

Abstract
Calcium/calmodulin-dependent protein kinase II (CaMK II) is found throughout the CNS. It regulates calcium signaling in synaptic transmission by phosphorylating various proteins, including neuronal membrane receptors and intracellular transcription factors. Inflammation or injuries to peripheral tissues cause long-lasting increases in the responses of central nociceptive neurons to innocuous and noxious stimuli. This change can occur independently of alterations in the responsiveness of primary afferent neurons and has been termed central sensitization. Central sensitization is a form of activity-dependent plasticity and results from interactions in a set of intracellular signaling pathways, which modulate nociceptive transmission. Here we demonstrate an increased expression and phosphorylation of CaMK II in rat spinal dorsal horn neurons after noxious stimulation by intradermal injection of capsaicin. Local administration of a CaMK II inhibitor in the spinal cord significantly inhibits the enhancement of responses of spinal nociceptive neurons and changes in exploratory behavior evoked by capsaicin injection. In addition, spinal CaMK II activity enhances phosphorylation of AMPA receptor GluR1 subunits during central sensitization produced by capsaicin injection. This study reveals that CaMK II contributes to central sensitization in a manner similar to its role in the processes underlying long-term potentiation.
- CaMK II
- central sensitization
- long-term potentiation
- protein phosphorylation
- intradermal capsaicin injection
- spinal cord
Dorsal horn neurons undergo anatomical and functional plastic changes after strong noxious stimulation (Willis, 2001a,b). Central sensitization is an enhanced responsiveness of central nociceptive neurons to innocuous and noxious stimuli. Central sensitization caused by capsaicin injection involves the activation of AMPA receptors, NMDA receptors, metabotropic glutamate receptors, and peptide receptors (Willis, 2001a,b). As a consequence of these synaptic events, a number of signal transduction pathways, involving such protein kinases as protein kinase A (PKA), protein kinase C (PKC), and protein kinase G (PKG), are activated (Dubner and Ruda, 1992; Lin et al., 1996, 1997; Malmberg et al., 1997), as are several transcription factors, including c-fos and the cAMP response element-binding protein (CREB) (Ji and Rupp, 1997; Wu et al., 2000, 2002). Among the membrane proteins phosphorylated by the activated protein kinases are NR1 subunits of NMDA receptors (Zou et al., 2000). Phosphorylation of NMDA receptors helps explain the enhanced responsiveness of NMDA receptors during central sensitization (Zou et al., 2000; Willis, 2001b).
Central sensitization resembles long-term potentiation (LTP) in the hippocampus (Willis, 1997, 2001b). For example, LTP initiation often involves excitation of NMDA and metabotropic glutamate receptors and activation of several protein kinases (Malenka and Nicoll, 1999;Soderling and Derkach, 2000). Enhanced excitatory transmission during LTP reflects a greater responsiveness of postsynaptic AMPA receptors, caused by either increased sensitivity of individual receptors or insertion of more AMPA receptors in the postsynaptic membrane (Fukunaga et al., 1996; McEachern and Shaw, 1996; Malenka and Nicoll, 1999;Soderling and Derkach, 2000).
A key initiating event in LTP is activation of calcium–calmodulin protein kinase II (CaMK II) (Fukunaga et al., 1996; Malenka and Nicoll, 1999; Kennedy, 2000; Soderling and Derkach, 2000). An increase in intracellular Ca2+ concentration, partly by influx through NMDA receptor channels, activates calmodulin, which in turn triggers the activation of CaMK II, causing it to bind to the postsynaptic density. Inhibitors of this kinase or genetic deletion of CaMK II subunits prevent LTP, and phosphorylation of CaMK II at Thr-286 allows it to remain active even after the Ca2+ signal returns to baseline (Malenka and Nicoll, 1999; Soderling and Derkach, 2000). Phosphorylation of GluR1 subunits of AMPA receptors enhances AMPA receptor mediated single-channel conductance (Li et al., 1999; Broutman and Baudry, 2001) and promotes the insertion of AMPA receptors into the postsynaptic membrane (Barria et al., 1997; Mammen et al., 1997;Soderling and Derkach, 2000). In addition, CaMK II stimulates CREB-dependent transcription by phosphorylation of this and other transcription factors (Curtis and Finkbeiner, 1999; Soderling and Derkach, 2000; Wu et al., 2002).
No previous evidence has linked the activation of CaMK II to central sensitization, although CaMK II is expressed in the spinal cord (Fukunaga et al., 1996; Li et al., 1999). Given the key role of CaMK II in LTP and our hypothesis that central sensitization is a spinal cord form of LTP, we decided to examine the role of CaMK II in the central sensitization produced by intradermal injection of capsaicin, using Western blots, immunostaining, electrophysiological recordings, and behavioral experiments.
