


Abstract
Activation or blockade of cannabinoid CB1 receptors markedly alters many effects of opioids. In the present study, we investigated whether the cannabinoid antagonist (N-piperidinyl-5-(4-chlorophenyl)-1-(2,4-dichlorophenyl)-4-methylpyrazole-3-carboxamide (SR-141716A) could alter the reinforcing effects of heroin in rats. A Δ9-tetrahydrocannabinol (THC) drug-discrimination procedure was first used to determine effective CB1 antagonist doses of SR-141716A and optimal pretreatment times for self-administration studies. Subsequently, Sprague-Dawley rats learned to self-administer heroin under three different schedules of intravenous drug injection: a continuous reinforcement schedule [fixed ratio (FR)1], a five-response, fixed ratio schedule (FR5), and a progressive ratio schedule. Then, SR-141716A (1 mg/kg i.p.) was administered 60 min before the start of the session for three consecutive daily sessions. SR-141716A markedly decreased heroin self-administration under the progressive ratio schedule at heroin doses ranging from 12.5 to 100 μg/kg/injection. In contrast, SR-141716A had no effect on heroin self-administration under the FR1 schedule at heroin doses of 50 or 100 μg/kg/injection, but produced small decreases in self-administration at lower doses (25 and 12.5 μg/kg/injection). Consistent with a behavioral economics evaluation, SR-141716A produced a small but significant decrease in self-administration of the higher 50 μg/kg/injection dose of heroin when the fixed ratio requirement was raised to five (FR5). Thus, blockade of CB1 receptors differentially decreased the reinforcing efficacy of heroin depending on the number of responses required for each injection (price). These findings indicate a facilitatory modulation of opioid reward by endogenous cannabinoid activity and provide support for the use of cannabinoid CB1 antagonists as medications for heroin addiction.
A wide range of functional interactions between opioid and cannabinoid systems have been found in the areas of nociception, physical dependence development, direct measures of reward, such as intravenous drug self-administration behavior, and indirect measures of reward, such as drug-induced alterations in thresholds for electrical brain self-stimulation and conditioned place preference (Manzanares et al., 1999; Maldonado, 2002). Interactions reported have largely involved opioid modulation of cannabinoid effects. For example, self-administration of cannabinoid CB1 receptor agonists is reduced by pretreatment with an opioid antagonist in mice and squirrel monkeys (Goldberg et al., 2001; Navarro et al., 2001). Also, development of a preference for a distinctive compartment associated with THC administration is lost in μ-receptor knockout mice (Ghozland et al., 2002), and reduction by THC of the threshold for electrical brain self-stimulation is prevented by naloxone administration (Gardner and Lowinson, 1991). Studies of cannabinoid effects on opioid reinforcement indicate that opioid-cannabinoid interactions can also be bidirectional. For example, it has been reported that cannabinoid CB1 receptor knockout mice do not learn to self-administer morphine (Ledent et al., 1999) and do not develop conditioned preferences for places associated with opiate injection (Martin et al., 2000). Also, acute administration of the cannabinoid CB1 antagonist SR-141716A has been reported to precipitate signs of withdrawal in morphine-dependent rats (Navarro et al., 1998).
Recently, it has been reported that acute administration of the cannabinoid CB1 receptor antagonist SR-141716A can reduce intravenous heroin self-administration by rats (Navarro et al., 2001). This result, like the results of a number of experiments using other techniques, has been attributed to a reduction in the reinforcing effects of heroin. However, the self-administration part of that study, like many others in the rat self-administration literature, was limited to the evaluation of the effects of SR-141716A on self-administration of a single injection dose of heroin, only under a schedule of continuous reinforcement (FR1) and for a single session. Under such a restricted range of conditions, a critical interpretation of the treatment effects can be difficult.
It has been suggested that comparing the effects of pharmacological treatments under simple FR schedules and under more complex progressive ratio schedules, in which the number of responses required for injection increases progressively within a session, can greatly facilitate interpretation of observed changes in self-administration behavior (Arnold and Roberts, 1997). Progressive ratio schedules allow an estimation of the maximal effort an individual will perform under a specified set of conditions to obtain a particular reinforcing event and, thus, are believed to provide one measure of the motivational strength of the reinforcing event (Hodos, 1961). Also, because drug self-administration in animals is characterized by an inverted U-shaped dose-response curve, a decrease in the number of reinforcements obtained could reflect either a decrease or an increase in the drug's reinforcing effects, depending on the dose of drug chosen for study. Evaluation of the effects of pharmacological treatments on a complete self-administration, dose-response curve can help in drawing more meaningful conclusions. Finally, drug abuse and addiction are chronic or subchronic phenomena, and it is impossible to predict whether acute, single-session changes in self-administration would persist over time.
In the present study, we first investigated the effects of different doses of SR-141716A on food-maintained responding under a drug-discrimination procedure in which rats learned to discriminate between injections of THC and injections of vehicle. This allowed us to select a minimal dose of SR-141716A and an optimal pretreatment time for near maximal blockade of THC's discriminative effects (an indication of CB1 receptor blockade) for subsequent use in the heroin self-administration studies. Then, we compared the effects of pretreatment with SR-141716A on intravenous heroin self-administration by rats under continuous reinforcement (FR1) and progressive ratio schedules of drug injection. Under both types of schedule, full heroin dose-response functions were determined, with and without SR-141716A pretreatment, and each treatment condition was studied for three consecutive daily sessions. In addition, a control group of rats responded under a similar progressive ratio schedule of food reinforcement to assess selectivity of SR-141716A treatment effects. Finally, because the effects of SR-141716A seemed dependent on the schedule used (FR1 versus progressive ratio) and, thus, on the “price” of heroin, the number of responses required for reinforcement was increased from one to five (FR5) in a third group of rats at an injection dose of heroin that produced maximal responding under the progressive ratio schedule.
Materials and Methods
Subjects. Male Sprague-Dawley rats (Charles River Laboratories, Inc., Wilmington, MA) experimentally naive at the start of the study and initially weighing 300 to 350 g were used for all experiments. All rats were housed individually in a temperature- and humidity-controlled room and were maintained on a 12-h light/dark cycle; the lights were on from 6:45 AM to 6:45 PM. Experiments were conducted during the light phase. Animals were maintained in facilities fully accredited by the American Association for the Accreditation of Laboratory Animal Care; all experimentation was conducted in accordance with the guidelines of the Institutional Care and Use Committee of the Intramural Research Program, National Institute on Drug Abuse, National Institutes of Health, and the directives of the Principles of Laboratory Animal Care (National Institutes of Health publication 85-23, revised 1985).
