


Abstract
Previous studies have shown that 5-hydroxytryptamine (5-HT) can modulate the hyperpolarization-activated nonselective cation current (Ih) to elicit a membrane depolarization in neurons. However, the receptor subtype involved in this response remains controversial. In the accompanying study, we have identified a 5-HT7 receptor-mediated depolarization in the anterodorsal nucleus of the thalamus (ADn). In the present study, we have examined the possible role of Ih in mediating this 5-HT7receptor-mediated depolarization. We used the blind tight-seal patch clamp technique to examine the ability of 5-HT to modulate Ih in the ADn. We found that 5-HT induced a shift in the voltage dependence of Ih to more depolarized potentials. The pharmacology of the receptor mediating this effect was consistent with that of a 5-HT7 receptor. Since the 5-HT7receptor is coupled positively to adenylate cyclase, we examined the cAMP dependence of the 5-HT-induced modulation of Ih. Intracellular addition of cAMP mimicked and occluded the 5-HT response. Conversely, in the presence of the protein kinase inhibitors H-8 and staurosporine, ADn neurons still expressed a 5-HT-induced shift in the voltage dependence of Ih. These results suggest that 5-HT regulates Ih in the ADn through a cAMP-dependent but protein kinase A (PKA)-independent mechanism. To determine the contribution of Ih to the 5-HT7receptor-mediated depolarization, we used the selective Ihblocker ZD7288. This compound greatly reduced the depolarizing response elicited by activation of 5-HT7 receptors. We conclude that 5-HT7 receptors depolarize ADn neurons primarily by increasing Ih through a cAMP-dependent, PKA-independent mechanism.
In the accompanying article (Chapin and Andrade, 2001), we reported that activation of the 5-HT7 receptor elicits a membrane depolarization and associated inward current in the anterodorsal nucleus of the thalamus (ADn). In many cases, 5-HT-induced depolarizations are elicited through modulation of the hyperpolarization-activated nonselective cation current, Ih (Bobker and Williams, 1989; Pape and McCormick, 1989; Takahashi and Berger, 1990; Larkman and Kelly, 1992;Spain, 1994; Cardenas et al., 1999). Therefore, we tested the possibility that Ih may, at least in part, mediate the depolarization elicited by 5-HT7receptors.
Ih is a widely distributed nonselective cation current that is activated on hyperpolarization and can display very slow activation kinetics. Under physiological conditions, Ih reverses at approximately −20 to −30 mV (Yanagihara and Irisawa, 1980; Takahashi and Berger et al., 1990; Li et al., 1993) and activates at hyperpolarized potentials with half-activation (V0.5) values ranging from −92 mV (Larkman and Kelly, 1992) to −75 mV (Banks et al., 1993). Its time constant for activation in a physiological preparation can be slower than 2 s (Pape and McCormick, 1989). Ih is also blocked by ZD7288 with reasonable selectivity and this compound can be used to differentiate it from other currents (BoSmith et al., 1993; Harris and Constanti, 1995). A final characteristic of Ih is its regulation by cAMP. Increased intracellular cAMP shifts the voltage dependence of activation of Ih to more depolarized potentials (DiFrancesco and Tortora, 1991) and, in some cases, increases its maximal conductance (Tokimasa and Akasu, 1990; Accili et al., 1997). Since under physiological conditions Ih is an inward current, these effects result in a greater number of Ih channels open at rest. This change becomes manifested as a membrane depolarization.
Ih-mediated depolarizations signaled through cAMP secondary to the activation of 5-HT receptors have been identified in several brain regions, including thalamic geniculate nuclei (Pape and McCormick, 1989), the brainstem nucleus prepositus hyppoglossi (Bobker and Williams, 1989), the medial nucleus of the trapezoid body (Banks et al., 1993), the cerebral cortex (Spain, 1994), and facial and spinal motor neurons (Takahashi and Berger, 1990; Larkman and Kelly, 1992). However, the specific serotonin receptor subtype involved in these responses remains controversial (Bobker and Williams, 1989; McCormick and Pape, 1990; Takahashi and Berger, 1990; Larkman and Kelly, 1992). Given the involvement of cAMP in regulating Ih, it seems most likely that the receptor involved would belong to the 5-HT4, 5-HT6, or 5-HT7 subtypes. However, this conjecture has not been rigorously tested.