Preliminary findings have been reported (Fang et al., 2001; Wu et al., 2001b).
MATERIALS AND METHODS
Experimental animals and induction of pain. Male Sprague Dawley rats (270–340 gm) were used. Capsaicin (3%, 50 μl) was injected into the plantar surface of the foot on the left side in anesthetized rats (Sluka et al., 1997; Wu et al., 2001a). The experiments were approved by the Institutional Animal Care and Use Committee and were consistent with the ethical guidelines of the National Institutes of Health.
Intrathecal and microdialysis fiber drug administration.After they were anesthetized with sodium pentobarbital (50 mg/kg), rats were implanted with an intrathecal catheter (32 gauge; Micor, Allison Park, PA) placed in the spinal subarachnoid space at the T12–L1 vertebral level for intrathecal administration of drugs. The rats were placed in a stereotaxic device, and the suboccipital region was exposed via a midline incision. The dura over the cisterna magna was incised vertically. The catheter was gently advanced ∼7.3–7.5 cm into the spinal subarachnoid space. The tip of a catheter was placed at the T12–L1 vertebral level, which was estimated to be the L4/5 level of the spinal cord. All animals were returned to the Animal Care Facility for 5 d to allow them to recover from the surgical procedure. After recovery, the end of the intrathecally inserted catheter was connected to an osmotic pump (Alzet; Alza Scientific Products, Palo Alto, CA) for the exploratory behavior study or to an infusion pump for the Western blot study. Artificial CSF (ACSF), KN-92, or KN-93 (Alexis, San Diego, CA) were infused. For electrophysiological experiments, a microdialysis fiber was used to deliver ACSF or the CAMK II inhibitor KN-93. Cuprophan hollow fibers (150 μm inner diameter, 9-μm-thick wall, 9 kDa molecular cutoff; Spectrum Medical Inc., Great Falls, MT) were used. Two 1 mm marks separated by a 3 mm space were drawn on a 30-cm-length Cuprophan fiber. A polyethylene sleeve (PE50 tubing) was placed around the fiber on each side of the marks. A few drops of silicon rubber were applied on each side of the marks, but not over the middle 3 mm zone used for dialysis, and the sleeve was then moved back and forth to seal the membrane uniformly, including the two 1 mm marks. A thin layer of silicon rubber (3140 RTN coating; Dow Corning, Midland, MI) was applied along the remaining length of the fiber. The fiber was left to dry in the air at room temperature overnight. Any fiber that exceeded 250 μm in diameter was discarded. The dull end of a stainless steel dissecting pin (0.1 mm diameter, 0.5 mm in length) was inserted into the lumen at one end of the dialysis fiber and glued in place with cyanoacrylate. The tip of the dissecting pin attached to the dialysis fiber was grasped in forceps and inserted into the spinal cord at a point approximately one-third the distance between the dorsolateral sulcus and the dentate ligament and out through the other side at the same level. The 3 mm collection zone was placed across the dorsal horns bilaterally. The fiber end and pin were cut off, and the fiber was glued to the end of a premeasured PE 20 tubing that was connected to a 5 ml syringe. The syringe was mounted on a Harvard infusion pump that maintained a constant infusion rate of 5 μl/min.
Western immunoblotting. Spinal cord tissue (L3–L6) was collected. In some experiments, the dorsal quadrants were separated from the ventral quadrants on each side so that the dorsal cord on each side could be examined independently of the ventral cord. The tissue was removed and put into liquid nitrogen immediately, followed by homogenation in 50 mol/l Tris buffer, pH 7.4 (0.1 mol/l EGTA, 0.14 μl/ml β-mercapto-ethanol, 100 mol/l PMSF, and 0.2 mg/ml trypsin inhibitor). The homogenate was centrifuged at 13,000 ×g for 15 min at 4°C. The supernatant for the cytosolic fraction was decanted from the pellet and used for all Western blot analyses. The concentration of protein in the homogenate was measured using the BCA kit (Pierce, Rockford, IL). Equal amounts of protein (60 μg) were size fractionated by SDS-PAGE and transferred onto a polyvinylidene difluoride membrane. The blots were incubated with primary monoclonal antibodies to CaMK IIα (Zymed, San Francisco, CA), phospho-CaMK IIα (Affinity BioReagents, Golden, CO), and polyclonal antibodies GluR1 and phospho-GluR1 (Ser-831 and Ser-845; Upstate Biotechnology, Lake Placid, NY). As an internal control, the expression of β-actin (Sigma, St. Louis, MO) was tested in all groups. The density of bands was quantified by AlphaEase software.