For drug discrimination studies, approximately 1 week after arrival, rats' weights were gradually reduced to approximately 80% of free-feeding levels by limiting daily access to food and were kept at 80% throughout the study. This was consistent with our previous drug discrimination studies (Yasar et al., 1993). For i.v. heroin self-administration studies, rats had free access to food until recovery from surgery for implantation of i.v. catheters. One day before the start of daily self-administration sessions, food was restricted to approximately 15 g/day. Again, this provided consistency with a previous study of FR1 and progressive ratio heroin self-administration from our laboratory (Panlilio and Schindler, 2000). Rats working for food under a progressive ratio schedule were initially deprived to about 90% of their weight but after the acquisition of food responding, they were allowed to eat about 20 g of food pellets each day and their weight increased about 1 g/day. This level of food restriction resulted in breaking points similar to those obtained with 25 and 50 μg/kg/injection doses of heroin under the progressive ratio schedule. Food was always given shortly after the end of daily experimental sessions. Water was available ad libitum for all animals.
Catheterization. For rats trained with self-administration procedures, catheters were implanted in the right jugular vein under aseptic conditions using i.p. ketamine (60 mg/kg) and xylazine (10 mg/kg) anesthesia. Catheters consisted of approximately 4 cm of silastic tubing (0.044-mm i.d., 0.814-mm o.d.) connected to vinyl tubing (0.5-mm i.d., 1.0-mm o.d.; Tygon) with a 23-gauge stainless steel tube and bonded with polyethylene shrink tubing. A 10-mm-long section of Tygon tubing (5-mm o.d.) was glued to the catheter at the midpoint of the vinyl tubing and used as a subcutaneous anchor when the catheter exited the skin in the neck. The right external jugular vein was exposed by blunt dissection and the silastic portion of the catheter was inserted into the vein and sutured into place. The vinyl portion of the catheter was passed subcutaneously to the back of the neck, where the tip exited and was obturated with a modified 23-gauge needle. The incision was closed with stainless wound clips. A 20-mm plastic bolt was cemented with dental acrylic to four stainless jeweler's screws implanted in the skull. During experimental sessions, the catheter was connected to the injection pump via tubing passing through a metal spring that was attached to the plastic bolt on the rat's head. Catheters were flushed before and after each session with 0.1 ml of saline solution.
Drug Discrimination Apparatus and Procedure. Standard operant conditioning chambers (Coulbourn Instruments, Lehigh Valley, PA) were used. Each chamber contained a white house light and two levers, which were separated by a recessed tray into which a pellet dispenser could deliver 45-mg food pellets (F0021; Bioserv, Frenchtown, NJ). Each press of a lever with a force of 0.4 N through 1 mm was recorded as a response and was accompanied by an audible click. The operant-conditioning chambers were controlled by microcomputers using MED-PC software package (MED Associates, East Fairfield, VT).
Rats were trained, as described previously (Yasar et al., 1993), under a discrete-trial schedule of food-pellet delivery to respond on one lever after an injection of a training dose of 3 mg/kg THC and on the other lever after an injection of 2 ml/kg THC vehicle. Injections of THC or vehicle were given i.p. 30 min before the start of the session. At the start of the session, a white house light was turned on, and in its presence the rats were required to make 10 consecutive responses (fixed ratio 10 schedule of food delivery; FR10) on the lever appropriate to the presession treatment. The completion of 10 consecutive responses on the correct lever produced delivery of a 45-mg food pellet and initiated a 45-s time-out during which lever-press responses had no programmed consequences and the chamber was dark. Responses on the incorrect lever had no programmed consequences other than to reset the FR requirement on the correct lever. After each time-out, the white house light was again turned on and the next trial began. Each session ended after completion of 20 FR trials or after 30 min elapsed, whichever occurred first.
Discrimination training sessions were conducted 5 days/week under a double alternation schedule (i.e., DDSSDDSS, etc., D, drug, THC; S, saline). Training continued until there were eight consecutive sessions during which rats completed at least 90% of their responses during the session on the correct lever and no more than four responses occurred on the incorrect lever during the first trial. Test sessions were then initiated.
Test sessions were identical to training sessions with the exception that 10 consecutive responses on either one of the two levers ended the trial. Switching responding from one lever to the other lever reset the ratio requirement. In a test phase, a single alternation schedule was introduced and test sessions were usually conducted on Tuesdays and Fridays. Thus, a 2-week sequence starting on Monday was DTSDTSTDST (T, test). In this way, test sessions occurred with equal probability after saline and drug sessions. Test sessions were conducted only if the criteria of 90% accuracy and not more than four incorrect responses during the first trial were maintained in the two preceding training sessions.
Heroin Self-Administration Apparatus and Procedure. Experimental chambers (30 × 24 × 29 cm; Coulbourn Instruments) were enclosed individually in sound-attenuation chests. Each experimental chamber had a metal grid floor, a recessed food tray (not used for heroin-trained rats, but used for food-trained rats, as described below), and two nose-poke holes in the right wall. Previous data indicated that, in the absence of reinforcement, neither hole is favored (Panlilio and Schindler, 2000). A 4500-Hz auditory stimulus was provided by a Sonalert (model 628) operated at 8.75 V. Illumination was provided by a shielded white light bulb (type no. 1820) at all times except during injection and time-out periods, when both the tone and the light were pulsed at a rate of 5 Hz, with one stimulus on while the other was off. Drug solution was delivered through Tygon tubing, protected by a metal spring, and suspended through the ceiling of the experimental chamber from a single-channel fluid swivel. This tubing was attached to a syringe pump (MED Associates) that delivered fluid at a rate of 3.19 ml/min using a 10-ml syringe. Unit doses were manipulated by computerized adjustment of the duration of the injection (0.5–2 s). Experimental events were controlled by microcomputers using MED Associates interface and software (MED Associates).