In the preceding study, we identified a serotonin-induced depolarization in the ADn that is mediated by receptors of the 5-HT7 subtype. We hypothesized that part or all of this 5-HT7 receptor-mediated depolarization/inward current could be mediated by modulation of Ih in a cAMP-dependent manner. Therefore, in the present study, we first examined whether 5-HT regulated Ih in the ADn. We then tested for involvement of a 5-HT7 receptor in this response using a pharmacological approach. Finally, we directly examined the role of Ih in mediating the 5-HT7receptor-induced inward current in this region. From the results of these experiments, we conclude that 5-HT7receptors depolarize ADn neurons by regulating Ihthrough a cAMP-dependent, but PKA-independent, mechanism.
Materials and Methods
The methods for preparation of brain slices and electrophysiological recordings were essentially as outlined in the accompanying article (Chapin and Andrade, 2001). To generate Ih activation curves, cells were held at −40 mV. Current-voltage relationships were generated by applying 4.0- to 5.6-s long hyperpolarizing pulses every 10 to 20 s to increasingly hyperpolarized steps (in 5–10 mV increments). Ihwas measured by subtracting the instantaneous current from the steady-state current. The amplitude of Ih was normalized to the maximal value.
Data Analysis.
Data were analyzed using Origin 6.0 (Microcal Software, Northampton, MA). The voltage activation curve for each cell was fitted to the Boltzmann equation (I/Imax = 1{1 + exp[(Vm −V0.5)s−1]}−1), whereI/Imax is the normalized current,Vm is the command voltage,V0.5 is the half-activation voltage, and s is the slope factor. The slope factors and theV0.5 values were allowed to vary for analysis. Statistical data were tested using GB-STAT 6.0 or GraphPad Prism. Fitting of the time constant of activation was done using a second-order exponential decay equation (I= Io + A1e−(x−xo)/τ1 +A2e−(x−xo)/τ2) where I is the current, x represents time in ms, and τ is the time constant. Except where indicated, data are presented as means ± S.E.M.
Results
As illustrated in Fig. 1A, cells of the ADn express an Ih (n > 50 cells). We characterized this current by applying voltage steps to increasingly hyperpolarized potentials from a holding potential of −40 mV (Fig. 1A). Steps to voltages negative to −60 mV revealed a slowly developing inward relaxation. This is the physiological profile expected for Ih. We confirmed that this slowly developing inward current corresponded to Ih by its sensitivity to ZD7288.

Ih in the ADn is regulated by serotonin. A, hyperpolarizing current pulses elicit a slowly activating inward current (Ih). Bath administration of serotonin (10 μM) increases the amplitude of this current (symbols).Vh = −40 mV, steps to −50 mV, −65 mV, and −80 mV. Holding current at −40 mV = 220 pA. The time constants at −65 mV were τ1 = 189 ± 20, τ2 = 1820 ± 170; the time constants at −80 mV were τ1 = 109 ± 1, τ2 = 1043 ± 10). B, voltage dependence of Ih determined in this same cell. Serotonin shifts the voltage dependence of this current in the depolarizing direction by approximately 8 mV. Data illustrated in this plot were normalized to the maximal Ih amplitude.
Ih in the ADn displays many common characteristics of this current found in other regions. We determined the voltage activation of this current in the ADn by measuring Ih at each voltage tested and normalizing these to the greatest magnitude determined. We estimated the half-activation (V0.5) of Ih by fitting the data with the Boltzmann equation (see Materials and Methods). The average V0.5 of activation under control conditions was −81 ± 1.3 mV (Fig.2, n = 12 cells). Furthermore, the time constant of Ih activation increased with increasing hyperpolarization and could be fit by a second-order exponential decay equation. These results are consistent with the properties of Ih seen in other studies (Banks et al., 1993; Solomon and Nerbonne, 1993).