Immunohistochemistry and image analysis. Different groups of animals were deeply anesthetized with sodium pentobarbital (50 mg/kg, i.p) and perfused at 5, 15, or 30 min after stimulation. The perfusion fluid, pumped through the aorta after 200 ml of warm heparinized saline, was 4% paraformaldehyde at 4°C. The lumbar segments of the spinal cord were removed and placed in fresh fixative for 4 hr at 4°C. The spinal segments were left in 30% sucrose in phosphate buffer overnight and frozen sectioned on a sliding microtome at 30 μm. After blocking by 3% normal goat serum, sections were incubated in a solution of antibodies to CaMK IIα and phospho-CaMK IIα overnight. The immunoreactivity was detected by biotin-labeled secondary antibody on the second day. Sections from control and experimental animals were incubated simultaneously in the same antibody solution. Quantitative image analysis for relative optical density of CaMK IIα and phospho-CaMK IIα in the superficial laminas of the dorsal horn (laminas I-III) was performed usingBioquant software.
Electrophysiology. Rats used in electrophysiology experiments were anesthetized with sodium pentobarbital (50 mg/kg). Laminectomy was performed to expose the spinal lumbar enlargement for extracellular recording. Neuronal responses were recorded with a carbon filament electrode (3–4 MΩ). The effect of KN-93 was tested on the central sensitization of dorsal horn neurons after intradermal injection of capsaicin in two groups of animals. In one group, capsaicin was injected after pretreatment with KN-93 infused through the microdialysis fiber for 30–40 min (100 μm) while the injection was given without KN-93 pretreatment in the control group. The background activity and responses to mechanical stimuli (brush, press, pinch) were recorded before and after KN-93 infusion as well as 15 and 30 min after capsaicin injection. Recorded data were analyzed off-line from peristimulus time histograms processed by Spike 2 software. Responses to mechanical stimuli applied to the receptive field for 10 sec were calculated by subtracting the background activity to yield a net increase in discharge rate. The responses before capsaicin injection were normalized to baseline, which was set to 100%. The responses induced by capsaicin were expressed as a percentage of the baseline.
Exploratory behavior studies. Rats were individually placed into a computerized activity box that was equipped with monitors for detecting exploratory activity in terms of the frequency at which the rat interrupted one of six beams of ultraviolet light during a period of 45 min. A computer tracked the activity of the animals and integrated the data into a report that assessed the exploratory activity over time. For this study, the activity was summed over 45 min. The behavioral activity was described in terms of such parameters as entries, distance moved, and resting activity (Palecek et al., 2002). Capsaicin was injected intradermally in one foot 30 min before placing an animal in the activity box. Rats were briefly anesthetized with halothane during the injection. Data are presented as a percentage of values seen in pretreated animals. Student's t test was used to assess differences between vehicle and capsaicin injection groups.
RESULTS
Increased expression of CaMK II and phospho-CaMK II in spinal cord after capsaicin injection
By using the Western blot technique, we determined if the expression of CaMK II and phospho-CaMK II in the lumbar segments (L3–L6) of the spinal cord was increased at 5, 15, and 60 min after capsaicin injection (Fig.1a,c). CaMK II expression increased significantly by 15 min (Fig. 1a), and phosphorylated CaMK IIα increased significantly by 5 min (Fig.1c) after the intradermal capsaicin injection. No significant change was found at any time point in either the expression of CaMK IIα or phospho-CaMK IIα in animals given a vehicle injection. To localize further the expression of CaMK II in the spinal cord to the sensory part of the spinal cord, we separated and isolated spinal cord tissue dorsal to the central canal at 15 min after capsaicin injection and performed Western blots. We again found that at 15 min after capsaicin injection, the expression of both CaMK II (Fig.1b) (2.24 ± 0.18 vs 1.01 ± 0.03) and phospho-CaMK II protein (Fig. 1d) (2.87 ± 0.48 vs. 1.19 ± 0.14) was significantly increased in the dorsal part of the spinal cord ipsilateral to the injection site. However, no increase in CaMK II or phospho-CaMK II was detected on the contralateral side in capsaicin-injected rats when compared with vehicle-treated animals. As an internal control, we checked for a change in the expression of β-actin after capsaicin injection and found none (data not shown).

Expression of spinal cord CaMK II and phospho-CaMK II after capsaicin injection. Immunoblots of rat spinal cord tissue (L3–L6) with CaMK IIα (a) and phospho-CaMK IIα (c) antibody at 5, 15, and 60 min after capsaicin (solid bars) or vehicle injection (open bars). Bar graphs summarize the relative density of the bands [the densities of the bands from vehicle-treated rats at 5 min were normalized with respect to CaMK IIα (a) and phospho-CaMK IIα (c) after capsaicin injection (n = 4–6)]. Immunoblots of tissue containing the spinal dorsal horn, but not the ventral horn with CaMK IIα (b) and phospho-CaMK IIα (d) antibody at 15 min after capsaicin injection showed increases on the ipsilateral (Ips.) side (solid bar), compared with that from the contralateral (Con.) side (open bar) in the vehicle control (n = 4–6). All data are expressed as mean ± SEM and were analyzed by Student's t test; *p < 0.05. β-Actin levels were unchanged after capsaicin injection (data not shown).