Heroin Self-Administration under an FR1 Schedule. Self-administration sessions began approximately 1 week after catheter implantation. For the first two sessions, which lasted 3 h each, 50 injections of heroin at a dose of 25 μg/kg/injection were delivered independently of the rat's behavior. During these sessions, the nose-poke holes were removed from the chamber and injections were delivered at random intervals, averaging 225 s. Each injection of heroin was paired with flashing of the house light for 5 s. Beginning with the third session, the nose-poke holes were reinstalled, and an FR1 schedule was instituted. A single response in the left nose-poke hole immediately activated the injection pump and also caused the house light to flash and a tone to pulse for 5 s. During this 5-s time-out period, responding had no programmed consequences. After the time-out, the next response in the left hole was reinforced. Responses in the nose-poke hole on the right side were recorded but never reinforced. Sessions lasted 2.5 h. During initial training, a unit dose of 100 μg/kg/injection heroin was made available under the FR1 schedule described above. All rats began self-administering heroin within a few sessions. When consistent self-administration responding was obtained, rats were either moved to a lower dose of heroin or kept on the same dose until a stable baseline was reached and then they were tested with SR-141716A. Baseline responding was considered stable when the number of injections per session did not differ more than 10% for three consecutive days. Animals consistently increased the numbers of injections to compensate for decreases in the dose, and they were tested after they reached a new stable baseline (within three to five sessions). Not all rats received all treatments. As long as the catheter was patent at the end of one test condition, rats were switched to another dose and test condition.
Heroin Self-Administration under an FR5 Schedule. Rats first learned to respond for heroin under an FR1 schedule at a dose of 50 μg/kg/injection, as described above. When they showed consistent self-administration, the schedule was changed to FR5. Under this schedule, the first nose-poke in a session resulted in one injection of heroin but each subsequent injection of heroin required five nose-pokes. Within three to five sessions, rats reached a stable baseline of responding and they were then tested with SR-141716A. Baseline responding was considered stable when the number of injections per session did not differ more than 10% for three consecutive days.
Heroin Self-Administration under a Progressive Ratio Schedule. Rats first learned to respond for heroin under an FR1 schedule at a dose of 50 μg/kg/injection, as described above. When they showed consistent self-administration, they were directly switched to the progressive ratio schedule. Within three to five sessions, they reached stability and they were then either tested with SR-141716A or switched to a different heroin dose. Baseline responding was considered stable when the number of injections per session did not differ more than 10% for three consecutive days.
Under the progressive ratio schedule of i.v. drug injection, the response requirement increased with each successive injection. The steps of the exponential progression were the same as those used by Roberts and Bennett (1993), based on the following equation: response ratio = [5eX(0.2×infusion number)] – 5, rounded to the nearest integer. Thus, the values of the steps were 1, 2, 4, 6, 9, 12, 15, 20, 25, 32, 40, 50, 62, 77, 95, 118, 145, 178, 219, 268, 328, 402, 492, 603, and 737. Sessions under the progressive ratio schedule lasted5hor until 30 min passed without a response, which almost always occurred within less than 5 h. This 30-min nonresponse criterion was about 50% longer than the longest single interinfusion interval (around 20 min) recorded under any dose with the FR1 schedule of injection. The breaking point was defined as the step number of the last ratio completed before 30 min of nonresponse or at the end of the 5 h. Not all rats received all treatments. As long as the catheter was patent at the end of one test condition, rats were switched to another dose and test condition.
Food Reinforcement Apparatus and Procedure. The apparatus and the stimuli associated with the progressive ratio schedule of food delivery were exactly the same as described above for heroin injection.
Food Self-Administration under the Progressive Ratio Schedule. Rats learned to respond for food during one to three sessions under a FR1 schedule. In most cases, after the first day, in less than 30 min, rats earned more than 100 pellets. After these initial sessions, rats were directly switched to the progressive ratio schedule. Sessions under the progressive ratio schedule lasted until 5 min passed without a response, which almost always occurred within less than 1 h. This 5-min nonresponse criterion was chosen to obtain breaking points similar to those obtained in the heroin self-administration experiments. Responding was typically very high the first day and slowly decreased and stabilized within five to eight sessions. When a stable baseline was reached, animals were tested with SR-141716A. Baseline responding was considered stable when the number of food pellets earned per session did not differ by more than 10% for three consecutive sessions. Between test sessions, there were at least two control sessions to reestablish baseline responding. Usually, rats immediately returned to the original baseline level of responding so that three tests could be conducted over a 2-week period.
Drugs. Δ9-THC (National Institute on Drug Abuse, Baltimore, MD; 50 mg/ml in ethanol) was dissolved in a solution of 40% (w/v) cyclodextrin (Sigma/RBI, Natick, MA). Heroin (National Institute on Drug Abuse) was dissolved in distilled water. Unit doses were manipulated by adjusting the duration of the injection (1–4 s). SR-141716A (National Institute on Drug Abuse) was suspended in 0.3% Tween 80 in saline and administered i.p. in a volume of 2 ml/kg.
Data Analysis. Drug discrimination data were expressed as the percentage of the total responses on both levers that were made on the THC-appropriate lever. Response rate data were expressed as responses per second averaged over the session, with responding during time-out periods not included in calculations. The data from sessions during which rats did not complete at least one fixed ratio were excluded from analysis of drug-lever selection. All results are presented as group means (±S.E.M.). Statistical analysis on the effect of SR-141716A on drug-lever selection was done by using one-way ANOVA for repeated measures. Significant main effects were further analyzed post hoc by paired comparisons with the effects of the training dose of THC (3 mg/kg i.p.) alone using post hoc Dunnett's test. Changes were considered to be significant when P < 0.05.
Self-administration data for the effects of SR-141716A, vehicle pretreatment or saline extinction were analyzed as last ratio completed per session (±S.E.M.) after a logarithmic transformation for the progressive-ratio experiments; number of responses (±S.E.M.) and number of injections (±S.E.M.) were used for the fixed-ratio (FR1 and FR5) experiments. Data were subjected to a repeatedmeasures ANOVA, followed when appropriate by post hoc Dunnett's test with comparisons with the baseline condition. Changes were considered significant when P < 0.05.
Multilevel analysis with maximum likelihood estimation was performed with SAS software (SAS Institute, Cary, NC) using PROC MIXED (Singer, 1998) to analyze the drug intake at different unit dose of heroin during self-administration sessions under the progressive ratio schedule. This procedure has the flexibility to handle repeated measures data sets in which some subjects were not tested under all conditions.
