putamen (peak Talairach coordinates (−28, 2, 10), volume=133 voxels, peak t=7.6) (Figures 1 and 2b). This effect was lateralized, as the left anterior/posterior putamen showed a significantly stronger correlation than the mirrored anterior/posterior cluster on the right (see section on ‘Tests of Laterality’ below). No negative peaks were found in any of the SPM analyses.

Change in DAR occupancy: group effects and correlation with craving score. The figure shows the locations of the significant clusters found using SPM. Images show significant clusters at a threshold of p=0.005, minimum cluster size of 21 voxels. (a) The change in DAR occupancy in subjects who reported craving is greater than in subjects who did not report craving. This result was found using SPM with linearized SRTM-based parametric maps. Cluster is located in the right anterior putamen, Talairach coordinates for peak voxel (28, 0, 4). (b) Change in DAR occupancy correlated positively and significantly with subjective craving using MRTM2-based parametric maps. Cluster is located in the left anterior putamen, Talairach coordinates for peak voxe (−28, 2, 10). (c) Change in DAR occupancy correlation with subject craving using the LSRTM approach to parametric map construction. Cluster is located in the left anterior putamen but extends posteriorly, Talairach coordinates for two peak voxels (−36, −8, 4) and (−28, 14, 2). LSRTM also showed a peak on the right, at coordinates (4, 18, −2), but this cluster was not significant at the threshold chosen for our analysis. However, this peak suggests a bilateral effect, the detection of which is limited by sample size.

Correlation of change in DAR occupancy with craving score. Using the voxel-wise correlation in SPM2, we identified a cluster within the left anterior putamen where there was a significant correlation between change of DAR occupancy and subjective craving among participants who reported cue-induced craving (N=11).
SPM Analysis with LSRTM
When the same correlation analysis was run using the parametric maps generated by LSRTM approach, we found a similar result: DAR occupancy correlated positively with mean craving score among cravers in a left anterior/posterior putamen cluster with two continuous peaks (Talairach coordinates (−36, −8, 4), peak t=9.8 and (−28, 14, 2), peak t=5.4) (Table 3, Figure 2c). Furthermore, we also found a small cluster on the right where DAR occupancy correlated with mean craving with a peak at Talairach coordinates (4, 18, −2), but which was not significant at the threshold chosen for our SPM analysis.
Test of Laterality
We further tested the laterality of the correlation directly involving the putamen. First, we extracted change of DAR occupancy values from a VOI defined by the significant left anterior putamen cluster that emerged from the SPM correlation analysis (using MRTM2-based maps). The same VOI was then flipped to the right hemisphere and overlaid on the right anterior putamen. The change of DAR occupancy values for left and right anterior putamen and the individual craving scores were analyzed in a regression analysis, in which the left anterior putamen showed a significant correlation (r=0.76, p<0.0001) between the change in DAR occupancy and cocaine craving scores, whereas the right anterior putamen did not (r=0.11, p=0.9). Additionally, using a left–right interaction regression model, a comparison of the regression on the right vs left anterior putamen showed that the relationship between DAR occupancy and craving score was significantly stronger on the left anterior putamen than the right (p=0.005). This analysis suggested the left laterality of the correlation between self-reported craving and DAR occupancy. We did, however, show a small (33 voxels) correlation cluster on the right, using the LSRTM parametric approach for constructing maps. This finding suggests that a bilateral effect may in fact be present but not highly significant in our sample due to power considerations (see limitations below). Furthermore, the left putamen cluster found with the LSRTM-based maps extended into the posterior putamen (Figure 2c). Thus, the LSRTM approach suggests that our results may be bilateral and extend into the posterior putamen.
Analysis of Baseline BP and BP Change
To further extend our analyses of percent change in BP (and DAR occupancy), we tested for differences in BP at baseline (ie, during the neutral-cues PET scan) between subjects who craved and those who did not. Data obtained using the VOI method showed no statistically significant differences between the two groups. In addition, when we used SPM to compare the baseline BP images of the two groups, we also found no significant clusters.
We used change in BP from the neutral cues scan to the cocaine cues scan as an outcome measure in an SPM analysis. When applied to craving subjects only, SPM showed a cluster in the right anterior putamen, where the BP during the neutral cues session was significantly greater than the BP during the cocaine cues session (peak Talairach coordinates (20, 12, 2), t=3.57, and p=0.005, uncorrected; cluster size=24 voxels). When the same test was applied to noncravers, no significant clusters were found. These results were confirmed using the second method for BP map construction (Zhou et al, 2003). Therefore, the group difference in DAR at this location was in fact due to the BP reduction in the craving subjects and not to a change in BP in noncraving subjects. The findings also confirmed that the group difference in DAR was not an artifact of the division operation.
In addition, we identified a significant cluster in the left anterior putamen, where the BP difference (ie, BP of neutral cues session—BP of cocaine cues session) of craving subjects was correlated positively with mean craving score (peak Talairach coordinates (−24, 8, 2), t=6.56, p<0.005, uncorrected; cluster size=22 voxels). Again, this result was confirmed with the LSRTM method of binding map construction, and also confirmed the correlation we found with the change of DAR occupancy at this location.