Immunocytochemical localization of CaMK II and phospho-CaMK II expression in spinal cord
To localize and to confirm the increased expression of CaMK II and its phosphorylated form in the dorsal horn of the spinal cord in response to capsaicin injection, immunohistochemical studies were performed. The distributions of CaMK II and phospho-CaMK II after a vehicle injection are shown in Figure2a–c and h–j, respectively. After capsaicin injection, immunoreactivity for CaMK II (Fig. 2d–f) and phospho-CaMK II (Fig.2k–m) increased in the superficial dorsal horn on the ipsilateral side. CaMK II and its phosphorylated form were expressed both in terminals in the superficial layers of the dorsal horn and in cell bodies in deeper laminas. Retrogradely labeled spinothalamic tract (STT) cells were also found to express CaMK II and phospho-CaMK II (our unpublished observation). The time courses of the increases in immunohistochemical expression of CaMK II (Fig. 2g) and of phospho-CaMK II (Fig. 2n) were similar to those found in the Western blot studies (Fig. 1a,c), and image analysis confirmed that the changes were significant (Fig. 2g,n).

Immunostaining and image analysis of CaMK II and phospho-CaMK II in the dorsal horn after induction of central sensitization. Immunostaining for CaMK II (d–f) on the side of rat spinal cord ipsilateral to injection increased by 15 min (e) and at 60 min (f) after capsaicin injection. No obvious difference was found in capsaicin-injected animals (d) and vehicle-injected rats (a) at 5 min (b, 15 min; c, 60 min). Expression of phospho-CaMK II increased after capsaicin injection (k, 5 min; l, 15 min; m, 60 min) compared with the vehicle-injected rats (h–j). Image analysis showed that differences were significant (g, CaMK II; n, phospho-CaMK II; open bar, vehicle treatment; solid bar, capsaicin injection;n = 4; *p < 0.05). Scale bars, 100 μm.
Electrophysiological evidence of involvement of CaMK II in central sensitization
To test whether CaMK II activation is related to central sensitization after intense noxious stimulation, we made electrophysiological recordings of the background activity and responses of dorsal horn neurons to innocuous and noxious mechanical stimuli after capsaicin injection, with or without inhibition of CaMK II. A selective CaMK II inhibitor, KN-93, was administered through a microdialysis fiber that was implanted across the dorsal horn. In control animals without KN-93 pretreatment, the intradermal capsaicin injection caused a significant increase in the background activity and responses to innocuous and noxious stimulation (Fig.3a,c) (background: 442.4 ± 77.9%; brush: 293.0 ± 29.8%; press: 307.8 ± 61.2%; pinch: 271.9 ± 37.6%), which is consistent with previous data collected from both rats and primates (Willis 1993; Wu et al., 2001a).

Effect of CaMK II inhibitor KN-93 on the responses of spinal dorsal horn neurons to mechanical stimuli. a,Rate histograms represent changes in the responses to mechanical stimuli of a dorsal horn neuron after intradermal injection of capsaicin. Top row, Baseline activity and responses to brush, press, and pinch. Second row, Responses 15–30 min after capsaicin injection. b, Rate histograms represent changes in the responses to mechanical stimuli of a dorsal horn neuron after intradermal injection of capsaicin after pretreatment with KN-93 administered through a microdialysis fiber. Top row, Baseline background activity and responses to brush, press, pinch. Second row, Responses after infusion of KN-93 within the dorsal horn. Third row, Responses after capsaicin injection with KN-93 pretreatment. c, Bar graph showing the effects of KN-93 (100 μm) on the responses of dorsal horn neurons (n = 6) to mechanical stimuli after intradermal injection of capsaicin. *p < 0.05; **p < 0.01, compared with the percentage in rats without pretreatment by KN-93.Open bar, Vehicle treatment; solid bar, capsaicin injection.
We found that KN-93 (100 μm) significantly blocked the increases in background activity and in the responses of dorsal horn cells to innocuous and noxious stimulation caused by capsaicin injection (Fig. 3b,c) (background: 187 ± 48.5%; brush: 109 ± 11.3%; press: 122.7 ± 6.4%; pinch: 117.4 ± 8.8%). These data are consistent with the hypothesis that activation of CaMK II contributes to central sensitization.
