Article Text

Abstract
Background Conditions characterised by chronic visceral pain represent a significant healthcare burden with limited treatment options. While animal models have provided insights into potential mechanisms of visceral nociception and identified candidate drug targets, these have not translated into successful treatments in humans.
Objective To develop an in vitro afferent nerve preparation using surgically excised freshly isolated human colon and vermiform appendix-mesentery tissues.
Methods Non-inflamed appendix (n=18) and colon (n=9) were collected from patients undergoing right and left hemicolectomy. Electrophysiological recordings were made from mesenteric nerves and the tissue stimulated chemically and mechanically.
Results Ongoing neuronal activity was sparse and where units occurred peak firing rates were: colon (2.0±0.4 spikes/s, n=4) and appendix (2.4±0.6 spikes/s, n=9). Afferent nerves innervating the appendix responded with a significant increase in activity following stimulation with inflammatory mediators (73±10.6 vs 3.0±0.3 spikes/s, n=6, p<0.001, inflammatory mediator vs baseline) and capsaicin (63±15.8 vs 2±0.3 spikes/s, n=3, p<0.001, capsaicin vs buffer). Afferent nerves innervating the colon responded with increased activity to blunt probing of the serosal surface.
Conclusions This first-in-human study demonstrates afferent nerve recordings from human gut tissue ex vivo and shows that tissue may be stimulated both chemically and mechanically to study neuronal responses. Collectively, the results provide preliminary evidence to validate this in vitro human tissue model as one that may aid future disease mechanistic studies and candidate drug testing.
- Visceral pain
- human
- afferent
- electrophysiology
- electrophysiology
- nerve - gut interactions
- visceral nociception
Statistics from Altmetric.com
- Visceral pain
- human
- afferent
- electrophysiology
- electrophysiology
- nerve - gut interactions
- visceral nociception
Significance of this study
What is already known about this subject?
Visceral pain is a significant clinical problem for which there are few currently effective treatments.
Rodent studies have examined visceral afferent electrophysiological responses to a variety of stimuli, contributing to the understanding of disease mechanisms and peripheral pain signalling in the gastrointestinal tract.
These methods have not been employed using human tissue.
What are the new findings?
Spontaneous and stimulated afferent fibre activities were recorded in mesenteric nerves from the colon and appendix.
These represent the first such recordings in any human tissue.
How might it impact on clinical practice in the foreseeable future?
The results support the potential development of a preclinical model to evaluate molecular targets involved in mediating human visceral pain.
The model provides a platform on which candidate molecules that have been successful in animal models might be tested in humans in vitro to provide further certainty of bio-efficacy before expensive clinical trials.
Introduction
Abdominal pain is one of the commonest medical presentations,1 with abdominal or pelvic viscera commonly implicated or proved to be the site of origin. Chronic abdominal pain represents a particular problem owing to the paucity of treatments that are not complicated or limited by gastrointestinal (GI) side effects such as nausea and constipation. Whereas inflammatory, obstructive, ischaemic or infiltrative processes may lead to chronic pain, patients with unexplained symptoms are much more common.2 A high proportion of these patients may be defined as having a ‘functional gastrointestinal disorders’, such as ‘irritable bowel syndrome’ on the basis of the association of recurrent, unexplained pain with other GI symptoms.3 Such patients are responsible for up to 40% of patients seen in secondary GI practice with attendant costs estimated at US$40B/year in 10 industrialised nations.4 Such conditions remain orphan areas of research and have been identified by specialist organisations as major burdens to healthcare providers.5
Studies of human GI tissue have been developed to examine the mechanisms underlying secretion and motility, including direct recordings of activity in submucosal neurons of the enteric nervous system.6–9 These studies provide an important functional translational step between studies in rodents and in vivo studies in humans, thereby allowing potential therapeutic targets to be evaluated at an early stage before the development of compounds suitable for clinical use. Although in vitro recordings of extrinsic sensory nerve activity from the rodent GI tract are well established,10 11 the development of such recordings from human tissue remains a challenge. As a consequence, human studies of visceral afferent signalling are currently limited to in vivo studies employing intraluminal stimuli in patients or healthy volunteers (with biomarkers of response limited to symptom reporting or central nervous system imaging) and hence can only be performed once drugs suitable for clinical use are available.12
This report presents preliminary experience and data for afferent recordings from freshly excised human colon and appendix. To our knowledge, this is the first published report of human afferent ex vivo recordings from an end organ in the viscera or in any human tissue.
