


Abstract
Mibefradil is a Ca2+ channel antagonist that inhibits both T-type and high-voltage-activated Ca2+ channels. We previously showed that block of high-voltage-activated channels by mibefradil occurs through the production of an active metabolite by intracellular hydrolysis. In the present study, we modified the structure of mibefradil to develop a nonhydrolyzable analog, (1S, 2S)-2-(2-(N-[(3-benzimidazol-2-yl)propyl]-N-methylamino)ethyl)-6-fluoro-1,2,3,4-tetrahydro-1-isopropyl-2-naphtyl cyclopropanecarboxylate dihydrochloride (NNC 55-0396), that exerts a selective inhibitory effect on T-type channels. The acute IC50 of NNC 55-0396 to block recombinant α1G T-type channels in human embryonic kidney 293 cells was ∼7 μM, whereas 100 μM NNC 55-0396 had no detectable effect on high-voltage-activated channels in INS-1 cells. NNC 55-0396 did not affect the voltage-dependent activation of T-type Ca2+ currents but changed the slope of the steady-state inactivation curve. Block of T-type Ca2+ current was partially relieved by membrane hyperpolarization and enhanced at a high-stimulus frequency. Washing NNC 55-0396 out of the recording chamber did not reverse the T-type Ca2+ current activity, suggesting that the compound dissolves in or passes through the plasma membrane to exert its effect; however, intracellular perfusion of the compound did not block T-type Ca2+ currents, arguing against a cytoplasmic route of action. After incubating cells from an insulin-secreting cell line (INS-1) with NNC 55-0396 for 20 min, mass spectrometry did not detect the mibefradil metabolite that causes L-type Ca2+ channel inhibition. We conclude that NNC 55-0396, by virtue of its modified structure, does not produce the metabolite that causes inhibition of L-type Ca2+ channels, thus rendering it more selective to T-type Ca2+ channels.
Voltage-gated Ca2+ channels are transmembrane proteins involved in the regulation of cellular excitability and intracellular Ca2+ signaling. Calcium channels have been divided into various categories based on functional and pharmacological criteria. High-voltage-activated (HVA) channels, which have been further subdivided into L-, N-, P/Q-, and R-types, require strong depolarizations for activation, whereas low-voltage-activated or T-type channels activate over a much more negative voltage range and exhibit unique inactivation and deactivation kinetics (Armstrong and Matteson, 1985; Catterall, 1998; Perez-Reyes, 1998). The main structural component of the voltage-gated calcium channel is the α1 subunit, which forms the pore and the channel gates. Molecular cloning has identified 10 α1 subtypes. α1A–α1E and α1S encode HVA channels, whereas α1G–α1I encode T-type channels. Pharmacological agents that act selectively on α1 subtypes have been key in studying calcium channel function in many physiological systems; however, a selective antagonist of T-type Ca2+ channels is not yet available.
Mibefradil, a tetralol derivative chemically distinct from other Ca2+ channel antagonists, has been reported to block T-type Ca2+ channel currents in many tissues, including heart (Madle et al., 2001), brain (McDonough and Bean, 1998), and vascular smooth muscle (Bian and Hermsmeyer, 1993; Mishra and Hermsmeyer, 1994; Schmitt et al., 1995). Mibefradil was also reported to block HVA Ca2+ channels, including α1A, α1B, α1C, and α1E (Bezprozvanny and Tsien, 1995; Jiménez et al., 2000). We previously demonstrated that mibefradil potently blocked HVA Ca2+ channels in a rat insulin-secreting cell line (INS-1 cells) via a mechanism involving intracellular hydrolysis of mibefradil to produce an active metabolite (Wu et al., 2000).
The cardiovascular effects of mibefradil are interesting; for instance, it decreases heart rate without a negative inotropic effect (Osterrieder and Holck, 1989; Cremers et al., 1997), and its action is not associated with the reflex activation of neurohormonal and sympathetic systems (Ernst and Kelly, 1998). These properties differ from other clinically important Ca2+ antagonists such as nifedipine, diltiazem, and verapamil, which in the heart selectively inhibit L-type (α1C) Ca2+ channels. However, it is unclear whether the unique effects of mibefradil are caused by the blockade of T-type Ca2+ channels since mibefradil also blocks the L-type Ca2+ currents in the cardiomyocytes (Leuranguer et al., 2001). Therefore, identifying a more selective T-type Ca2+ channel antagonist will be useful for the study of cardiac voltagegated calcium channels and may promote the development of a new class of therapeutically beneficial compounds.
In a previous study, we demonstrated that hydrolysis of the ester side chain of mibefradil resulted in a compound (des-methoxyacetyl mibefradil or Ro 40-5966) that exhibited an L-type Ca2+ channel-specific inhibitory effect (Wu et al., 2000). We thus proposed that modifications in this ester side chain, which decreased hydrolysis, might result in compounds with a lower potency in blocking L-type Ca2+ channels and selective action on T-type channels. To test this hypothesis, we examined the effects of several novel mibefradil derivates on T-type and HVA Ca2+ currents in whole-cell voltage-clamp recordings. We used HEK 293 cells stably transfected with the α1G subtype of T-type Ca2+ channel (HEK/α1G) and INS-1 cells to test the effects of our new compounds on T-type and HVA Ca2+ channels, respectively. INS-1 cells express a variety of HVA Ca2+ currents, including P/Q-, N-, L-, and R-types (Horvath et al., 1998), with α1D contributing most of the currents (Liu et al., 2002; Huang et al., 2004).
