

Abstract
Mitogen-activated protein (MAP) kinase plays important roles in the establishment of long-term potentiation both in vitroand in living animals. MAP kinase is activated in response to a broad range of stimuli, including calcium influx through NMDA receptor and L-type calcium channel, cAMP, and neurotrophins. To investigate the role of Ras in the activation of MAP kinase and cAMP response element-binding protein (CREB) in hippocampal neurons, we inhibited Ras function by overexpressing a Ras GTPase-activating protein, Gap1m, or dominant negative Ras by means of adenovirus vectors. Gap1m expression almost completely suppressed MAP kinase activation in response to NMDA, calcium ionophore, membrane depolarization, forskolin, and brain-derived neurotrophic factor (BDNF). Dominant negative Ras also showed similar effects. On the other hand, Rap1GAP did not significantly inhibit the forskolin-induced activation of MAP kinase. In contrast to MAP kinase activation, the inactivation of Ras activity did not inhibit significantly NMDA-induced CREB phosphorylation, whereas BDNF-induced CREB phosphorylation was inhibited almost completely. These results demonstrate that Ras transduces signals elicited by a broad range of stimuli to MAP kinase in hippocampal neurons and further suggest that CREB phosphorylation depends on multiple pathways.
Ras is a small guanine nucleotide-binding protein the function of which is essential for the proliferation, differentiation, and survival of various types of cells. Ras is a molecular switch that recycles between inactive GDP-bound and active GTP-bound states, and the latter returns to the inactive state by the intrinsic GTPase activity accelerated by GTPase-activating proteins (GAPs) (Vojtek and Der, 1998; for review, see Zwartkruis and Bos, 1999). Ras transduces signals from various stimuli to downstream effectors, including Raf [an activator for mitogen-activated protein kinase (MAP kinase)/extracellular-regulated kinase (ERK) pathway], phosphatidylinositol-3 kinase (PI3-kinase), and Ral guanine nucleotide dissociation stimulator (RalGDS), inducing many cellular responses (Vojtek and Der, 1998; Zwartkruis and Bos, 1999).
In the CNS, Ras and its regulators are highly expressed and some have been shown to localize at synapses (Chen et al., 1998; Ye et al. 2000). The phenotypes of mutant mice for Ha-Ras (Manabe et al. 2000), RasGRF (Brambilla et al., 1997), and NF1 (Silva et al., 1997) genes have been reported, the former of which shows enhanced long-term potentiation (LTP), and the latter two show some defects in memory consolidation. However, the cell biological mechanisms of Ras function in neuronal cells remain to be clarified.
Calcium influx through NMDA receptor and L-type calcium channel is essential for the establishment of LTP (for review, see Ghosh and Greenberg, 1995; Bading et al., 1997). Neurotrophins and cAMP signaling are requisite for LTP formation (Korte et al., 1995; Wong et al., 1999). All of these signaling pathways activate MAP kinase, which also is indispensable for the establishment of LTP and learning in living animals (English and Sweatt, 1997; Atkins et al., 1998). Therefore, it is of particular interest to analyze the mechanism of MAP kinase activation in neuronal signaling.
Many signals converge on MAP kinase (for review, see English et al., 1999; Curtis and Finkbeiner, 1999; Impey et al., 1999). Growth factors activate MAP kinase in a Ras-dependent manner, whereas protein kinase C and Mos kinase activate MAP kinase in a Ras-independent manner. Calcium- and cAMP-induced activation of MAP kinase is a matter of disagreement. Several studies have described Ras-dependent activation (Rosen et al., 1994; Busca et al. 2000), whereas others have demonstrated that Rap1, a Ras-related small GTPase, mediates the activation (Vossler et al., 1997; Grewal et al. 2000). However, because these studies were performed using cultured cell lines, the mechanism of MAP kinase activation should be examined using neuronal cells.
In this study we examined the requirement of Ras for the activation of MAP kinase and CREB induced by NMDA, membrane depolarization, forskolin, and BDNF in hippocampal neurons. We inhibited Ras activity using adenoviruses expressing a Ras GAP, Gap1m, or dominant negative Ras. The results described here clearly indicate that Ras is requisite for MAP kinase activation in response to a broad range of stimuli in hippocampal neurons and further suggest that CREB phosphorylation depends on multiple pathways.
MATERIALS AND METHODS
Adenoviruses and expression vectors. Adenoviruses expressing Gap1m(Adex-Gap1m), a dominant negative Ras (AdRasY57 in which the 57th residue of Ha-Ras, aspartate, was replaced with tyrosine) (Ueno et al., 1997), and Rap1GAP (Adex-Rap1GAP) were constructed according to Miyake et al. (1996). Adex-LacZ was kindly donated by Dr. I. Saito (Laboratory of Molecular Genetics, University of Tokyo). Adenoviruses were purified by a CsCl gradient ultracentrifugation method (Kanegae et al., 1994). pEFBOS-MycGap1m and pEFBOS-MycΔGap1m were produced by inserting Myc-tagged full-length Gap1m(Maekawa et al., 1994) or Gap1m lacking GAP domain into a mammalian expression vector, pEFBOS (Mizushima and Nagata, 1990). Rap1GAPII cDNA (Mochizuki et al., 1999) was a kind gift of Dr. M. Matsuda (Department of Pathology, Research Institute, International Medical Center of Japan).
