3’-O-Methylorobol Inhibits the Voltage-Gated Sodium Channel Nav1.7 with Anti-Itch Efficacy in A Histamine-Dependent Itch Mouse Model


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
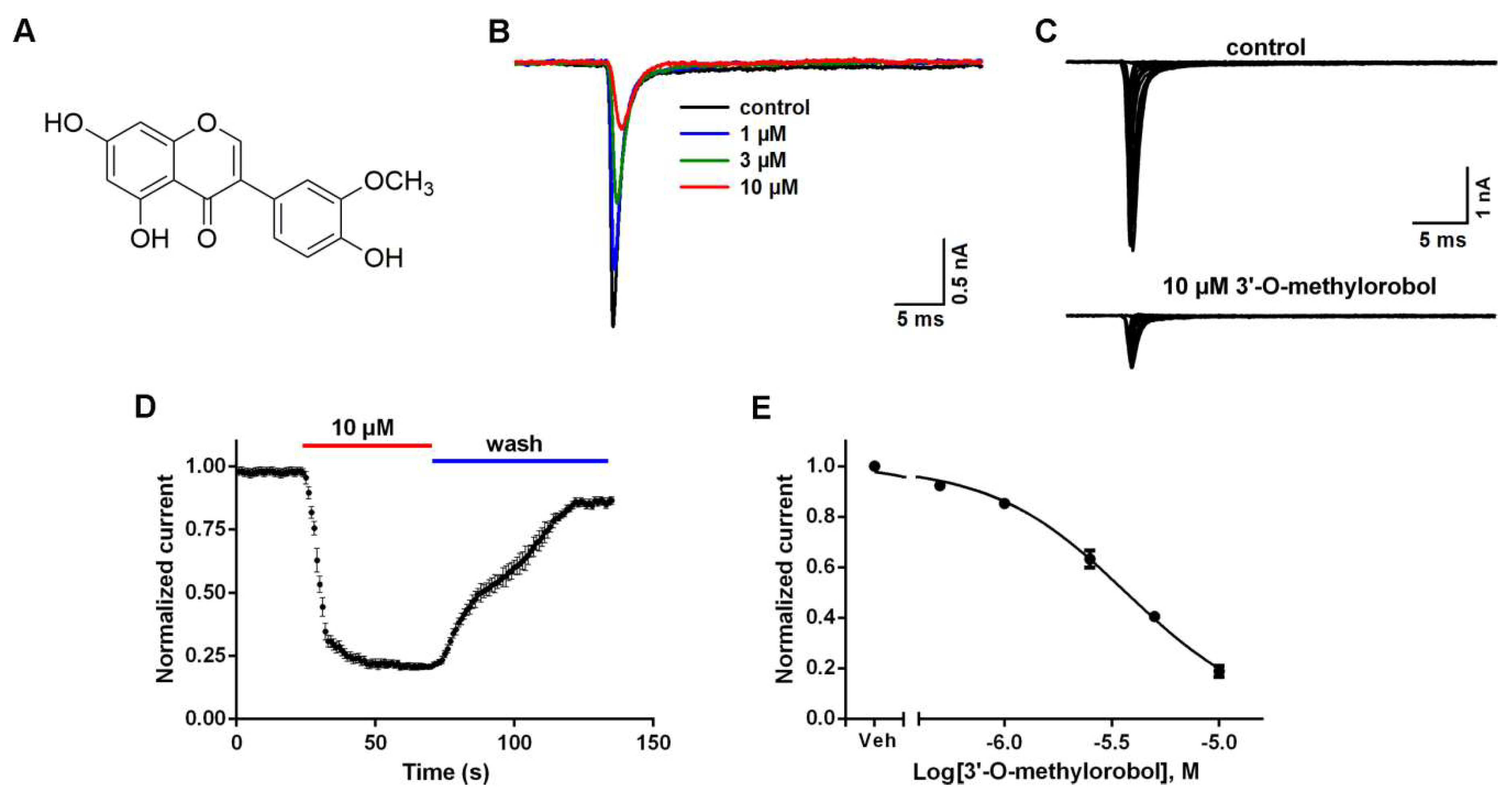
2.1. Inhibitory Effects of 3’-O-Methylorobol on Nav1.7 Stably Expressed in CHO Cells
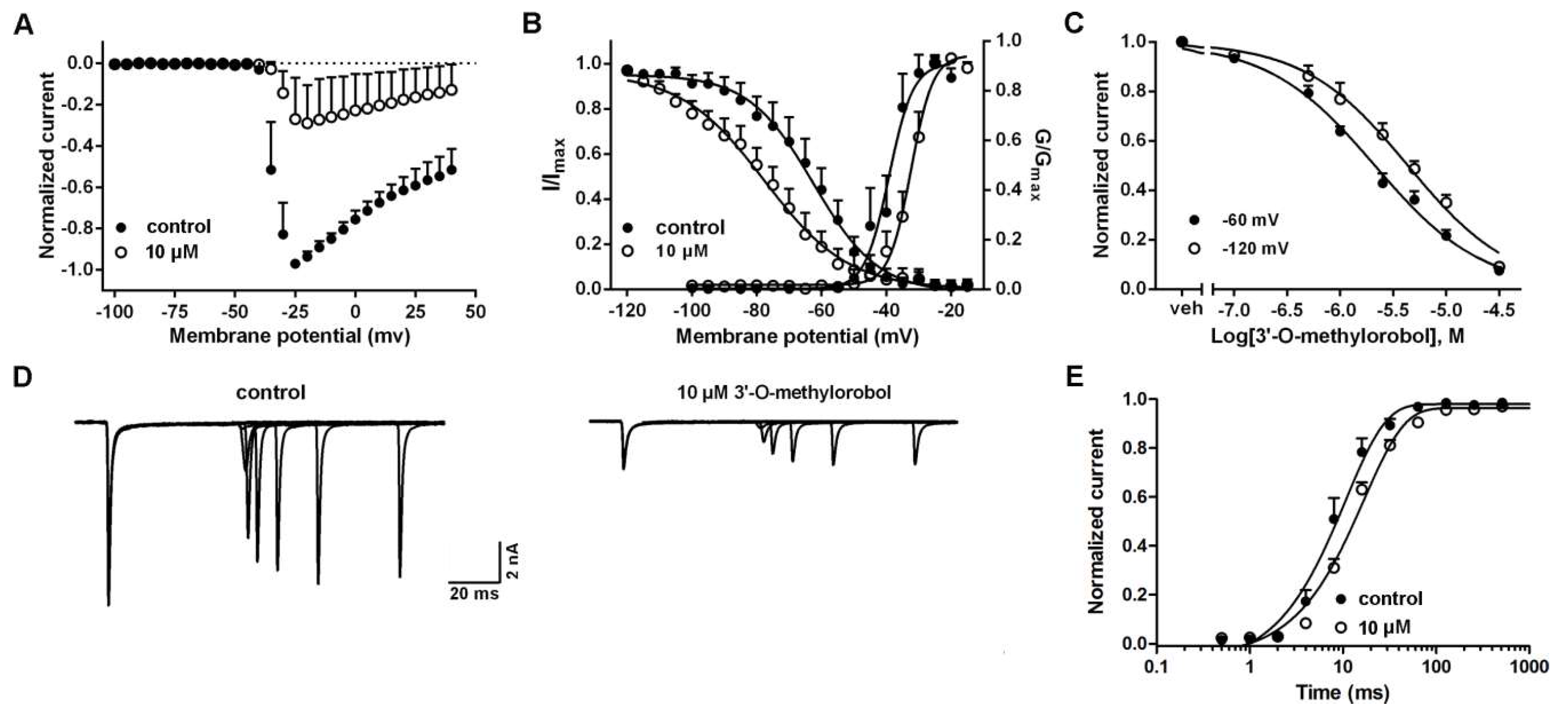
2.2. Influences of 3’-O-Methylorobol on the Channel Kinetics of Nav1.7 Stably Expressed in CHO Cells
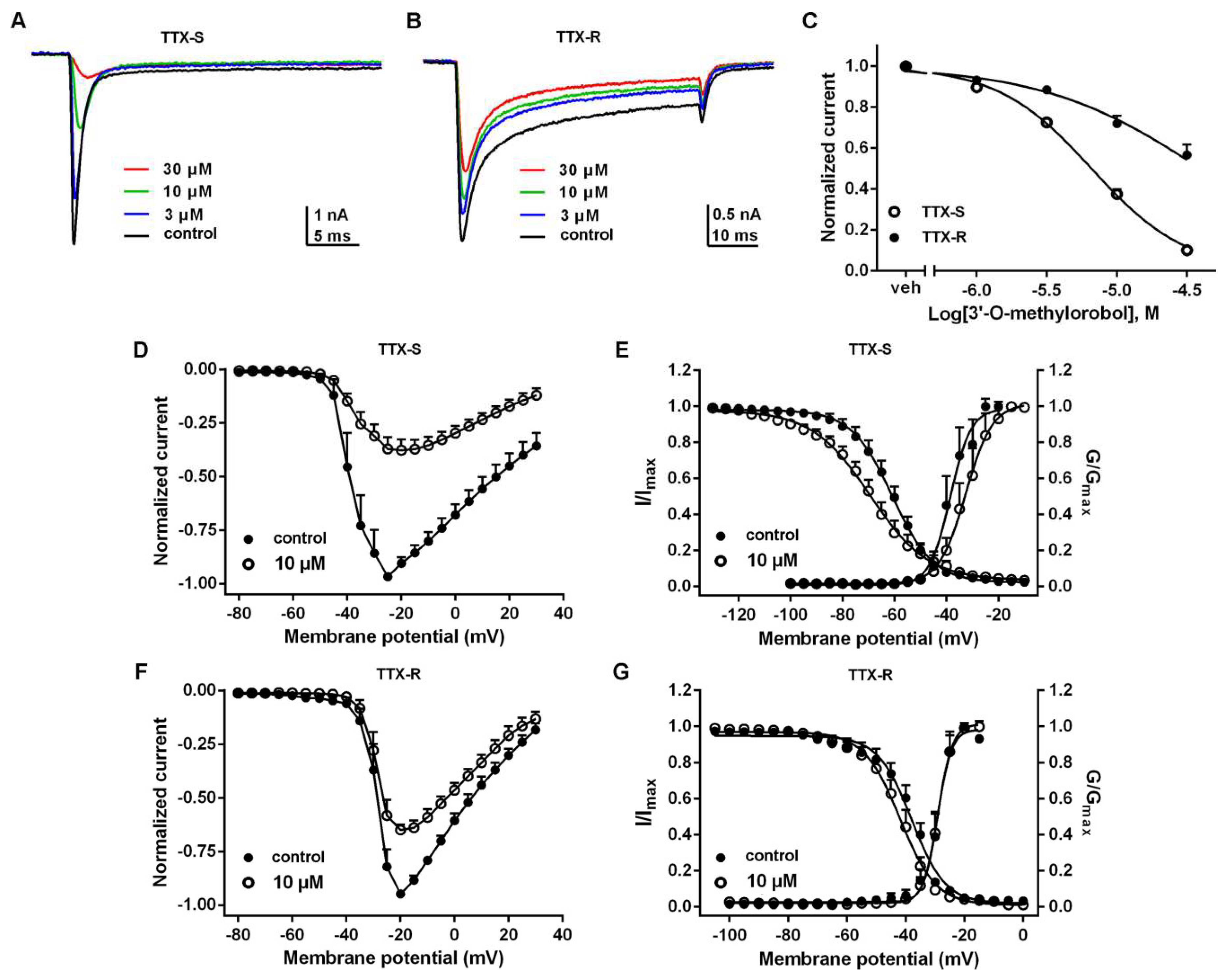
2.3. Effects of 3’-O-Methylorobol on TTX-S and TTX-R Na+ Currents in DRG Neurons
2.4. Effects of 3’-O-Methylorobol on Action Potentials Firing in Primary Cultured DRG Neurons
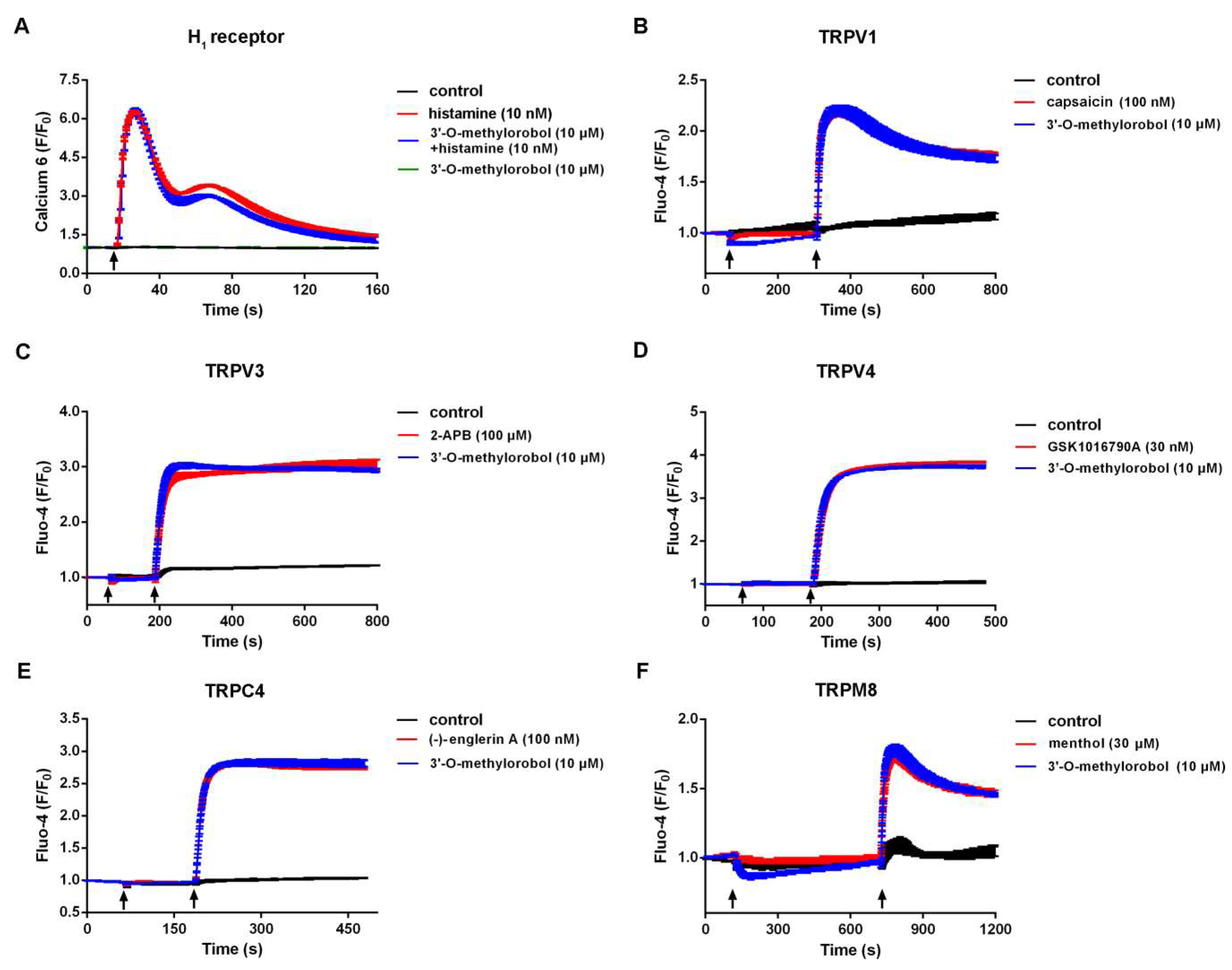
2.5. Effect of 3’-O-Methylorobol on Histamine-Dependent Itch Receptor and Ion Channels
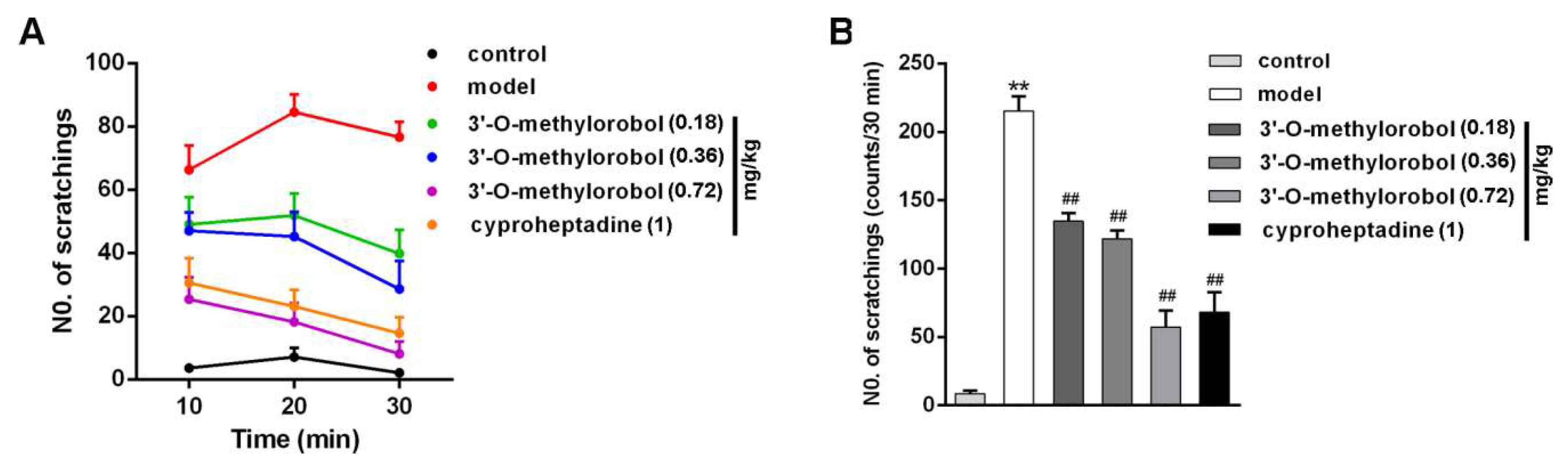
2.6. 3’-O-Methylorobol Ameliorates Itch in Mouse Model of Histamine-Dependent Itch Induced by Compound 48/80
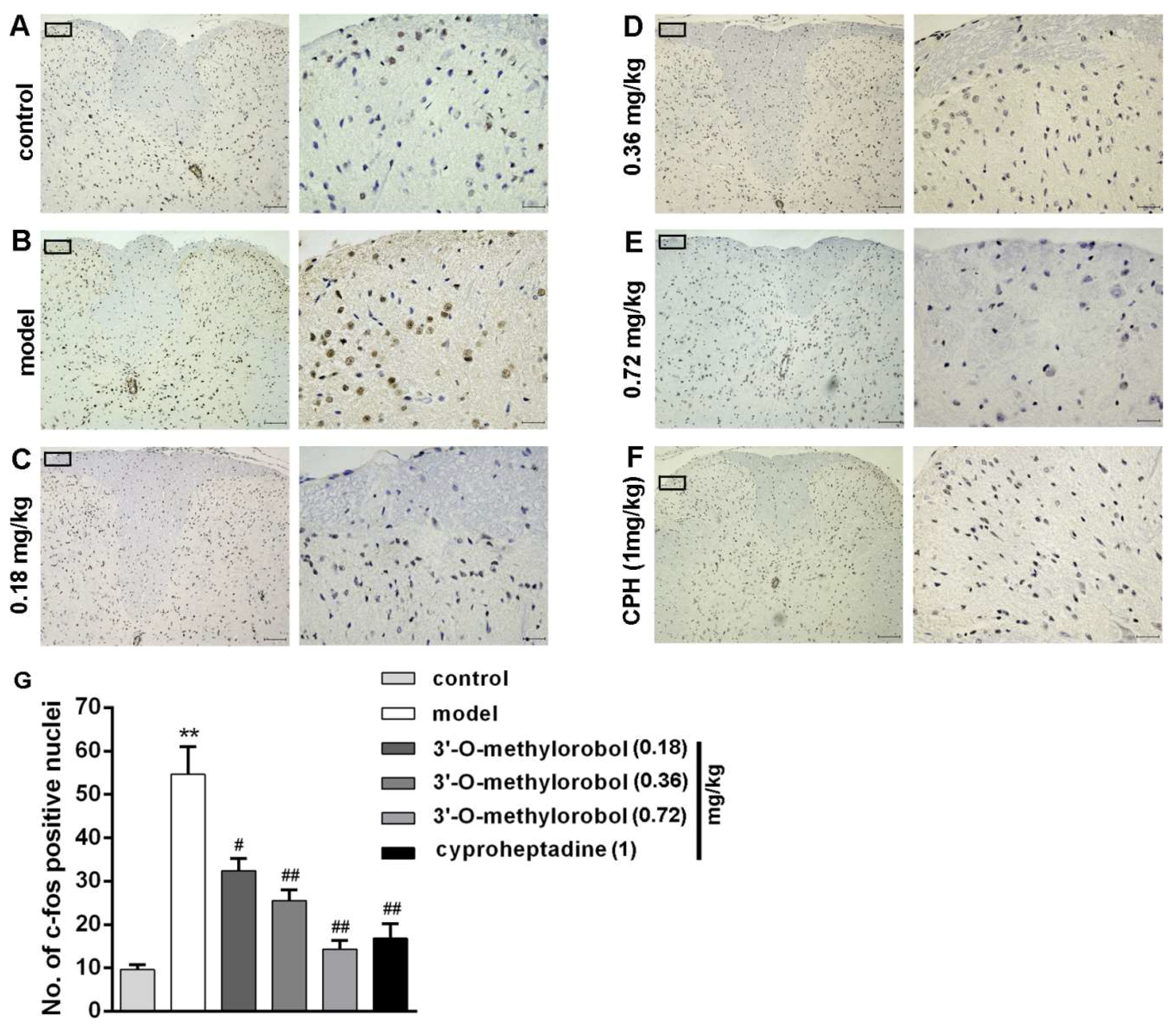
2.7. Effect of 3’-O-Methylorobol on Itch-Evoked C-fos Expression
3. Discussion
4. Materials and Methods
4.1. Materials and Animals
4.2. Cultures of CHO, HEK293, and ND7/23 Cells
4.3. Primary Cultures of Dorsal Root Ganglion Neurons
4.4. Whole-Cell Voltage-Clamp Electrophysiology
4.5. Intracellular Ca2+ Concentration Determination
4.6. Compound 48/80-Induced Histamine-Dependent Itch Model
4.7. C-fos Immunohistochemistry
4.8. Data Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| APs | Action potentials |
| CI | Confidence interval |
| DRG | Dorsal root ganglion |
| FLIPR | Fluorescence imaging plate reader |
| IC50 | Half-maximal inhibitory concentration |
| I–V | Current–voltage |