DISCUSSION
In this study, among individuals with histories of cocaine abuse, self-reported cocaine craving in response to cue exposure was positively correlated with a change of DAR occupancy in the left putamen. These results provide direct evidence that DAR occupancy in human dorsal striatum increases proportionately with subjective increases in cocaine craving. This study also shows group differences in cravers vs noncravers for DAR occupancy during the presentation of cocaine cues.
Thus, unlike previous neuroimaging studies which demonstrated brain activation or changes in glucose metabolism in response to cocaine-associated cues (Bonson et al, 2002; Breiter and Rosen, 1999; Childress et al, 1999; Garavan et al, 2000; Grant et al, 1996; Kilts et al, 2001, 2004; Maas et al, 1998; Risinger et al, 2005; Wexler et al, 2001), our finding is linked directly to dopamine. There have been studies of dopamine in relation to cues for drinking alcohol (Heinz et al, 2004) or eating food (Wang et al, 2001), but those studies used single-scan measures of BP correlations with subjective effects (ie, BP with euphoric drug effects (Udo de Haes et al, 2005). Additionally, only a few other studies have directly demonstrated dynamic changes in human brain dopamine in relation to motivational states (Koepp et al, 1998) unrelated to drug abuse.
Several mechanisms might account for the increased DAR occupancy observed in the present study during cocaine craving. Recent research suggests that a change in dopamine release focuses the attention of an individual towards cues that predict salient stimuli (Franken, 2003). Within dopamine target areas, the ventral striatum (nucleus accumbens) is active during early exposure to reward when reward is unpredicted, whereas the dorsal striatum (putamen and caudate nucleus) is engaged by cues after the linking of cues and actions leading to reward have been established (Robbins and Everitt, 2002; Gerdeman et al, 2003).
It is well-known that the dorsal striatum is active during ritualized or habitual behavior (Yin et al, 2004; McDonald and White, 1993; Harlan and Garcia, 1998; Gerdeman et al, 2003; Faure et al, 2005; Berke, 2003), including drug-taking. More specifically, changes in DAR occupancy within dorsal striatal structures have been proposed to play a role in habit formation (Gerdeman et al, 2003; Berke, 2003), which is the tendency of an organism to repeat well-learned behaviors without requiring cognitive interventions. It has been theorized by Robbins and Everitt that while the initial positive effects of cocaine activate structures in the ventral striatum, a transition occurs when habitual drug-seeking behaviors are established and activation of the dorsal striatum predominates (Robbins and Everitt, 2002). Recent evidence has also shown that stimulation of dopamine and glutamate receptors in the dorsal striatum is critical for well-established drug seeking that depends on cocaine-associated stimuli (Vanderschuren et al, 2005).
Berke (2003) has suggested that habit formation is under prefrontal cortical control, allowing an organism to exert cognitive control over adaptive decision-making. Normally, the prefrontal cortex acts on both ventral and dorsal components of the striatum. However, studies with rodents show that this integrated circuit can be disrupted (Floresco et al, 2003). As tonic and phasic changes in intrasynaptic dopamine release occur in response to continued cocaine administration, there is a decrease in prefrontal control over ventral striatal structures (Goto and Grace, 2005) where the primary rewarding effects of the drug occur. Instead, the hippocampus becomes the primary region controlling the ventral striatum, which leaves the prefrontal cortex to act solely on the dorsal striatum. This change could lead to an increase in DAR occupancy in the putamen, which via a presynaptic action (West and Grace, 2002) would feed back on the prefrontal cortical input, further reducing cognitive control over habitual behaviors like drug taking. Thus, the craving-induced elevations of DAR occupancy in cocaine abusers that exhibit cue-induced craving in the present study may serve to further drive non-adaptive continuation of cocaine use in the face of adverse consequences because of the reduction in cognitive control (Schiltz, 2006).
There is also evidence to suggest that repeated experience with environmental contingencies resulting in reward may be necessary for involvement of putamen. For example, putamen activation has been observed in fMRI studies when healthy individuals are shown images of persons they love (Bartels and Zeki, 2000) and when alcohol-dependent subjects view alcohol-related cues (Grüsser et al, 2004; Heinz et al, 2004). Similarly, studies have shown that changes of dopamine release in the dorsal striatum rely on the degree of history with cocaine (Ito et al, 2002), such that putamen activation occurs only after chronic, but not acute, cocaine administration (Porrino et al, 2004; Saka et al, 2004). Putamen involvement may be the result of a shift in activation with prolonged cocaine administration, where the change of DAR occupancy occurs primarily in the ventral striatum during initial cocaine exposure but becomes more dorsal with chronic cocaine use (Ito et al, 2002). This shift in regional DAR occupancy is consistent with a ventral-to-dorsal progression in cerebral glucose metabolism following prolonged cocaine administration in primates (Porrino et al, 2004).