Patients and methods
Patients
All experiments were performed in accordance with human ethics regulations (NREC 09/H0704/2). Fresh, non-inflamed colon (total n=9) and appendix specimens (total n=18) were obtained from patients (19 male and eight female, median age 60, range 17–84 years) undergoing right or left hemicolectomy at Barts and The London NHS Trust and Homerton University Hospital NHS Foundation Trust. All specimens were obtained from patients with non-obstructive tumours not involving the base of the appendix (right side) and not occurring in the context of inflammatory bowel disease. Two patients had had chemo-radiotherapy for rectal cancer. Colonic specimens were taken away from the tumour, resection margins or lymphatic drainage field with the permission of the histopathologist following macroscopic examination (appendices were returned to the morbid anatomy department after completion of studies). Scattered diverticula were present in some left-sided specimens but avoided in specimen acquisition.
Tissues: broad rationale
The colon is available in quantity from patients undergoing segmental resection for cancer (right, left, sigmoid), and is disposed to ‘flatbed’ testing of mechanical stimuli. Extrinsic nerves of the appendix are embryologically identical to those of the right colon (caecum) being derived from the same progenitors and branching from the same nerve (along ileocolic artery). Furthermore, the appendix is a common site of acute inflammatory pain, is readily available in quantity both in normal (colon cancer) and inflamed states (for future immunological studies).13 It has its own mesentery containing a dominant vessel (appendicular artery) with associated nerves for study (figure 1A) and is of a ‘manageable’ size for use in organ bath experiments (a little akin to the rat colon), including the use of through and through intraluminal perfusion of chemical agents and balloon catheter distension.

Human appendix and mesentery. (A) Appendix in situ with arrows showing mesenteric blood vessels. (B) Organ bath in which electrophysiology recordings were made with a suction electrode (1). Luminal perfusion is facilitated by input (2) and output (3) tubes to which the appendix was secured.
Colon and appendix preparations
Specimens with associated mesentery were resected according to standard surgical protocol. A section of full-thickness colonic wall (approximately 3–4 cm long and 1–2 cm either side of the mesenteric fan) was sharp dissected from the mesenteric aspect with intact mesentery (>3 cm). The appendix was removed in its entirety from the right colon at its base with the full mesentery after division of the bloodless fold of Treves. Specimens were immediately placed in buffer and kept cold on wet ice before being transferred to a bespoke organ bath which was lined with Sylgard (Dow Corning, Dow Corning Corp. MI, USA). The blunt end of appendix was cut and the open ends were attached to input and output tubes. The bath was continually superfused with oxygenated HEPES buffer (in mM: 140.0 NaCl, 10.0 HEPES hemi Na2+, 2.10 CaCl2, 2.12 MgCl2, 2.50 KCl, 10.0 D-glucose; Sigma-Aldrich Inc., MO, USA) at a flow rate of 9 ml/min with temperature maintained at 28–32°C. The colon was pinned flat with the serosal side up and the bath was superfused with carbogenated KREBS buffer (in mM: NaCl 120; KCl 5.9; NaH2PO4 1.2; MgSO4 1.2; NaHCO315.4; CaCl2 2.5; glucose 11.5; Sigma-Aldrich) at the same flow rate and temperature as for the appendix preparation.
Electrophysiological recordings
For the appendix, the mesentery was pinned out proximal to the tissue and the mesentery carefully dissected free. For the colon, mesenteric blood vessels which entered the gut wall were identified and surrounding mesentery was dissected away and the remaining mesentery, containing blood vessels, was pinned to the base of the recording chamber. Colonic nerves running in close proximity to blood vessels were carefully dissected and isolated. Nerves isolated from both preparations were drawn into a borosilicate glass suction electrode (Harvard Apparatus) filled with buffer. Nerve activity was recorded on a Neurolog headstage (Neurolog Ltd), amplified (gain 5K), filtered (band pass 100–2000 Hz) and data acquired (20 kHz sampling rate) to a desktop computer on a Micro 1401 MKII (Cambridge Electronic Design) interface running Spike2 (Cambridge Electronic Design) software. Action potentials were counted using a spike processor (Digitimer) and the threshold level for spike processing was set to the smallest identifiable spike. Nerve activity was expressed as a rate histogram displaying nerve discharge/s in 1 s bin widths.