| PDL | Poly-d-lysine |
| PLL | Poly-l-lysine |
| TRP | Transient receptor potential |
| TTX-R | Tetrodotoxin-resistant |
| TTX-S | Tetrodotoxin-sensitive |
| V1/2 | Half-maximal voltage |
| VGSC | Voltage-gated sodium channel |
References
- Davidson, S.; Giesler, G.J. The multiple pathways for itch and their interactions with pain. Trends Neurosci. 2010, 33, 550–558. [Google Scholar] [CrossRef] [PubMed]
- Bautista, D.M.; Wilson, S.R.; Hoon, M.A. Why we scratch an itch: The molecules, cells and circuits of itch. Nat. Neurosci. 2014, 17, 175–182. [Google Scholar] [CrossRef] [PubMed]
- Tarikci, N.; Kocatürk, E.; Güngör, Ş.; Topal, I.O.; Can, P.Ü.; Singer, R. Pruritus in systemic diseases: A review of etiological factors and new treatment modalities. Sci. World J. 2015, 2015, 803752. [Google Scholar] [CrossRef] [PubMed]
- LaMotte, R.H.; Dong, X.; Ringkamp, M. Sensory neurons and circuits mediating itch. Nat. Rev. Neurosci. 2014, 15, 19–31. [Google Scholar] [CrossRef]
- Roberson, D.P.; Gudes, S.; Sprague, J.M.; Patoski, H.A.; Robson, V.K.; Blasl, F.; Duan, B.; Oh, S.B.; Bean, B.P.; Ma, Q.; et al. Activity-dependent silencing reveals functionally distinct itch-generating sensory neurons. Nat. Neurosci. 2013, 16, 910–918. [Google Scholar] [CrossRef]
- Green, D.; Dong, X. The cell biology of acute itch. J. Cell Biol. 2016, 213, 155–161. [Google Scholar] [CrossRef]
- Imamachi, N.; Park, G.H.; Lee, H.; Anderson, D.J.; Simon, M.I.; Basbaum, A.I.; Han, S.K. TRPV1-expressing primary afferents generate behavioral responses to pruritogens via multiple mechanisms. Proc. Natl. Acad. Sci. USA 2009, 106, 11330–11335. [Google Scholar] [CrossRef]