In a previous study of cerebral glucose metabolism, exposure to audiovisual cocaine cues resulted in craving that was correlated with increases in [18F]FDG-6-P in the right superior frontal gyrus, the left orbitofrontal cortex, and left amygdala (Bonson et al, 2002). Notably, the superior frontal gyrus projects to the same region of the dorsal striatum where the cue-related change of DAR occupancy was observed in the present study. Thus, the dorsal striatum may function as a convergence site for cortical and dopaminergic afferent drives during cue-induced craving. In contrast, the orbitofrontal cortex and amygdala project to the ventral striatum, where no change of DAR occupancy was observed in the present study. However, recent anatomical studies indicate that the ventral striatum is the source of descending projections to the portions of the substania nigra that contain dopaminergic projections to the dorsal striatum. The ventral striata of the primate project not only to the dopamine neurons that project back to the ventral striatum, but also innervate the dopaminergic neurons that project to more dorsal, motor striatal regions such as the putamen (Haber et al, 2000). In these ways, the present results complement prior functional imaging studies by providing an integrative view of the neuronal circuitry activated during cue-induced drug craving. They may also explain why metabolic activation of the more dorsal dopamine receptive brain regions was not observed in our previous studies of cerebral glucose metabolism (Bonson et al, 2002; Grant et al, 1996).
Another issue that remains to be resolved is the interpretation of the laterality of our findings in the putamen. Although the group difference in change in DAR occupancy was seen in the right putamen, the left putamen showed the correlation between the increase in DAR occupancy and craving score (Figures 1 and 2). Prior imaging studies of cocaine craving (Childress et al, 1999; Kilts et al, 2001, 2004), including our own studies of glucose metabolism (Bonson et al, 2002; Grant et al, 1996) have also shown laterality in terms of cue-induced craving. Cocaine administration produced greater changes of glucose metabolism in the right than in the left putamen (London et al, 1990). The more robust correlation of craving with a change of DAR occupancy in the left compared to the right putamen suggests that dopaminergic function in the left putamen contributes to the central processing and labeling of neurochemical changes that may be greater on the right. Alternatively, it is possible that the dopamine activity observed in the right anterior putamen may induce a change in the functionality of the left anterior putamen, or the effect may be bilateral (see Limitations below).
Knowledge of the level of basal extracellular dopamine prior to the cue exposure might be relevant to the interpretation of our results. As measured here, the BP reflects the density of available D2 and D3 receptors. Baseline BP levels in cravers and noncravers were similar with no significant group differences in BP as tested by SPM. It follows, therefore, that the primary contribution to the observed decrease in BP is an increase in DAR occupancy after cue exposure. Whereas the noncravers exhibited no significant change in BP between baseline and post-cue measurements, the cravers showed a significant difference. These results, therefore, support the notion that the change of DAR occupancy was not due to baseline differences between craving and noncraving subjects.
LIMITATIONS
In the correlation analysis of change in DAR occupancy and mean craving score, we included only those subjects who had a non-zero craving score. That is, only subjects who showed a positive increase in craving during the cocaine cues scan compared to the neutral cues scan were included. We limited the analysis of correlation to this subgroup because including users with a zero craving score (ie, no cue-induced response) may introduce aspects of the change in DAR occupancy that are independent of any craving focus and therefore were not part of the tested hypothesis that craving of any level (as defined in this paper) increases in a monotonic, dose-dependent manner with the change in DAR occupancy.
In the present study, as in our previous published FDG studies of cocaine craving (Bonson et al, 2002; Grant et al, 1996), scan order always consisted of the neutral cues in the first session in order to prevent associations of the study setting with the cocaine-related stimuli. The potential confounding effects of study setting contamination were considered to outweigh potential session order effects such as novelty of both the PET imaging and cue presentation in the first scan, which we attempted to minimize by familiarizing subjects with the PET technique and explaining in detail the cue challenge procedures prior to the first scan. In addition, diurnal fluctuations in dopamine amongst our subjects could be a possible confounds, but such minor fluctuations are unlikely to affect the results.
Cigarette smoking in our study sample could be a potential confound. All but one of the participants reported regular nicotine use, with an average of 6.5±1.1 (SD) cigarettes smoked per day. Between craving groups, there was no significant difference in cigarettes smoked (8.6±1.4 in non cravers vs 5.12±1.2 in cravers, p>0.1). We therefore can conclude that if nicotine withdrawal affected changes in DAR occupancy, it did so similarly across craving groups.
In the present study, the cocaine cues were sufficiently evocative to induce craving in 58% of the subjects. Notably, however, the degree of craving (change score from neutral cues session to cocaine cues session) in these subjects (2.5) was lower than the craving score our previous FDG study (4.1) using identical cocaine cues (Bonson et al, 2002). However, in both studies, the mean peak craving scores were similar (3.9 vs 5.1), and the maximal individual craving change score in both studies was similar (8.0 vs 8.7) (Bonson et al, 2002). The percent of subjects who experienced craving in the present study (58%) is also greater than that typically reported in studies of drug/alcohol cues, where 33% experience no craving (change score of 0), 33% experience a low degree of craving (change score of less than 2.0) and 33% experience a high degree of craving (change score of greater than 2.0) (Avants et al, 1995; Bonson et al, 2002; Grant et al, 1996; Newlin, 1992; Rohsenow et al, 1990). These differences in craving may be accountable by certain limitations in the present methodology.