Results
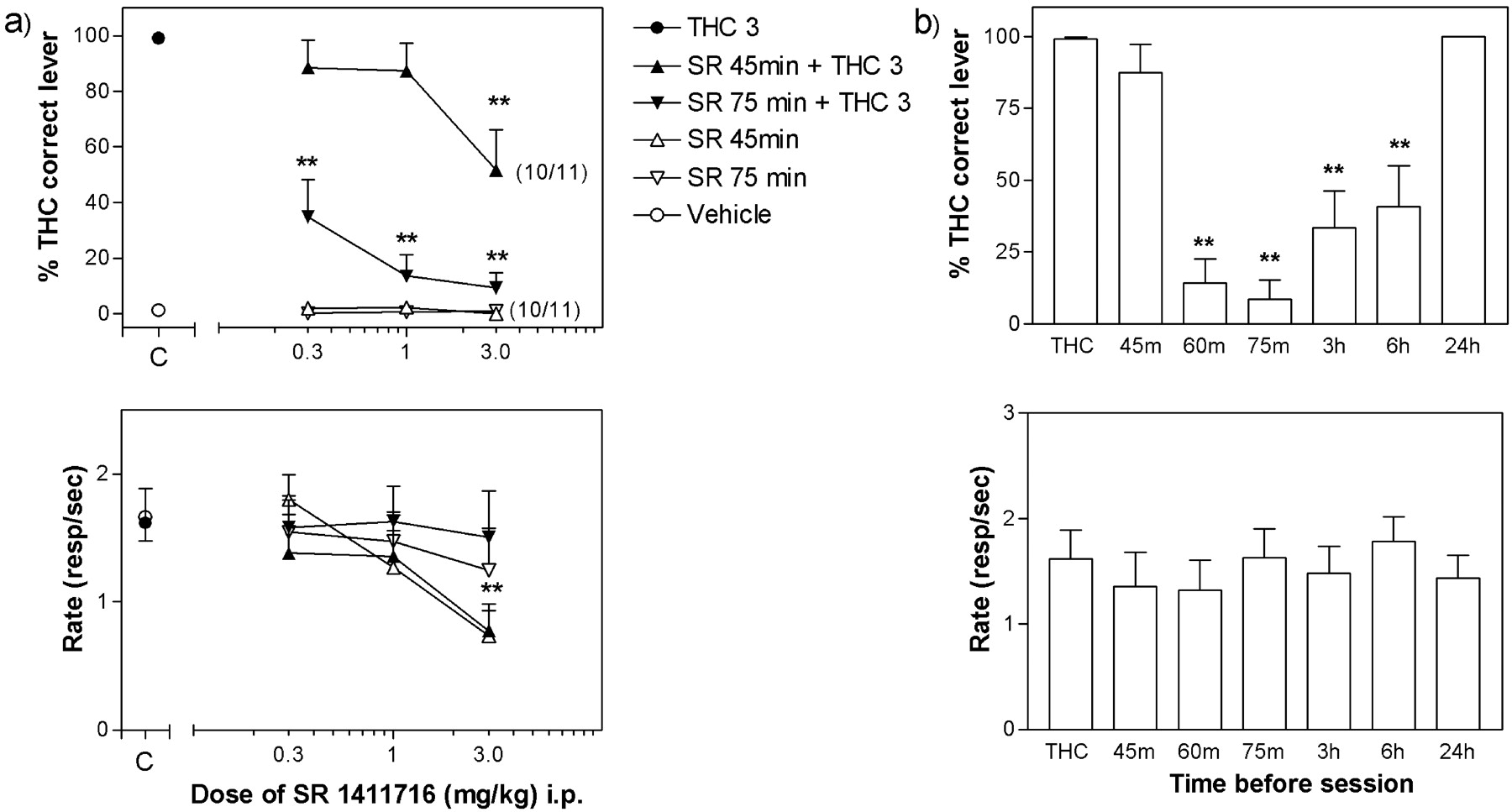
SR-141716A Antagonism of the THC Discriminative Stimulus Effects. The training dose of THC (3 mg/kg), administered 30 min before the session, produced about 100% drug lever selection, whereas drug lever selection was nearly 0% after vehicle administration (Fig. 1a, top). SR-141716A, given alone both 45 and 75 min before the session, produced vehicle-like responding. When SR-141716A was given 15 min before THC (45 min before the session), it only produced a partial blockade (about 50% drug lever selection) of the discriminative stimulus effects of THC [F(3,27) = 6.135, P < 0.005] at the highest tested dose of SR-141716A (3 mg/kg). This dose of SR-141716A produced a significant decrease in the rate of responding both alone [F(3,30) = 5.402, P < 0.01] and in combination with THC [F(3,30) = 5.175, P < 0.05] (Fig. 1a, bottom). In contrast, when given 45 min before THC (75 min before the session), SR-141716A produced an almost complete blockade (less than 20% of THC lever selection) of THC effects [F(3,30) = 38.559, P < 0.0001], with the dose of 1 mg/kg being as effective as the higher dose of 3 mg/kg. SR-1411716 had no effect on the rate of responding when administered alone or in combination with THC 75 min before the session.

Effects of SR-141716A pretreatment on the discriminative stimulus effects of THC. Ordinates, percentage of responses on the lever associated with THC administration (top) and overall rate of lever pressing during the session (bottom). Abscissa, dose of SR-141716A in milligrams per kilogram, log scale (a) or pretreatment time before the session in minutes or hours (b). Dose-response and pretreatment-time data are means (±S.E.M.) from 11 rats. The numbers in parentheses at the 3-mg/kg dose of SR-141716A show the number of rats that completed at least one fixed ratio during the session relative to the total number of rats tested. b, time-course effects of pretreatment with 1 mg/kg i.p. SR-141716A on THC drug discrimination performance. *, P < 0.05; **, P < 0.01. Post hoc comparison with the effect of the training dose of THC after significant ANOVA for repeated measures main effect, Dunnett's test.
A dose of 1 mg/kg SR-141716A was chosen to perform a time-course study (Fig. 1b). This dose of SR-141716A was not effective when given 45 min before the session. When given 60 or 75 min before the session, 1 mg/kg SR-141716A produced a complete blockade of THC discriminative effects [F(7,70) = 21.924, P < 0.001]. The blockade, even though partial, was still present when SR-141716A was administered 3 and 6 h before the session. Finally, no effect of SR-141716A was found 24 h after administration. At none of the time points did SR-141716A at the dose of 1 mg/kg produce significant changes in the rate of responding.