Experimental protocol
After the isolation of a mesenteric nerve bundle, the preparation was observed for about 10 min so as to record spontaneous activity. No experimental procedure was carried out during this time. In appendix preparations, experimenter-induced activation of nerve fibres was then assessed using 1 ml of inflammatory soup (composed of PGE2 10 μM, histamine 10 μM, serotonin 1 μM, ATP 5 mM, adenosine 1 mM and bradykinin 1 μM) applied directly on to the serosal surface of the tissue, or intraluminal infusion of capsaicin (10 μM) or buffer (2 ml/min). Repeated stimulation was carried out with 5-min intervals between each application.
In colonic preparations a large diameter (0.80 mm) von Frey hair was used to bluntly probe (rather than exert to a point force at its torsional capacity) the serosal surface of the colon for mechanical responses. This hair was chosen to guarantee stimulation of nerve endings throughout the thickness of the colonic wall. After 5 min of stabilisation, the serosal surface of the colon was systematically probed in a circular and radially decreasing fashion, starting approximately 1 cm to the left of the recording nerve entry point into the tissue.
Data and statistical analysis
Baseline activity was determined as the peak firing rate during the 10 min stabilisation. Similarly, the peak firing rate was determined after application of either inflammatory soup or capsaicin. Peak responses to inflammatory soup were compared with peak baseline firing rate, whereas peak increases to capsaicin responses were compared with buffer infusions. Data are presented as mean±SEM. Paired comparisons of data before and after stimuli were made using the Student paired t-test with significance set at p<0.05.
Results
General observations on viability of tissues and neuronal recordings