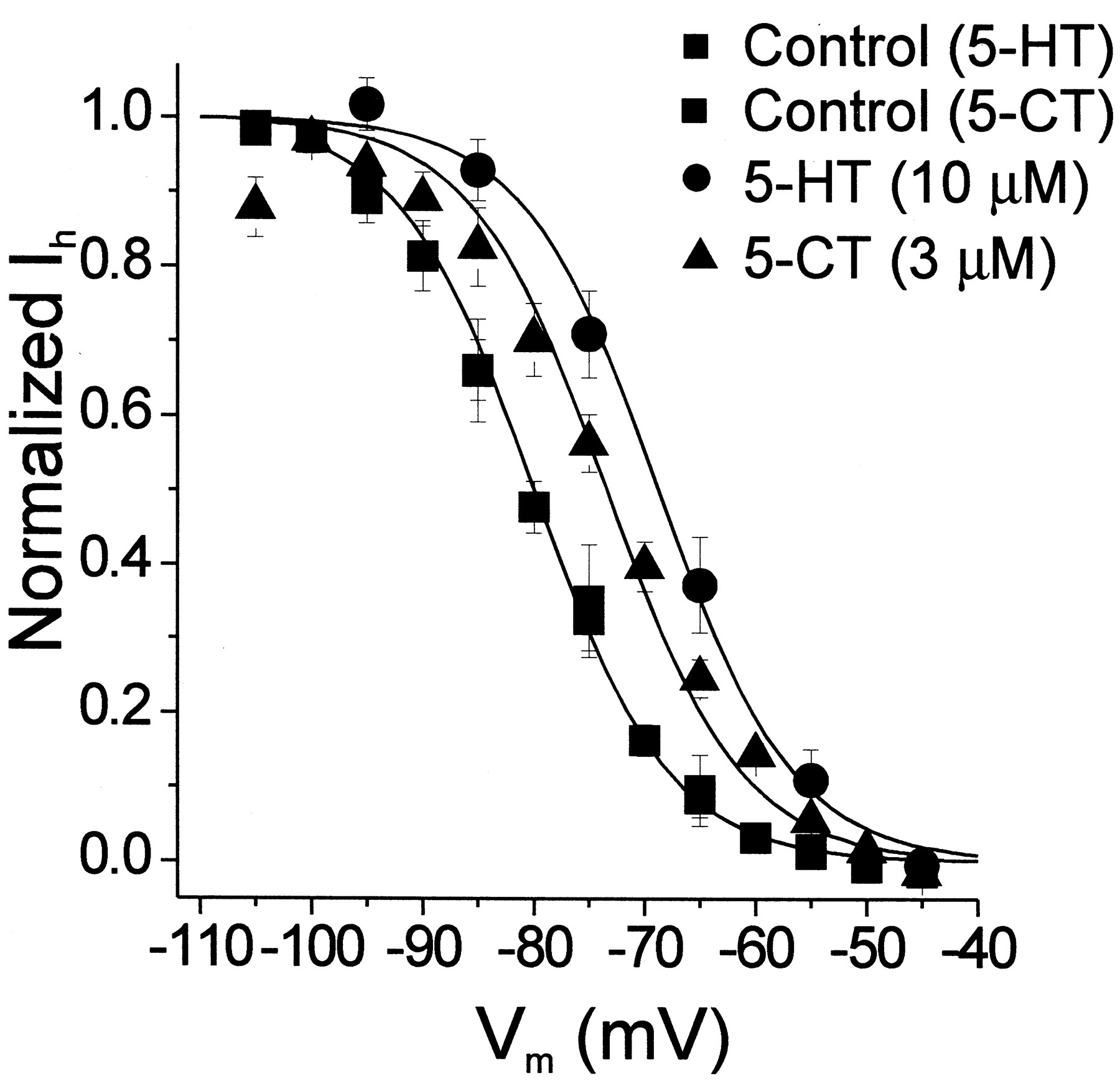
Effects of 5-HT and 5-CT on the voltage dependence of Ih. Bath administration of 5-HT (10 μM) shifts the Ih activation curve in the depolarizing direction by approximately 10 mV. Bath administration of 5-CT (3 μM) similarly shifts the voltage dependence of Ih by approximately 7 mV (n = 4 cells). All of the 5-CT experiments and one of the 5-HT experiments were conducted in the presence of 10 μM cyanopindolol. ▪, controls (n = 7 cells for 5-HT and n = 6 cells for 5-CT); ●, 5-HT (n = 7 cells); and ▴ 5-CT (n= 6 cells). Error bars represent the S.E.M.
We next examined the effect of bath application of 5-HT on the activation curve for Ih. Bath application of 5-HT (10 μM) induced an approximately 10 mV depolarizing shift in the voltage dependence of activation of this current (Figs. 1 and 2). This shift in the voltage dependence of activation is evident in the raw traces as an increase in the amplitude of Ih at more depolarized potentials (Fig. 1A). TheV0.5 for Ih in the presence of serotonin (10 μM) was −71 ± 2.3 mV (n = 7 cells). Thus, 5-HT increases Ih at resting membrane potentials by shifting its voltage dependence.