Effects of SR-141716A on Heroin Self-Administration under the FR1 Schedule. Rats readily learned to nose-poke for i.v. injections of heroin (100 μg/kg/injection). When the dose of heroin was reduced, the number of injections self-administered per session increased (Fig. 2a) and the latency between consecutive injections decreased (Fig. 2c), resulting in maintenance of a fairly constant intake. The mean heroin intake per session was 1.539 ± 0.14 mg/kg at a dose of 100 μg/kg/injection, 1.37 ± 0.24 mg/kg at a dose of 50 μg/kg/injection, 1.18 ± 0.25 mg/kg at a dose of 25 μg/kg/ injection, and 0.91 ± 0.07 at a dose of 12.5 μg/kg/injection (Fig. 2b). When SR-141716A (1 mg/kg i.p.) was given 1 h before each session for three consecutive sessions, it had no effect on self-administration of high doses of heroin (50 and 100 μg/kg/injection) (Fig. 2, a and b). In contrast, SR-141716A significantly decreased self-administration of lower heroin doses of 25 μg/kg/injection [F(3,18) = 7.288, P < 0.005] and 12.5 μg/kg/injection heroin [F(3,18) = 5.236, P < 0.01] (Fig. 2, a and b). The number of heroin injections self-administered progressively decreased over three consecutive sessions with SR-141716A pretreatment. Analysis of interinjection intervals showed a significant effect of SR-141716A on interinjection interval at the 12.5 μg/kg/injection dose of heroin [one-way ANOVA repeated measures, treatment × distribution, F(2,30) = 2.946, P < 0.0001], which consisted of a downward shift of the interinjection-frequency curve (Fig. 2c). When saline was substituted for heroin, the number of injections self-administered per session increased on the 1st day but decreased on the 2nd and 3rd days (Fig. 2a, top right).

Self-administration of different doses of heroin under a continuous reinforcement (FR1) schedule and effects of three consecutive sessions with SR-141716A pretreatment (1 mg/kg i.p. 60 min before session). a, effect on number of injections. Data are expressed as mean (±S.E.M.) of the number of injections per session during baseline (BL) conditions and during three consecutive sessions with vehicle or SR-141716A pretreatment (1 mg/kg i.p. 60 min before session) or during three consecutive sessions with saline substituted for heroin (a, top right). b, effect on heroin intake. Data are expressed as mean (±S.E.M.) of the number of total heroin intake per session in milligrams per kilogram during baseline conditions (filled symbols) and during the last of three consecutive sessions with SR-141716A pretreatment (open symbols). c, effects on frequency distribution of interinjection intervals. Points represent the mean number of heroin injections that occurred after each indicated interinjection interval (n = 6–7/group). The empty circles represent the last of three consecutive sessions with SR-141716A pretreatment (1 mg/kg i.p. 60 min before session), at a heroin dose of 12.5 μg/kg/injection. *, P < 0.05; **, P < 0.01. Post hoc comparison BL versus treatment sessions after significant ANOVA for repeated measures main effect, Dunnett's test.
Heroin Self-Administration under the Progressive Ratio Schedule. When rats were changed from the FR1 schedule to the progressive ratio schedule, they readily increased their rate of responding. The total number of responses (Fig. 3a) and number of injections per session (Fig. 3b), as well as the breaking point, as measured by last ratio completed (Fig. 4a), showed similar inverted U-shaped dose-response curves with all three measures. The lowest (12.5 μg/kg/injection) and highest (100 μg/kg/injection) doses of heroin maintained similar low breaking points and number of injections and responses per session, whereas the two intermediate doses (25 and 50 μg/kg/injection) maintained higher breaking points, number of injections, and responses per session. Representative cumulative response records of a rat responding at each of the four doses of heroin are shown in Fig. 5a. Note that maximal responding was maintained at the 50 μg/kg/injection dose of heroin and that both responses per session and the time to cessation of responding within a session (indicated by arrows in Fig. 5) showed dose-related decreases with decreases in heroin dose. The highest doses (50 and 100 μg/kg/injection) maintained more consistent self-administration, considering individual animal variability from session to session and variability between animals. Also, the dose-effect curve for heroin intake per session was linearly dose-dependent [F(3,36) = 22.83, P < 0.001] with the dose of 100 μg/kg/injection producing an intake about 50% higher than the lower dose of 50 μg/kg/injection (Fig. 3c). All animals reached a stable baseline in a few sessions when trained at these doses. On the other hand, heroin doses of 12.5 and 25 μg/kg/injection produced less consistent behavior.

Self-administration of different doses of heroin under a progressive ratio schedule and effects of three consecutive sessions with SR-141716A pretreatment. Data are expressed as mean (±S.E.M.) of number of responses per session (a), number of injections per session (b), and total intake of heroin in milligrams per kilogram per session (c) during baseline conditions (filled symbols) and during the last session with SR-141716A 1 mg/kg i.p. 60 min before session pretreatment (open symbols) (n = 5–8/group).

a, effects of SR-141716A on self-administration of different doses of heroin under a progressive ratio schedule. Data are expressed as means (±S.E.M.) of the last ratio completed (breaking point) during baseline conditions (BL) and during three consecutive sessions with SR-141716A pretreatment (1 mg/kg i.p. 60 min before session) for each group (n = 5–8). b, effects of vehicle or SR-141716A pretreatment on heroin self-administration, effects of substituting saline for heroin (extinction), and effects of SR-141716A on food-maintained responding under the progressive ratio schedule. Data are expressed as means (±S.E.M.) of the last ratio completed (breaking point) during baseline conditions (BL) and during three consecutive sessions with vehicle or SR-141716A pretreatment (1 mg/kg i.p. 60 min before session) or during three consecutive sessions of saline extinction (ext) (n = 5–8). *, P < 0.05; **, P < 0.01. Post hoc comparison BL versus treatment sessions after significant ANOVA for repeated measures main effect, Dunnett's test.

Representative cumulative-response records of heroin self-administration under the progressive ratio schedule. Ordinates represent cumulative number of nose-poke responses and abscissa represent time. Vertical lines indicate heroin injections. Arrows indicate the end of the session. a, self-administration of different unit doses of heroin. At doses of 12.5, 25, 50, and 100 μg/kg/injection, the number of injections of heroin self-administered per session was 9, 17, 21, and 16, the last ratio completed was 25, 145, 328, and 118 and the total number of responses per session was 104, 784, 1716, and 582, respectively. B, self-administration of 50 μg/kg injections of heroin during baseline conditions (top record) and pretreatment with 1 mg/kg SR-141716A (middle records) and during saline extinction (bottom records). Note that decreases in responding and the time to cessation of responding within the session were more pronounced after saline extinction than after SR-141716A pretreatment.