Although colonic and appendicular tissues were regularly available and no patient refused to participate, the proportion of those in whom suitable nerves were dissectable and spontaneous activity recordable was 4/9 for colon and 9/18 for appendices (overall yield 48% of 27 patients). Factors that predicted difficulties included obesity (due to fat deposition in the mesentery); however, there was no clearly evident effect of intraoperative ischaemic or transport time durations. Other practicalities that may influence future use included cancellation of operation and change in operative procedure, primarily owing to tumour unresectability.
Spontaneous neuronal firing can be measured in the mesentery of the colon and appendix
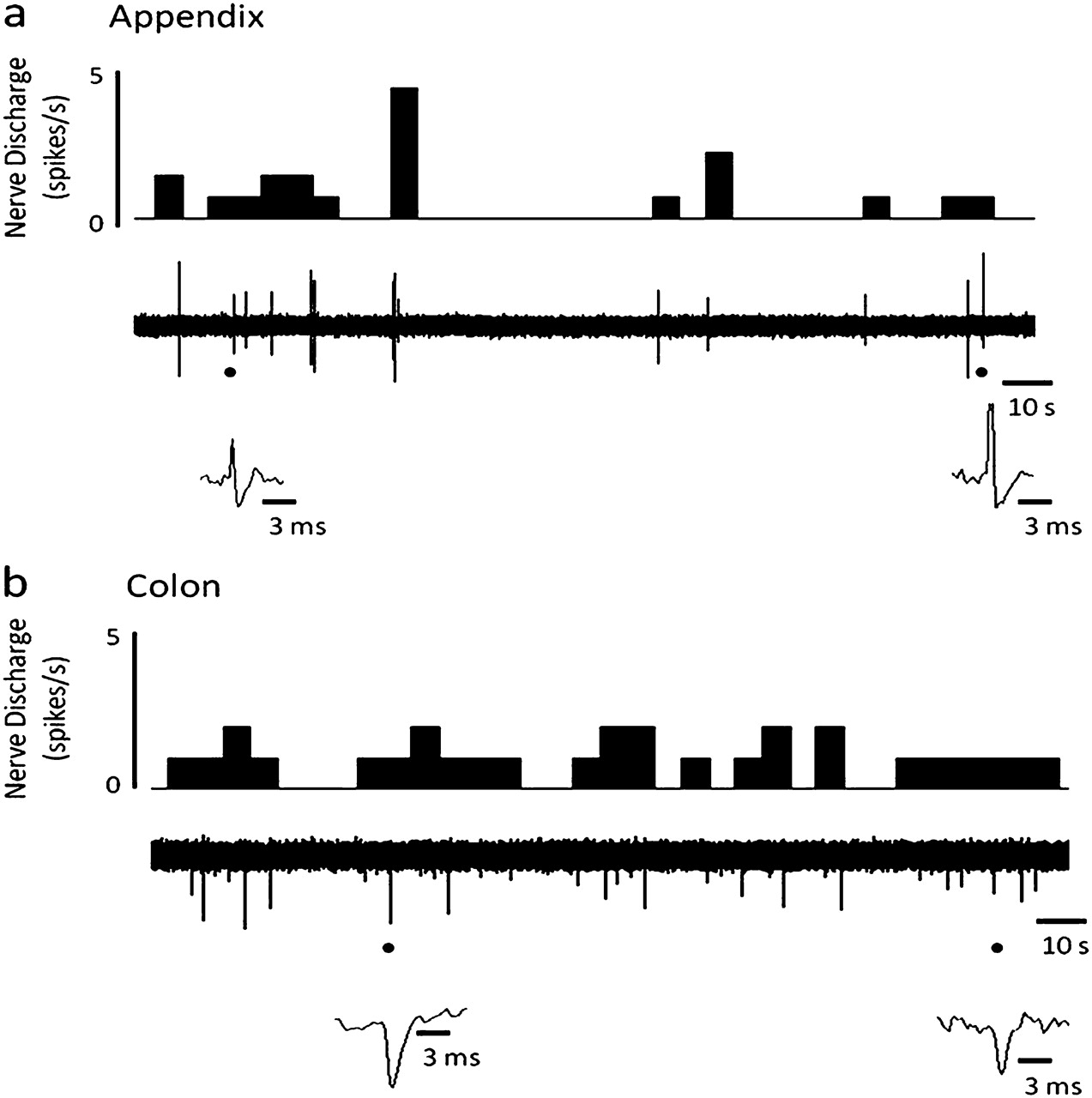
Superficial nerves innervating the wall of normal colon and appendix (figure 1A) displayed spontaneous but irregular neuronal activity ex vivo (figure 2A,B) with consistent (inter-specimen) peak multi-unit afferent neuronal activity (colon: 2.0±0.4 spikes/s, n=4; appendix: 2.4±0.6 spikes/s, n=9) recorded. Such spontaneous discharge was, however, relatively infrequent with lag periods of quiescent firing where 60 s without activity were commonly seen. In the absence of repetitive stimulation, spontaneous afferent activity continued on average for 2–3 h.
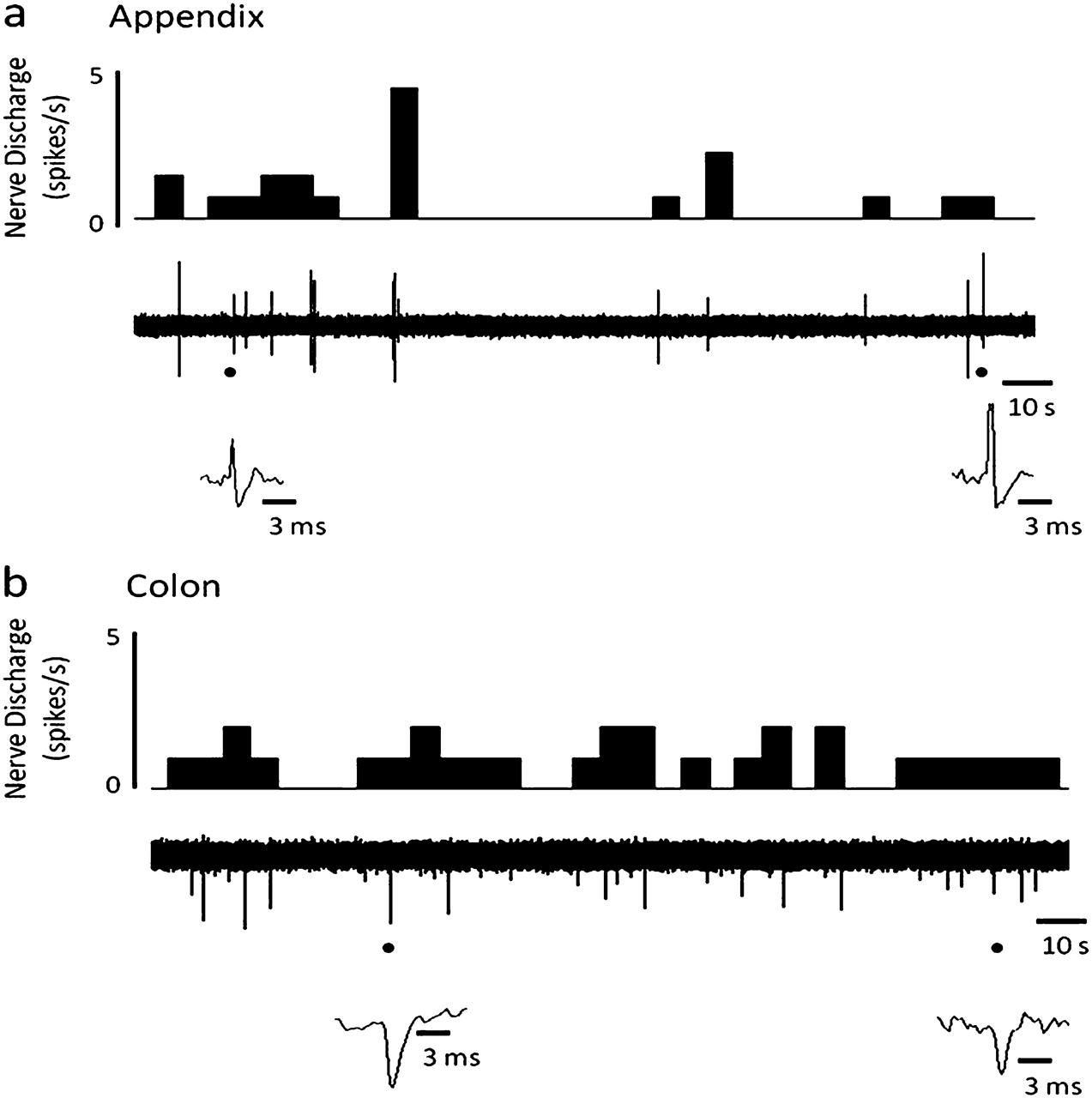
Spontaneous multi-unit neuronal activity of normal human appendix and colon. Raw traces and rate histograms demonstrating spontaneous neuronal firing of the mesenteric nerve innervating (A) the appendix; (B) the colon.
Inflammatory mediators induce increases in appendix afferent activity
Direct application of inflammatory soup resulted in robust transient increases in afferent activity compared with baseline activity (73±10.6 vs 3.0±0.3 spikes/s, n=6, p<0.001) (figure 3A,B). Although the firing rate during stimulation was too high for accurate waveform templates to be formed, an increase in the activity of units with ongoing activity and the recruitment of previously silent units could be clearly seen from the neurogram. Repeated stimulation (at 5-min intervals) of the nerve with inflammatory soup resulted in a decrease in the magnitude of the afferent fibre response with each subsequent stimulation (stimulation 1: 82±12.8 spikes/s, n=4; stimulation 2: 47±4.9 spikes/s, n=4; stimulation 3: 26 spikes/s, n=2) (figure 3C).