Cell culture and adenovirus infection. Hippocampal neurons were prepared from embryonic day 18–19 rat embryos as described (Brewer, 1995). Cells were dissociated by trypsin and plated on 35-mm-diameter plates (5 × 105 cells per plate) precoated with poly-l-lysine (0.5 mg/ml) in DMEM containing 10% fetal calf serum. At 1 day in vitro (DIV) the medium was changed to Neurobasal Medium (Life Technologies, Gaithersburg, MD) containing B27 supplement and 3 μm cytosine arabinoside. At DIV 6 the cells were infected with adenoviruses at multiplicity of infection (moi) 500.
Two days after infection, the culture medium was replaced by balanced salt solution containing (in mm): 20 HEPES-NaOH, pH 7.5, 130 NaCl, 5.4 KCl, 2 CaCl2, and 5.5 glucose, and after 2 hr incubation, cells were stimulated with reagents for various periods of time that are specified in the figure legends. For NMDA stimulation, 6-cyano-7-nitro-quinoxaline-2,3-dione (CNQX) (40 μm) and nimodipine (5 μm) were added 20 min before NMDA treatment to inhibit AMPA receptor and L-type calcium channel, respectively. When cells were depolarized with KCl (60 mm), dl-2-amino-5-phosphonopentanoic acid (AP5) (100 μm) and CNQX were added. Forskolin (100 μm) treatment was performed in the presence of three channel antagonists. Tetrodotoxin (1 μm) was added to cultures 15 hr before stimulation to reduce spontaneous synaptic activity. More than 90% of cells were judged to be neurons by the staining for microtubule-associated protein 2 (MAP2) and glial fibrillary acidic protein (GFAP). BDNF was kindly supplied by Dr. C. Nakayama (Sumitomo Pharmaceuticals Research Center).
PC12 cells were maintained in DMEM containing 10% fetal calf serum and 5% horse serum. Cells were infected with adenoviruses at moi 100 and incubated for 2 d, then stimulated with nerve growth factor (NGF) for 5 min. DNA transfection was performed using Lipofectamine 2000 reagent (Life Technologies) according to the manufacturer's protocol.
Western blotting. The cells were lysed in SDS sample buffer and boiled for 10 min. The lysates (corresponding to 3 × 104 cells) were subjected to 10% SDS-PAGE, and proteins in the gel were transferred onto polyvinylidene difluoride membranes (Millipore, Bedford, MA). The filters were blocked with Tris-buffered saline containing 0.1% NP40 and 5% low-fat dry milk and incubated with the primary antibodies for 2–4 hr and then with horseradish peroxidase (HRP)-conjugated secondary antibodies for 1 hr at room temperature. The signals were visualized by using an ECL chemoluminescence kit (Amersham-Pharmacia Biotech, Arlington Heights, IL). The following primary antibodies were used: rabbit anti-phospho-MAP kinase (NEB), rabbit anti-MAP kinase (UBI), rabbit anti-phospho-CREB (UBI), rabbit anti-phospho-MEK (NEB), and mouse anti-Myc tag, 9E10 (Santa Cruz Biotechnology, Santa Cruz, CA). HRP-conjugated secondary antibodies were from Amersham-Pharmacia Biotech.
Ras pull-down assay. The relative amount of Ras-GTP was determined according to a method described by de Rooij and Bos (1997)using Ras binding domain of Raf fused to glutathioneS-transferase (GST) (a gift of Dr. M. Matsuda) and monoclonal antibody against Ras, NCC-RAS-004 (Kanai et al., 1987) (a gift of Dr. S. Hirohashi at Pathology Division, National Cancer Center Research Institute).
Rap1 pull-down assay. COS7 cells grown on 60 mm dishes (1 × 106 cells per plate) were transfected with pCXN2-FLAG-Rap1 (Mochizuki et al., 1999) or pCXN2-FLAG-Rap1 plus pCAGGS-C3G (Gotoh et al., 1995) and infected with Adex-LacZ or Adex-Rap1GAP at moi 10. Two days later, the cell lysates were prepared and incubated with GST-RalGDS (Franke et al., 1997) to isolate Rap1-GTP. The relative amount of active Rap1 was determined by Western blot using anti-Rap1 antibody (Santa Cruz Biotechnology). Cortical neurons were prepared as described above and plated on 90-mm-diameter dishes at the density of 2 × 107 cells per plate. At DIV 4, cells were infected with either Adex-LacZ or Adex-Rap1GAP at 10 moi, and the cells were further cultured for 2 d. Then the cells were stimulated with a mixture of forskolin (100 μm) and 12-O-tetradecanoylphorbol-13-acetate (TPA; 1 μm) and depolarized with KCl (60 mm) for 15 min. The cell lysates from three dishes were combined and processed to the Rap1 pull-down assay as above.