Materials and Methods
Cell Culture. INS-1 cells, an insulin-secreting cell line derived from rat pancreatic β-cells (Asfari et al., 1992) were cultured in RPMI 1640 medium containing 10% fetal bovine serum, 100 U/ml penicillin, 100 μg/ml streptomycin, and 50 μM mercaptoethanol in an atmosphere of 5% CO2 in air at 37°C for 2 to 5 days before recording.
Creation of HEK 293 Cell Lines Stably Expressing Recombinant T-Type Ca2+ Channels. An α1G cDNA originally cloned from rat pancreatic β-cells (Zhuang et al., 2000) in vector pcDNA3.1 hygro(-) (Invitrogen, Carlsbad, CA) was transfected into HEK cells using the FuGENE kit (Roche Diagnostics, Indianapolis, IN). Cell lines stably expressing Cav3.1 were obtained after transfection using standard cell cloning techniques (Freshney, 1983). HEK 293 cells stably transfected with α1G cDNA (HEK 293/α1G) were incubated in Dulbecco's modified Eagle's medium, supplemented with 10% fetal bovine serum, 100 U/ml penicillin, 100 μg/ml streptomycin, and 0.5 mg/ml hygromycin-B in an atmosphere of 5% CO2 in air at 37°C for 2 to 5 days before recording.
Electrophysiological Recording. Whole-cell recordings were carried out by the standard “giga-seal” patch-clamp technique. The whole-cell recording pipettes were made of hemocapillaries (Warner Instrument, Hamden, CT), pulled by a two-stage puller (PC-10; Narishige, Greenvale, NY), and heat-polished with a microforge (MF-200; World Precision Instruments, Sarasota, FL) before use. Pipette resistance was in the range of 2 to 5 MΩ in our internal solution. The recordings were performed at room temperature (22–25°C). Electrical currents were recorded using an EPC-9 patch-clamp amplifier (HEKA, Lambrecht/Pfalz, Germany) and filtered at 2.9 kHz. Data were acquired with PULSE/PULSEFIT software (HEKA). Voltage-dependent currents were corrected for linear leak and residual capacitance by using an on-line P/n subtraction paradigm. In the whole-cell configuration, T-type Ca2+ currents were recorded at -20 or -10 mV when the holding potential was -70 mV. The HVA Ca2+ current was measured at 10 mV, with a holding potential of -40 mV.
Solutions. The extracellular solution used in the whole-cell Ca2+ current recording contained 10 mM CaCl2, 110 mM tetraethylammonium-Cl, 10 mM CsCl, 10 mM HEPES, 40 mM sucrose, 0.5 mM 3,4-diaminopyridine, pH 7.3. The intracellular solution contained 130 mM N-methyl-d-glucamine, 20 mM EGTA (free acid), 5 mM 1,2-bis(2-aminophenoxy)ethane-N,N,N′,N′-tetraacetate, 10 mM HEPES, 6 mM MgCl2, and 4 mM Ca(OH)2, with pH adjusted to 7.4 with methanesulfonate. Mg-ATP (2 mM) was included in the pipette solution to minimize rundown of L-type Ca2+ currents. For perforated-patch experiments, 200 μg/ml nystatin was used. The pipette was filled with nystatin-containing intracellular solution, and gentle suction was used to achieve gigaohm resistance. The access resistance gradually decreased within 5 min after the gigaohm-seal formation, and then currents were recorded after stabilization. The extracellular solution contained 26 mM sucrose, 130 mM tetraethylammonium-Cl, 10 mM HEPES, 5 mM KCl, 2 mM CaCl2, 2 mM MgCl2, pH 7.3. The pipette solution contained 65 mM CsOH, 65 mM Cs-methanesulfonate, 20 mM sucrose, 10 mM HEPES, 10 mM MgCl2, and 1 mM Ca(OH)2, with pH 7.4.
Mass Spectrometric Analysis. Mass spectrometric analysis was performed on a PerSeptive Voyager-DE MALDI-TOF instrument (Applied Biosystems, Foster City, CA). Cultured INS-1 cells were treated with 20 μM NNC 55-0396 for various lengths of time under each experimental condition. After incubation, cells were washed with PBS three times, scraped into Eppendorf tubes with 1 ml of PBS, and centrifuged at 1000g for 5 min. The cell pellets were collected and redissolved by sonication (10 s) in 0.5 ml of PBS for mass spectrometric analysis. The sample was taken up in α-cyano-4-hydroxcinnamic acid (Aldrich, Milwaukee, MI), which was used as the matrix. Ten microliters of each sample were mixed with 70 μl of the matrix. One microliter of the mixture was spotted on a plate for analysis on the MALDI-TOF instrument. Several positive ion spectra were recorded in the mass range m/z 820 to 99 at a mass resolution of 1000 and a scan speed of 2 s/decade. For NNC 55-0396, m/z 492 was the dominant ion (M + H). For calibration, a standard solution of 50 μM NNC 55-0396 was subjected to mass spectrometric analysis. The relative amount of NNC 55-0396 was determined by calibrating the intensities of NNC 55-0396 with the intensity of standard solution.