Responses to application of inflammatory soup from nerves innervating the human appendix. (A) Raw trace and histogram illustrating the increase in nerve activity following serosal application of inflammatory mediators (PGE2 10 μM, histamine 10 μM, serotonin 1 μM, ATP 5 mM, adenosine 1 mM and bradykinin 1 μM). (B) Stimulation-induced peak firing rate was significantly increased compared with baseline activity (p<0.001, paired t-test, n=6). (C) Repeated stimulation (with 5 min interval between each stimulation) with inflammatory soup resulted in decreased peak firing rate (stimulation 1 and 2, n=4; stimulation 3, n=2).
Capsaicin induces neuronal afferent activity in the appendix
Luminal application of capsaicin (10 μM) resulted in a significant increase in afferent firing (figure 4A,B) compared with buffer control (63±15.8 vs 2.0±0.3 spikes/s, n=3, p<0.001). Similar to the ‘inflammatory soup’ stimulus, although the firing rate during stimulation with capsaicin was too high for accurate waveform templates to be formed, an increase in the activity of units with ongoing activity and the recruitment of previously silent units could be clearly seen from the neurogram.

Responses to application of TRPV1 agonist capsaicin of nerves innervating the human appendix. (A) Raw trace and histogram illustrating the increase in nerve activity following luminal perfusion of 10 μM capsaicin. (B) Stimulation of nerves with capsaicin induces a significant increase in neuronal activity compared with luminal perfusion of buffer (p<0.001, paired t-test, n=3).
Responses to blunt probing of the colon
After 5 min of stabilisation, the von Frey hair was applied to the colon segment (n=4) in a systematic manner until a response was elicited. In two of these, repeated responses to blunt probing identified hot spots (figure 5).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Responses to mechanical stimulation of nerves innervating the human colon. Raw trace and histogram illustrating the increase in nerve activity following mechanical stimulation of the human colon. Arrows indicate points at which a von Frey hair (0.8 mm diameter) was applied to probe the serosal surface of the colon.
