


Abstract
The negative affective aspects of nicotine withdrawal have been hypothesized to contribute to tobacco dependence. In the present studies in rats, brain stimulation reward thresholds, conditioned place aversions, and somatic signs of withdrawal were used to investigate the role of central and peripheral nicotinic acetylcholine and opioid receptors in nicotine withdrawal. Rats prepared with s.c. osmotic mini-pumps delivering 9.0 mg/kg/day nicotine hydrogen tartrate or saline were administered various doses of the nicotinic antagonists mecamylamine (s.c.), chlorisondamine (s.c. or i.c.v.), dihydro-β-erythroidine (s.c.), or the opiate antagonist naloxone (s.c.). Nicotine-treated rats receiving mecamylamine or i.c.v. chlorisondamine exhibited elevated thresholds and more somatic signs than saline-treated rats. Nicotine-treated rats receiving s.c. chlorisondamine, at doses that do not readily cross the blood-brain barrier, exhibited more somatic signs than saline-treated rats with no threshold elevations. Naloxone administration produced threshold elevations and somatic signs only at high doses that induced similar magnitude effects in both nicotine- and saline-treated subjects. Mecamylamine or dihydro-β-erythroidine administration induced conditioned place aversions in nicotine-treated rats but required higher doses than those needed to precipitate threshold elevations. In contrast, naloxone administration induced conditioned place aversions at lower doses than those required to precipitate threshold elevations and somatic signs. These data provide evidence for a dissociation between centrally mediated elevations in reward thresholds and somatic signs that are both centrally and peripherally mediated. Furthermore, threshold elevations and somatic signs of withdrawal appear to be mediated by cholinergic neurotransmission, whereas conditioned place aversions appear to be primarily mediated by the opioid system.
Nicotine withdrawal in humans is characterized by somatic symptoms such as bradycardia, insomnia, gastrointestinal discomfort, and increased appetite, leading to weight gain (Hughes et al., 1991) as well as affective symptoms, including depressed mood, irritability, anxiety, frustration, difficulty concentrating, and craving for tobacco (American Psychiatric Association, 1994). Although the somatic symptoms of nicotine withdrawal may contribute to smoking behavior, it has been hypothesized that affective signs are of greater motivational significance in contributing to relapse and continued nicotine use than the somatic symptoms of nicotine withdrawal (Koob et al., 1993; Markou et al., 1998). Thus, animal models of the affective aspects of nicotine withdrawal are important tools for understanding the neurobiological bases of nicotine dependence and for developing effective treatment strategies to facilitate nicotine abstinence.
In the rat, withdrawal from chronic nicotine exposure results in the emergence of somatic signs, including abdominal constrictions, facial fasciculation, increased eyeblinks, and ptosis (Malin et al., 1992;Hildebrand et al., 1997; Epping-Jordan et al., 1998). This somatic syndrome also can be precipitated by the nicotinic acetylcholine receptor (nAChR) antagonists mecamylamine, hexamethonium, chlorisondamine, and dihydro-β-erythroidine (DHβE) (Malin et al., 1994, 1997, 1998; Hildebrand et al., 1997) in rats chronically exposed to nicotine. Interestingly, the somatic signs of nicotine withdrawal also have been precipitated in nicotine-treated rats by the opiate antagonist naloxone (Malin et al., 1993). Studies have sought to address the relative involvement of the central and peripheral nervous systems in mediating the somatic signs of nicotine withdrawal and have yielded evidence for contribution from both sources (Hildebrand et al., 1997; Malin et al., 1997, 1998). In nicotine-treated rats, central administration of the competitive nAChR antagonist DHβE and the noncompetitive nAChR antagonist hexamethonium precipitated somatic signs of nicotine abstinence, whereas systemic administration of hexamethonium, a nAChR antagonist that does not readily cross the blood-brain barrier (Gosling and Lu, 1969), did not precipitate a significant somatic syndrome (Malin et al., 1997, 1998). Although these results suggest that somatic signs of nicotine withdrawal are centrally mediated, a peripherally, but not centrally, active dose of the noncompetitive nAChR antagonist chlorisondamine precipitated somatic signs of nicotine withdrawal (Hildebrand et al., 1997). Hence, somatic symptoms of nicotine withdrawal are mediated by the inactivation of nAChRs through the termination of chronic nicotine or precipitated by centrally or peripherally active nAChR antagonists.
Nevertheless, overt somatic signs of nicotine withdrawal may not accurately reflect the affective or motivational state of the animal. Changes in reward function, motivation, and affect are difficult to assess in animals; however, brain stimulation reward thresholds have been shown to be a valid and reliable measure of the diminished reward and motivation associated with withdrawal from several drugs of abuse, including cocaine, amphetamine, opiates, and ethanol (Leith and Barrett, 1976; Markou and Koob, 1991; Schulteis et al., 1994, 1995; Lin et al., 1999). Elevations in brain stimulation reward thresholds also were observed during spontaneous withdrawal from chronic nicotine administration and DHβE-precipitated withdrawal (Epping-Jordan et al., 1998).
Clinical and preclinical studies indicate that negative affective states experienced during drug withdrawal can become associated with previously neutral environmental stimuli and that these conditioned stimuli gain motivational significance in the maintenance of drug use and relapse during periods of abstinence (Wikler, 1973; Childress et al., 1994; Schulteis et al., 1998). For example, precipitated opiate withdrawal in rats produced an aversive motivational state that became associated with environmental cues and led to a conditioned place aversion (Hand et al., 1988; Stinus et al., 1990; Schulteis et al., 1994, 1998; Spanagel et al., 1994). Consequently, precipitated nicotine withdrawal-induced conditioned place aversions also provide a sensitive measure of affective changes produced by nicotine abstinence.
The purpose of these studies was to delineate the role of the central and peripheral nervous systems in mediating the motivational aspects of precipitated nicotine withdrawal. The effects of systemically administered competitive opiate antagonist naloxone or the noncompetitive nAChR antagonist mecamylamine, which both cross the blood-brain barrier (Ngai et al., 1976; Martin et al., 1989, respectively), or systemically or centrally administered noncompetitive nAChR antagonist chlorisondamine, which does not readily cross the blood-brain barrier (Gosling and Lu, 1969), on brain reward function and somatic signs were measured in nicotine- and saline-treated rats. Additional studies examined whether various doses of the nAChR antagonists mecamylamine or DHβE, or the opiate receptor antagonist naloxone, also induce conditioned place aversions in nicotine- and saline-treated rats.