- Palkar, R.; Ongun, S.; Catich, E.; Li, N.; Borad, N.; Sarkisian, A.; McKemy, D.D. Cooling relief of acute and chronic itch requires TRPM8 channels and neurons. J. Invest. Dermatol. 2018, 138, 1391–1399. [Google Scholar] [CrossRef]
- Yamamoto-Kasai, E.; Imura, K.; Yasui, K.; Shichijou, M.; Oshima, I.; Hirasawa, T.; Sakata, T.; Yoshioka, T. TRPV3 as a therapeutic target for itch. J. Invest. Dermatol. 2012, 132, 2109–2112. [Google Scholar] [CrossRef]
- Chen, Y.; Fang, Q.; Wang, Z.; Zhang, J.Y.; MacLeod, A.S.; Hall, R.P.; Liedtke, W.B. Transient receptor potential vanilloid 4 ion channel functions as a pruriceptor in epidermal keratinocytes to evoke histaminergic itch. J. Biol. Chem. 2016, 291, 10252–10262. [Google Scholar] [CrossRef]
- Lee, Y.W.; Won, C.H.; Jung, K.; Nam, H.J.; Choi, G.; Park, Y.H.; Park, M.; Kim, B. Efficacy and safety of PAC-14028 cream - a novel, topical, nonsteroidal, selective TRPV1 antagonist in patients with mild-to-moderate atopic dermatitis: A phase II b randomized trial. Br. J. Dermatol. 2019, 180, 1030–1038. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Sun, X.; Qi, H.; Ma, Q.; Zhou, Q.; Wang, W.; Wang, K. Pharmacological inhibition of temperature-sensitive and Ca2+-permeable TRPV3 channel by a natural forsythoside B attenuates pruritus and cytotoxicity of keratinocytes. J. Pharmacol. Exp. Ther. 2018, 368, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.Y.; Sun, L.L.; Qi, H.; Gao, Q.; Wang, G.X.; Wei, N.N.; Wang, K. Antipruritic effect of natural coumarin osthole through selective inhibition of thermosensitive TRPV3 channel in the skin. Mol. Pharmacol. 2018, 94, 1164–1173. [Google Scholar] [CrossRef] [PubMed]