Effects of SR-141716A on Heroin Self-Administration under the Progressive Ratio Schedule. A 1-mg/kg dose of SR-141716A, given i.p. 1 h before the session for three consecutive sessions, significantly decreased heroin self-administration at the 25 μg/kg/injection [F(3,18) = 4.560, P < 0.05], 50 μg/kg/injection [F(3,21) = 8.171, P < 0.001], and 100 μg/kg/injection [F(3,18) = 9.139, P < 0.001] doses (Figs. 3, a–c, and 4a). At the lowest dose of 12.5 μg/kg/injection of heroin, SR-141716A decreased responding over three consecutive sessions, but results did not reach significance. Representative cumulative-response records of a rat responding for heroin under the progressive ratio schedule at the 50 μg/kg/injection dose of heroin under baseline conditions and on the first and the last of 3 days of pretreatment with 1 mg/kg SR-141716A are shown in Fig. 5b. Note that under baseline conditions, 17 injections of heroin were self-administered, the last ratio completed (breaking point) was 145, and the total number of responses in the session was 811. When SR-141716A was given 60 min before the session, the number of heroin injections in the session dropped to 16 and 14, the breaking point dropped to 118 and 77, and the total number of responses in the session dropped to 634 and 468, respectively, on the 1st and 3rd day of treatment.
Effect of Vehicle Pretreatment and Saline Substitution on Heroin Self-Administration under the Progressive Ratio Schedule. In rats self-administering 50 μg/kg injections of heroin under the progressive ratio schedule, pretreatment with the vehicle for SR-141716A for three consecutive sessions had no effect on responding (Fig. 4b). When saline was substituted for heroin, the breaking point significantly decreased (F(3,12) = 21.424, P < 0.0001) (Fig. 4b). This effect was relatively moderate on the 1st day but very pronounced on the 2nd and 3rd day. Representative cumulative response records of a rat responding for 50 μg/kg injections of heroin under baseline conditions and on the first and the last of 3 days of saline extinction are shown in Fig. 5b. Note that under baseline conditions 17 injections of heroin were self-administered, the last ratio completed (breaking point) was 145, and the total number of responses in the session was 811. When saline was substituted for heroin, the number of injections in the session dropped to 10 and 8, the breaking point dropped to 32 and 20, and the total number of responses in the session dropped to 151 and 81, respectively, on the 1st and 3rd days of extinction.
Effects of SR-141716A on Food-Maintained Responding under the Progressive Ratio Schedule. Pretreatment with 1.0 mg/kg SR-141716A for three consecutive sessions did not significantly affect responding for food under a progressive ratio schedule (Fig. 4b). There was a small but nonsignificant decrease in responding for food on the 1st day with SR-141716A pretreatment, but no further changes were seen on the 2nd or 3rd days of SR-141716A pretreatment.
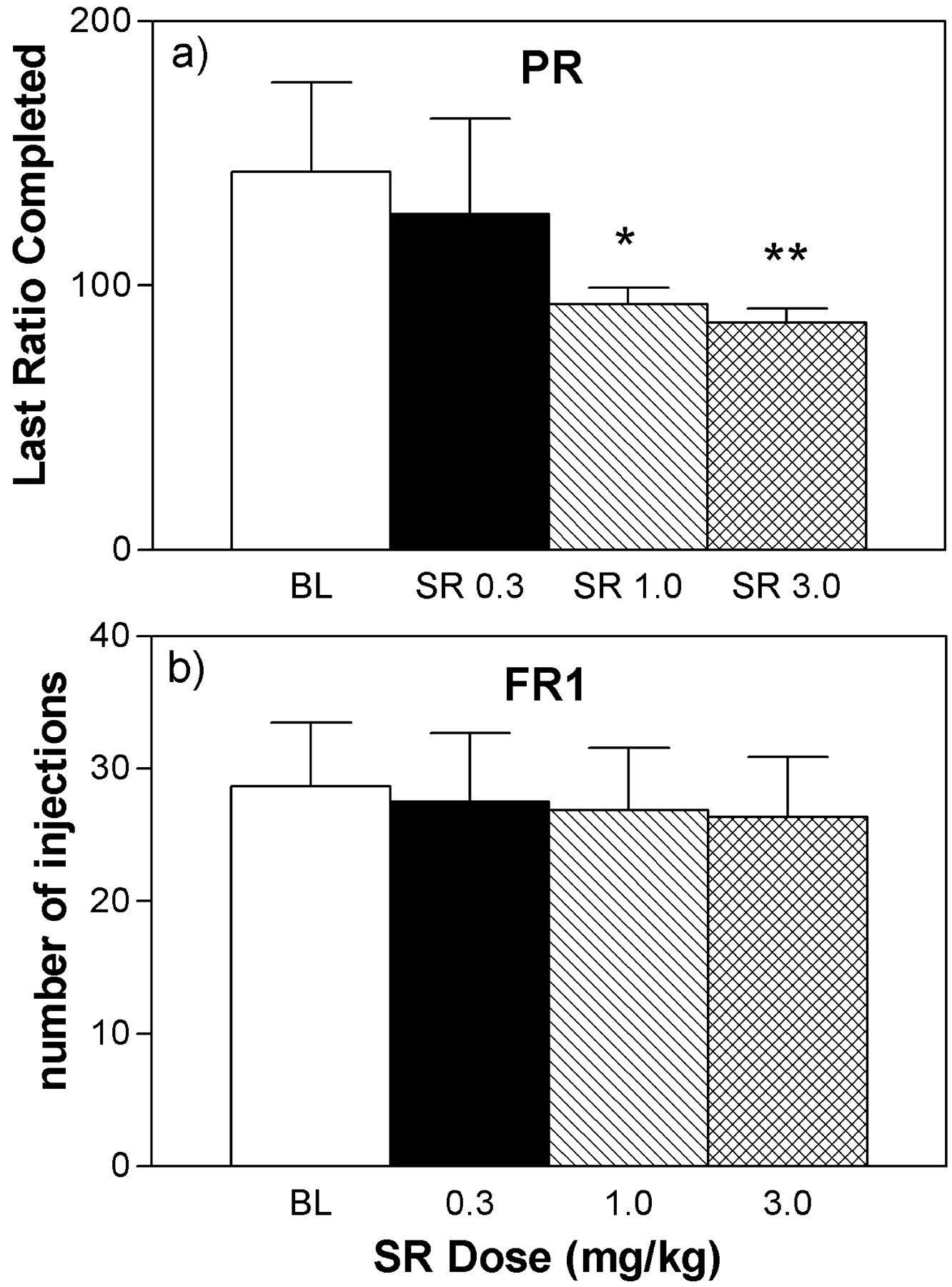
Effects of Different Doses of SR-141716A on Self-Administration of 50 μg/kg/injection of Heroin under FR1 and Progressive Ratio Schedules.When doses of 0.3, 1, and 3 mg/kg SR-141716A were given 1 h before the session for a single session, there were dose-dependent decreases in responding for 50 μg/kg injections of heroin under the progressive ratio schedule, as measured by last ratio completed [F(3,12) = 8.72, P < 0.005] (Fig. 6a). However, the dose of 1 mg/kg SR-141716A produced a near maximal reduction in responding, compared with the high dose of 3 mg/kg. In contrast, under the FR1 schedule, SR-141716A had no effect on responding for 50 μg/kg injections of heroin at any of the tested doses (Fig. 6b).