Immunostaining. Cells were fixed in 4% paraformaldehyde for 7 min at room temperature and permeabilized with methanol for 7 min at −20°C. Cells were incubated with appropriate first antibodies at room temperature for 2 hr and with fluorescein isothiocyanate- and rhodamine-conjugated secondary antibodies (Jackson Laboratories) for 1 hr.
RESULTS
To investigate the function of Ras in the signal transduction in neuronal cells, we constructed adenovirus vectors that express a GAP for Ras, Adex-Gap1m, or AdRasY57. We first examined whether Adex-Gap1m suppressed Ras function in PC12 rat pheochromocytoma cells (Fig.1). PC12 cells were infected with either Adex-Gap1m or a control virus, Adex-LacZ expressing β-galactosidase, at moi 100 and incubated for 2 d. The cells were then stimulated with various concentrations of NGF for 5 min, and the activation of Ras and MAP kinase was examined by a pull-down assay using Ras binding domain of Raf fused to GST to detect active GTP-bound Ras (de Rooij and Bos, 1997), and by Western blot using an active form-specific antibody to MAP kinase (Fig.1A). The active Ras in Adex-Gap1m-infected cells was barely detectable at moderate concentrations of NGF and significantly lower than in control cells even at 100 ng/ml NGF.

Inhibition of Ras/MAP kinase pathway by Adex-Gap1m (A) and pEFBOS-MycGap1m (B) in PC12 cells. A, PC12 cells were infected with Adex-LacZ or Adex-Gap1m at moi 100 and incubated for 2 d. Cells were stimulated with the indicated concentrations of NGF for 5 min. The relative amount of active Ras and phosphorylated MAP kinase (pMAPK) was analyzed as described in Materials and Methods. B, PC12 cells were transfected with pEFBOS-MycGap1m or pEFBOS-MycΔGap1m and incubated for 2 d. Cells were treated with 100 ng/ml NGF for another 2 d and immunostained for Myc-tagged proteins (myc) or neurofilaments (NF). Arrows indicate cells expressing Myc-tagged proteins.
Consistent with this inhibition, Adex-Gap1m markedly inhibited the activation of MAP kinase by NGF (Fig. 1A). We also observed that Adex-Gap1m completely inhibited MAP kinase activation induced by membrane depolarization in PC12 cells, indicating that this MAP kinase activation is also a Ras-dependent process (data not shown). The result was consistent with the result reported by Rosen et al. (1994) that depolarization-induced activation of MAP kinase is Ras-dependent in PC12 cells.
As a next step, the effect of Gap1mexpression on neurite outgrowth of PC12 cells by NGF was examined (Fig.1B). The cells were transfected with a mammalian expression plasmid, pEFBOS-MycGap1m or pEFBOS-MycΔGap1m(Gap1m lacking GAP domain), incubated for 2 d, and then treated with 100 ng/ml NGF for another 2 d. Cells expressing the full-length Gap1m did not show NGF-induced neurite outgrowth, but cells expressing ΔGap1m still extended neurites. These results indicated that both Adex-Gap1m and pEFBOS-MycGap1m suppressed Ras function in PC12 cells.
We then examined whether Adex-Gap1msimilarly inhibited Ras function in hippocampal neurons (Fig.2). Hippocampal neurons at DIV 8 were stimulated with 10 μm NMDA or 100 ng/ml BDNF for 5 min. The amount of active GTP-bound Ras in the cells was increased by both treatments (Fig. 2A). Infection with Adex-Gap1m but not Adex-LacZ markedly diminished the amount of active Ras in both untreated and BDNF-treated cells. When the expression of MycGap1m was examined by immunostaining of the culture with anti-Myc antibody, 9E10, almost all the cells expressed MycGap1m(data not shown).

Inhibition of Ras activation by Adex-Gap1m (A) and MAP kinase activation by pEFBOS-MycGap1m(B) in hippocampal neurons. A, Hippocampal neurons at DIV 6 were infected with Adex-LacZ (top panel) or with either Adex-LacZ or Adex-Gap1m (bottom panel) at moi 500. Two days later, the cells were stimulated with 10 μm NMDA or 100 ng/ml BDNF for 5 min, and the relative amount of active Ras (indicated by arrows) was determined as in Figure 1. B, Cells were transfected with pEFBOS-MycGap1m and incubated for 2 d. Cells were then stimulated with BDNF (100 ng/ml) or NMDA (10 μm) for 5 min and immunostained with anti-Myc (myc) or anti-phospho-MAP kinase (pMAPK) antibodies. Myc-positive cells and negative cells are indicated by arrows andarrowheads, respectively.