Materials and Methods
Subjects
For all experiments conducted at The Scripps Research Institute (La Jolla, CA; i.e., all experiments except conditioned place aversion), male Wistar rats bred at the Beckman Laboratories of The Scripps Research Institute from a Wistar stock originally obtained from Charles River, NY, were used. At Beckman Laboratories, rats were bred with a circular pair random system of breeding to maintain genetic heterogeneity. New breeders were obtained from Charles River as determined by our internal Genetics Advisory Board. For the place aversion experiments, conducted at the University of Bordeaux 2, Unité Mixte de Recherche Centre National de la Recherche Scientifique 5541 (Bordeaux, France), male Sprague-Dawley rats (IFFA CREDO, Lyon, France) were used. Animals were group-housed (two to four per cage) in temperature and humidity-controlled vivariums on a 12-h reverse light/dark cycle with ad libitum access to food and water. Place aversion testing occurred during the light phase of the light/dark cycle. All other behavioral testing occurred during the dark phase of the light/dark cycle. For 1 week after arrival, animals were allowed to habituate to their new environment and were handled twice at the end of this week. All subjects were treated in accordance with the National Institutes of Health guidelines regarding the principles of animal care. Animal facilities and experimental protocols were in accordance with the Association for the Assessment and Accreditation of Laboratory Animal Care.
Drugs
(−)-Nicotine hydrogen tartrate salt ([−]-1-methyl-2-[3-pyridyl] pyrrolidine), mecamylamine hydrochloride, and naloxone hydrochloride were purchased from Sigma Chemical Co., St. Louis, MO; dihydro-β-erythroidine hydrobromide was purchased from Research Biochemicals, Natick, MA; and chlorisondamine diiodide was purchased from Tocris, Ballwin, MO. All drugs were dissolved in physiological saline (0.9% sodium chloride). All s.c. injections were administered in a volume of 1 ml/kg b.wt. All intraventricular injections were administered in a volume of 5 μl over 30 s. Drug doses refer to the salt form.
Apparatus
Brain Stimulation.
Brain stimulation training and testing took place in eight Plexiglas operant chambers (25 × 31 × 24 cm) each housed in a sound-attenuating box (Med Associates, St. Albans, VT). The operant chamber consisted of floors constructed of parallel aluminum rods spaced 1.25 cm apart. One wall contained a metal wheel manipulandum that required 0.2 N force to rotate it one-quarter of a turn. The wheel (5 cm in width) extended out of the wall ∼3 cm. Each testing chamber was enclosed within a larger, sound- and visual-attenuated chamber (62 × 63 × 43 cm). Brain stimulation was delivered by constant current stimulators (Stimtech model 1200; San Diego Instruments, San Diego, CA). Subjects were connected to the stimulation circuit through flexible bipolar leads (Plastics One, Roanoke, VA) attached to gold-contact swivel commutators (model SL2C; Plastics One) mounted above the chamber. The stimulation parameters, data collection, and all test session functions were controlled by a microcomputer.
Conditioned Place Aversion.
The apparatus consisted of three rectangular boxes (40 × 33 × 34 cm) spaced at 120° angles and all accessible from a triangular central compartment. Distinctive visual and tactile cues distinguished the three compartments (Stinus et al., 1990). Each compartment was equipped with a series of infrared photocells to allow automatic detection of the rat's position at all times. The apparatus was located in a sound-attenuated testing room, with white noise (75 dB) to mask external noise and illuminated by three 15-W red lights located 1.5 m above each compartment.
Surgeries
Electrode and Cannula Implantation.
Rats were anesthetized with a Halothane/oxygen mixture (1–3% Halothane) then secured in a Kopf stereotaxic frame (David Kopf Instruments, Tujunga, CA) with the tooth bar elevated 5.0 mm above the intraaural line. A stainless steel bipolar electrode (model MS303/2; Plastics One) cut to 11 mm in length, with a diameter of 0.25 mm was implanted into the medial forebrain bundle at the level of the posterior lateral hypothalamus according to the following coordinates: A-P, −0.5 mm from bregma; M-L, ±1.7 mm; and D-V, −8.3 mm from dura with the incisor bar +5.0 mm above the interaural line (Pellegrino et al., 1979). For subjects in the i.c.v. experiments (n = 16), a stainless steel guide cannula (23-gauge, 7 mm in length) was lowered to 2 mm above the lateral ventricle according to the following coordinates: A-P, −0.6 mm from bregma; M-L, ±2.0 mm; and D-V, −3.2 mm from skull with the incisor bar +5.0 mm above the interaural line (Pellegrino et al., 1979). The electrode or electrode/cannula assembly was anchored by four stainless steel screws, embedded in dental acrylic (Teets methyl methacrylate denture material; CoOral-Lite Mfg. Co., Diamond Springs, CA). A dummy stylet, extending to the injection site was inserted to keep foreign objects out of the cannula and to prevent cannula blockage. The surgical wound was flushed with a solution of saline and Gentamicin (0.3 ml of 40 mg/ml Gentamicin sulfate in 0.6 ml of physiological saline), closed with silk sutures, followed by application of Bacitracin ointment. Animals were allowed to recover at least 7 days before any behavioral testing or drug administration.
Mini-Pump Surgery.
Rats were anesthetized with a Halothane/oxygen mixture (1–3% Halothane) and prepared with Alzet osmotic mini-pumps [model 2 ML2 (14-day); Alza Corporation, Palo Alto, CA] placed s.c. (back of the animal parallel to the spine). Pumps were filled with either physiological saline or nicotine tartrate solution. The concentration of nicotine tartrate salt solution was adjusted according to animal weight, resulting in ∼9 mg/kg/day (3.16 mg/kg, free base) for 14 days. The surgical wound was closed with 9-mm stainless steel wound clips (Becton Dickinson Primary Care Diagnostics, Sparks, MD) and treated with Bacitracin ointment.
Intracerebroventricular Drug Administration
An 8.5-mm injector (30-gauge, stainless steel) was connected to 70 cm of calibrated polyethylene-10 tubing preloaded with drug solution and lowered into the ventricle via the guide cannula. All drug injections were made by gravity induced by raising the tubing above the animal's head until flow began. Five microliters of solution was administered by gravity over 30 s and the injector was left in place an additional 30 s, after which the dummy stylet was replaced.