Statistics and Curve Fitting. Nonlinear regression analysis was used to fit concentration-response data to a sigmoid relationship, Y = 100/(1 + 10^((log IC50 - X) slope), where slope is the Hill slope parameter, IC50 is the concentration producing 50% blockage, and X is the drug concentration. Voltage-dependent activation and steady-state inactivation curves were generated by normalizing the currents with the maximal amplitude in each cell and fitting the data with the Boltzmann equation, 1/(1 + exp((V - V1/2)/k), where k represents the slope and V1/2 represents the voltage corresponding to half-activation of the channels. Student's t test was used to compare V1/2 and k values determined from fits of the data with this equation. The data fitting and statistical analysis were performed with Prism version 4 (GraphPad Software, San Diego, CA). In the figures where the data are presented as symbols and error bars, the values are mean ± S.E.M.
General Chemical Procedure. Reagents, starting materials, and solvents were purchased from common commercial suppliers and were used as received. All dry solvents were dried over molecular sieves (0.3 or 0.4 nm). Evaporation was carried out on a rotary evaporator at bath temperatures <40°C and under appropriate vacuum. Flash chromatography was carried out on a Biotage FLASH 40 (Biotage AB, Uppsala, Sweden) using Biotage FLASH columns (KP-SIL 60 Å particle size, 32–63 μM). Melting points were determined with a Büchi B545 apparatus (Büchi, Flawil, Switzerland) and are uncorrected. Proton NMR spectra were recorded at ambient temperature using a Bruker AVANCE DPX 200 (Bruker, Newark, DE) (200 MHz) and DPX 300 (300 MHz), with tetramethylsilane as an internal standard for proton spectra. Chemical shifts are given in ppm (δ), and splitting patterns are designated as follows: s, singlet; d, doublet; dd, double doublet; t, triplet; dt, double triplet; q, quartet; quint, quintet; m, multiplet; and br, broad. The 70-electron-volt EI solid mass spectra were recorded on a Finnigan MAT TSQ 70 mass spectrometer (Thermo Finnigan, San Jose, CA). Reactions were followed by thin layer chromatography performed on silica gel 60 F254 (Merck, Darmstadt, Germany) or ALUGRAM SIL G/UV254 (Macherey-Nagel, Düren, Germany) thin layer chromatography aluminum sheets.
Synthesis of NNC 55-0396. Methoxyacetic acid (2(S)-[2-[N-[3-(2-benzimidazolyl)propyl]-N-methylamino]ethyl]-6-fluoro-1(S)-isopropyl-1,2,3,4-tetrahydro-2-naphthyl ester dihydrochloride) (mibefradil, 0.570 g) in ethanol (96%, 5 ml) and aqueous sodium hydroxide (1 N, 5 ml) was refluxed for 2 h. The cold reaction mixture was concentrated in vacuo. The residue was partitioned between water and dichloromethane. The aqueous layer was extracted with dichloromethane. The combined organic layers were dried with sodium sulfate and concentrated to give 2-(2-[[3-(1-benzimidazol-2-yl)-propyl]methylamino]ethyl)-6-fluoro-1-isopropyl-1,2,3,4-tetrahydro-2-naphthalinol as a clear syrup (100%, 0.43 g). 1H NMR (CDCl3): δ 7.57 (br, 2H); 7.23 (m, 2H); 6.97 (m, 1H); 6.58 (m, 2H); 3.07–2.83 (m, 3H); 2.75 (m, 1H); 2.6 (m, 4H); 2.5–2.2 (s + m, 3H + 3H); 2.06 (quint, 2H); 1.81 (br dd, 1H); 1.50 (m, 2H); 1.20 (d, 3H); 0.53 ppm (d, 3H).
2-(2-{[3-(1-Benzimidazol-2-yl)-propyl]methylamino}ethyl)-6-fluoro-1-isopropyl-1,2,3,4-tetrahydro-2-naphthalinol (0.110 g) was dissolved in 1 ml of dichloromethane. Diisopropylethylamine (0.045 ml) and 0.071 ml of cyclopropanecarbonyl chloride was added. After stirring for 19 h, the reaction mixture aqueous saturated sodium hydrogencarbonate was added. The aqueous layer was extracted twice with dichloromethane. The combined organic layers were dried with sodium sulfate and concentrated in vacuo. The residue was purified by flash chromatography using dichloromethane/methanol 6:1 as an eluent to give the free base as a syrup. This product was dissolved in ethanol, and aqueous hydrochloride (1 N, 0.38 ml) was added. After stirring for 30 min, the mixture was concentrated. The residue was crystallized from ethyl acetate to give the title compound as a white powder (34%, 50 mg). Mp: 134 to 141°C. EI SP/MS: 491 (M+). 1H NMR (DMSO— d6): δ 7.77 (m, 2H); 7.52 (m, 2H); 7.07 (m, 1H); 6.96 (broad d, 2H); 3.3 (m, 2H); 3.2 (m, 4H); 3.0 (m, 2H); 2.9 (m, 1H); 2.67 (s, 3H); 2.45 (m, 1H); 2.38 (m, 2H); 2.15–1.45 (m, 4H); 1.57 (m, 1H); 1.04 (d, 3H); 0.90 (m, 4H); 0.48 ppm (d, 3H).