Having shown that Gap1m was capable of downregulating Ras function in neuronal cells, we examined whether NMDA- and BDNF-induced MAP kinase activation was Ras dependent. Neuronal cultures were transfected with pEFBOS-MycGap1m and stimulated with BDNF or NMDA 2 d after transfection. Cells were then immunostained with the antibody specific to the phosphorylated form of MAP kinase (Fig.2B). Cells expressing Gap1m (indicated by arrows) did not show immunoreactivity of active MAP kinase in response to BDNF or NMDA, whereas surrounding cells that did not express Gap1m were positive in the immunostaining (arrowheads). The result clearly indicated that Ras activity is necessary for BDNF- and NMDA-induced MAP kinase activation.
We further analyzed biochemically the involvement of Ras function in the phosphorylation of MAP kinase family and CREB in hippocampal neurons by infection of Adex-LacZ, Adex-Gap1m, or AdRasY57 expressing dominant negative Ras (Ueno et al., 1997) (Fig.3). As reported previously, application of NMDA, KCl, ionomycin, and BDNF induced phosphorylation of MAP kinase, MEK (a direct activator for MAP kinase), and CREB in cultured hippocampal neurons that had been infected with a control virus, Adex-LacZ. Depletion of extracellular calcium by EGTA completely abolished MAP kinase phosphorylation induced by NMDA, indicating that calcium influx was critical for MAP kinase phosphorylation in this system (data not shown).

Inhibition of MAP kinase activation by NMDA, membrane depolarization, ionomycin, and BDNF by Adex-Gap1m and AdRasY57. Hippocampal neurons (DIV 6) were infected with Adex-LacZ (LacZ), Adex-Gap1m (Gap1m), or AdRasY57 (dnRas) at moi 500 and incubated for 2 d. The cells were then stimulated with NMDA (100 μm), KCl (60 mm), ionomycin (0.5 μg/ml), or BDNF (100 ng/ml) for 5 min. The cell lysates (corresponding to 3 × 104 cells) were subjected to Western blots using antibodies against the phosphorylated form of MAP kinase (pMAPK), MEK (pMEK), and CREB (pCREB), as well as the antibody recognizing both dephosphorylated and phosphorylated forms of MAP kinase (total MAP kinase). Representative data of three independent experiments are shown.
The expression of Gap1m almost completely suppressed the phosphorylation of MAP kinase and MEK by NMDA, KCl, or ionomycin (Fig. 3). The result demonstrates that Ras function is indispensable for the activation of MAP kinase induced by calcium influx in these cells. BDNF stimulation of MAP kinase phosphorylation was also a Ras-dependent process. Dominant negative Ras inhibited the activation of MAP kinase and MEK in a manner similar to that of Gap1m except at a lower magnitude.
Because it has been shown that the MAP kinase pathway phosphorylates CREB (English and Sweatt, 1996; Impey et al., 1998; Hardingham et al., 1999), we examined the effect of Gap1mexpression on the phosphorylation of CREB (Fig. 3). Phosphorylation of CREB induced by BDNF was suppressed by Gap1m or dominant negative Ras in proportion to the inhibition of MAP kinase activation (Fig. 3). In contrast, CREB phosphorylation induced by NMDA, KCl, or ionomycin was not significantly inhibited by Gap1m or dominant negative Ras.
We further examined quantitatively the effect of Gap1m on the phosphorylation of MAP kinase and CREB induced by NMDA or BDNF at various concentrations (Fig.4). Gap1mdecreased the basal level of phosphorylated MAP kinase and almost completely inhibited the activation of MAP kinase by NMDA and BDNF. However, although BDNF-induced CREB phosphorylation was decreased to the basal level in Gap1m-expressing cells, NMDA-induced CREB phosphorylation was marginally inhibited.

Dose dependency of NMDA and BDNF and the effect of Adex-Gap1m infection on the phosphorylation of MAP kinase and CREB. Neurons were infected with Adex-LacZ (gray bars) or Adex-Gap1m(black bars) at moi 500 and stimulated with various concentrations of BDNF or NMDA for 5 min as in Figure 3(n = 3). The relative amounts of phosphorylated MAP kinase and CREB are quantitated using NIH Image software and shown with SDs.
The results described above suggest that a kinase or kinases other than the MAP kinase pathway may phosphorylate CREB in NMDA-stimulated cells. However, it was possible that Gap1minhibition on MAP kinase activation was not complete and that the residual activity might be responsible for CREB phosphorylation. Therefore, we further studied the effect of MAP kinase inhibition on the phosphorylation of CREB by using pharmacological inhibitors (Figs. 5,6). First, cells were treated with PD098059, an inhibitor of MEK (Pang et al., 1995). Little but measurable activation of MAP kinase was still observed either in cells pretreated with 50 μm PD098059 or in cells infected with Adex-Gap1m. The combination of PD098059 and Adex-Gap1m resulted in the almost complete inhibition of MAP kinase activation by NMDA stimulation. However, phosphorylation of CREB by NMDA was impaired only partially. Under the same condition, BDNF-induced CREB phosphorylation was suppressed almost completely (data not shown). We also examined the effect of U0126, which is more potent and specific than PD098059 in the inhibition of the MAP kinase pathway (DeSilva et al., 1998) (Fig. 6). In U0126-treated cells, MAP kinase activation was hardly visible, but CREB phosphorylation by NMDA treatment was similar to that observed in control cells. In contrast, both BDNF-induced MAP kinase activation and CREB phosphorylation were suppressed to the basal level under the same condition (Fig. 6). These results taken together strongly suggest that other signaling pathways leading to CREB phosphorylation may be functioning in this system.