Changes in pain behavior by CaMK II inhibitor
We have proposed that central sensitization underlies the secondary mechanical allodynia and hyperalgesia that follows intradermal capsaicin injection. To test the effect of CaMK II on pain-related behavior triggered by capsaicin injection, we used a computerized photobeam activity system to measure changes in exploratory behavior in rats (Palecek et al., 2002). This allowed us to determine the effect of inhibition of CaMK II activity by intrathecal administration of KN-93. As a control, we administered KN-92, the inactive isomer of KN-93. As previously reported, capsaicin injection causes a decrease in the entry number and traveled distance and an increase in the resting time in naive rats (Palecek et al., 2002). We confirmed that the injection of capsaicin significantly reduced the number of entries (Fig. 4a) (102.78 ± 13.4 vs 60.8 ± 11.27) and the distance traveled (Fig. 4b) (99.7. ± 7.9 vs 71.9 ± 7.4) and increased the resting time (data for resting time not shown here) after intrathecal treatment with KN-92. Data from rats treated with KN-92 were not significantly different from those from capsaicin-injected naı̈ve rats or ACSF-treated rats. This is consistent with our previous report (Palecek et al., 2002). However, administration of KN-93 through an intrathecal catheter prevented the reduction in exploratory behavior in response to capsaicin injection (Fig.4a) (97.29 ± 11.27 vs 103.31 ± 17.94) (Fig.4b) (116.0 ± 19.3 vs 117.9 ± 17.9). This change was significant. The results suggest that CaMK II contributes to the changes in exploratory behavior induced by capsaicin injection. This study provides evidence that CaMK II is an important step in the signal transduction pathways that cause central sensitization after noxious peripheral stimulation, as demonstrated at a behavioral level. Blocking CaMK II activity effectively prevents behavioral changes that we believe reflect the presence of hyperalgesia and allodynia.

Effect of CaMK II activity on changes in exploratory behavior in rats after capsaicin injection. Noxious stimulation by capsaicin injection significantly decreases the number of entries and distance traveled in KN-92-treated rats, but not in vehicle-injected rats. KN-93 prevents the decreases in entries (a) and traveled distance (b) in capsaicin-injected rats (n = 10–16; *p < 0.05; Student's t test). Data presented as a percentage of values seen in pretreated animals. Open bar, Vehicle injection; solid bar, capsaicin injection.
Activation of AMPA receptors by CaMK II
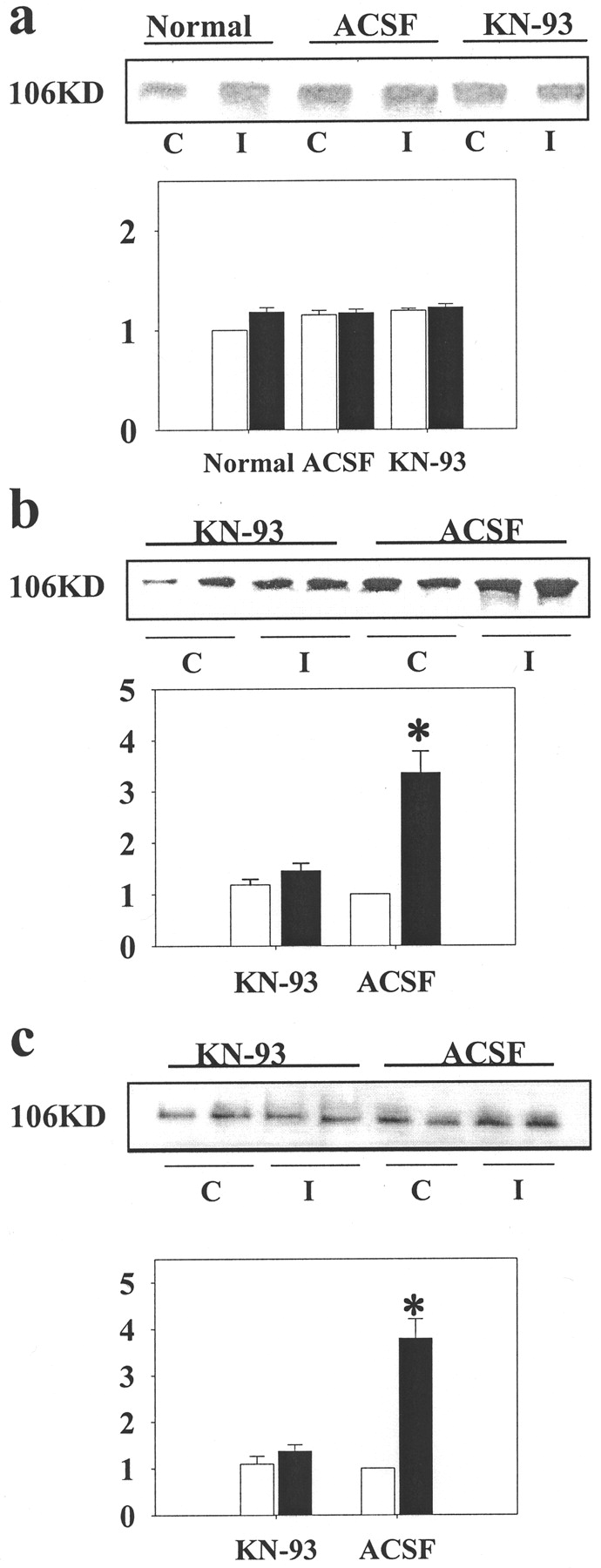
Central sensitization induced by injection of capsaicin activates a number of protein kinases and changes the sensitivity of ionotropic glutamate receptors (Dougherty and Willis, 1992; Malmberg et al., 1997; Zou et al., 2000; Willis, 2001a,b). These interactions have recently been discussed (Strack et al., 1997; Willis 1997, 2001a,b). To investigate further the influence of CaMK II and its activation on AMPA receptors, we used Western blots to test if AMPA receptor GluR1 subunits in spinal cord tissue are phosphorylated after intradermal capsaicin injection. We found that capsaicin injection significantly enhanced the phosphorylation of GluR1 subunits of AMPA receptors at both the Serine-831 (Fig. 5b) (compared to the ACSF treated control group) and Serine-845 (Fig.5c) (compared with the ACSF-treated control