Synthesis of (1S,2S)-2-(2-(N-[(3-Benzimidazol-2-yl)propyl]-N-methylamino)ethyl)-6-fluoro-1,2,3,4-tetrahydro-1-isopro-pyl-2-naphtyl valeroate dihydrochloride (NNC 55-0395). NNC 55-0395 was prepared by an analogous procedure: Mp 118 to 121°C. EI SP/MS: 507 (M+).1H NMR (DMSO-d6): δ 7.75 (m, 2H); 7.49 (m, 2H); 7.08 (m, 1H); 6.96 (br d, 2H); 3.15 (m, 4H); 2.95 (m, 3H); 2.7 (s, 3H); 2.48 (dt, 2H); 2.23 (m, 2H); 2.0 (m, 4H); 1.50 (p, 2H); 1.35 (quint, 2H); 1.02 (d, 3H); 0.90 (t, 3H); 0.35 ppm (d, 3H).
Synthesis of (1S,2S)-2-(2-(N-[(3-Benzimidazol-2-yl)propyl]-N-methylamino)ethyl)-6-fluoro-1,2,3,4-tetrahydro-1-isopro-pyl-2-naphtyl isobutyrate dihydrochloride (NNC 55-0397). NNC 55-0397 was prepared by an analogous procedure: Mp 114 to 117°C. EI SP/MS: 493 (M+). 1H NMR (DMSO-d6): δ 7.77 (m, 2H); 7.52 (m, 2H); 7.07 (m, 1H); 6.96 (br d, 2H); 3.43 (m, 1H); 3.3-3.05 (m, 5H); 2.97 (m, 2H); 2.88 (m, 1H); 2.70 (s, 3H); 2.61 (m, 1H); 2.45 (m, 1H); 2.3 (m, 2H);2.15–1.85 (m, 4H); 1.15 (d, 6H); 1.00 (d, 3H); 0.45 ppm (d, 3H).
Results
Synthesis of NNC 55-0396 and Other Mibefradil Analogs. We synthesized three analogs of mibefradil by replacing the ester chain (methoxyacetyl) group NNC 55-0395, NNC 55-0396, and NNC 55-0397 (Fig. 1). All compounds were synthesized from mibefradil in two steps. Alkaline hydrolysis of mibefradil and subsequent treatment with valeroyl chloride, cyclopropanecarbonyl chloride, and isobutyryl chloride gave the desired compounds NNC 55-0395, NNC 55-0396, and NNC 55-0397 upon workup, respectively.

Synthesis of NNC 55-0396 and other analogs from mibefradil and chemical structures of NNC 55-0395, NNC 55-0396, and NNC 55-0397. The side chain of mibefradil inside the dashed box is replaced by the new structures indicated by arrows.
Pharmacological-Screening Effect of NNC 55-0395, NNC 55-0396, and NNC 55-0397 on T-Type and HVA Ca2+ Currents. We used perforated whole-cell patch-clamp to examine the effects of NNC 55-0395, NNC 55-0396, and NNC 55-0397 on HVA Ca2+ current in INS-1 cells. In these experiments, the resting membrane potential was held at -40 mV to eliminate T-type Ca2+ current. The inhibitory potency of the mibefradil analogs on HVA Ca2+ current was examined at dosages of 0.1, 1, 10, and 100 μM approximately 10 min after the drug perfusion. The results showed that both NNC 55-0395 and NNC 55-0397 had an inhibitory effect on HVA Ca2+ currents at 100 μM (Fig. 2, A and C), whereas no inhibition on the HVA Ca2+ current was detected with NNC 55-0396 at the same concentration (Fig. 2B).

Pharmacological-screening effect of NNC 55-0395, NNC 55-0396, and NNC 55-0397 on HVA and T-type Ca2+ currents. A, B, and C, dose-dependent effects of the three compounds on HVA currents recorded with perforated whole-cell patch-clamp in INS-1 cells with a holding potential of -40 mV and a test pulse at 10 mV for 200 ms for every 30 to 40 s. The relative current values were obtained by normalizing the current amplitudes with respect to those recorded under the drug-free condition. D, E, and F, dose-dependent effects of the three compounds on T-type Ca2+ currents recorded with whole-cell patch-clamp in HEK 293/α1G cells with a holding potential of -70 mV and a testing potential of -20 mV for 200 ms (n = 3–5 for each set of data).