The effect of PD098059 and Adex-Gap1m on the activation of MAP kinase and CREB. Cells (DIV 6) were infected with Adex-LacZ (LacZ) or Adex-Gap1m (Gap1m) and incubated for 2 d. Cells pretreated with either 50 μm PD098059 or a control vehicle solution for 30 min were stimulated with 1, 3, and 10 μm NMDA for 5 min (n = 3). The phosphorylation of MAP kinase and CREB was examined as in Figure 3 (top panel) and quantitatively shown (bottom panel).

The effect of a MEK inhibitor, U0126, on NMDA- and BDNF-induced CREB phosphorylation. Cells (DIV 6) pretreated with 20 or 40 μm U0126 for 30 min were stimulated with 10 μm NMDA or 10 μg/ml BDNF for 5 min (n = 3). The phosphorylation of MAP kinase and CREB was examined as in Figure 5.
It is reported that NGF induces the rapid and transient activation of MAP kinase in a Ras-dependent manner and the prolonged activation of MAP kinase through Rap1 activation (York et al., 1998). Therefore, we examined the effect of Gap1m expression on NMDA-induced phosphorylation of MAP kinase and CREB at various time points (Fig. 7). Biochemical analysis showed that Gap1m stimulates GTPase activity of Ras but not that of Rap1 (Maekawa et al., 1993). MAP kinase phosphorylation induced by NMDA reached a maximum level within 2 min, and the level was sustained for up to 60 min in Adex-LacZ-infected cells. CREB phosphorylation followed a similar activation profile and then gradually declined during a period of 60 min. Gap1m suppressed the phosphorylation of MAP kinase induced by NMDA at all time points. Unlike NGF stimulation of MAP kinase in PC12 cells, the activation of MAP kinase by NMDA was Ras dependent in both the early and prolonged phases. Consistent with the result in the previous section, CREB phosphorylation was not inhibited significantly by Gap1mexpression.

The effect of Adex-Gap1m on the phosphorylation of MAP kinase and CREB in NMDA-stimulated hippocampal neurons. Neuronal cells (DIV 6) were infected with Adex-LacZ (▪) or Adex-Gap1m (●) and stimulated with NMDA (10 μm) for the indicated periods of time. The cell lysates were subjected to Western blots as in Figure 3, and the relative amounts of phosphorylated MAP kinase and CREB were quantitated (n = 3).
cAMP is an important secondary messenger in neurons, because adenylyl cyclase is essential for long-term potentiation (Wong et al., 1999). In PC12 cells, cAMP activates MAP kinase in a Rap1-dependent manner (Vossler et al., 1997; Grewal et al. 2000). However, recent reports described Ras activation by a cAMP-inducing reagent, forskolin, in cortical neurons (Ambrosini et al. 2000) and in melanocytes (Busca et al. 2000). Therefore, we sought to determine which small GTPase played a major role in the activation of MAP kinase in our system by using Adex-Gap1m and Adex-Rap1GAP.