Pharmacology of the 5-HT-Induced Shift in the Voltage Dependence of Ih Activation.
To test a possible role for the 5-HT7 receptor subtype in mediating the serotonin-induced shift in the voltage dependence of Ih, we used agonists and antagonists capable of distinguishing between serotonin receptor subtypes. One of the defining characteristics of the 5-HT7 receptor is its sensitivity to 5-carboxamidotryptamine (5-CT; Lovenberg et al., 1993; Ruat et al., 1993). Therefore, we first tested the effects of this compound on Ih. As illustrated in Fig. 2, bath administration of 5-CT (3 μM) mimicked 5-HT and elicited a shift in the voltage dependence of Ih(V0.5 = −74 ± 1.5 mV,n = 6 cells; t test, p < 0.01). A second pharmacological characteristic of the 5-HT7 receptor is its sensitivity to 8-hydroxydipropylaminotetralin (8-OHDPAT). This ligand has a high affinity for the 5-HT1A receptor (Hoyer et al., 1994) and a moderate affinity for the 5-HT6 and 5-HT7 receptors (Shen et al., 1993), yet has a negligible affinity for most other serotonin receptor subtypes. Bath application of 100 μM 8-OHDPAT to neurons of the ADn had no detectable effect on the voltage dependence of Ih, but effectively blocked the response to 5-HT (n = 4 cells, ANOVA F = 0.14,p > 0.5 not shown). Finally, the 5-HT7 receptor also exhibits very high affinity for LSD, a compound that appears to function as a low intrinsic activity partial agonist when tested on 5-HT7receptors expressed in heterologous systems (Ruat et al., 1993). Bath administration of LSD (1 μM) had no detectable effect on the voltage dependence of Ih, but blocked the effect of 5-HT (n = 2 cells). These results exclude several 5-HT receptor subtypes, including most 5-HT1receptors, as well as receptors of the 5-HT2, 5-HT3, and 5-HT4 subtypes. As such, they are consistent with the possible involvement of a receptor of the 5-HT7 subtype, but they do not rule out the involvement of receptors of the 5-HT1A, 5-HT5, or 5-HT6 subtypes.
Mesulergine and cyanopindolol are useful agents for identifying 5-HT7 receptors in functional assays. Mesulergine exhibits low nanomolar affinity for the 5-HT7receptor, but micromolar affinity for those of the 5-HT5 and 5-HT6 subtypes (Hoyer et al., 1994) and therefore can be used to distinguish 5-HT7 from 5-HT5 and 5-HT6 receptors. Cyanopindolol, in contrast, exhibits high affinity for the 5-HT1A receptor, but negligible affinity for receptors of the 5-HT7 subtype. Therefore, we next tested both of these antagonists. Bath application of cyanopindolol (10 μM) by itself had no detectable effect on the voltage dependence of Ih and failed to antagonize the effects of 5-HT or 5-CT (Fig. 2, n = 4 cells). This failure was unlikely to reflect inadequate penetration of cyanopindolol into the slice because when administered at this same concentration to hippocampal slices, under essentially identical recording conditions, it readily inhibits 5-HT1A responses (Chapin and Andrade, 2001). In contrast to the lack of effect of cyanopindolol, bath administration of mesulergine (10 μM) completely blocked the 5-CT-induced shift in the voltage dependence of activation of Ih (Fig. 3A). These results are inconsistent with the involvement of 5-HT1A, 5-HT5, or 5-HT6 receptors and suggested the involvement of a receptor of the 5-HT7 subtype. More definitive characterization was performed using the selective 5-HT7 antagonist SB-269770. Bath administration of this compound (1–10 μM) blocked the shift in the voltage dependence of activation induced by 5-HT (Fig. 3B). This indicates that 5-HT modulates Ih through the 5-HT7 receptor subtype.

Mesulergine and SB-269770 block the ability of serotonin to elicit a shift in the voltage dependence of Ih. A, bath administration of mesulergine (10 μM) completely antagonizes the ability of 5-CT (3 μM) to elicit a shift in the voltage dependence of Ih. The mesulergine experiment was conducted in the presence of 10 μM cyanopindolol;n = 3 cells tested. B, similarly, bath administration of SB-269770 (3–10 μM) is without effect on the voltage dependence of Ih, but antagonizes the effect of serotonin (10 μM). n = 4 cells. Error bars represent the S.E.M.