Effects of different doses of SR-141716A on self-administration of 50 μg/kg injections of heroin under a progressive ratio (PR) schedule (a) and under a continuous reinforcement (FR1) schedule (b). Data under the progressive ratio schedule are expressed as mean (±S.E.M.) of the last ratio completed during baseline conditions and during single sessions with SR-141716A pretreatment (n = 6). Under the FR1 schedule, data are expressed as mean (±S.E.M.) of the number of injections per session during baseline conditions and during single sessions with SR-141716A pretreatment (n = 6). *, P < 0.05; **, P < 0.01. Post hoc comparison BL versus treatment sessions after significant ANOVA for repeated measures main effect, Dunnett's test.
Effects of SR-141716A on Self-Administration of Heroin under an FR5 Schedule. Rats readily increased their responding when the FR requirement was changed from 1 to 5 and the number of injections self-administered per session remained relatively unchanged. An i.p. dose of 1 mg/kg SR-141716A, which had no effect on self-administration responding under the FR1 schedule, significantly decreased the number of injections self-administered [F(3,15) = 5.780, P < 0.001] (Fig. 7) under the FR5 schedule.

Effects of SR-141716A on i.v. self-administration of 50 μg/kg injections of heroin under an FR5 schedule. Data are expressed as mean (±S.E.M.) of the number of injections per session during baseline conditions and during three consecutive sessions with saline or SR-141716A pretreatment (1 mg/kg i.p. 60 min before session) (n = 6). *, P < 0.05; **, P < 0.01. Post hoc comparison baseline versus treatment sessions after significant ANOVA for repeated measures main effect, Dunnett's test.
Discussion
Heroin generally serves as an effective reinforcer of operant behavior in both humans and experimental animals (for review, see van Ree et al., 1999). In our study, rats rapidly acquired stable self-administration behavior under the FR1 schedule when each response produced an injection of 100 μg/kg heroin, and they readily increased the number of responses when either the dose was decreased or the response requirement was increased. Under the progressive ratio schedule, the dose-effect curve for responses per session and for maximal ratio size completed before responding for heroin ceased (the breaking point) had inverted U shapes. The lowest and highest injection doses of heroin (12.5 and 100 μg/kg) produced similar low numbers of responses and breaking points, whereas intermediate injection doses of 25 and 50 μg/kg heroin produced more responding and higher breaking points, consistent with previous findings under progressive ratio schedules of heroin injection in rats (Roberts and Bennett, 1993; Panlilio and Schindler, 2000).
Before studying the effects of SR-141716A on heroin self-administration, a drug-discrimination study was conducted using 3 mg/kg i.p. Δ9-THC as a training drug and dose, to select an appropriate SR-141716A dose and pretreatment time. Consistent with previous studies (Wiley et al., 1995a,b), we found that a 1-mg/kg dose of SR-141716A was as effective as a higher 3-mg/kg dose in blocking the discriminative stimulus effects of the training dose of Δ9-THC, and unlike the higher 3-mg/kg dose, it did not depress responding for food at any time point. We also found that a long (30 and 45 min before the injection of THC), but not a short (15 min before the injection of THC) pretreatment time produced complete blockade of the discriminative effects of THC. We then verified that SR-141716A was still effective in blocking the THC effects after 2.5 and 5.5 h (about the length of our self-administration sessions), but its effects disappeared after 24 h. Consequently, the dose of 1 mg/kg SR-141716A with a pretreatment time of 60 min was selected for repeated testing, because it seemed to be a dose with maximal CB-1 blocking actions but with no nonspecific disruptive effects on food-maintained responding, and would not be likely to produce accumulation over successive days of testing.
Blockade of CB1 receptors by SR-141716A markedly reduced responding for intravenous heroin injections under the progressive ratio schedule, but had little effect on responding for food under a similar progressive ratio schedule. Reductions in the number of responses per session, number of injections per session, and in the breaking point (the last ratio completed before responding ceased) under the progressive ratio schedule occurred across a wide range of heroin doses (12.5–100 μg/kg/injection) and were maintained over three consecutive sessions with SR-141716A pretreatment. In contrast to effects under the progressive ratio schedule, SR-141716A only reduced responding for low 12.5 and 25 μg/kg injection doses of heroin under the FR1 schedule, and this effect became clearly significant only after the third consecutive session with SR-141716A pretreatment. Analysis of interinjection intervals in the FR1 schedules indicated that administration of 1 mg/kg SR-141716 produced a significant decrease in responding for heroin (only at the dose of 12.5 μg/kg/injection) without modifying the interinjection pattern (frequency distribution) of self-administered injections. It has been suggested that such an effect would indicate that the compound does not interact directly with the mechanism of action of the self-administered drug, whereas a shift in the interinjection pattern would indicate a direct interaction with its primary mechanism of action (Brebner et al., 2000).
The fact that heroin self-administration was affected in a different manner under these three schedules is consistent with a behavioral economic analysis (Bickel et al., 2000), with the effects of SR-141716A on heroin self-administration being strong under a progressive ratio schedule (high price of heroin), weaker under an FR1 schedule of self-administration of low doses of heroin (12.5 and 25 μg/kg injection) (lower prices of heroin), and null under an FR1 schedule of self-administration of higher doses of heroin (50 and 100 μg/kg injection) (very low prices of heroin). When price of heroin under the FR schedule was increased by increasing the response requirement 5-fold to FR5 for the 50 μg/kg injection dose of heroin, SR-141716A did significantly decrease responding. Thus, the effects of SR-141716A in our study seem to depend on the price of heroin with only high prices being sensitive to its effects.