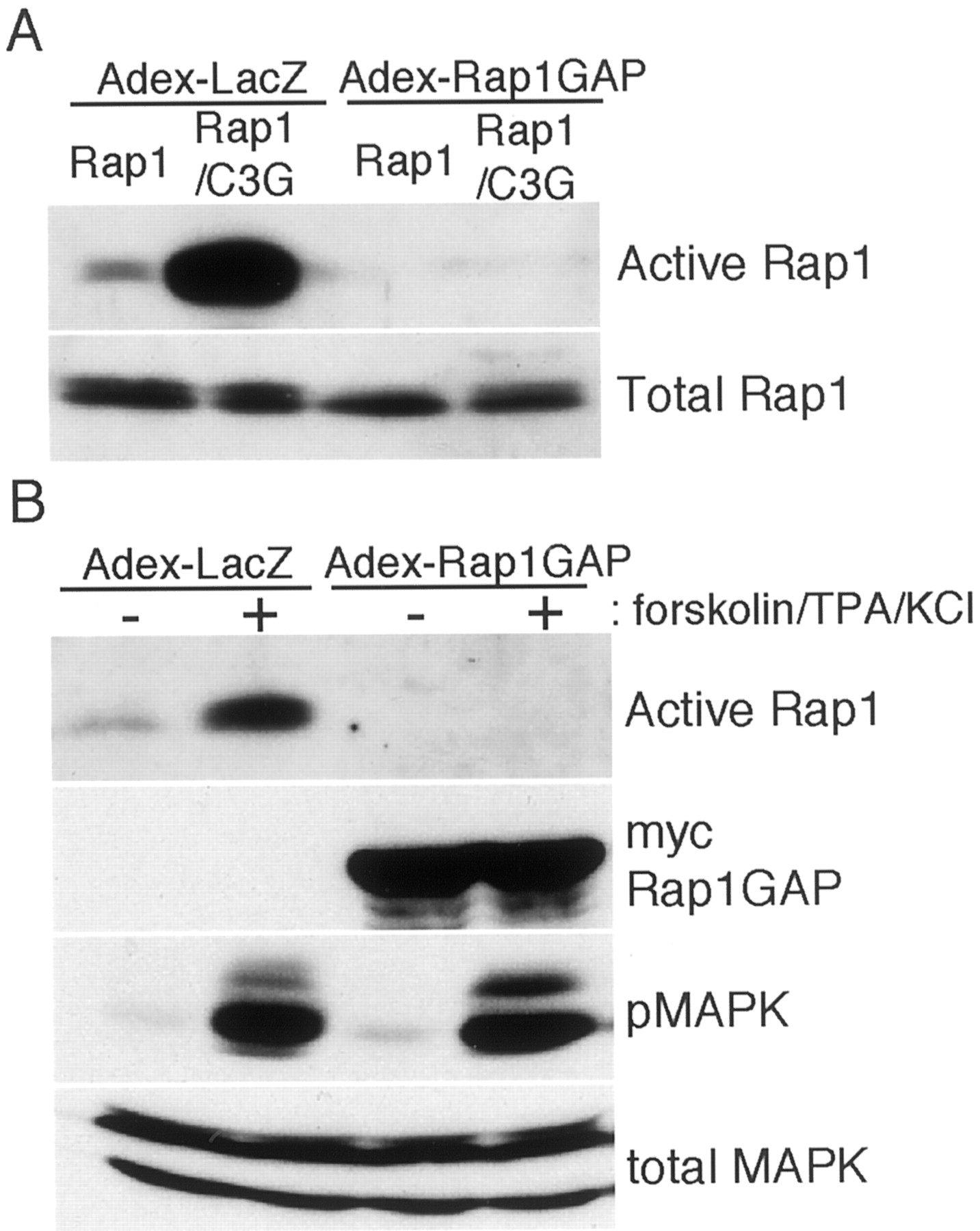
In COS7 cells, expression of C3G, a guanine nucleotide exchange factor for Rap1 (Gotoh et al., 1995), markedly enhanced the amount of active Rap1 (Fig. 8A). Infection of COS7 cells with Adex-Rap1GAP at moi 10 decreased the amount of active Rap1 below the limits of our detection. In cortical neurons, a mixed stimuli including forskolin, TPA, and KCl depolarization activated Rap1 as shown in Figure 8B. As in COS7 cells, Adex-Rap1GAP infection at moi 10 decreased the amount of Rap1-GTP to undetectable levels in both untreated and stimulated cells. Although Rap1 activation was completely blocked by Adex-Rap1GAP, the activation of MAP kinase was still observed. Therefore, we then examined the effect of Ras inactivation on the activation of MAP kinase by the cAMP pathway.

Inhibition of Rap1 activation by Adex-Rap1GAP.A, COS7 cells grown on 60 mm dishes (1 × 106 cells per plate) were transfected with pCXN2-FLAG-Rap1 (Mochizuki et al., 1999) or pCXN2-FLAG-Rap1 plus pCAGGS-C3G (Gotoh et al., 1995) and infected with Adex-LacZ or Adex-Rap1GAP at moi 10. Two days later, the active Rap1 and total Rap1 were analyzed as described in Materials and Methods. B, Cortical neurons (2 × 107 cells per 90-mm-diameter dishes) at DIV 4 were infected with either Adex-LacZ or Adex-Rap1GAP (expressing Myc-tagged Rap1GAP) at 10 moi, and the cells were further cultured for 2 d. Cells were stimulated with a mixture of forskolin (100 μm) and TPA (1 μm) and then KCl depolarized (60 mm) for 15 min. Cell lysates from three dishes were combined, and the relative amounts of active Rap1 in the lysates were determined by pull-down assay. The expression of Myc-Rap1GAP, the activation of MAP kinase (pMAPK), and the amount of total MAP kinase were analyzed as in Figure 3. Representative data of two independent experiments are shown.
Treatment of hippocampal neurons by forskolin activated MAP kinase at 5 and 30 min after the treatment (Fig. 9). At both time points, Adex-Gap1m almost completely inhibited MAP kinase activation. In contrast, inactivation of Rap1 by Adex-Rap1GAP did not show the marked inhibition. The result suggests that Ras is a key molecule in mediating cAMP signal to MAP kinase and that Rap1 does not contribute mainly to MAP kinase activation. Because cAMP-dependent protein kinase directly phosphorylates CREB (Gonzalez and Montminy, 1989), CREB phosphorylation was not inhibited by either Adex-Gap1m or Adex-Rap1GAP. We also examined whether the inhibition of Rap1 activity by Adex-Rap1GAP affected the activation of MAP kinase by NMDA or membrane depolarization. In neither case did Adex-Rap1GAP significantly decrease the levels of MAP kinase activation (data not shown).