Given that our experimental session was three times as long than in our previous FDG studies (Bonson et al, 2002; Grant et al, 1996), subjects were re-exposed nine times to the same short videotape of cocaine use, which may have produced boredom or frustration that competed with craving responses. Subjects were also lying on a flat scanner bed during cue exposure, rather than upright in a comfortable overstuffed chair as in our previous study. In addition, although subjects heard a continuously changing audiotape describing how cocaine abusers use and respond positively to cocaine, these stories may not have been adequately representative of personal drug use habits to be effective for all subjects. Finally, subjects only heard the evocative cocaine script twice during each PET session, reminding them to orient to the cocaine cues as if they were experiencing them. It is possible that 45-min intervals between reminders may have limited craving responses if subjects stopped personalizing the cocaine cues. In the end, individual differences may underscore the view that drug abuse is not solely due to the pharmacological effects of a drug, but involves biological and psychological factors that may vary considerably across individuals.
An additional limitation of this study may be the statistical power of tests for laterality. We powered this study to examine the comparison between cravers and non-cravers and correlations with the craving score, and not laterality differences. It is conceivable that a larger sample size would reveal the bilateral nature of the change if present. We specifically used two different parametric mapping approaches to address issues of power and statistics. Indeed, the LSRTM parametric approach of obtaining the BP maps showed small but significant clusters in the right putamen as well as in the left putamen, suggesting that the effect is bilateral. With greater power, it is possible that the peak in the right putamen seen with the LSRTM approach would be statistically significant. This may also be true for the group differences in DAR occupancy. With greater power, it is possible that the peak in the right putamen seen with the LSRTM approach would be statistically significant. This may also be true for the group differences and correlation of craving with DAR occupancy consistent with a bilateral effect. However the ultimate evidence of laterality is more appropriate for future studies with larger samples but does not weaken the evidence of the unilateral findings reported here.
CONCLUSIONS
The present study suggests that changes in DAR occupancy can provide a surrogate marker of drug craving. In addition, the methodology used in this study to measure binding potential changes in dopaminergic systems during cocaine craving may prove useful in investigations of other motivational states linked to dopamine, including reward associated with addiction to drugs (Wexler et al, 2001), food (Wang et al, 2001, 2002, 2004; Wise, 2004), sex (Bocher et al, 2001; Mouras et al, 2003; Redouté et al, 2000), alcohol (Grüsser et al, 2004; Heinz et al, 2004), and gambling (Hollander et al, 2005; Reuter et al, 2005; Crockford et al, 2005).
The relationship of cue-induced cocaine craving and increased DAR occupancy extends our knowledge of the mechanisms by which environmental stimuli can contribute to addictive disorders. Our findings demonstrate the importance of dorsal striatal dopamine systems in automated (habitual) behaviors (Berke and Hyman, 2000; Schultz, 2001). We have also presented the cue-induced changes in DAR occupancy within the context of the tonic-phasic dopamine system and discussed how this system may be disrupted through drug use. Given the prior evidence that responsivity to cocaine cues can outlast withdrawal symptoms and can occur despite abstinence from drugs (O'Brien et al, 1992), an understanding of the mechanism of the cue-induced craving is highly relevant to treatment for stimulant dependence.
References
Alexander GE, Crutcher MD, DeLong MR (1990). Basal ganglia-thalamocortical circuits: parallel substrates for motor, oculomotor, ‘prefrontal’ and ‘limbic’ functions. Prog Brain Res 85: 119–146.
Amara SG, Sonders MS (1998). Neurotransmitter transporters as molecular targets for addictive drugs. Drug Alcohol Depend 51: 87–96.
Avants SK, Margolin A, Kosten TR, Cooney NL (1995). Differences between responders and nonresponders to cocaine cues in the laboratory. Addict Behav 20: 215–224.
Bartels A, Zeki S (2000). The neural basis of romantic love. NeuroReport 11: 3829–3834.
Baumann B, Danos P, Krell D, Diekmann S, Leschinger A, Stauch R et al (1999). Reduced volume of limbic system-affiliated basal ganglia in mood disorders: preliminary data from a postmortem study. J Neuropsychiatry Clin Neurosci 11: 71–78.
Berger SP, Hall S, Mickalian JD, Reid MS, Crawford CA, Delucchi K et al (1996). Haloperidol antagonism of cue-elicited cocaine craving. Lancet 347: 504–508.
Berke JD (2003). Learning and memory mechanisms involved in compulsive drug use and relapse. In: Wang J (ed). Drugs of Abuse: Analysis of Neurological Effects. Humana Press: Totowa.