- Chew, L.A.; Bellampalli, S.S.; Dustrude, E.T.; Khanna, R. Mining the Nav1.7 interactome: Opportunities for chronic pain therapeutics. Biochem. Pharmacol. 2019, 163, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Devigili, G.; Eleopra, R.; Pierro, T.; Lombardi, R.; Rinaldo, S.; Lettieri, C.; Faber, C.G.; Merkies, I.S.; Waxman, S.G.; Lauria, G. Paroxysmal itch caused by gain-of-function Nav1. 7 mutation. Pain 2014, 155, 1702–1707. [Google Scholar] [CrossRef] [PubMed]
- Snyder, L.M.; Ross, S.E.; Belfer, I. An SCN9A variant, known to cause pain, is now found to cause itch. Pain 2014, 155, 1677–1678. [Google Scholar] [CrossRef]
- Lee, J.H.; Park, C.K.; Chen, G.; Han, Q.; Xie, R.G.; Liu, T.; Ji, R.R.; Lee, S.Y. A monoclonal antibody that targets a Nav1. 7 channel voltage sensor for pain and itch relief. Cell 2014, 157, 1393–1404. [Google Scholar] [CrossRef]
- Cheng, R.X.; Feng, Y.; Liu, D.; Wang, Z.H.; Zhang, J.T.; Chen, L.H.; Su, C.J.; Wang, B.; Huang, Y.; Ji, R.R.; et al. The role of Nav1.7 and methylglyoxal-mediated activation of TRPA1 in itch and hypoalgesia in a murine model of type 1 diabetes. Theranostics 2019, 9, 4287–4307. [Google Scholar] [CrossRef]
- Hoeck, E.A.; Marker, J.B.; Gazerani, P.H.; Andersen, H.; Arendt-Nielsen, L. Preclinical and human surrogate models of itch. Exp. Dermatol. 2016, 25, 750–757. [Google Scholar] [CrossRef]
- Xie, Z.; Hu, H. TRP channels as drug targets to relieve itch. Pharmaceuticals 2018, 11, 100. [Google Scholar] [CrossRef]
- Ahmad, S.; Riaz, N.; Saleem, M.; Jabbar, A.; Nisar-Ur-Rehman; Ashraf, M. Antioxidant flavonoids from Alhagi maurorum. J. Asian Nat. Prod. Res. 2010, 12, 138–143. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Yang, M.; Ma, X. Flavonoids from roots of Flemingia philippinensis. Zhongguo Zhong Yao Za Zhi 2009, 34, 724–726. [Google Scholar] [PubMed]