Next, we determined the effects of NNC 55-0395, NNC 55-0396, and NNC 55-0397 on T-type Ca2+ current. These experiments were conducted by measuring the effects of the compounds on whole-cell current in HEK 293/α1G cells. The dose-dependent inhibition of compounds NNC 55-0395, NNC 55-0396, and NNC 55-0397 on T-type Ca2+ current was examined at dosages ranging from 1 to 100 μM. NNC 55-0396 and NNC 55-0397 blocked more than 50% of the T-type Ca2+ current at 8 μM (Fig. 2, E and F), whereas NNC 55-0395 inhibited less than 50% of the T-type Ca2+ current at 64 μM (Fig. 2D).
Both NNC 55-0395 and NNC 55-0397 blocked HVA Ca2+ channel currents in our screening experiments and were thus eliminated from further characterization. Compound NNC 55-0396, which blocked T-type Ca2+ current but not HVA Ca2+ currents, was selected for subsequent analysis.
Characterization of the Inhibitory Effects of NNC 55-0396 on T-Type Ca2+ Current. To further characterize NNC 55-0396, we used whole-cell patch-clamp and a bath perfusion system to examine its dose-dependent effects on T-type and HVA Ca2+ currents. At 8 μM, NNC 55-0396 reduced more than 50% of the peak of T-type Ca2+ current compared with the control in HEK 293/α1G cells (Fig. 3A); however, NNC 55-0396 at 100 μM did not block the HVA Ca2+ current in INS-1 cells (Fig. 3B), whereas this HVA Ca2+ current could be inhibited by 10 μM nifedipine, a selective L-type Ca2+ channel blocker (Fig. 3C). Pooled data of the effects of NNC 55-0396 on T-type and HVA Ca2+ currents are shown in Fig. 3D. After bathing HEK 293/α1G cells with NNC 55-0396 at 8 μM or bathing INS-1 cells with NNC 55-0396 at 100 μM for over 8 min, T-type Ca2+ currents were inhibited by 60%, whereas no decrease in the HVA Ca2+ current was observed.

The effects of NNC 55-0396 on T-type and HVA Ca2+ channels. A, current traces of T-type Ca2+ currents in an HEK 293/α1G cell before and after application of 8 μM NNC 55-0396. The holding potential was -70 mV, and the test potential was -20 mV. B, current traces of HVA Ca2+ currents in an INS-1 cell before and after 100 μM NNC 55-0396. The currents were recorded at 10 mV for 200 ms when held at -40 with perforated whole-cell patch-clamp. The traces were filtered at 1 kHz. C, Ca2+ current was inhibited by 10 μM nifedipine in an INS-1 cell. The currents were measured at 10 mV before and after nifedipine (Nif) administration with a holding potential of -70 mV. D, time-dependent effects of NNC 55-0396 on HVA and T-type Ca2+ currents. The open circles represent relative currents of HVA Ca2+ channels recorded in INS-1 cells before and after 100 μM NNC 55-0396 administration (n = 6), whereas the solid circles represent relative currents recorded in HEK 293/α1G cells before and after 8 μM NNC 55-0396 administration (n = 4). NNC 55-0396 was applied to the bath 3 min after the beginning of the recording. E, dose-dependent inhibitory effects of NNC 55-0396 (open circles, n = 6) and mibefradil (solid circles, n = 4) on T-type Ca2+ currents recorded in HEK/α1G cells. The smooth lines are consistent with the Hill equation, with IC50 values of 6.8 and 10.08 μM for NNC 55-0396 and mibefradil, respectively. The Hill slopes are -2.39 and -4.26 for NNC 55-0396 and mibefradil, respectively. F, comparison of current densities (pA/pF) among HEK/α1G cells incubated in various concentrations of NNC 55-0396 for 30 to 60 min. The peak-current amplitudes were measured at -20 mV when held at -70 mV (n = 7–11 for each set of data). The asterisk represents p < 0.01 of the data compared with the control (0 nM NNC 55-0396).
The inhibitory potency of NNC 55-0396 on T-type Ca2+ currents in HEK 293/α1G was also compared with that of mibefradil, as shown in Fig. 3E. By fitting the data with a sigmoid dose-response relationship equation, NNC 55-0396 and mibefradil blocked T-type Ca2+ current with IC50 values of 6.8 and 10.08, respectively. This result suggests that NNC 55-0396 retains potency similar to mibefradil as an inhibitor of T-type Ca2+ current.
Long-term exposure of HEK/α1G cells to NNC 55-0396 showed a decrease in current density of T-type Ca2+ channels. Using whole-cell patch-clamp, we measured T-type Ca2+ current density in the HEK/α1G cells that had bathed in the extracellular solutions containing 0, 10, 100, and 1000 nM NNC 55-0396 for 30 to 60 min (Fig. 3F). All peak-current amplitudes and slow capacitances were obtained during the first minute after breaking in; thus, the effect of the frequency-dependent inhibitory effect was minimal. Our data showed a slow yet more potent inhibitory effect of NNC 55-0396 on the T-type Ca2+ currents.