Berke JD, Hyman SE (2000). Addiction, dopamine, and the molecular mechanisms of memory. Neuron 25: 515–532.
Bocher M, Chisin R, Parag Y, Freedman N, Meir Weil Y, Lester H et al (2001). Cerebral activation associated with sexual arousal in response to a pornographic clip: A 15O-H2O PET study in heterosexual men. Neuroimage 14: 105–117.
Bonson KR, Grant SJ, Contoreggi CS, Links JM, Metcalfe J, Weyl HL et al (2002). Neural systems and cue-induced cocaine craving. Neuropsychopharmacology 26: 376–386.
Breiter HC, Rosen BR (1999). Functional magnetic resonance imaging of brain reward circuitry in the human. Ann NY Acad Sci 877: 523–547.
Buchert R, Varga J, Mester J (2004). Limitations of bi-linear regression analysis for the determination of receptor occupancy with positron emission tomography. Nucl Med Commun 25: 451–459.
Cannon CM, Bseikri MR (2004). Is dopamine required for natural reward? Physiol Behav 81: 741–748.
Childress AR, Mozley PD, McElgin W, Fitzgerald J, Reivich M, O'Brien CP (1999). Limbic activation during cue-induced cocaine craving. Am J Psychiatry 156: 11–18.
Collignon A, Maas LC, Delaere P, Vandermeulen D, Suetens P, Marchal G (1995). Automated multi-modality image registration based on information theory. In: Bizais Y, Di Paola R (eds). Proc. Information Processing. Kluwer Academic Publishers: Dordrecht, The Netherlands.
Crockford DN, Goodyear B, Edwards J, Quickfall J, el-Guebaly N (2005). Cue-induced brain activity in pathological gamblers. Biol Psychiatry 58: 787–795.
Dolan RJ, Fletcher P, Frith CD, Friston KJ, Frackowiak RS, Grasby PM (1995). Dopaminergic modulation of impaired cognitive activation in the anterior cingulate cortex in schizophrenia. Nature 378: 180–182.
Ehrin E, Farde L, de Paulis T, Eriksson L, Greitz T, Johnström P et al (1985). Preparation of 11C-labelled Raclopride, a new potent dopamine receptor antagonist: preliminary PET studies of cerebral dopamine receptors in the monkey. Int J Appl Radiat Isot 36: 269–273.
Faure A, Haberland U, Condé F, El Massioui N (2005). Lesion to the nigrostriatal dopamine system disrupts stimulus-response habit formation. J Neurosci 25: 2771–2780.
Fiorillo CD, Tobler PN, Schultz W (2003). Discrete coding of reward probability and uncertainty by dopamine neurons. Science 299: 1898–1902.
Floresco SB, West AR, Ash B, Moore H, Grace AA (2003). Afferent modulation of dopamine neuron firing differentially regulates tonic and phasic dopamine transmission. Nat Neurosci 6: 968–973.
Franken IHA (2003). Drug craving and addiction: integrating psychological and neuropsychopharmacological approaches. Prog Neuropsychopharmacol Biol Psychiatry 27: 563–579.
Friston KJ, Frith CD, Liddle PF, Frackowiak RSJ (1991). Comparing functional (PET) images: The assessment of significant change. J Cereb Blood Flow Metab 11: 690–699.
Garavan H, Pankiewicz J, Bloom A, Cho JK, Sperry L, Ross TJ et al (2000). Cue-induced cocaine craving: neuroanatomical specificity for drug users and drug stimuli. Am J Psychiatry 157: 1789–1798.
Gerdeman GL, Partridge JG, Lupica CR, Lovinger DM (2003). It could be habit forming: drugs of abuse and striatal synaptic plasticity. Trends Neurosci 26: 184–192.
Gerfen CR, Wilson CJ (1996). The basal ganglia. In: Swanson LW, Björklund A, Hökfelt T (eds). Integrated Systems of the CNS, Part III. Elsevier: New York. pp 371–468.
Gjedde A, Wong DF, Rosa-Neto P, Cumming P (2005). Mapping neuroreceptors at work: on the definition and interpretation of binding potentials after 20 years of progress. Int Rev Neurobiol 63: 1–20.
Goto Y, Grace AA (2005). Dopamine-dependent interactions between limbic and prefrontal cortical plasticity in the nucleus accumbens: disruption by cocaine sensitization. Neuron 47: 255–266.
Grant S, London ED, Newlin DB, Villemagne VL, Liu X, Contoreggi C et al (1996). Activation of memory circuits during cue-elicited cocaine craving. Proc Natl Acad Sci USA 93: 12040–12045.
Grüsser SM, Wrase J, Klein S, Hermann D, Smolka MN, Ruf M et al (2004). Cue-induced activation of the striatum and medial prefrontal cortex is associated with subsequent relapse in abstinent alcoholics. Psychopharmacology (Berlin) 175: 296–302.
Haber SN, Fudge JL, McFarland NR (2000). Striatonigrostriatal pathways in primates form an ascending spiral from the shell to the dorsolateral striatum. J Neurosci 20: 2369–2382.