- Li, L.; Deng, X.; Zhang, L.; Shu, P.; Qin, M. A new coumestan with immunosuppressive activities from Flemingia philippinensis. Fitoterapia 2011, 82, 615–619. [Google Scholar] [CrossRef] [PubMed]
- Umehara, K.; Nemoto, K.; Kimijima, K.; Matsushita, A.; Terada, E.; Monthakantirat, O.; De-Eknamkul, W.; Miyase, T.; Warashina, T.; Degawa, M.; et al. Estrogenic constituents of the heartwood of Dalbergia parviflora. Phytochemistry 2008, 69, 546–552. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Xiang, C.; Liang, H.; Zhao, Y.Y. Chemical constituents of isoflavones from vine stems of Millettia nitita var. hirsutissima. Zhongguo Zhong Yao Za Zhi 2007, 32, 321–322. [Google Scholar] [PubMed]
- Zhang, S.; Cheng, J.; Chen, W.; Ling, X.; Zhao, Y.; Feng, J.; Xiang, C.; Liang, H. Interactions between thrombin and natural products of Millettia nitita var. hirsutissima using capillary zone electrophoresis. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2009, 877, 4107–4114. [Google Scholar] [CrossRef]
- Nkengfack, A.E.; Fomum, Z.T.; Ubillas, R.; Sanson, D.R.; Tempesta, M.S. Extractives from Erythrina eriotriocha. J. Nat. Prod. 1990, 53, 509–512. [Google Scholar] [CrossRef]
- Lee, M.K.; Ha, N.R.; Yang, H.; Sung, S.H.; Kim, Y.C. Stimulatory constituents of Eclipta prostrata on mouse osteoblast differentiation. Phytother. Res. 2009, 23, 129–131. [Google Scholar] [CrossRef]
- Ho, C.; O’Leary, M.E. Single-cell analysis of sodium channel expression in dorsal root ganglion neurons. Mol. Cell. Neurosci. 2011, 46, 159–166. [Google Scholar] [CrossRef]