group) sites on the side ipsilateral to the injection. However, no significant increase was found on the contralateral side (Fig. 5b,c), and GluR1 protein itself did not show any change in response to the noxious stimulation (Fig. 5a). This means that any enhanced responsiveness of AMPA receptors after capsaicin injection would have been caused by enhanced protein phosphorylation rather than by increased production of the protein. However, this does not rule out an increased insertion of AMPA receptors into the postsynaptic membrane (Strack et al., 1997; Li et al., 1999; Malenka and Nicoll, 1999;Soderling and Derkach, 2000). The increased phosphorylation was significantly blocked at both Serine-831 (Fig. 5b) (1.36 ± 0.14 vs 3.79 ± 0.42) and Serine-845 (Fig.5c) (1.75 ± 0.16 vs 3.36 ± 0.44) sites in tissue from the side ipsilateral to the capsaicin injection when the selective CaMK II inhibitor, KN-93, was administered intrathecally. These data clearly demonstrate that enhanced phosphorylation of AMPA receptors, specifically of GluR1 subunits, in the spinal cord dorsal horn is triggered by intense peripheral noxious stimulation induced by capsaicin injection and that CaMK II directly regulates the phosphorylation state of AMPA receptors during central sensitization.

Effect of CaMK II activity on the phosphorylation of AMPA receptor GluR1 subunit in spinal cord tissue during central sensitization. Spinal GluR1 protein expression (a) did not show any changes on either the ipsilateral (solid bar) or contralateral (open bar) sides at 30 min after capsaicin injection. There was an increase in phosphorylation of AMPA receptor GluR1 subunits at Serine-831 (b) and Serine-845 (c) on the ipsilateral side in the lumbar segments of the spinal cord at 30 min after capsaicin injection in the hindpaw, compared with the contralateral side (c) (n = 4–6). However, KN-93 significantly blocked the increased GluR1 receptor phosphorylation both at Serine-831 (b) and Serine-845 (c) sites on the ipsilateral side (I) of lumbar spinal cord at 30 min after capsaicin injection (n = 4–6). *p < 0.05, relative to the value of that from contralateral side of ACSF-treated animals. β-Actin showed no significant change on either side after injection of capsaicin or vehicle (data not shown).
DISCUSSION
Multiple approaches in this study demonstrated the involvement of CaMK II in the central sensitization of nociceptive dorsal horn neurons that follows an intradermal injection of capsaicin. We found that there is both an increase in the amount of this protein and an increase in its phosphorylation after capsaicin injection. Administration of the CaMK II inhibitor, KN-93, blocked the increase in electrophysiological responses of nociceptive dorsal horn neurons expected after capsaicin, showing that these are dependent on CaMK II activity. KN-93 also prevented the changes in exploratory behavior that result from capsaicin injection, providing evidence that CaMK II contributes to the behavioral changes associated with central sensitization. CaMK II was also found to mediate the phosphorylation of GluR1 receptors, which may contribute to the enhanced responses to mechanical stimulation of the skin observed during central sensitization. These results suggest that CaMK II is importantly involved in the intracellular signal transduction pathways underlying the central sensitization induced by intradermal capsaicin injection.