Discussion
This study offers first proof of principle that electrophysiological afferent responses can be obtained in a human in vitro nerve organ preparation using basic chemical (inflammatory soup and capsaicin) and mechanical stimuli.
The study is necessarily limited by the small number of tissues successfully studied, reflecting the inherent difficulties of acquiring fresh suitable material (overall yield from consenting patients was 48%). Although for some patients the cause for failure was obvious—for example, mesenteric fat deposition and failure to isolate a suitable nerve, in others the cause was less clear and had no obvious relationship to type of operation, ischaemic time or transfer time to organ bath. Such problems may be overcome by further experience and protocol modification but currently represent limitations to the widespread use of the technique. These effects will, however, be mitigated by the ability to record from multiple preparations from the same tissue specimen, as is the case with experiments using human muscle strips to study efferent neuronal effects on contraction and secretion.14 15
In this study, two classical algogenic stimuli—an ‘inflammatory soup’ containing chemical mediators released from inflamed tissue, and the transient receptor potential vanilloid 1 ligand, capsaicin—were used to examine neuronal responses to chemical stimuli. The components of the inflammatory soup were used to mimic the chemical milieu found in tissues following damage/inflammation, while capsaicin, a well-established naturally occurring ligand of the transient receptor potential vanilloid 1,16 was used as an exemplar of nociceptor activation. Application of either stimulus resulted in robust, transient increases of mesenteric nerve activity, consisting of increased activation of spontaneously active units and recruitment of previously silent units. Sensitisation of visceral afferents has been observed after chemical stimulation, with various components of the inflammatory soup used in this study, as well as capsaicin.17 Therefore, we provide evidence to show that inflammatory mediators, as well as the vanilloid capsaicin, activate peripheral nerves innervating the human appendix. Furthermore, silent afferents have a key role in sensing painful stimuli and their activation following chemical stimulation has been described in the viscera.18 This is in keeping with the clinical observation that non-diseased intra-abdominal viscera are basically insensate to mucosal stimuli—for example, biopsies, diathermy etc, but become sensitised in inflamed disease states.2 Collectively, the results of this study provide functional evidence for the role of excitatory receptors in sensitising peripheral neurons in GI conditions characterised by inflammation and pain. Indeed, these results suggest that receptors which have been shown to mediate visceral pain sensation in animal models—for example, TRPA1, TRPV4, and PAR2/4, may similarly be tested in this in vitro model to determine their role in human GI pain.19–23
Mechanical stimulation by blunt probing of the serosal surface of the colon resulted in activation of mechanosensitive endings. It is accepted that the use of a single wide diameter von Frey hair was rudimentary. This hair was chosen to reliably overcome constraints caused by the much greater thickness of human colonic tissue (compared with rodents). It is possible that we may have directly stimulated the axon by this blunt probing; however, mechanosensitive sites were sparse and focal. Furthermore, sites were non-adjacent; adjacency would be expected if probing of a linear projecting axon was occurring. This early report did not include detailed characterisation of fibre response profiles by spike sorting (high threshold, low threshold and wide dynamic range), or of sites of nerve endings as determined by dye filling.24 Such studies are planned and will be necessary to yield further characterisation data on human visceral mechanosensitivity.
This study does not represent the only direct recording from human sensory nerves; previous in vivo studies have used transcutaneous microneurography in the somatic nervous system25 and studies of afferent activity have also been performed on isolated human vagal and somatic C fibre axons.26 27 However, this study has significant advantages which complement the existing publications on human studies. First, unlike studies on isolated human nerves, we were able to study the transduction of stimuli at the nerve-terminal organ level as opposed to the activation of isolated nerves by channels expressed along the axon, which may not reflect their function at afferent fibre terminals. Second, we were able to study a broader range of mechanisms underlying the activation of afferent fibres at their terminal organ than is currently possible with in vivo human microneurography studies. These advantages will permit an early indication of mechanisms important in the activation of human sensory nerves and, potentially, direct therapeutic research towards mechanisms which have a much higher probability of providing clinical utility.
References
Footnotes
See Commentary, p 151
MP and DB are joint first authors on this paper.
Funding This work was supported by CORE Digestive Diseases (CHK, QA and MP) and The Royal College of Surgeons (SS).
Competing interests None.
Ethics approval This study was conducted with the approval of the East London and The City HA Local Research Ethics Committee (NREC 09/H0704/2).
Provenance and peer review Not commissioned; externally peer reviewed.