Molecular Mechanism of the Shift in the Voltage Dependence of Ih Activation.
Previous studies have shown that 5-HT7 receptors couple to stimulation of adenylate cyclase and the cAMP signaling pathway (Lovenberg et al., 1993; Meyerhof et al., 1993; Shen et al., 1993;Heidmann et al., 1998). Hence, a 5-HT7-mediated response may be signaled by increases in cAMP. Therefore, we examined the possible involvement of cAMP in signaling the effect of 5-HT in the ADn. To test this possibility, we added 1 mM cAMP directly into the intracellular solution used for whole-cell recording. Intracellular perfusion of cAMP produced a large shift in the voltage dependence of Ih (Fig. 4), such that the V0.5 of Ih under these conditions was shifted to −66.4 mV ± 1.8 mV (n = 4 cells tested). This value was significantly different from the V0.5obtained without cAMP in the intracellular recording solution (ANOVA,F = 11.06, p < 0.001, control versus cAMP, Tukey's post hoc test, p < 0.01). This shift in voltage dependence was associated with a significant increase in holding current at near-rest membrane potentials (−60 to −70 mV; Fig.4). Thus, cAMP mimics the effects of serotonin on Ih in the ADn. This is consistent with the well known regulation of Ih by cAMP (Bobker and Williams, 1989; Pape and McCormick, 1989; DiFrancesco and Tortora, 1991).

cAMP mimics and occludes the ability of 5-HT to elicit a shift in the voltage dependence of Ih. A, when 1 mM cAMP is included in the intracellular recording solution, the voltage dependence of Ih is shifted in the depolarizing direction with respect to that obtained under control conditions by approximately 15 mV. Administration of serotonin (10 μM) under these conditions does not elicit any further shift in the voltage dependence of this current. The control and 5-HT curves have been redrawn from Fig. 2 for illustration purposes. ■, control (n = 7 cells); ○, 5-HT (n = 7 cells); and ▪, cAMP (1 mM) added to the recording solution (n = 4 cells); ●, 5-HT tested in the presence of raised intracellular cAMP (n = 4 cells). B, box chart summarizing the effects of cAMP and 5-HT illustrated in A. Inclusion of cAMP in the recording solution elicited a significant shift in theV0.5 of activation for Ih with respect to control. Box represents mean. Error bars represent S.E.M.
If serotonin signaled its effect on Ih in the ADn through cAMP, then the effect of serotonin should be occluded by inclusion of cAMP in the electrode. Therefore, we examined the ability of cAMP to block the 5-HT-induced shift in the voltage dependence of Ih after intracellular perfusion with cAMP. As illustrated in Fig. 4, under these conditions, serotonin failed to produce any further shift in the voltage dependence of activation of Ih. In fact, theV0.5 for Ihrecorded in the presence of cAMP in the electrode was unaltered by the application of 10 μM serotonin (cAMP = 66 ± 1.8 mV; serotonin in cAMP 66 ± 2.2 mV). Hence, cAMP occluded a further response to 5-HT. These results are consistent with 5-HT shifting the voltage activation of Ih via cAMP in the ADn.
Intracellular cAMP can regulate Ih channels directly (DiFrancesco and Tortora, 1991) by binding to the channel itself (Santoro et al., 2000), or indirectly via PKA through an unidentified mechanism (Accili et al., 1997). To distinguish between these two possibilities, we used the relatively broad-spectrum protein kinase inhibitors staurosporine (1 μM) and H-8 (200 μM). To ensure that the inhibitors would reach equilibrium, we preincubated the slices with the inhibitors for 1 to 6 h before recording. These experimental manipulations have been shown previously to be effective in blocking PKA-mediated responses in brain slices (Chang et al., 1991,1995; Torres et al. 1995). We found that staurosporine and H-8 both failed to inhibit the ability of 5-HT to induced a shift in the voltage dependence of Ih (p > 0.5, Fig.5). These results suggest that the 5-HT shift in the voltage dependence of Ih does not require activation of PKA in the ADn.