Central sensitization
The animal model that we used in this study is intradermal injection of capsaicin into the rat paw. Intradermal injection of capsaicin in humans causes pain, primary mechanical and heat hyperalgesia, and secondary mechanical hyperalgesia and allodynia (LaMotte et al., 1991, 1992). Similar injections result in central sensitization in nonhuman primates (Simone et al., 1991; Dougherty and Willis, 1992; Sluka et al., 1997; Willis, 2001a,b). Central sensitization is a term used to describe the increased responsiveness of nociceptive neurons in the spinal cord after intense peripheral stimulation (Kenshalo et al., 1982). Central sensitization, which is reflected in the sensitization of nociceptive dorsal horn neurons, including primate STT neurons, is a reasonable explanation for secondary hyperalgesia and allodynia (Hardy et al., 1952; Simone et al., 1991; Willis 2001a,b; Wu et al., 2001a). After intradermal administration of capsaicin in rats, central sensitization also occurs in unidentified nociceptive neurons of the dorsal horn (Wu et al., 2001a), and there are changes in behavioral responses that have been interpreted to reflect secondary mechanical allodynia and hyperalgesia (Sluka and Willis, 1997; Palecek et al., 2002).
Our laboratory has provided evidence that central sensitization in the spinal cord depends on activation of glutamate receptors, several protein kinases, as well as other enzymes and transcription factors, such as nitric oxide synthase, c-fos, and CREB (Lin et al., 1996, 1997; Wu et al., 2000, 2001a, 2002).
Studies in humans, as well as in rats, have shown that the peak of the secondary hyperalgesia and allodynia occurs between 15 and 30 min after the injection of capsaicin (Hardy et al., 1952; Sluka et al., 1997;Willis, 2001a,b). The present investigation showed that CaMK II phosphorylation increases by 5 min after capsaicin injection and that CaMK II itself is increased by 15 min. The increase in the phosphorylation of GluR1 by CaMK II after capsaicin happens no later than 30 min after injection. The time course of these changes thus coincides with the time course of the onset of secondary hyperalgesia and allodynia. Therefore, the activation of CaMK II may initiate secondary hyperalgesia and allodynia.
Long-term potentiation in the spinal cord
The observation that CaMK II and its phosphorylation are important components of the mechanism of the central sensitization that follows noxious peripheral stimulation is consistent with our hypothesis that central sensitization is a spinal cord form of LTP.
LTP was first described by Bliss and Lømo (1973) in the hippocampus. Numerous studies have since been done on the mechanisms of LTP in the hippocampus (O'Dell and Kandel, 1994; Huang et al., 1996; Bach et al., 1999; Zhuo et al., 1999; Kasahara et al., 2001). However, there have also been studies showing that an LTP-like phenomenon can be demonstrated in dorsal horn neurons of the spinal cord (Randic et al., 1993; Liu and Sandkuhler, 1995, 1997, 1998; Lozier and Kendig, 1995;Liu and Sandkuhler, 1995, 1997, 1998; Svendsen et al., 1997; Rygh et al., 1999).
CaMK II is a very important component of the mechanism underlying LTP in the hippocampus. It has been shown that after a transient Ca2+ influx through NMDA receptors, phosphorylation of CaMK II and its persistent activation occur in LTP (Fukunaga et al., 1996). CaMK II becomes highly concentrated in the postsynaptic density (PSD). The PSD is a subcellular structure that contains abundance of proteins, including receptors associated with signal transduction pathways (Cotman et al., 1974; Cohen et al., 1977). CaMK II helps control the function of postsynaptic membrane receptors and the signal transduction events that follow receptor activation (Siekevitz, 1985; Miller and Kennedy, 1985, 1986; Kennedy, 1993). CaMK II makes up 1% of the total PSD protein, and it interacts with other important signal molecules after induction of LTP (Kennedy et al., 1983; Husi et al., 2000; Walikonis et al., 2000).
The AMPA receptor is one of the crucial proteins that interact with CaMK II in LTP (McGlade-McCulloh et al., 1993; Lisman and Zhabotinsky, 2001). GluR1 is one of the four subunits of AMPA receptors, is expressed in the hippocampus (Squire, 1992), and is crucial to LTP (Zamanillo et al., 1999; Lee at al., 2000; Broutman and Baudry, 2001;Liao et al., 2001). Activity of GluR1 is regulated by protein phosphorylation at its Serine-831 and Serine-845 sites (Carvalho et al., 2000; Soderling and Derkach, 2000; Vinade and Dosemeci, 2000). CaMK II mediates GluR1 phosphorylation in the hippocampal region in both LTP and LTD (Barria et al., 1997; Mammen et al., 1997; Kameyama et al., 1998; Lee et al., 1998, 2000, Wang and Kelly, 2001). It helps anchor the AMPA receptors onto the PSD and enhances AMPA-mediated synaptic transmission by increasing the single-channel conductance of existing AMPA receptors (Derkach et al., 1999; Lisman and Zhabotinsky, 2001).