Harlan RE, Garcia MM (1998). Drugs of abuse and immediate-early genes in the forebrain. Mol Neurobiol 16: 221–267.
Heinz A, Siessmeier T, Wrase J, Hermann D, Klein S, Grüsser-Sinopoli SM et al (2004). Correlation between dopamine D2 receptors in the ventral striatum and central processing of alcohol cues and craving. Am J Psychiatry 161: 1783–1789.
Hollander E, Pallanti S, Balcini-Rossi N, Sood E, Baker BR, Buchsbaum MS (2005). Imaging monetary reward in pathological gamblers. World J Biol Psychiatry 6: 113–120.
Ichise M, Liow JS, Lu JQ, Takano A, Model K, Toyama H et al (2003). Linearized reference tissue parametric imaging methods: application to [11C]DASB positron emission tomography studies of the serotonin transporter in human brain. J Cereb Blood Flow Metab 23: 1096–1112.
Ichise M, Toyama H, Innis RB, Carson RE (2002). Strategies to improve neuroreceptor parameter estimation by linear regression analysis. J Cereb Blood Flow Metab 22: 1271–1281.
Ito R, Dalley JW, Robbins TW, Everitt BJ (2002). Dopamine release in the dorsal striatum during cocaine-seeking behavior under the control of a drug-associated cue. J Neurosci 22: 6247–6253.
Kilts CD, Gross RE, Ely TD, Drexler KPG (2004). The neural correlates of cue-induced craving in cocaine-dependent women. Am J Psychiatry 161: 233–241.
Kilts CD, Schweitzer JB, Quinn CK, Gross RE, Faber TL, Muhammad F et al (2001). Neural activity related to drug craving in cocaine addiction. Arch Gen Psychiatry 58: 334–341.
Knoblich G, Curtis D, Faustman WO, Zarcone V, Stewart S, Mefford I et al (1992). Increased CSF HVA with craving in long-term abstinent cocaine abusers. Biol Psychiatry 32: 96–100.
Koepp MJ, Gunn RN, Lawrence AD, Cunningham VJ, Dagher A, Jones T et al (1998). Evidence for striatal dopamine release during a video game. Nature 393: 266–268.
Lammertsma AA, Hume SP (1996). Simplified reference tissue model for PET receptor studies. Neuroimage 4 (3 Part 1): 153–158.
Lin KP, Huang SC, Yu DC, Melega W, Barrio JR, Phelps ME (1996). Automated image registration for FDOPA PET studies. Phys Med Biol 41: 2775–2788.
Logan J, Fowler JS, Volkow ND, Wang GJ, Ding YS, Alexoff DL (1996). Distribution volume ratios without blood sampling from graphical analysis of PET data. J Cereb Blood Flow Metab 16: 834–840.
Logan J, Fowler JS, Volkow ND, Wolf AP, Dewey SL, Schlyer DJ et al (1990). Graphical analysis of reversible radioligand binding from time-activity measurements applied to [N-11C-methyl]-(−)-cocaine PET studies in human subjects. J Cereb Blood Flow Metab 10: 740–747.
London ED, Cascella NG, Wong DF, Phillips RL, Dannals RF, Links JM et al (1990). Cocaine-induced reduction of glucose utilization in human brain. A study using positron emission tomography and [fluorine 18]-fluorodeoxyglucose. Arch Gen Psychiatry 47: 567–574.
Maas LC, Lukas SE, Kaufman MJ, Weiss RD, Daniels SL, Rogers VW et al (1998). Functional magnetic resonance imaging of human brain activation during cue-induced cocaine craving. Am J Psychiatry 155: 124–126.
Martin SD, Yeragani VK, Lodhi R, Galloway MP (1989). Clinical ratings and plasma HVA during cocaine abstinence. Biol Psychiatry 26: 356–362.
Martinez D, Slifstein M, Broft A, Mawlawi O, Hwang DR, Huang Y et al (2003). Imaging human mesolimbic dopamine transmission with positron emission tomography. Part II: amphetamine-induced dopamine release in the functional subdivisions of the striatum. J Cereb Blood Flow Metab 23: 285–300.
McDonald RJ, White NM (1993). A triple dissociation of memory systems: hippocampus, amygdala, and dorsal striatum. Behav Neurosci 107: 3–22.
Meyer JH, Gunn RN, Myers R, Grasby PM (1999). Assessment of spatial normalization of PET ligand images using ligand-specific templates. Neuroimage 9: 545–553.
Mintun MA, Raichle ME, Kilbourn MR, Wooten GF, Welch MJ (1984). A quantitative model for the in vivo assessment of drug binding sites with positron emission tomography. Ann Neurol 15: 217–227.
Mouras H, Stoléru S, Bittoun J, Glutron D, Pélégrini-Issac M, Paradis AL et al (2003). Brain processing of visual sexual stimuli in healthy men: a functional magnetic resonance imaging study. Neuroimage 20: 855–869.