Effect of the protein kinase inhibitors staurosporine and H-8 on the ability of 5-HT to shift the voltage dependence of Ih. Slices were incubated in either staurosporine or H-8 for at least 1 h to ensure penetration of the blocker to the cell. This protocol has been shown previously to be effective in inhibiting PKA in brain slices. Administration of 5-HT (10 μM) elicited a significant shift in the voltage dependence of Ih even in the presence of the protein kinase inhibitors (p < 0.05). Each point is the average of three cells in either control or in the presence of 5-HT. ▪, Control; ●, 5-HT.
The Role of Ih in the 5-HT7-Induced Inward Current.
In the last set of experiments, we wished to ascertain what role the modulation of Ih played in the 5-HT7 receptor-mediated inward current described in the previous paper (Chapin and Andrade, 2001). To determine how much of the 5-HT7-induced inward current is due to modulation of Ih, we took advantage of the selective Ih blocker ZD7288 (BoSmith et al., 1993). As illustrated in Fig. 6A, administration of ZD7288 (25–100 μM) greatly reduced, but did not completely block, the 5-HT7 receptor-induced inward current (Fig. 6). In a group of four cells tested using this protocol, the modulation of Ih accounted for 71% ± 6% of the 5-HT-induced inward current (Fig. 7). The residual 5-HT7-induced inward current that remained after ZD7288 was unlikely to be an effect on Ih since ZD7288 completely blocked Ih measured using a hyperpolarizing pulse (Fig. 6B, inset). From these experiments, we conclude that an effect on Ih accounts for a large fraction, but not all, of the 5-HT7-induced inward current.

Effect of the Ih blocker ZD7288 on the 5-HT7 receptor-induced inward current. A, administration of 5-HT (10 μM) elicits an inward current in an ADn neuron. We have previously shown that this effect is mediated by the activation of receptors of the 5-HT7 subtype (Chapin and Andrade, 2001).Vh = −60 mV, Iholding = 20 pA. Bath administration of ZD7288 (100 μM) greatly reduces the inward current elicited by a second administration of serotonin to the same cell. This inhibition does not reflect desensitization, because this response to serotonin does not show detectable desensitization under our recording conditions (Chapin and Andrade, 2001). B, quantitative comparison of the 5-HT7receptor-mediated inward current recorded under control conditions or in the presence of ZD7288 (25–100 μM). ZD7288 significantly reduced the effect of serotonin (n = 4 cells,p <0 .05) and completely blocked Ihestimated using a hyperpolarizing step from −40 to −80 mV (inset,p < 0.001).
Discussion
In the preceding study, we showed that 5-HT7receptor stimulation induces a membrane depolarization and inward current in neurons of the ADn. In the present study, we examined the possibility that this inward current may result from a modulation of Ih. We found that 5-HT in the ADn shifts the voltage dependence of activation of Ih and that the pharmacology of the receptor mediating this effect has the characteristics of a 5-HT7 receptor. In addition, we found that, consistent with the involvement of a 5-HT7 receptor in this response, the effect on Ih is likely mediated via cAMP. Finally, we found that the selective Ih blocked ZD7822 greatly reduced the 5-HT7 receptor-mediated inward current seen in ADn neurons. Combined, these results suggest that 5-HT7 receptors signal a depolarization in the ADn primarily by increasing Ih through a cAMP-dependent mechanism.
Pharmacology of the 5-HT-Induced Modulation of Ih.
Previous studies in a variety of central neurons have described a serotonin-induced depolarization mediated by an increase in Ih (Bobker and Williams, 1989; Pape and McCormick, 1989; Takahashi and Berger, 1990; Larkman and Kelly, 1992;Spain, 1994). Many of these same studies have shown that this effect is mediated through cAMP. Therefore, since 5-HT7receptors are known to couple to Gαs, it seemed reasonable to hypothesize that Ih could mediate at least part of the 5-HT7-induced depolarization in the ADn.
To begin examining this possibility, we first ascertained whether neurons of the ADn expressed Ih. Hyperpolarizing pulses revealed the presence of a slowly developing inward current that was sensitive to ZD7288, thus identifying this current as Ih. The expression of Ih in these cells is consistent with the expression of Ih channel subunits in the ADn (Santoro et al., 2000). Then, we examined whether serotonin regulated Ih in the ADn. Bath administration of serotonin increased Ih by shifting its voltage sensitivity in the depolarizing direction by approximately 10 mV. This effect is similar to, albeit considerably larger than, that seen in other thalamic nuclei (Pape and McCormick, 1989).