Similarities between central sensitization and LTP
There are a number of similarities between LTP and central sensitization (Willis, 1997, 2001a,b). Both can involve NMDA, non-NMDA, and metabotropic glutamate receptors, increased influx of Ca2+, and activation of several second messenger pathways, such as the PKA, PKC, and NO/PKG pathways (Malmberg et al., 1997; Zou et al., 2000; Willis, 2001a,b). The sensitivity of excitatory amino acid receptors increases and that of the inhibitory amino acid receptors decreases in both processes (Willis 2001a,b). CaMK II is responsible for the initiation of hippocampal LTP (Malenka and Nicoll, 1999) after activation of calmodulin by increased intracellular Ca2+ concentration (Reymann et al., 1988;Malenka and Nicoll, 1999).
Based on our current study, it is evident that CaMK II is also crucial for the development of central sensitization of nociceptive dorsal horn neurons. Central sensitization, like LTP, not only involves the activation of other signal molecules, but also the induction of gene expression through protein phosphorylation, thereby strengthening cellular responses to peripheral stimulation. Phosphorylation of GluR1 subunits of AMPA receptors and phosphorylation of transcription factors, like CREB, presumably to alter gene expression in late stages of long-lasting plastic changes, are observed both in LTP and in central sensitization (Soderling, 2000; Wu et al., 2002).
The time courses of the early stages of LTP and of central sensitization are also similar. The phosphorylation of CaMK II in LTP studied in an in vitro preparation starts at 5 min (Soderling and Derkach, 2000), whereas capsaicin-induced CaMK II phosphorylation follows a similar time course. The observations from studies of LTP and from our study of central sensitization have demonstrated an increase in CaMK II expression within 15 min after stimulation (Ouyang et al., 1999; Kennedy, 2000). In the brain, CaMK II-mediated AMPA receptor phosphorylation needs to last for ∼30 min to have a maximal effect on LTP (Soderling and Derkach, 2000). In the present study, the increase in phospho-CaMK II lasts for at least 60 min. Inhibition of CaMK II activity blocks the phosphorylation of GluR1 subunits in both in LTP and capsaicin-induced central sensitization.
Whether GluR1 phosphorylation is involved in central sensitization and whether CaMK II mediates this process in neurons in the spinal cord has not been clear. The fact that CaMK II mediates phosphorylation of GluR1 in the spinal cord after peripheral noxious stimulation provides additional evidence that the process of central sensitization of spinal nociceptive neurons is similar to LTP.
In this study, we found that GluR1 is phosphorylated at both the Serine 831 and Serine 845 sites after intradermal capsaicin injection. Moreover, a selective CaMK II inhibitor, KN-93, partially blocked the increased phosphorylation at both sites. Phosphorylation of GluR1 at Serine-831 by CaMK II in central sensitization is consistent with the results of studies of LTP in the hippocampus. However, we do not know why the CaMK II inhibitor, KN-93, partially blocked the phosphorylation of GluR1 at the Serine-845 site, because this is a PKA phosphorylation site. KN-93 may have done this indirectly. For example, the Ca2+-calmodulin complex can stimulate adenylate cyclase, which produces more cAMP and PKA (Soderling and Derkach, 2000). There is other evidence that CaMK II induces phosphorylation of NMDA channels, which causes an enhancement in Ca2+ influx through the channel (Kitamura et al., 1993). Administration of KN-93 would prevent this and cause less Ca2+ influx through the NMDA channels, reducing the availability of the Ca2+-calmodulin complex. Less Ca2+-calmodulin complex in the cells after administration of the CaMK II inhibitor would presumably result in less adenylate cyclase and less phosphorylation of GluR1 at the Serine-845 site by PKA. Another possibility is that there may be cross-reactivity between antibodies to phospho-GluR1 at the Serine-831 and Serine-845 sites.
In conclusion, this study not only provides evidence that CaMK II is a key factor in central sensitization in the spinal cord, but it also strengthens our hypothesis that central sensitization is a spinal cord form of LTP. Central sensitization may be considered as a memory of a painful event by the spinal cord. The idea that central sensitization in the spinal cord resembles LTP can provide attractive possibilities for further research on central sensitization.
Footnotes
↵* L.F. and J.W. contributed equally to this work.
This work is supported by National Institutes of Health Grants NS 09743 and NS11255 and by the J. E. Kempner Scholarship Fund. We thank Griselda Gonzales for helping with the figures.
Correspondence should be addressed to Prof. William D. Willis, Department of Anatomy and Neurosciences, Marine Biomedical Institute, The University of Texas Medical Branch, Galveston, TX 77555-1069. E-mail: wdwillis{at}utmb.edu.
L. Fang's present address: Division of Neurosurgery, Department of Surgery, University of Texas Medical Branch, Galveston, TX 77555-0517.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}