Histologies
At the completion of the i.c.v. chlorisondamine experiment, all cannula placements were verified by the injection of toluidine blue (Kodak Co., NY), followed by histological examination. Only subjects with proper cannula placements were included in the data analyses.
Brain Stimulation Reward Threshold Procedure
The procedure used was a modification of the Kornetsky and Esposito discrete-trial current-threshold procedure (Kornetsky and Esposito, 1979; Markou and Koob, 1993). At the beginning of each trial, rats received a noncontingent electrical stimulus after which the animals had 7.5 s to turn the wheel manipulandum one-quarter of a turn to receive a contingent stimulus identical in all parameters to the noncontingent stimulus, ending the trial. If a response did not occur within the 7.5 s, the trial also ended. The intertrial interval ranged from 7.5 to 12.5 s, averaging 10 s. Any responding during the intertrial interval resulted in a 12.5-s delay before the initiation of the next trial. After the intertrial interval, another trial began with the presentation of a noncontingent electrical stimulus. Stimulus intensities were presented in alternating descending and ascending series with a step size of 5 μA. Animals were offered three trials at each current intensity with the starting current of the first descending series set 30 to 40 μA above the subject's baseline threshold level estimated at the end of preliminary training. Subjects completed four series (two descending and two ascending) during each daily testing session which lasted ∼30 min. The current threshold was defined as the midpoint between the current intensity level at which the animal made two or more positive responses out of three stimulus presentations and the level where the animal made less than two positive responses. The animal's estimated current threshold for the session was defined as the mean of the four individual series thresholds. In addition to the threshold measure, a measure of performance, response latency, was obtained from the discrete-trial current-threshold paradigm. Response latency was defined as the time between the delivery of the noncontingent stimulus (i.e., initiation of the stimulus) and the animal's response on the wheel manipulandum. Mean response latency was defined as the mean latency of responding during all trials when the animal responded within the 7.5-s interval after the noncontingent stimulus.
Ratings of Somatic Signs of Withdrawal
Each rat was placed in a plastic opaque cylindrical container (30 × 29 cm) in which the rat could move around freely. Each subject was observed for 10 min during which time the subject was filmed with a video recording device. The frequency and time of occurrence of the following signs were recorded: body shakes, chews, cheek tremors, escape attempts, eyeblinks, foot licks, gasps, genital licks, head shakes, ptosis, scratches, teeth chattering, writhes, and yawns on a checklist of nicotine abstinence signs, derived from standard checklists of opiate withdrawal signs (Malin et al., 1992). The categories of “abdominal constrictions” included gasps and writhes; “facial fasciculation” included cheek tremors, chews, and teeth chattering; and “miscellaneous other signs” included shakes, escape attempts, licks, scratches, and yawns. Multiple successive counts of any sign required a distinct pause between episodes. Ptosis, if present continuously, was only counted once per minute. The total number of somatic signs per 10-min observation period was defined as the sum of individual occurrences of the above-mentioned withdrawal signs. Subjects were habituated to the observation room and containers for 10 min over 3 days before the first antagonist injection.
Experimental Procedure: Brain Stimulation and Somatic Signs of Withdrawal
Naive rats were used for each experiment. Each experiment investigated the effects of various doses of an antagonist on reward thresholds and/or somatic signs with a within-subject design for antagonist dose and between-subject design for treatment (seeStatistical Analyses and Table1). After recovery from surgery, rats were trained to respond in the self-stimulation paradigm described above. Rats were tested until stable thresholds were achieved, defined as <10% variation within a 5-day period. Once stable thresholds had been achieved, rats were prepared with 14-day osmotic mini-pumps filled with either saline or nicotine hydrogen tartrate dissolved in saline. On day 6 of chronic nicotine or saline exposure, rats received the first injection of mecamylamine (0.29–1.14 mg/kg s.c.,n = 9 nicotine, n = 8 saline), chlorisondamine (0.1–1.0 mg/kg s.c., n = 8 nicotine,n = 9 saline or 2.5–10.0 μg i.c.v.,n = 9 nicotine, n = 7 saline), or naloxone (0.03–0.48 mg/kg s.c., n = 8 nicotine,n = 8 saline) dissolved in 0.9% physiological saline, 15 min before threshold testing, with a within-subjects repeated-measures Latin-square design that included a saline injection. Rats were required to return to baseline threshold levels before subsequent injections. Mecamylamine and naloxone were administered with 48 h between injections. Based on the results of pilot studies, chlorisondamine (s.c. and i.c.v.) was administered with 72 h between injections. On day 13 (mecamylamine and naloxone experiments) or day 14 (chlorisondamine experiments) of chronic nicotine or saline exposure, after a baseline threshold session, rats were prepared with a second 14-day mini-pump (Table 1). Nicotine-treated rats were prepared with the second mini-pump on the opposite side of the spine to prevent tissue damage from the nicotine solution, whereas the second saline pumps were placed in the same location as the original saline mini-pumps. Although the saline pumps were implanted in the same site, it was not expected that this procedure would contribute to differences between nicotine- and saline-treated rats because saline does not produce any effects. After one baseline threshold session, rats received the first injection in a second Latin-square design for mecamylamine (1.72–3.43 mg/kg s.c.) or naloxone (1.0–4.0 mg/kg s.c.), each including an additional saline injection as part of the Latin-square design. In the chlorisondamine experiments, after preparation with a second 14-day mini-pump and a baseline threshold session, rats received the final dose in the first Latin-square design. At the end of the Latin-square in the s.c. chlorisondamine experiment, all subjects received two additional injections of chlorisondamine (0.2 and 0.3 mg/kg s.c.) with 72 h between injections. At the end of the Latin-square in the i.c.v. chlorisondamine experiment, all rats were administered a single dose of chlorisondamine (1.0 μg i.c.v.).