Figure 4A illustrates that there was no significant difference in the conductance-voltage relationship (G/V) curves of T-type Ca2+ currents between the control cells and cells incubated with 8 μM NNC 55-0396. The conductance (G) was calculated from the current (I) divided by the driving force (Vdrive = Vmembrane - Vreversal), where the Vreversal was hypothetically assigned as 70 mV. Curves were generated by fitting the data with the Boltzmann equation. The V1/2 and k values are -32.61 ± 1.0 and 4.16 ± 0.9 for the controls (n = 3) and -32.12 ± 0.8 and 4.33 ± 0.73 for cells incubated with NNC 55-0396 (n = 3), respectively. The steady-state inactivation curves of T-type Ca2+ current were also generated with the Boltzmann equation (Fig. 4B). The V1/2 values are -59.21 ± 0.38 and -62.84 ± 0.94 for the controls (n = 4) and cells incubated with NNC 55-0396 (n = 6), respectively. The slope k values are 4.92 ± 0.33 with a 95% confidence interval between 4.16 to 5.69 for the controls and 7.95 ± 0.72 with a 95% confidence interval between 6.31 to 9.58 for cells incubated with NNC 55-0396, respectively. Therefore, the steady-state inactivation curve of T-type Ca2+ current is flattened when NNC 55-0396 is present.

Effects of NNC 55-0396 on voltage-dependent activation (G/V) and steady-state inactivation of T-type Ca2+ channel. A, voltage-dependent activation (G/V) curves of T-type Ca2+ channel with (solid circles) and without (open circles) the presence of 8 μM NNC 55-0396 were fit by the Boltzmann equation (n = 3). B, steady-state inactivation curves of T-type Ca2+ channel with (solid circles) and without (open circles) the presence of 8 μM NNC 55-0396 were fit by the Boltzmann equation (n = 6). Steady-state inactivation was determined by applying a prestimulating pulse of 1.5 s at various voltages immediately before the test pulse at -20 mV. For both A and B, currents were recorded in HEK 293/α1G cells, and the cell membrane was held at -70 mV.
State-Dependent Block of T-Type Ca2+ Current by NNC 55-0396. The inhibitory mechanism of NNC 55-0396 on T-type Ca2+ current was further investigated by testing the effect of changing the holding potential and stimulation frequency. As shown in Fig. 5A, in the absence of NNC 55-0396, changing the holding potential from -120 to -80 mV did not alter the peak amplitude of T-type Ca2+ current in an HEK 293/α1G cell, elicited by depolarizations to -10 mV every 10 s. In contrast, after the addition of 8 μM NNC 55-0396, switching the membrane potential from -80 to -120 mV caused an increase in the current amplitude, suggesting a partial relief of block at the more hyperpolarized holding potential. Similarly, increasing the pulse rate from 0.05 to 0.5 Hz had little effect on current amplitude without NNC 55-0396, whereas the rate of inhibition of T-type Ca2+ current was accelerated by increasing the frequency of stimulation in the presence of the drug (Fig. 5B). Similar results of both voltage-dependent and frequency-dependent blockade were observed in four experiments.

State-dependent inhibition of T-type Ca2+ channels by NNC 55-0396. A, whole-cell Ca2+ currents were measured at -10 mV when held at -120 and -80 mV, as indicated by the bar above the graph. The pulses were applied every 10 s. B, whole-cell Ca2+ currents were measured at -10 mV when held at -70 mV at stimulation frequencies of 0.05 and 0.5 Hz, as indicated by the bar above the graph. NNC and the arrows indicate the time when NNC 55-0396 was perfused into the bath.