- Zhang, F.; Xu, X.; Li, T.; Liu, Z. Shellfish toxins targeting voltage-gated sodium channels. Mar. Drugs 2013, 11, 4698–4723. [Google Scholar] [CrossRef]
- Wu, Y.; Ma, H.; Zhang, F.; Zhang, C.; Zou, X.; Cao, Z. Selective voltage-gated sodium channel peptide toxins from animal venom: Pharmacological probes and analgesic drug development. Acs Chem. Neurosci. 2018, 9, 187–197. [Google Scholar] [CrossRef] [PubMed]
- Bosmans, F.; Swartz, K.J. Targeting voltage sensors in sodium channels with spider toxins. Trends Pharmacol. Sci. 2010, 31, 175–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaji, N.; Little, M.J.; Nishio, H.; Billen, B.; Villegas, E.; Nishiuchi, Y.; Tytgat, J.; Nicholson, G.M.; Corzo, G. Synthesis, solution structure, and phylum selectivity of a spider δ-toxin that slows inactivation of specific voltage-gated sodium channel subtypes. J. Biol. Chem. 2009, 284, 24568–24582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corzo, G.; Sabo, J.K.; Bosmans, F.; Billen, B.; Villegas, E.; Tytgat, J.; Norton, R.S. Solution structure and alanine scan of a spider toxin that affects the activation of mammalian voltage-gated sodium channels. J. Biol. Chem. 2007, 282, 4643–4652. [Google Scholar] [CrossRef] [Green Version]
- Yang, N.N.; Shi, H.; Yu, G.; Wang, C.M.; Zhu, C.; Yang, Y.; Yuan, X.L.; Tang, M.; Wang, Z.L.; Gegen, T.; et al. Osthole inhibits histamine-dependent itch via modulating TRPV1 activity. Sci. Rep. 2016, 6, 25657. [Google Scholar] [CrossRef] [Green Version]