Experimental design for nicotine or saline pump implantation and subsequent administration of mecamylamine or naloxone (total of eight injections each, including two of saline), or chlorisondamine (s.c., total of six injections, including one of saline; i.c.v., total of five injections, including one of saline)
The doses of mecamylamine selected were of equimolar concentration to those of DHβE previously reported to precipitate elevations in brain stimulation reward thresholds in nicotine-treated rats (Epping-Jordan et al., 1998). Chlorisondamine doses were selected based on results from previous behavioral studies with this compound (Clarke, 1984;El-Bizri et al., 1995; Reuben et al., 1998). Naloxone doses were selected based on previous reports of naloxone-precipitated elevations in brain stimulation reward thresholds in opiate-dependent rats (Schulteis et al., 1994), and naloxone-precipitated somatic signs of nicotine withdrawal (Malin et al., 1993).
Immediately after each threshold testing session, ∼45 min after the antagonist or saline injection, each rat was observed for somatic signs of nicotine withdrawal for 10 min (see above). In a second “somatic-only” naloxone experiment, rats (n = 8 nicotine, n = 8 saline) were prepared with osmotic mini-pumps delivering either saline or nicotine hydrogen tartrate dissolved in saline. On day 6 of chronic nicotine or saline exposure, rats received the first injection of a Latin-square design of naloxone (1.0–4.0 mg/kg s.c.), or saline as specified by the Latin-square design, 15 min before somatic observations. Doses of naloxone were administered with 48 h between injections. On day 13 of chronic nicotine or saline exposure, rats were prepared with a second mini-pump, and 48 h after mini-pump replacement, all rats received a single injection of naloxone (8.0 mg/kg s.c.) and were observed for somatic withdrawal signs. The purpose of this study was to assess the ability of naloxone to precipitate somatic withdrawal signs shortly after naloxone administration (15 min, comparable with the pretreatment time used by Malin et al., 1993) and shorter than the 45-min pretreatment time used in the previous naloxone study reported herein.
Place Aversion Conditioning Procedure
The place conditioning experiments consisted of three phases: preconditioning, conditioning, and testing. In the preconditioning phase (3 days after implantation of mini-pumps), animals were placed in the center triangular compartment and allowed to freely explore the apparatus for 20 min. The two compartments in which the most similar time was spent were randomly paired with an antagonist dose [DHβE (0.25–10 mg/kg s.c.), mecamylamine (0.5–6.0 mg/kg s.c.), naloxone (30–240 μg/kg s.c.)], or saline. The third compartment was not paired with an injection. This selection and pairing procedure allowed for a minimization of the imbalance in time spent in the antagonist- versus saline-paired compartments. In the conditioning phase, rats received injections of saline on days 4, 6, and 8 postpump implantation immediately before being confined to the preselected saline-paired compartment for 20 min. On days 5, 7, and 9 postpump implantation, the rats received one of several doses of DHβE, mecamylamine, or naloxone immediately before confinement in the antagonist-paired compartment for 20 min (three pairings of antagonist with compartment). The testing phase consisted of a 20-min free exploration of the entire apparatus on day 10 postpump implantation, 24 h after the last drug dosing. The difference in time spent in the antagonist-paired compartment during the testing phase and preconditioning phase served as a measure of place aversion. A between-subjects design was used for the factor of antagonist dose.
Statistical Analyses
All reward threshold and somatic withdrawal sign data were analyzed with mixed-factor ANOVA. Two-way repeated-measures ANOVAs were used to analyze threshold data, response latency, overall somatic signs, and the individual somatic withdrawal signs (e.g., abdominal constrictions, facial fasciculation, eyeblinks, ptosis, and miscellaneous other signs) with two levels of the between-subjects factor of group (nicotine or saline) and antagonist dose as the within-subjects factor. Chlorisondamine (s.c. and i.c.v.) data were analyzed with 3-way ANOVAs with the additional within-subjects factor of time postinjection, corresponding to individual time points of testing and observation. To test for possible confounding effects of the order of drug administration, the repeated-measures ANOVAs were recalculated with the order of the treatments as the independent variable. Prepump implantation baseline thresholds and in-between injections baseline thresholds were subjected to one-factor repeated-measures ANOVA to assess the stability of baseline responding and potential carryover effects throughout the course of drug treatment regimens for all experiments. After significance in the overall ANOVA, post hoc comparisons among means were conducted with Tukey's honestly significant difference post hoc test after a significant effect of the between factor group or the Newman-Keuls post hoc test after a significant interaction effect or an effect of antagonist dose. Nonparametric statistics were used to analyze the place aversion conditioning data because these data were not normally distributed, and thus, violated one of the assumptions required for parametric statistics. Place aversion conditioning data were analyzed with the nonparametric Wilcoxon signed ranks test comparing the time spent in the receptor antagonist-paired compartment before conditioning versus after conditioning; the subgroup variables were the pretreatments, either mecamylamine, DHβE, or naloxone. The level of statistical significance was set at P < .05.
Results
Order and Carryover Effects.
No effect of dose order was noted for the reward threshold measure (Fs = 0.54–1.04, NS), the latency measure (Fs = 0.42–1.76, NS), or somatic signs (Fs = 0.65–0.98, NS) in any of the experiments. There were no differences in the mean baseline thresholds between nicotine- and saline-treated animals during acute nicotine or saline exposure (5 days) and no significant carryover effects of any antagonist administered in any of the experiments during in-between drug baseline days (Fs = 0.15–0.95, NS). The mean baseline threshold values for the subjects before any manipulations were 143.23 ± 10.10 μA (mecamylamine), 144.14 ± 11.88 μA (chlorisondamine s.c.), 147.89 ± 9.43 μA (chlorisondamine i.c.v.), and 159.94 ± 11.97 μA (naloxone).
Mecamylamine and DHβE.
Subcutaneous administration of the nicotinic receptor antagonist mecamylamine produced a dose-dependent elevation in brain stimulation reward thresholds in nicotine-treated but not in saline-treated rats. This effect was reflected in a statistically significant effect of group (F1,15 = 12.20, P < .01), a significant effect of dose (F6,90 = 8.65, P < .01), and a significant group × dose interaction (F6,90 = 6.94, P < .01). Tukey post hoc tests indicated that nicotine-treated animals had significantly elevated reward thresholds compared with saline-treated animals at all doses above 0.29 mg/kg mecamylamine. Newman-Keuls post hoc tests revealed statistically significant elevations in thresholds after 1.14 to 3.43 mg/kg mecamylamine compared with those after injection of 0.57 mg/kg, P < .05, indicating a dose-dependent effect of mecamylamine (Fig.1A).