Newlin DB (1992). A comparison of drug conditioning and craving for alcohol and cocaine. Recent Dev Alcohol 10: 147–164.
O'Brien CP, Childress AR, McLellan AT, Ehrman R (1992). Classical conditioning in drug-dependent humans. Ann NY Acad Sci 654: 400–415.
O'Brien CP, Childress AR, McLellan T, Ehrman R (1990). Integrating systemic cue exposure with standard treatment in recovering drug dependent patients. Addict Behav 15: 355–365.
Oswald LM, Wong DF, McCaul M, Zhou Y, Kuwabara H, Choi L et al (2005). Relationships among ventral striatal dopamine release, cortisol secretion, and subjective responses to amphetamine. Neuropsychopharmacology 30: 821–832.
Porrino LJ, Lyons D, Smith HR, Daunais JB, Nader MA (2004). Cocaine self-administration produces a progressive involvement of limbic, association, and sensorimotor striatal domains. J Neurosci 24: 3554–3562.
Redouté J, Stoléru S, Grégoire MC, Costes N, Cinotti L, Lavenne F et al (2000). Brain processing of visual sexual stimuli in human males. Hum Brain Mapp 11: 162–177.
Reuter J, Raedler T, Rose M, Hand I, Glascher J, Buchel C (2005). Pathological gambling is linked to reduced activation of the mesolimbic reward system. Nat Neurosci 8: 147–148.
Risinger RC, Salmeron BJ, Ross TJ, Amen SL, Sanfilipo M, Hoffmann RG et al (2005). Neural correlates of high and craving during cocaine self-administration using BOLD fMRI. Neuroimage 26: 1097–1108.
Robbins TW, Everitt BJ (2002). Limbic-striatal memory systems and drug addiction. Neurobiol Learn Mem 78: 625–636.
Rohsenow DJ, Niaura RS, Childress AR, Abrams DB, Monti PM (1990). Cue reactivity in addictive behaviors: theoretical and treatment implications. Int J Addict 25: 957–993.
Roy A, Berman J, Williams R, Kuhn C, Gonzalez B (2002). Higher levels of CSF homovanillic acid in recently abstinent cocaine-dependent patients. Am J Psychiatry 159: 1053–1055.
Saka E, Goodrich C, Harlan P, Madras BK, Graybiel AM (2004). Repetitive behaviors in monkeys are linked to specific striatal activation patterns. J Neurosci 24: 7557–7565.
Schiltz CA (2006). Habitual responding and the dorsal striatum. J Neurosci 26: 1891–1892.
Schultz W (2001). Reward signaling by dopamine neurons. Neuroscientist 7: 293–302.
Sherer MA, Kumor KM, Jaffe JH (1989). Effects of intravenous cocaine are partially attenuated by haloperidol. Psychiatry Res 27: 117–125.
Talairach J, Tournoux P (1988). Co-Planar Stereotaxic Atlas of the Human Brain 3-Dimensional Proportional System: An Approach To Cerebral Imaging. G. Thieme: Stuttgart.
Udo de Haes JI, Kortekaas R, Van Waarde A, Maguire RP, Pruim J, den Boer JA (2005). Assessment of methylphenidate-induced changes in binding of continuously infused [11C]-raclopride in healthy human subjects: correlation with subjective effects. Psychopharmacology (Berlin) 183: 322–330.
Vanderschuren LJ, Di Ciano P, Everitt BJ (2005). Involvement of the dorsal striatum in cue-controlled cocaine seeking. J Neurosci 25: 8665–8670.
Volkow ND, Wang GJ, Telang F, Fowler JS, Logan J, Childress AR et al (2006). Cocaine cues and dopamine in dorsal striatum: mechanism of craving in cocaine addiction. J Neurosci 26: 6583–6588.
Volkow ND, Fowler JS, Wang GJ (2004). The addicted human brain viewed in the light of imaging studies: brain circuits and treatment strategies. Neuropharmacology 47 (Suppl 1): 3–13.
Wang GJ, Volkow ND, Felder C, Fowler JS, Levy AV, Pappas NR et al (2002). Enhanced resting activity of the oral somatosensory cortex in obese subjects. NeuroReport 13: 1151–1155.
Wang GJ, Volkow ND, Logan J, Pappas NR, Wong CT, Zhu W et al (2001). Brain dopamine and obesity. Lancet 357: 354–357.
Wang GJ, Volkow ND, Thanos PK, Fowler JS (2004). Similarity between obesity and drug addiction as assessed by neurofunctional imaging: a concept review. J Addict Dis 23: 39–53.
West AR, Grace AA (2002). Opposite influences of endogenous dopamine D1 and D2 receptor activation on activity states and electrophysiological properties of striatal neurons: studies combining in vivo intracellular recordings and reverse microdialysis. J Neurosci 22: 294–304.
Wexler BE, Gottschalk CH, Fulbright RK, Prohovnik I, Lacadie CM, Rounsaville BJ et al (2001). Functional magnetic resonance imaging of cocaine craving. Am J Psychiatry 158: 86–95.