Next, we examined whether the pharmacology of this effect on Ih was consistent with the involvement of a 5-HT7 receptor. The ability of serotonin to shift the voltage dependence of Ih was mimicked by 5-CT and inhibited by 8-OHDPAT and LSD, whereas the effect of 5-CT was antagonized by mesulergine, but not by cyanopindolol. This is the pharmacological profile expected for a receptor of the 5-HT7 subtype. The only potential discrepancy is that 8-OHDPAT and LSD have been reported to be partial agonists at the 5-HT7 receptor (Lovenberg et al., 1993; Ruat et al., 1993), whereas they function as antagonists at the receptor regulating Ih in the ADn. This discrepancy could easily reflect differences in the strength of receptor G-protein coupling. LSD exhibits very low intrinsic activity at 5-HT7receptors even when heterologously expressed in Chinese Hamster ovary cells (Ruat et al., 1993) and can be expected to function as an antagonist in a more weakly coupled system. Similarly, 8-OHDPAT has been reported to exhibit a reasonably high intrinsic activity when tested in heterologous expression systems (50–90%; Lovenberg et al., 1993; Plassat et al., 1993; Adham et al., 1998), but a lower intrinsic activity when tested on natively expressed 5-HT7receptors (Hirst et al., 1997; Thomas et al., 1999; Chapin and Andrade, 2001). If the 8-OHDPAT-induced shift in theV0.5 of Ih was similar in magnitude to its effect on the 5-HT7-induced inward current (approximately 20% of the maximal 5-HT response), then the response would be below the ability of our assay to detect agonist activity. As such, these results support the involvement of a 5-HT7 receptor in mediating the effects of serotonin on Ih in the ADn.
The results above exclude some receptors, most notably those of the 5-HT1, 5-HT2, 5-HT3, and 5-HT4 subtypes. Furthermore, the effectiveness of mesulergine, which is more than 100-fold selective for the 5-HT7 receptor versus 5-HT5 and 5-HT6 (Hoyer et al., 1994), strongly argues against the involvement of a receptor of the 5-HT5 or 5-HT6 subtype. Additional evidence supporting the involvement of 5-HT7 receptors in mediating the effect of 5-HT on Ih was obtained with the selective 5-HT7 antagonist SB-269770 (Bacon and Beck, 2000;Lovell et al., 2000), which blocked the 5-HT-induced response. Unfortunately, the shift in voltage dependence of Ih does not lend itself to the kind of quantitative pharmacological analysis required for a definitive receptor subtype identification. Therefore, we explored additional avenues to test the possible involvement of 5-HT7receptors in the regulation of Ih in the ADn.
Signal Transduction Mechanism of the 5-HT7-Induced Shift in the Voltage Dependence of Activation of Ih.
Because the 5-HT7 receptor is coupled to Gαs and hence adenylate cyclase and cAMP production (Adham et al., 1998), it seemed reasonable to expect that responses signaled by a 5-HT7 receptor be mediated via cAMP. In the present study, we found that cAMP can mimic and occlude the serotonin-induced shift of the voltage activation of Ih. This is consistent with a response that is mediated through the cAMP-signaling cascade. This is in accord with previous studies showing that cAMP can regulate Ih (DiFrancesco and Tortora, 1991) and that serotonin regulates Ih through cAMP in other brain areas (Bobker and Williams, 1989; Pape and McCormick, 1989).
Previous studies have shown that Ih can be regulated by cAMP through PKA-independent, as well as PKA-dependent mechanisms. In their now classic work, DiFrancesco and Tortora (1991)demonstrated that, in the heart, cAMP directly acts on the Ih channel to shift the voltage dependence of activation. This conclusion has been substantiated by the cloning of four Ih channel subunits (HCN1–4), each containing a cAMP binding domain in their sequence (Gauss et al., 1998;Ludwig et al., 1998, 1999; Santoro et al., 1998). In addition, parallel studies have shown that protein kinase inhibitors can reduce, and protein phosphatase inhibitors can increase, the magnitude of Ih current (Chang et al., 1991; Accili et al., 1997). This modulation by phosphorylation seems to be different than the direct modulation by cAMP and may reflect the participation of HCN channel subunits currently not cloned, or alternatively, a separate means of modulation of Ih channels. Given these two modes of regulation, we tested whether cAMP could be activating a protein kinase, presumably PKA, to increase Ih. To this effect, we used the kinase inhibitors H-8 and staurosporine. These nonselective inhibitors would not allow us to identify the protein kinase involved in the response, but a response that relies on PKA activation should be blocked by these inhibitors (Chang et al., 1991; Torres et al., 1995). In the present experiments, 5-HT could still shift the voltage activation curve of Ih to more depolarized potentials even in the presence of these inhibitors. These results suggest that 5-HT modulates the Ihcurrent in the ADn through a cAMP-dependent, but PKA-independent, mechanism. Similar conclusions have been reached by others working in other regions of the brain (Bois et al., 1997; Larkman and Kelly, 1997).