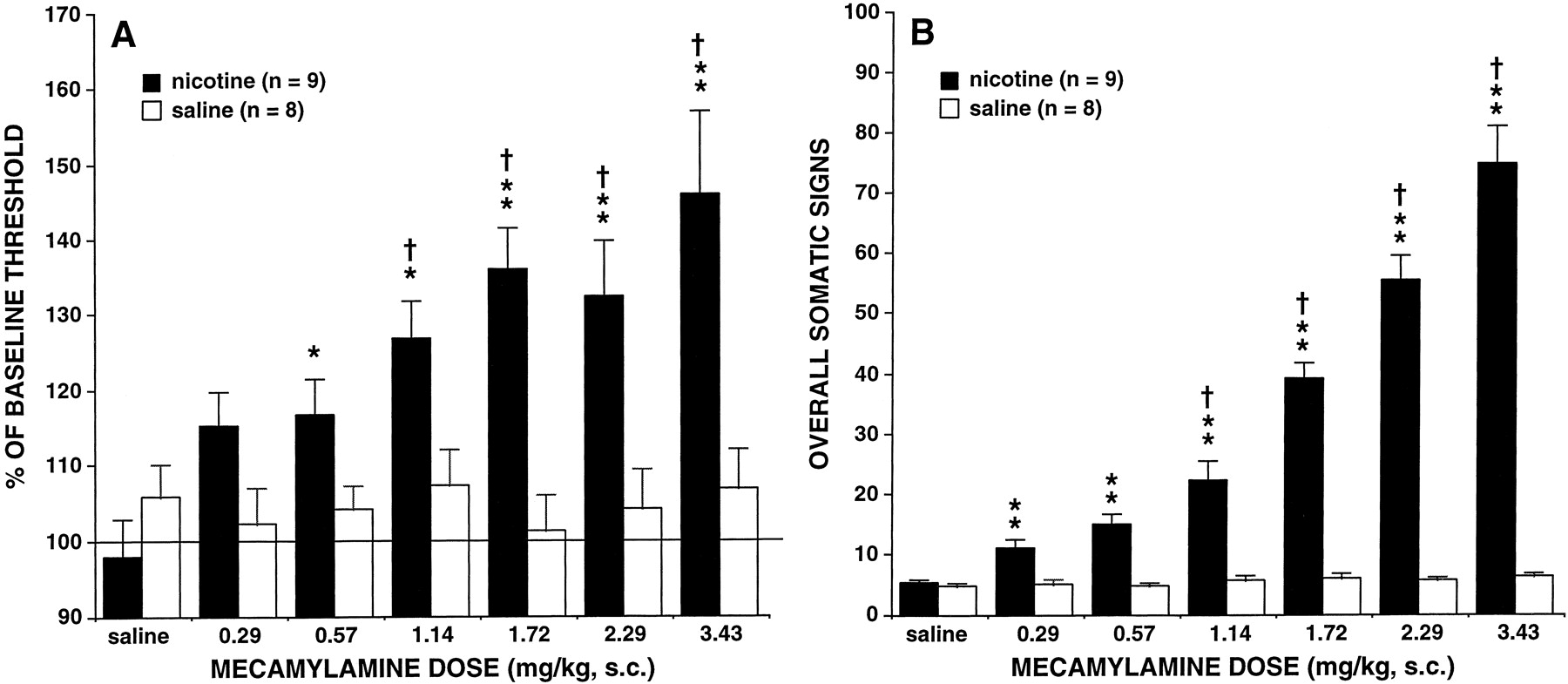
A, effects of s.c. mecamylamine on brain stimulation reward thresholds (mean ± S.E.) in nicotine (n = 9) and saline-treated (n = 8) rats. Data (mean ± S.E.) are expressed as a percentage of the mean 5-day baseline threshold. B, overall somatic withdrawal signs (mean ± S.E.) during a 10-min observation precipitated by mecamylamine during chronic nicotine (9.0 mg/kg/day, salt form,n = 9) or saline administration (n = 8). Asterisks indicate statistically significant differences between nicotine- and saline-treated rats,*P < .05,**P < .01. †, indicates statistically significant differences compared with administration of 0.57 mg/kg, indicating a dose-dependent effect of mecamylamine,†P < .05.
As shown in Fig. 1B, s.c. administration of mecamylamine produced increases in the overall number of somatic withdrawal signs reflected in a statistically significant effect of group (F1,15 = 236.93, P < .01), a significant effect of dose (F6,90 = 65.02, P < .01), and a significant group × dose interaction (F6,90 = 59.59, P < .01). Tukey post hoc tests indicated that nicotine-treated animals displayed significantly more somatic signs of nicotine withdrawal than saline-treated animals after all doses of mecamylamine. Newman-Keuls post hoc tests revealed significant increases in somatic signs after 1.14 to 3.43 mg/kg mecamylamine compared with those after injection of 0.57 mg/kg, P < .05, indicating a dose-dependent effect of mecamylamine. Compared with saline-treated animals, nicotine-treated animals showed statistically significant increases in the following categories of abstinence signs: abdominal constrictions, facial fasciculation, eyeblinks, ptosis, and miscellaneous other signs that included escape attempts, foot licks, genital grooming, shakes, scratches, and yawns (Fig. 2, A and B).

Individual categories of somatic withdrawal signs (mean ± S.E.) during a 10-min observation ∼45 min after an s.c. injection of mecamylamine in nicotine-treated (n = 9) (A) or saline-treated (n = 8) rats(B), or ∼45 min after an s.c. injection of chlorisondamine in nicotine-treated (n = 8) (C) or saline-treated (n = 9) rats (D), or ∼45 min after an i.c.v. injection of chlorisondamine in nicotine-treated (n = 9) (E) or saline-treated (n = 7) rats (F). Miscellaneous signs included escape attempts, foot licks, genital grooming, shakes, scratches, and yawns. Asterisks indicate statistically significant differences between nicotine- and saline-treated rats, *P < .05,**P < .01.
As shown in Fig. 3A, mecamylamine induced a statistically significant place aversion in nicotine-treated rats after administration of 4 mg/kg s.c. mecamylamine (Wilcoxon T = 2.98, P < .01) and in both nicotine- and saline-treated rats after administration of 6 mg/kg, s.c. mecamylamine (Wilcoxon T = 2.84, P < .01; T = 2.04,P < .05, respectively). As shown in Fig. 3B, DHβE induced a statistically significant place aversion in nicotine-treated rats after administration of 10 mg/kg s.c. DHβE (Wilcoxon T = 2.82, P < .01).