Wise RA (2004). Dopamine and food reward: back to the elements. Am J Physiol Regul Integr Comp Physiol 286: R13.
Wu Y, Carson RE (2002). Noise reduction in the simplified reference tissue model for neuroreceptor functional imaging. J Cereb Blood Flow Metab 22: 1440–1452.
Worsley KJ, Evans AC, Marrett S, Neelin P (1992). A three-dimensional statistical analysis for CBF activation studies in human brain. J Cereb Blood Flow Metab 12: 900–918.
Yin HH, Knowlton BJ, Balleine BW (2004). Lesions of dorsolateral striatum preserve outcome expectancy but disrupt habit formation in instrumental learning. Eur J Neurosci 19: 181–189.
Zhou Y, Endres CJ, Brasic JR, Huang SC, Wong DF (2003). Linear regression with spatial constraint to generate parametric images of ligand-receptor dynamic PET studies with a simplified reference tissue model. Neuroimage 18: 975–989.
Acknowledgements
Grant support: PHS Grant NIH K24DA00412, DA11080, AA12839, NS 38927 to DW, Danish National Science Foundation Center of Excellence grant to AG. Special Thanks to L Sims, M Thomas, B Gay, A Crabb, RF Dannals, H Ravert, W Mathews, M Pomper, and S Tata for scientific and technical assistance.
Author information
Authors and Affiliations
Corresponding author
Additional information
NOTE ADDED IN PROOF
Findings similar to those reported here and in our preliminary results (see below) were recently reported by Volkow et al (2006).
Preliminary findings from this study were initially presented as abstracts in 2003 and 2004, cited below:
1. 50th Annual Meeting, Society for Nuclear Medicine, New Orleans, Louisiana, June 21–25, 2003.
Citation: DF Wong, JS Lee, A Maini, Y Zhou, H Kuwabara, C Endres, J Brasic, AS Dogan, D Schretlen, M Alexander, E Kimes, M Ernst, D Jasinski, ED London, S Zukin. Cue induced cocaine craving and dopamine release: Methodology and correlates. Journal of Nuclear Medicine 44(5):67P, 2003.
2. 65th Annual Meeting, College on problems of Drug Dependence, Bal Harbour, Florida, June 14–19, 2003.
Citation: DF Wong, JS Lee, Y Zhou, J Brasic, A Maini, H Kuwabara, AS Kimes, C Contoreggi, M Ernst, D Schretlen, D Jasinski, S Zukin, K Bonson and ED London. Intrasynaptic Dopamine Release and Cocaine Craving Induced by Video/Audio Cues. Abstracts, College on Problems of Drug Dependence, 65th Annual Meeting, #751. (2003)
3. 33rd Annual Meeting, Society for Neuroscience, New Orleans, Louisiana, November 8–12, 2003.
Citation: DF Wong, JS Lee, H Kuwabara, Y Zhou, J Brasic, A Maini, AS Kimes, C Contoreggi, M Ernst, D Schretlen, D Jasinski, S Zukin, ED London. Cocaine craving induced by video/audio cues and intrasynaptic dopamine release. Abstracts, Society for Neuroscience, Program No. 879.8. (2003)
4. 66th Annual Meeting, College for Problems on Drug Dependence, San Juan, Puerto Rico, June 12–17, 2004.
Citation: DF Wong, H Kuwabara, W Ye, A Kumar, Y Zhou, M Alexander, J Brasic, M Thomas, MA Maris, D Schretlen, ED London, R Jasinski. Cocaine craving correlates with psychostimulant-induced dopamine release and dopamine transporters. Abstracts, College on Problems of Drug Dependence, 66th Annual Meeting. (2004)
Rights and permissions
About this article
Cite this article
Wong, D., Kuwabara, H., Schretlen, D. et al. Increased Occupancy of Dopamine Receptors in Human Striatum during Cue-Elicited Cocaine Craving. Neuropsychopharmacol 31, 2716–2727 (2006). https://doi.org/10.1038/sj.npp.1301194
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/sj.npp.1301194
Keywords
This article is cited by
-
Acute and protracted abstinence from methamphetamine bidirectionally changes intrinsic excitability of indirect pathway spiny projection neurons in the dorsomedial striatum
Scientific Reports (2022)
-
Disruption of prepulse inhibition is associated with compulsive behavior severity and nucleus accumbens dopamine receptor changes in Sapap3 knockout mice
Scientific Reports (2021)
-
The influence of conditioned stimuli on [11C]-(+)-PHNO PET binding in tobacco smokers after a one week abstinence
Scientific Reports (2021)
-
Translational study of the whole transcriptome in rats and genetic polymorphisms in humans identifies LRP1B and VPS13A as key genes involved in tolerance to cocaine-induced motor disturbances
Translational Psychiatry (2020)
-
Effects of long-term cocaine self-administration on brain resting-state functional connectivity in nonhuman primates
Translational Psychiatry (2020)