Because there is evidence that SR-141716A can act as an inverse agonist at higher doses (Landsman et al., 1997; Pan et al., 1998), in our study we chose a dose of 1 mg/kg SR-141716A, the lowest dose with a maximal effect as a CB1 receptor antagonist (as judged by blockade of discriminated effects of a high 3-mg/kg dose of THC in the drug discrimination experiment). Increasing the dose from 1 to 3 mg/kg did not significantly change the effects of SR-141716A on heroin self-administration under the progressive ratio or the FR1 schedules. However, in our study, a 3-mg/kg dose of SR-141716A did markedly and significantly depress responding for food under an FR10 schedule when it was given alone 45 min before the start of a 30-min session. In a previous study by Navarro et al. (2001), a high 3-mg/kg dose of SR-141716A was found to decrease responding for heroin under an FR1 schedule. In that study, SR-141716A was given 30 min after the start of the session (the number of injections in subsequent 30-min blocks was then measured). Under these conditions, SR-141716A may have acted in a nonspecific manner disrupting the operant behavior of the rats, although a control experiment with rats responding for food under different experimental conditions found no effect of 3 mg/kg SR-141716A (Navarro et al., 2001).
It is worth noting that repeating a treatment over consecutive daily sessions is not a usual approach in rodent self-administration studies, although it provides important information about the effects of any pretreatment or manipulation. This seems clear from our saline-extinction control data under both the progressive ratio and the continuous reinforcement (FR1) schedules. When saline was substituted for heroin under the progressive ratio schedule, number of responses and number of injections per session and the breaking point showed a relatively small decrease on the 1st day, but the decreases were much larger on the 2nd and the 3rd day. Under the FR1 schedule, an increase in the number of injections on the 1st day was followed by a decrease on the 2nd and 3rd day, consistent with extinction of primary reinforcement. On the other hand, in rats responding for food on a progressive ratio schedule, SR-141716A produced a small but not significant decrease in responding on the 1st day of treatment but no further changes on the 2nd and the 3rd day. Thus, repeated pretreatment helps to avoid the possibility of introducing false positive or false negative results. In addition, because drug addiction is a chronic condition characterized by frequent relapse to drug use even after long drug-free periods, any possible treatment must be proven to be effective over time.
There are several possible neurochemical mechanisms that may mediate the attenuation of heroin's reinforcing effects by CB1 receptor blockade with SR-141716A. One possibility is that heroin releases endocannabinoids, such as anandamide, and this release plays a role in the reinforcing effects of heroin. Because procedures for measuring endogenous cannabinoids have only recently been established (Giuffrida and Piomelli, 1998), there is no direct evidence for such a mechanism. However, it has recently been shown that stimulation of dopamine D2 receptors produces release of anandamide in the striatum (Giuffrida et al., 1999). Because it is well established that heroin releases dopamine in the accumbens and striatum (Di Chiara and Imperato, 1988), this dopamine release could in turn release endocannabinoids.
Another possibility is that opioid and cannabinoid signaling converge at some point and blockade of cannabinoid CB1 receptors in some way decreases the effects of μ-agonists such as heroin. Opioid μ-receptors and cannabinoid CB1 receptors have a similar anatomical distribution in the brain (Herkenham et al., 1991; Matsuda et al., 1993), are both members of the G protein-coupled family of receptors, and they both modulate similar transduction systems (Reisine et al., 1996; Howlett, 2002). The convergence of the two signals could explain the interactions between cannabinoid and opioid systems. For example, it has been recently shown that the binding of SR-141716A to the cannabinoid CB1 receptor can sequester Gi/o proteins from a common pool and prevent other receptors from signaling (Vasquez and Lewis, 1999). Also, it has been shown that CB1 receptors and μ-opioid receptors are, at least in part, colocalized in the rat caudate putamen (Rodriguez et al., 2001).
There have been a number of studies in which pharmacological blockade of cannabinoid CB1 receptors or their depletion in CB1 knockout mice have been reported to influence the motivational effects of other drugs of abuse or of natural reinforcers, such as food. It has been reported, for example, that SR-141716A treatment can prevent the development of a conditioned place preference for a compartment associated with cocaine, morphine, or food (Chaperon et al., 1998). Also, SR-141716A treatment can decrease sensitivity to the reinforcing effects of electrical brain stimulation in rats (Deroche-Gamonet et al., 2001), decrease the self-administration of both methamphetamine (Vinklerova et al., 2002) and nicotine (Cohen et al., 2002) by rats, and prevent the reinstatement of extinguished cocaine self-administration responding by a priming injection of cocaine or by cocaine-associated “cues” (De Vries et al., 2001). However, some studies, including our study, have had contradictory findings. For example, in a study with CB1 knockout mice, morphine, but not cocaine, failed to produce a conditioned place preference (Martin et al., 2000). In another study, it was reported that CB1 knockout mice do not learn to self-administer morphine but do learn to self-administer cocaine, amphetamine, and nicotine (Cossu et al., 2001). Finally, in our study, the marked effect of SR-141716A was specific for heroin and not simply due to nonspecific effects on systems mediating motivation or reward, because no parallel effect of SR-141716A was found when comparable progressive ratio responding was maintained by food.
The present data demonstrate that the cannabinoid CB1 receptor antagonist SR-141716A produces a clear attenuation, but not a complete blockade, of the reinforcing effects of heroin as measured by decreases in the number of responses and injections per session under the three schedules of drug injection and by breaking points under the progressive ratio schedule. Together, our findings indicate a facilitatory modulation of opioid reinforcement by endogenous cannabinoid activity that is unmasked by CB1 receptor blockade. Because the breaking point measure with progressive ratio schedules of drug injection is often considered a measure of drug “craving” in the presence of the drug (Markou et al., 1993) and because breaking points under the comparable progressive ratio schedule of food delivery were unaltered by SR-141716A treatment, the present findings provide further support for the potential efficacy of SR-141716A in the prevention and treatment of opioid addiction (Navarro et al., 1998, 2001; Rubino et al., 2000).
Footnotes
-
DOI: 10.1124/jpet.102.047928.
-
ABBREVIATIONS: THC, Δ9-tetrahydrocannabinol; SR-141716A, (N-piperidinyl-5-(4-chlorophenyl)-1-(2,4-dichlorophenyl)-4-methylpyrazole-3-carboxamide; CB, cannabinoid; FR, fixed ratio; ANOVA, analysis of variance.
- Received December 20, 2002.
- Accepted March 17, 2003.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}