A, conditioned place aversion induced by s.c. administration of mecamylamine (n = 12–20 nicotine, n = 9–12 saline). B,conditioned place aversion induced by s.c. administration of DHβE (n = 6–12 nicotine, n = 6 saline). C, conditioned place aversion induced by s.c. administration of naloxone (n = 7–12 nicotine,n = 8–14 saline). Values represent the mean ± S.E. difference in time spent in the antagonist-paired compartment before conditioning versus after conditioning. Asterisks indicate statistically significant differences between the time spent in the antagonist-paired compartment before conditioning compared with after conditioning, *P < .05,**P < .01.
Chlorisondamine (s.c.).
Subcutaneous administration of the nicotinic receptor antagonist chlorisondamine did not precipitate elevations in brain stimulation reward thresholds (Fs = 0.46–1.58, NS; Fig. 4A). In contrast, as shown in Fig. 4B, s.c. administration of chlorisondamine produced increases in the overall number of somatic withdrawal signs in nicotine-treated rats reflected in a statistically significant effect of group (F1,15 = 13.84,P < .01), a significant effect of dose (F5,75 = 47.17, P < .01), a significant effect of time (F2,30 = 121.85, P < .01), and a significant group × dose × time interaction (F10,150 = 44.04, P < .01). Newman-Keuls post hoc tests indicated that nicotine-treated animals displayed significantly more somatic signs of nicotine withdrawal than saline-treated animals 15 min after the administration of 0.2 to 1.0 mg/kg chlorisondamine administration, P< .05. Compared with saline-treated animals, nicotine-treated animals showed statistically significant increases in the following categories of abstinence signs: abdominal constrictions, facial fasciculation, and ptosis (P < .05) (Fig. 2, C and D).

A, time course effects of s.c. chlorisondamine on brain stimulation reward thresholds (mean ± S.E.) in nicotine- (n = 8) and saline-treated (n = 9) rats. Data are expressed as a percentage of the mean 5-day baseline reward threshold. B, time course effects of overall somatic withdrawal signs (mean ± S.E.) precipitated by s.c. chlorisondamine during chronic nicotine (9.0 mg/kg/day, salt form, n = 8) or saline administration (n = 9). Each data point refers to a 10-min observation. Asterisks indicate statistically significant differences between nicotine- and saline-treated rats,*P < .05,**P < .01. Time points of testing: 15 m = 15 min, 24 h = 24 h, 48 h = 48 h.
Chlorisondamine (i.c.v.).
Intracerebroventricular administration of chlorisondamine produced an increase in brain stimulation reward thresholds in nicotine- and saline-treated rats reflected in a statistically significant effect of dose (F4,56 = 10.96, P < .01), a significant effect of time (F3,42 = 15.12, P < .01), a significant group × time interaction (F3,42 = 3.55, P < .05), and a significant dose × time interaction (F12,168 = 3.60, P < .01). Comparisons performed with the Newman-Keuls test indicated that nicotine-treated animals had significantly elevated reward thresholds compared with saline-treated animals 15 min after administration of 5.0 μg and 3 h after administration of 2.5 μg i.c.v. chlorisondamine, P < .05. Compared with i.c.v. saline administration at the same time point of testing, nicotine-treated animals had significantly elevated thresholds at the following time points postinjection: 15 min and 3 h after 2.5 μg (P < .05), 15 min and 3 h after 5.0 μg (P < .05), and 15 min and 3 h after 10.0 μg (P < .01 and P < .05, respectively). Saline-treated animals had significantly elevated thresholds compared with saline administration at the following time points postinjection: 24 h after 1.0 μg (P < .05) and 15 min and 3 h after 10.0 μg (P < .05 andP < .01, respectively) (Fig.5A).

A, time course effects of i.c.v. administration of chlorisondamine on brain stimulation reward thresholds (mean ± S.E.) in nicotine (n = 9) and saline-treated (n = 7) rats. Data are expressed as a percentage of the mean 5-day baseline reward threshold.B, time course effects of i.c.v. administration of chlorisondamine on overall somatic withdrawal signs (mean ± S.E.) during chronic nicotine (9.0 mg/kg/day, salt form,n = 9) or saline administration (n = 7). Each data point refers to a 10-min observation. Asterisks indicate statistically significant differences between nicotine- and saline-treated rats,*P < .05,**P < .01. † indicates statistically significant differences compared with saline administration at the same time point,†P < .05,††P < .01. Time points of testing: 15 m = 15 min, 3 h = 3 h, 24 h = 24 h, 48 h = 48 h.
As shown in Fig. 5B, i.c.v. administration of chlorisondamine produced increases in the overall number of somatic withdrawal signs with a statistically significant effect of group (F1,14 = 15.85, P < .01), a significant effect of dose (F4,56 = 26.33, P < .01), a significant effect of time (F3,42 = 53.77, P < .01), and a significant group × dose × time interaction (F12,168 = 4.61, P < .01). Newman-Keuls post hoc tests indicated that nicotine-treated animals displayed significantly more somatic signs of nicotine withdrawal than saline-treated animals at the following time points postinjection: 3 h and 24 h after 1.0 μg (P< .05), 15 min and 3 h after 2.5 μg (P < .01 and P < .05, respectively), 15 min after 5.0 μg (P < .01), and 15 min after 10.0 μg (P < .01). Compared with saline-treated animals, nicotine-treated animals showed statistically significant increases in the following categories of abstinence signs: abdominal constrictions and facial fasciculation (Fig. 2, E and F). Histological examination of the brains after dye injections revealed placements of cannulas in the lateral ventricles for all subjects.
Naloxone.
Subcutaneous administration of the opiate receptor antagonist naloxone precipitated elevations in brain stimulation reward thresholds in nicotine- and saline-treated rats reflected in a statistically significant effect of dose (F3,42 = 4.63, P < .01). Newman-Keuls post hoc tests indicated that 2.0 and 4.0 mg/kg naloxone increased reward thresholds compared with saline administration in both nicotine- and saline-treated rats (Fig.6A).