The effect of Adex-Gap1m or Adex-Rap1GAP on forskolin-induced activation of MAP kinase and CREB. Cells (DIV 6) were infected with the indicated adenoviruses at moi 500 and incubated for 2 d. Cells were stimulated with 100 μm forskolin for various times specified in the figure. At each point, activation of MAP kinase and CREB was examined as in Figure 3.
DISCUSSION
It has been shown that diverse signals activate MAP kinase in neuronal cells (for review, see Curtis and Finkbeiner, 1999; English et al., 1999; Impey et al., 1999). These include growth factors, cAMP, protein kinase C, and portions of G-protein-coupled receptors. However, except for growth factors, the involvement of Ras in the activation of MAP kinase has not been investigated thoroughly.
In this study we inhibited Ras activity by adenovirus-mediated gene expression, which enabled us to perform biochemical examinations in neuronal cells. Because of low efficiency of DNA transfection in neuronal cells, such analyses have not been performed. By using primary hippocampal cultures, we could stimulate the cells with defined reagents. The results shown here demonstrate that Ras is a prerequisite to the activation of MAP kinase by NMDA-induced calcium entry into hippocampal neuron cultures. Ras activity is also required for MAP kinase activation by depolarization with KCl or by ionomycin. BDNF- and forskolin-induced MAP kinase activation as well is Ras dependent. These results indicate that Ras transduces signals elicited by various stimuli to MAP kinase in hippocampal neurons.
Regarding calcium-induced signals, Bading and Greenberg (1991) andEnglish and Sweatt (1996) demonstrated that the calcium entry through NMDA receptor activates MAP kinase. Membrane depolarization activates Ras in PC12 cells (Rosen et al., 1994) and in cortical neurons (Farnsworth et al., 1995) and MAP kinase in a Ras-dependent manner in PC12 cells (Rosen et al., 1994). In contrast, another study reported that depolarization-induced activation of MAP kinase in PC12 cells is mediated by Rap1 (Grewal et al., 2000). Although the role of Rap1 in calcium signaling is still controversial, our results demonstrate that Ras plays a major role in both NMDA- and depolarization-induced MAP kinase activation in hippocampal neurons.
Downregulation of Ras activity inhibited MAP kinase activation by a broad range of stimuli as described above; however, the inactivation of Ras activity did not significantly inhibit the NMDA-induced CREB phosphorylation. The result suggests that other pathways leading to CREB phosphorylation may be activated by NMDA treatment. When BDNF-induced activation of MAP kinase was inhibited by Adex-Gap1m, CREB phosphorylation was similarly suppressed, suggesting that the Ras/MAP kinase pathway could phosphorylate CREB. However, further studies are necessary to determine whether the Ras/MAP kinase pathway is also involved in NMDA-induced CREB phosphorylation in our system. Several kinases, including cAMP-dependent protein kinase, calmodulin-dependent kinase (CaMK)II, CaMKIV, and RSK2 (activated downstream of MAP kinase), can phosphorylate the same residue of CREB, serine-133 (for review, seeSilva et al., 1998). Therefore, it may be possible that the inhibition of one signaling pathway would not result in the significant decrease of CREB phosphorylation when multiple signaling pathways would be activated.
Hardingham et al. (1999) reported that both CaMK- and MAP kinase-dependent pathways function in the phosphorylation of CREB induced by KCl depolarization, because the combination of KN62, an inhibitor for CaMKs, and PD098059, but not each alone, inhibited the phosphorylation of CREB. Impey et al. (1998) demonstrated that RSK2 but not CaMKIV mainly phosphorylates CREB in depolarized hippocampal neurons, and that PD098059 inhibited CREB phosphorylation. Chawla et al. (1998) reported that activated Ras could induce CREB phosphorylation in AtT20 cells. Deissoroth et al. (1996) demonstrated that CaMKs were CREB kinase activated by calcium influx through NMDA and AMPA receptors. The reasons for the differences among these studies are not clear at present, and they might reflect the differences in the model systems and stimulation protocols used in these studies. In the slice culture, PD098059 inhibits CREB phosphorylation induced by high-frequency stimulation (English and Sweatt, 1996; Impey et al., 1998). However, in slice cultures, the involvement of glutamate receptors other than NMDA receptor should be taken into consideration.