Interestingly, 5-HT7 receptors have also been recently reported to regulate calcium-activated potassium currents (AHP) in the CA3 region of the hippocampus (Bacon and Beck, 2000). It is known that cAMP and PKA mediates the ability of 5-HT to reduce the AHP in the hippocampus (Torres et al., 1995). Thus, this suggests the possibility that 5-HT7 receptors may signal their effects in the brain through PKA-dependent and PKA-independent mechanisms.
Ih and the 5-HT7 Receptor-Mediated Inward Current.
The above experiments suggested that serotonin increased Ih in the ADn through a cAMP-dependent mechanism, and that the receptor involved exhibited a pharmacological profile consistent with a 5-HT7 receptor. As such, these experiments suggested, but did not prove, that 5-HT7 receptors elicited a depolarization/inward current in the ADn by increasing Ih. To directly test this possibility, we used the selective Ihblocker ZD7822. Administration of this inhibitor reduced the 5-HT7 receptor-mediated inward current by about three-fourths. These results indicated that Ihchannels carry a large proportion of the 5-HT7receptor-induced inward current. Interestingly, a small residual current remained even after the Ih channels were blocked. This indicates that a second ionic mechanism may depolarize neurons of the ADn, but it is not clear whether this second component represents another receptor subtype or simply a second ionic mechanism by which the 5-HT7 receptor can alter cell properties. This uncertainty notwithstanding, the results outlined above identify the receptor signaling the increase in Ih in the ADn as belonging to the 5-HT7 subtype, and suggest that this effect on Ih accounts for most of the depolarization seen in the ADn in response to serotonin.
Serotonin has been shown previously to elicit a membrane depolarization by increasing Ih in a variety of neurons (Bobker and Williams, 1989; Pape and McCormick, 1989; Takahashi and Berger, 1990; Larkman and Kelly, 1992; Spain, 1994; Cardenas et al., 1999). However, the serotonin receptor subtype responsible for these actions has remained controversial. In most of these studies, the receptor has been reported to resemble the 5-HT1 subtype, although a more recent study on dorsal root ganglion cells has also suggested the possible involvement of a 5-HT7receptor (Cardenas et al., 1999). Such discrepancies are perhaps not unexpected, given the complex pharmacology of the 5-HT7 receptor. In the present study, we have used several distinct approaches to identify the receptor regulating Ih in the ADn. These approaches converge in identifying the serotonin receptor regulating Ihin the ADn as belonging to the 5-HT7 subtype. Admittedly, it is difficult to extrapolate with certitude from the ADn to other brain areas. However, these results nevertheless suggest that the “orphan” serotonin receptor regulating Ihin the brain corresponds to the 5-HT7 receptor.
Acknowledgment
We thank Dr. S. Haj-Dahmane for advice on the experiments.
Footnotes
-
Send reprint requests to: Dr. Rodrigo Andrade, Department of Psychiatry and Behavioral Neurosciences, Wayne State University School of Medicine, 2309 Scott Hall, Detroit, MI 48201. E-mail:randrade{at}med.wayne.edu
-
This work was supported in part by a research grant (Joe Young, Sr.) from the State of Michigan and by Grant MH43985 from the National Institute of Mental Health.
- Abbreviations:
- 5-HT
- 5-hydroxytryptamine
- ADn
- anterior dorsal nucleus of the thalamus
- Ih
- hyperpolarization-activated nonselective cation current
- 5-CT
- 5-carboxamidotryptamine
- 8-OHDPAT
- 8-hydroxydipropylaminotetralin
- PKA
- protein kinase A
- Received September 14, 2000.
- Accepted December 20, 2000.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}