- Funahashi, H.; Naono-Nakayama, R.; Ebihara, K.; Koganemaru, G.; Kuramashi, A.; Ikeda, T.; Nishimori, T.; Ishida, Y. Hemokinin-1 mediates pruriceptive processing in the rat spinal cord. Neuroscience 2014, 277, 206–216. [Google Scholar] [CrossRef]
- Inan, S.; Dun, N.J.; Cowan, A. Nalfurafine prevents 5’-guanidinonaltrindole- and compound 48/80-induced spinal c-fos expression and attenuates 5’-guanidinonaltrindole-elicited scratching behavior in mice. Neuroscience 2009, 163, 23–33. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.J.; Ji, R.R. c-Fos and pERK, which is a better marker for neuronal activation and central sensitization after noxious stimulation and tissue injury? Open Pain J. 2009, 2, 11–17. [Google Scholar] [CrossRef]
- de Almeida, M.L.; Gottlieb, O.R. Iso-and neo-flavonoids from Dalbergia inundata. Phytochemistry 1974, 13, 751–752. [Google Scholar] [CrossRef]
- Coleman, N.; Nguyen, H.M.; Cao, Z.; Brown, B.M.; Jenkins, D.P.; Zolkowska, D.; Chen, Y.J.; Tanaka, B.S.; Goldin, A.L.; Rogawski, M.A.; et al. The riluzole derivative 2-amino-6-trifluoromethylthio-benzothiazole (SKA-19), a mixed KCa2 activator and NaV blocker, is a potent novel anticonvulsant. Neurotherapeutics 2015, 12, 234–249. [Google Scholar] [CrossRef] [Green Version]
- Qu, C.R.; Ding, M.M.; Zhu, Y.M.; Lu, Y.G.; Du, J.; Miller, M.; Tian, J.B.; Zhu, J.M.; Xui, J.; Wen, M.; et al. Pyrazolopyrimidines as potent stimulators for transient receptor potential canonical 3/6/7 channels. J. Med. Chem. 2017, 60, 4680–4692. [Google Scholar] [CrossRef] [PubMed]