The mechanism of Ras activation by raised calcium has not been clarified yet; however, several pathways could be possible: (1) the activation of RasGRF by calmodulin (Farnsworth et al., 1995); (2) the activation of adenylyl cyclase by calmodulin followed by cAMP activation of CNrasGEF (Pham et al. 2000); (3) the inactivation of SynGAP, a negative regulator of Ras in neurons, by activated CaMKII (Chen et al., 1998); (4) the transactivation of EGF receptor-like molecule by calcium (Zwick et al., 1997); and (5) the direct activation of Ras by nitric oxide produced by calmodulin-activated NO synthase (Yun et al., 1998). Although we do not have any results concerning the above mentioned hypotheses, it would be quite intriguing to reveal the molecular mechanism of Ras activation by calcium in neuronal cells.
We have shown that Ras is also required for transduction of cAMP signal to MAP kinase activation. Ras activator, CNrasGEF (Pham et al. 2000), and Rap1 activator, Epac/cAMP-GEF (de Rooij et al., 1998; Kawasaki et al., 1998), are directly activated by cAMP independent of protein kinase A. Therefore, both Ras and Rap1 could activate MAP kinase in response to cAMP. However, our results showing that inhibition of Ras but not Rap1 markedly reduced the activation of MAP kinase by forskolin suggests Ras as a major transducer in cAMP activation of MAP kinase. Recently, Ambrosini et al. (2000) described Ras activation by forskolin in cortical neurons. Another Ras GEF, RasGRP, is activated by calcium and diacylglycerol (Ebinu et al., 1998). Therefore, Ras could be activated by receptor-type tyrosine kinases as well as by secondary messengers, calcium, cAMP, and diacylglycerol.
Gene targeting of both Ras activator, RasGRF (Brambilla et al., 1997), and Ras inactivator, NF1 (Silva et al., 1997), genes results in the impairment of memory consolidation. Because Ras recycles between active GTP-bound and inactive GDP-bound states, the inactivation of either an activator or an inactivator shifts the balance of the two states. Therefore, a dynamic balance between the two states may be important for Ras to function. Manabe et al. (2000) demonstrated that the inactivation of one of the Ras genes, Ha-ras gene, results in the enhanced LTP in hippocampal CA1 cells because of selective enhancement of NMDA synaptic responses, possibly by the increased tyrosine phosphorylation of NR2A and NR2B subunits. We inactivated Ras by an adenovirus gene delivery system instead of a gene-targeting strategy. This allows us a shorter inactivation period of Ras function as compared with gene targeting, minimizing possible side effects in the developmental stage. We currently try to introduce adenoviruses into living animals to perform electrophysiological experiments using hippocampal slices.
We inhibited Ras activity by expressing a GTPase-activating protein for Ras, Gap1m. Biochemical analyses showed that Gap1m stimulates GTPase activity of Ras and R-Ras but not that of Rap1 (Li et al., 1997). However, the effect of R-Ras inhibition on the MAP kinase activity in our system may not be significant, because R-Ras does not activate (Marte et al., 1997) or weakly activates (Kimmelman et al., 1997) MAP kinase. The expression of full-length Gap1m might cause some side effects other than the inactivation of Ras, because Gap1m has, besides a GTPase-activating domain for Ras, phospholipid-binding and plekstrin homology regions (Maekawa et al., 1994), and the observed effects could be attributed to these regions. However, this possibility seems unlikely, because the inhibition of MAP kinase was also observed by the expression of dominant negative Ras that inhibits Ras by a different mechanism (Feig, 1999).
In this study we have shown that MAP kinase activation by NMDA stimulation requires Ras activity. However, Ras function is not restricted solely to the activation of MAP kinase. Ras has at least two other well characterized effectors, PI3-kinase and RalGDS, both of which are necessary for transforming activity of Ras (Vojtek and Der, 1998; Zwartkruis and Bos, 1999). PI3-kinase and RalGDS activate Rac and Ral small GTPases, respectively. Both GTPases function in the reorganization of cytoskeletal structures, which might be critical for neuronal plasticity. Therefore, it may be reasonable to assume that Ras has another important function in neuronal cells. Our system should provide a powerful tool to analyze such roles of Ras in neuronal signaling.
Footnotes
This work was supported in part by a Grant-in-Aid for Scientific Research from The Ministry of Education, Science, and Culture of Japan. N.I. is a recipient of the domestic research fellowship from Japan Science and Technology Corporation. We thank Drs. C. Nakayama, M. Matsuda, Y. Niino, and I. Saito for materials.
Correspondence should be addressed to Dr. Seisuke Hattori, Division of Biochemistry and Cellular Biology, National Institute of Neuroscience, National Center of Neurology and Psychiatry, 4-1-1 Ogawahigashi, Kodaira, Tokyo 187-8502, Japan. E-mail: hattori{at}ncnp.go.jp.
K. Namikawa's and H. Kiyama's present address: Department of Anatomy, Osaka City University, Graduate School of Medicine, 1-4-5 Asahimachi, Abenoku, Osaka 545-8585, Japan.
H. Ueno's present address: Department of Biochemistry and Molecular Pathophysiology, University of Occupational and Environmental Health, School of Medicine, Kitakyusyu, 807-8555, Japan.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}