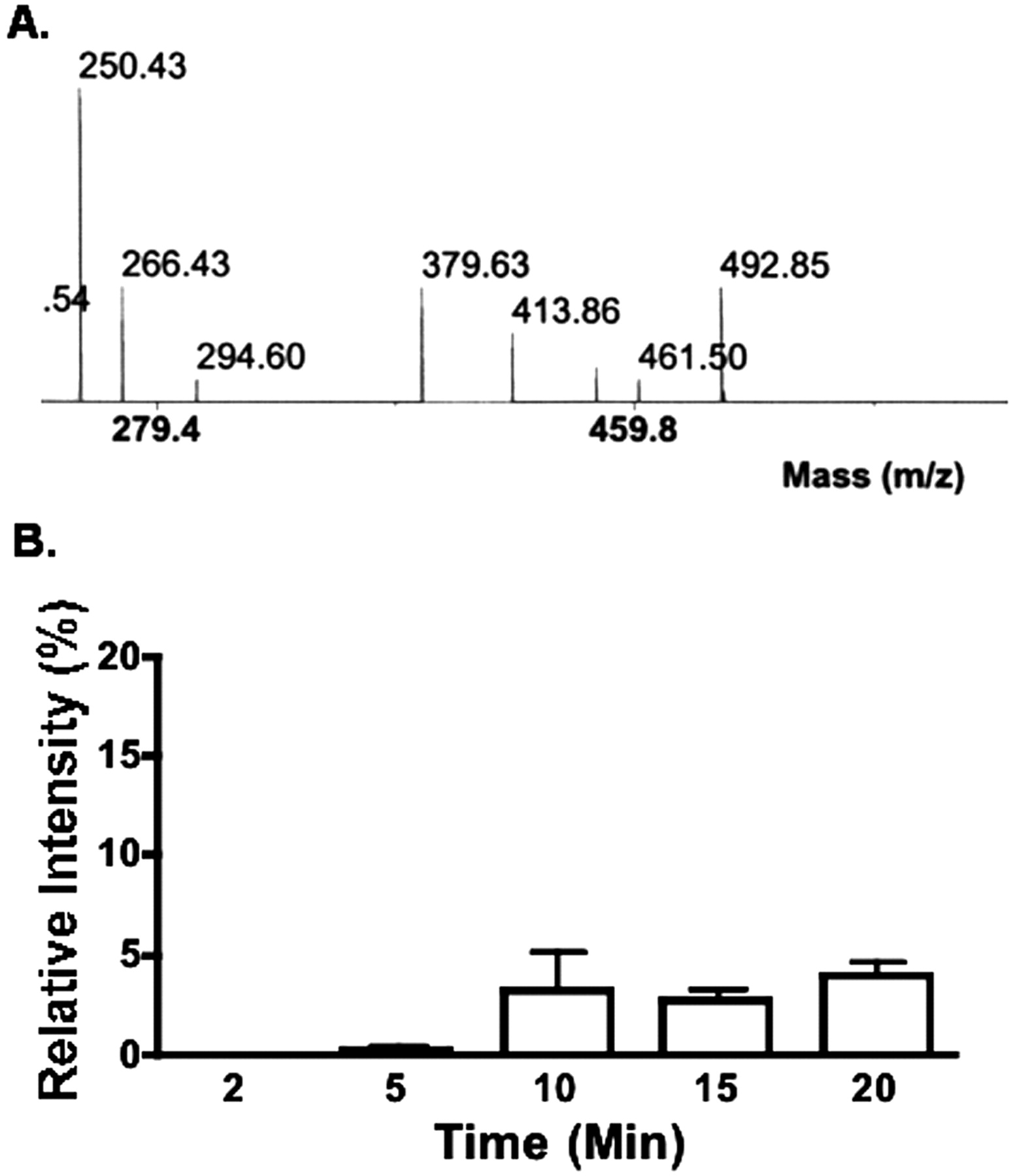
Mass Spectrometric Analysis of NNC 55-0396-Treated INS-1 Cells. Previously, we found that mibefradil blocked both T-type and HVA Ca2+ current in INS-1 cells (Wu et al., 2000). After entering the cells, mibefradil is broken down into metabolites. One of them is des-methoxyacetyl mibefradil, which exerts an inhibitory effect on HVA Ca2+ channels by acting on the channels from inside the cell. To investigate why NNC 55-0396 and mibefradil have different effects on HVA Ca2+ channels, we used mass spectrometric analysis to examine whether des-methoxyacetyl mibefradil was produced intracellularly when the cells were treated with NNC 55-0396. Figure 6A shows a section of mass spectrum of molecules in INS-1 cells that had been incubated with NNC 55-0396 for 20 min. Notably, no des-methoxyacetyl mibefradil (which peaks at 424 m/z) was detected in this preparation. This finding suggests that, unlike mibefradil, the compound NNC 55-0396 is not hydrolyzed into des-methoxyacetyl mibefradil. Thus, NNC 55-0396 does not inhibit HVA Ca2+ channels as mibefradil does. The amount of NNC 55-0396 (which peaks at 492 m/z), however, increased in the cells for longer periods than 10 min as shown in Fig. 6B.

Mass spectrometric analysis of INS-1 cells treated with NNC 55-0396. A, no ester side chain hydrolytic metabolite (peak is predicted at 424 m/z) was detected in INS-1 cells after 30 min of incubation with 20 μM NNC 55-0392, which peaked at 492 m/z. B, time-related relative intensity of NNC 55-0396 in INS-1 cells. Mass spectrometry was performed on the cells that had been incubated in 20 μM NNC 55-0396 for various time periods and then washed three times. The relative intensity is presented as the ratio of the energy power of the laser between the NNC 55-0396 found inside the cells and the standard solution containing 50 μM NNC 55-0396 (n = 4).
Accessibility of NNC 55-0396 on the T-Type Ca2+ Channel. The slow onset of NNC 55-0396 inhibition on T-type Ca2+ current and the increasing NNC 55-0396 accumulation in the cells with time suggest that this compound dissolves in or passes through the plasma membrane to exert its effect. To test this hypothesis, we examined the reversibility of the NNC 55-0396 blockade of T-type Ca2+ current. Whole-cell currents were evoked in HEK 293/α1G cells by test pulses to -10 mV from a holding potential of -70 mV. After establishing a steady current, small volumes of 8 μM NNC 55-0396 were delivered in close proximity to the recording cell with a quartz capillary positioned by a micromanipulator. The drug was washed out after more than a 50% inhibition of T-type Ca2+ current was achieved. As shown in Fig. 7A, the blockade of the T-type Ca2+ current by NNC 55-0396 was poorly reversible during a 10-min washout period in these experiments. Since the NNC 55-0396 blocks T-type Ca2+ current at a relatively slow rate and is poorly reversible by washing out, the drug binding site of the channel may be within transmembrane or intracellular domains of the channel.