A, effects of s.c. naloxone on brain stimulation reward thresholds (mean ± S.E.) in nicotine (n = 8) or saline-treated (n = 8) rats. Data are expressed as a percentage of the mean 5-day baseline threshold. B, overall somatic withdrawal signs (mean ± S.E.) during a 10-min observation period precipitated by s.c. injection of naloxone during chronic nicotine (9.0 mg/kg/day, salt form,n = 8) or saline administration (n = 8), ∼45 min postnaloxone injection. Inset shows overall somatic withdrawal signs precipitated 15 min after naloxone administration (n = 8 nicotine-treated rats, n = 8 saline-treated rats). #, indicates statistically significant increases for both nicotine and saline groups combined (P < .05) compared with saline administration (0 mg/kg).
After brain stimulation testing ∼45 min after injection, naloxone (0.03–4.0 mg/kg s.c.) did not precipitate somatic signs of nicotine withdrawal (Fs = 0–1.62, NS). In the somatic-only experiment, naloxone produced increases in somatic withdrawal signs in nicotine- and saline-treated rats with a statistically significant effect of dose (F4,56 = 36.97,P < .01). Newman-Keuls post hoc tests indicated that 8.0 mg/kg naloxone produced increases in the overall number of somatic withdrawal signs in both nicotine- and saline-treated animals compared with saline administration (Fig. 6B).
As shown in Fig. 3C, naloxone induced a statistically significant place aversion in nicotine-treated rats after administration of 0.12 mg/kg s.c. naloxone (Wilcoxon T = 2.13, P < .05) and in both nicotine- and saline-treated rats after administration of 0.24 mg/kg s.c. naloxone (Wilcoxon T = 2.27, P < .05; T = 2.42, P < .05, respectively).
Discussion
The results indicate that antagonist-precipitated nicotine withdrawal was associated with elevations in brain stimulation thresholds that reflect diminished sensitivity to rewarding stimuli, conditioned place aversions that reflect a conditioned negative affect to a distinct environment previously associated with nicotine withdrawal, and somatic signs. Furthermore, the data demonstrate that the decreases in reward observed during nicotine withdrawal are centrally mediated, whereas the somatic withdrawal syndrome appears to be both centrally and peripherally mediated. Finally, conditioned place aversions appear to reflect mostly decreased opioid neurotransmission, whereas threshold elevations and somatic signs appear to reflect primarily decreased cholinergic neurotransmission.
Systemic administration of the noncompetitive nonselective nAChR antagonist mecamylamine (Taylor, 1980) dose-dependently produced threshold elevations and increased somatic signs exclusively in nicotine-treated rats. This effect of mecamylamine on thresholds was similar in magnitude to that observed during spontaneous or DHβE-precipitated nicotine withdrawal, whereas the overall number of somatic signs precipitated by mecamylamine exceeded the numbers observed during spontaneous or DHβE-precipitated withdrawal (Malin et al., 1994, 1998; Epping-Jordan et al., 1998). In contrast to DHβE, which has high affinity and selectivity for the neuronal α4-receptor subunit that is found primarily in the brain (Harvey et al., 1996), mecamylamine is both centrally and peripherally active (Taylor, 1980). This may account for the more dramatic somatic syndrome precipitated by mecamylamine than DHβE (Tables 2, 3, and4). These results demonstrate that changes in nAChR function, at least partially, mediate the diminished reward and somatic signs of nicotine withdrawal. Furthermore, the results suggest that combined blockade of central and peripheral nAChRs with mecamylamine contributes to a more severe somatic syndrome than blockade of primarily central nAChRs with DHβE, which induced somatic signs only at doses that induced somatic signs in saline-treated rats also (Epping-Jordan et al., 1998).
Maximal antagonist effect in nicotine-treated rats
Rank order of effects in nicotine-treated rats
Maximal antagonist effect in saline-treated rats
In contrast to the relatively low doses of mecamylamine (0.57 mg/kg) or DHβE (2 mg/kg) (Epping-Jordan et al., 1998) required to induce threshold elevations, significantly higher doses of the same nAChR antagonists (4 mg/kg mecamylamine or 10 mg/kg DHβE) were required to induce conditioned place aversions in nicotine-treated rats (Table 2). Also in nicotine-treated rats, 0.57 mg/kg mecamylamine was sufficient to induce somatic signs, whereas DHβE induced somatic signs only at doses that also induced somatic signs in saline-treated subjects (≥4 mg/kg) (Epping-Jordan et al., 1998). Thus, reward thresholds appear to be a more sensitive measure of nAChR antagonist-precipitated nicotine withdrawal than conditioned place aversions or somatic signs.
As discussed above, blockade of primarily central nAChRs with DHβE was sufficient to produce threshold elevations in nicotine-treated rats without induction of somatic signs. Furthermore, systemic administration of the noncompetitive nAChR antagonist chlorisondamine at doses that do not readily penetrate the blood-brain barrier (Clarke, 1984; Clarke et al., 1994) did not elevate thresholds but increased somatic signs at all doses >0.2 mg/kg. There was a trend toward threshold elevations in nicotine-treated rats at the two highest doses tested. Nevertheless, this elevation of <10% is within the variability of the measure under baseline conditions. These findings indicate a dissociation between the threshold elevations and the somatic signs that are both associated with nicotine withdrawal (Tables 2, 3, and 4). Thus, the threshold elevations induced by mecamylamine in nicotine-treated rats or observed during spontaneous nicotine withdrawal are not likely to be a secondary effect of the induction of somatic signs.
In contrast to systemic administration of chlorisondamine, central administration of chlorisondamine at doses previously reported to block the acute effects of nicotine on locomotor activity, antinociception, and nicotine-evoked release of noradrenaline (Clarke, 1984; Bannon et al., 1998; Curzon et al., 1998; Reuben et al., 1998) led to both threshold elevations and increases in somatic signs in nicotine-treated rats. At the highest dose (10.0 μg i.c.v.), saline-treated rats also showed threshold elevations, similar to the threshold elevations observed after high doses of systemically administered DHβE in both saline- and nicotine-treated rats (Epping-Jordan et al., 1998). Collectively, these results suggest that tonic activity at nAChRs modulates brain stimulation reward, and reduced cholinergic neurotransmission decreases brain stimulation reward. A possible site for this modulation are the excitatory nAChRs located on ventral tegmental area dopaminergic neurons, as suggested by data showing that brain stimulation reward depends on activation of pedunculopontine cholinergic neurons that terminate on dopaminergic neurons in the ventral tegmental area (Yeomans et al., 1993).