- Gross, R.A.; Macdonald, R.L. Dynorphin A selectively reduces a large transient (N-type) calcium current of mouse dorsal root ganglion neurons in cell culture. Proc. Natl. Acad. Sci. USA 1987, 84, 5469–5473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, F.; Wu, Y.; Zou, X.H.; Tang, Q.L.; Zhao, F.; Cao, Z.Y. BmK AEP, an anti-epileptic peptide distinctly affects the gating of brain subtypes of voltage-gated sodium channels. Int. J. Mol. Sci. 2019, 20, 729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Y.; Zou, X.H.; Li, X.C.; Chen, J.; Jin, L.; Zhang, F.; Yu, B.Y.; Cao, Z.Y. Activation of sodium channels by α-scorpion toxin, BmK NT1, produced neurotoxicity in cerebellar granule cells: An association with intracellular Ca2+ overloading. Arch. Toxicol. 2017, 91, 935–948. [Google Scholar] [CrossRef]
- Shimada, S.G.; LaMotte, R.H. Behavioral differentiation between itch and pain in mouse. Pain 2008, 139, 681–687. [Google Scholar] [CrossRef] [PubMed] [Green Version]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, F.; Wu, Y.; Xue, S.; Wang, S.; Zhang, C.; Cao, Z. 3’-O-Methylorobol Inhibits the Voltage-Gated Sodium Channel Nav1.7 with Anti-Itch Efficacy in A Histamine-Dependent Itch Mouse Model. Int. J. Mol. Sci. 2019, 20, 6058. https://doi.org/10.3390/ijms20236058
Zhang F, Wu Y, Xue S, Wang S, Zhang C, Cao Z. 3’-O-Methylorobol Inhibits the Voltage-Gated Sodium Channel Nav1.7 with Anti-Itch Efficacy in A Histamine-Dependent Itch Mouse Model. International Journal of Molecular Sciences. 2019; 20(23):6058. https://doi.org/10.3390/ijms20236058
Chicago/Turabian StyleZhang, Fan, Ying Wu, Shuwen Xue, Shuangyan Wang, Chunlei Zhang, and Zhengyu Cao. 2019. "3’-O-Methylorobol Inhibits the Voltage-Gated Sodium Channel Nav1.7 with Anti-Itch Efficacy in A Histamine-Dependent Itch Mouse Model" International Journal of Molecular Sciences 20, no. 23: 6058. https://doi.org/10.3390/ijms20236058