Washout and intracellular application of NNC 55-0396. A, inhibitory effect of 8 μM NNC 55-0396 on HEK/α1 G T-type Ca2+ currents was not reversed after perfusion with the drug-free extracellular solution (n = 3). B, effect of intracellular application of NNC 55-0396 on HEK/α1G T-type Ca2+ currents. The currents were recorded by the whole-cell patch-clamp with a pipette solution containing 8 μM NNC 55-0396 (solid circles, n = 10) or a drug-free solution (open circles, n = 6). All recordings were conducted at a holding potential of -70 mV and a test pulse of -20 mV.
If the site of action of NNC 55-0396 is on the intracellular side of the membrane, we would expect that intracellular perfusion of the drug would effectively block T-type Ca2+ current. To test this possibility, whole-cell currents were recorded in HEK 293/α1G cells with 8 μM NNC 55-0396 added to the pipette solution. As shown in Fig. 7B, there was no significant difference between the current in HEK 293/α1G cells with (n = 10) and without (n = 6) NNC 55-0396 included in the pipette solution. For example, at the 10-min timepoint, the p value is 0.12 with two-tailed unpaired Student's t test analysis. These data are inconsistent with the idea that the site of action of NNC 55-0396 is on the cytoplasmic face of the T-type Ca2+ channel. Thus, NNC 55-0396 appears to exhibit a relatively slow and wash-resistant binding to a site, perhaps within the transmembrane domains of the channel.
Discussion
Mibefradil blocks both T-type Ca2+ and HVA Ca2+ channels. Although mibefradil appears to act directly on T-type channels, we previously reported that its effect on HVA channels is not direct but instead involves cell permeation and hydrolysis to an active metabolite that acts from the cytoplasmic side of the membrane (Wu et al., 2000). In the present study, we developed a new compound, NNC 55-0396, that has an inhibitory effect on T-type Ca2+ current in HEK/α1G cells but is not hydrolyzed to an active metabolite and does not block HVA Ca2+ currents in INS-1 cells. Thus, NNC 55-0396 is a selective inhibitor for T-type Ca2+ channels. Our data also suggest that the effect of NNC 55-0396 on T-type channels is state-dependent; i.e., block is partially relieved by membrane hyperpolarization and enhanced by high-frequency channel activation.
Our previous findings (Wu et al., 2000) suggest that the methoxyacetyl group of mibefradil is a critical determinant for binding to HVA channels since its hydrolysis is the critical step in the binding of the compound to HVA channels. In contrast, this moiety may play only a modest role in interacting with the receptor domain of T-type Ca2+ channels since all three alternations—NNC 55-0395, NNC 55-0396, and NNC 55-0397—retain the capability of blocking T-type Ca2+ current, although with varying potency. Our study indicates that modification in the methoxyacetyl group of mibefradil can improve its selectivity for T-type Ca2+ channels over the HVA Ca2+ channels without sacrificing the potency of T-type Ca2+ channel antagonism.
The primary objective of this study is to examine the structure-selectivity relationship of a small group of mibefradil derivatives. Therefore, we chose HEK/α1G as a clean model system. We do not know how effectively NNC 55-0396 will block other T-type Ca2+ channels, nor do we know its potency on α1G when other auxiliary subunits are present. However, we speculate that NNC 55-0396 will block α1H and α1I, for it has been shown that the IC50 values of mibefradil blocking α1G, α1H, and α1I are similar (Martin et al., 2000). In summary, our findings represent an important step toward development of a specific T-type Ca2+ inhibitor and thus have potentially important scientific and therapeutic implications.
Acknowledgments
We thank Dr. B. Z. Peterson for constructive criticism on this work. The mass spectrometric analyses were carried out in the Louisiana State University Health Sciences Center New Orleans Core Laboratories (New Orleans, LA).
Footnotes
-
Article, publication date, and citation information can be found at http://jpet.aspetjournals.org.
-
DOI: 10.1124/jpet.103.060814.
-
This study was supported by American Heart Association Grant 0151047B (to M.L.) and by Canadian Institutes of Health Research Grant MT13485 (to D.S.R.).
-
ABBREVIATIONS: HVA, high-voltage-activated; HEK, human embryonic kidney; NNC 55-0395, (1S,2S)-2-(2-(N-[(3-benzimidazol-2-yl)propyl]-N-methylamino)ethyl)-6-fluoro-1,2,3,4-tetrahydro-1-isopropyl-2-naphtyl valeroate dihydrochloride); NNC 55-0396, (1S,2S)-2-(2-(N-[(3-benzimidazol-2-yl)propyl]-N-methylamino)ethyl)-6-fluoro-1,2,3,4-tetrahydro-1-isopropyl-2-naphtyl cyclopropanecarboxylate dihydrochloride; NNC 55-0397, (1S,2S)-2-(2-(N-[(3-benzimidazol-2-yl)propyl]-N-methylamino)ethyl)-6-fluoro-1,2,3,4-tetrahydro-1-isopropyl-2-naphtyl isobutyrate dihydrochloride; PBS, phosphate-buffered saline; EI, electron impact; DMSO, dimethyl sulfoxide; G, conductance; MALDI-TOF, matrix-assisted laser desorption ionization/time of flight instrument.
- Received October 2, 2003.
- Accepted December 5, 2003.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}