It has been reported previously that the effects of centrally administered chlorisondamine are extremely long-lasting (up to 5 weeks) (Clarke, 1984; El-Bizri et al., 1995; Reuben et al., 1998). The current findings do not support a long-lasting receptor blockade by chlorisondamine because animals returned to baseline threshold levels within 48 h of injection. One explanation for this difference is the nicotine administration treatment regimen. In contrast to previous experiments that examined the effects of chlorisondamine on a single acute nicotine injection, the present study examined the effects of chlorisondamine in rats exposed to chronic continuous nicotine infusion. Such chronic nicotine exposure leads to receptor desensitization followed by an up-regulation in nAChRs (Wonnacott, 1990; Marks et al., 1992). It has been hypothesized that the balance of receptor activation, desensitization, and up-regulation serves to maintain baseline function within the systems affected by nicotine (Dani and Heinemann, 1996). Thus, in the presence of chronic nicotine there may be development of compensatory mechanisms that facilitate the reinstatement of baseline neuronal functioning after antagonist administration.
Even though the effects of chlorisondamine in the present studies were not as long-lasting as those reported previously, there were differential time courses of the effects seen after various doses of centrally administered chlorisondamine. More specifically, administration of 5.0 μg produced more rapid threshold elevations than administration of 2.5 μg of chlorisondamine (15 min and 3 h, respectively) in nicotine-treated rats. This difference may be due to the pharmacokinetics of chlorisondamine; the bisquaternary structure of chlorisondamine makes it difficult to diffuse through lipid layers. Thus, low chlorisondamine concentrations may require additional time for a sufficient quantity to reach the site of the nAChRs involved in reward processes, whereas higher concentrations may readily diffuse through brain tissue. In contrast to thresholds that were elevated 3 h after the administration of 2.5 or 5.0 μg i.c.v. chlorisondamine, somatic signs returned to baseline levels 3 h after administration of all chlorisondamine doses. In the presence of chronic nicotine, the systems that mediate the i.c.v. chlorisondamine-precipitated somatic signs may return to baseline activity levels faster than those mediating the reward signs of nicotine withdrawal. The observation that decreased brain reward function persists beyond the disappearance of somatic signs supports the hypothesis that reward and motivational changes during later withdrawal time points are more important contributors to relapse to tobacco smoking in humans than somatic signs.
Evidence suggesting that blockade of peripheral nAChRs contributes more to the mediation of the somatic signs than blockade of central nAChRs is provided by the finding that there were fewer overall somatic signs of withdrawal precipitated by i.c.v. than s.c. administration of chlorisondamine (Table 2). Nevertheless, there was no differential induction of the various types of somatic signs (Fig. 2). The current results, together with the finding that central administration of mecamylamine or hexamethenium also produced somatic signs of nicotine withdrawal (Malin et al., 1997; Hildebrand et al., 1999), indicate that the somatic syndrome associated with nicotine withdrawal is both centrally and peripherally mediated.
Similar to the cholinergic system, blockade of opioid receptors was sufficient to produce threshold elevations under baseline conditions. Systemic administration of 2 or 4 mg/kg of the opiate antagonist naloxone precipitated threshold elevations in both nicotine- and saline-treated rats with no differences between the two groups. Increased somatic signs were observed in both nicotine- and saline-treated rats only after receiving 8.0-mg/kg naloxone. The nondifferential effect of naloxone on reward thresholds in nicotine- and saline-treated rats, taken together with the finding that naloxone failed to block the threshold-lowering effect of nicotine (Huston-Lyons and Kornetsky, 1992), suggests that nicotine reward, measured by brain stimulation, is mediated by nonopiate systems. In contrast to thresholds and somatic signs, administration of a low naloxone dose (0.12 mg/kg) induced conditioned place aversions in nicotine-treated rats, whereas 0.24-mg/kg naloxone induced place aversions in both nicotine- and saline-treated rats. Thus, although the opiate system may be involved in the affective aspects of nicotine dependence as indicated by low dose naloxone-induced place aversions in nicotine-treated rats, the present data do not support an involvement of the opioid system in all aspects of nicotine withdrawal, such as those reflected in threshold elevations and somatic signs.
In summary, the present studies provide clear evidence for a dissociation between elevations in brain stimulation reward thresholds that are centrally mediated, and the somatic withdrawal syndrome that appears to be both centrally and peripherally mediated. This dissociation is further indicated by the observation that threshold elevations can occur without the appearance of somatic signs and vice versa. Finally, threshold elevations and somatic signs of withdrawal appear to be mediated by reduced cholinergic neurotransmission, whereas conditioned negative affect as measured by conditioned place aversions appears to be primarily mediated by reduced opioid neurotransmission.
Acknowledgments
This is publication 12541-NP from The Scripps Research Institute. We thank Drs. Amanda Roberts, John Walker, and Amanda Harrison for their consulting on the i.c.v. studies; Robert Lintz, Yves Liem, and Heather Jones for their excellent technical assistance; and Mike Arends for his meticulous editorial assistance.
Footnotes
-
Send reprint requests to: Athina Markou, Ph.D., Department of Neuropharmacology, The Scripps Research Institute, 10550 N. Torrey Pines Rd., CVN-7, La Jolla, CA 92037. E-mail: amarkou{at}scripps.edu
-
↵1 Shelly S. Watkins was supported by a predoctoral National Institute on Drug Abuse Individual National Research Service Award (DA05898). This work also was supported by Tobacco-Related Disease Research Program Grant 7RT-0004 from the State of California (to A.M.), a Novartis Research grant (to A.M.), National Institute on Drug Abuse Grant DA04398 (to G.F.K.), and by the University of Bordeaux 2, the Centre National de la Recherche Scientifique and the Conseil Regional d'Aquitaine (to L.S.).
-
↵2 Portions of these data were presented at the 28th Annual Society for Neuroscience meeting, Los Angeles, CA, November 1998, and at the 4th and 5th Annual Society for Research on Nicotine and Tobacco meetings, New Orleans, LA, March 1998 and San Diego, CA, March 1999, respectively.
- Abbreviations:
- nAChR
- nicotinic acetylcholine receptor
- DHβE
- dihydro-β-erythroidine
- Received August 23, 1999.
- Accepted November 16, 1999.
- The American Society for Pharmacology and Experimental Therapeutics





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}