

Abstract
Acute intrahippocampal infusion of brain-derived neurotrophic factor (BDNF) leads to long-term potentiation (BDNF-LTP) of synaptic transmission at medial perforant path→granule cell synapses in the rat dentate gyrus. Endogenous BDNF is implicated in the maintenance of high-frequency stimulation-induced LTP (HFS-LTP). However, the relationship between exogenous BDNF-LTP and HFS-LTP is unclear. First, we found that BDNF-LTP, like HFS-LTP, is associated with enhancement in both synaptic strength and granule cell excitability (EPSP–spike coupling). Second, treatment with a competitive NMDA receptor (NMDAR) antagonist blocked HFS-LTP but had no effect on the development or magnitude of BDNF-LTP. Thus, NMDAR activation is not required for the induction or expression of BDNF-LTP. Formation of stable, late phase HFS-LTP requires mRNA synthesis and is coupled to upregulation of the immediate early gene activity-regulated cytoskeleton-associated protein (Arc). Local infusion of the transcription inhibitor actinomycin D (ACD) 1 hr before or immediately before BDNF infusion inhibited BDNF-LTP and upregulation of Arc protein expression. ACD applied 2 hr after BDNF infusion had no effect, defining a critical time window of transcription-dependent synaptic strengthening. Finally, the functional role of BDNF-LTP was assessed in occlusion experiments with HFS-LTP. HFS-LTP was induced, and BDNF was infused at time points corresponding to early phase (1 hr) or late phase (4 hr) HFS-LTP. BDNF applied during the early phase led to normal BDNF-LTP. In contrast, BDNF-LTP was completely occluded during the late phase. The results strongly support a role for BDNF in triggering transcription-dependent, late phase LTP in the intact adult brain.
- long-term potentiation (LTP)
- synaptic plasticity
- neurotrophin
- brain-derived neurotrophic factor (BDNF)
- dentate gyrus
- hippocampus
- activity-regulated cytoskeleton-associated protein (Arc)
- gene expression
The neurotrophin family of secretory proteins play a diverse and broad role in regulating neuronal structure and function in the developing and adult nervous system (Thoenen, 1995;Bibel and Barde, 2000). Brain-derived neurotrophic factor (BDNF) is the most widely distributed neurotrophin in the adult brain. Although BDNF actions have been studied extensively in cell culture, the functions of BDNF in the adult brain have not been clearly defined. Growing evidence suggests that BDNF is important in activity-dependent synaptic plasticity, particularly in the context of long-term potentiation induced by high-frequency stimulation (HFS-LTP) (Schuman, 1999;Schinder and Poo, 2000; Binder et al., 2001; Bramham et al., 2002).
The role of endogenous BDNF in HFS-LTP has only been extensively studied at Schaffer collateral→CA1 synapses in the in vitro hippocampal slice preparation. Maintenance of HFS-LTP consists of at least two phases: an early, labile phase dependent on covalent modifications of existing proteins; and a late, stable phase requiring new mRNA and protein synthesis (Krug et al., 1984; Matthies et al., 1990; Bliss and Collingridge, 1993; Frey et al., 1996; Nguyen and Kandel, 1996). LTP maintenance is impaired by treatment with antibodies that inhibit activation of the BDNF receptor TrkB, or by deletion of the BDNF or TrkB genes (Figurov et al., 1996; Kang et al., 1997; Korte et al., 1998; Chen et al., 1999; Minichiello et al., 1999;Xu et al., 2000; Kossel et al., 2001; Patterson et al., 2001). Depending on the stimulation parameters used, relatively selective impairment in the ability to generate late, transcription-dependent LTP is seen (Kang et al., 1997; Korte et al., 1998; Minichiello et al., 1999; Patterson et al., 2001).
Exogenously applied BDNF has an impressive range of rapid, short-acting effects, including modulation of axon guidance, synaptic transmission, and membrane depolarization (Song and Poo, 1999; Schinder and Poo, 2000). In addition to these short-term effects, Kang and Schuman (1995,1996) and Kang et al. (1997) found that bath perfusion of hippocampal slices with BDNF induces a long-lasting enhancement of synaptic strength in the CA1 region. For reasons still unresolved, possibly related to the method of BDNF application (Kang et al., 1996), this finding has not yet been replicated (Patterson et al., 1996; Frerking et al., 1998; Schinder and Poo, 2000). However, when microinfused into the adult dentate gyrus in vivo, BDNF induces a lasting strengthening of transmission at perforant path→granule cell synapses (Messaoudi et al., 1998; Ying et al., 2002). We have termed this effect BDNF-induced LTP (BDNF-LTP).
The mechanism of BDNF action in synaptic plasticity is little understood. If BDNF participates in triggering late HFS-LTP, it should regulate new protein synthesis. New proteins could derive from translation of existing mRNA, from new transcription, or both. Kang and Schuman (1996) and Aakalu et al. (2001) have provided evidence that BDNF stimulates protein synthesis in dendrites from existing mRNA. However, the role of transcription in BDNF-LTP has not been examined. Moreover, the physiological relevance of exogenous BDNF-LTP and its relationship, if any, to mechanisms of HFS-LTP in the intact adult brain is unclear.
Here we provide evidence that (1) induction of BDNF-LTP, in contrast to HFS-LTP, does not require NMDA receptor (NMDAR) activation; (2) BDNF-LTP, like HFS-LTP, is associated with enhanced granule cell excitability [EPSP–spike (E–S) coupling] in addition to enhanced synaptic transmission; (3) BDNF-LTP, like late HFS-LTP, requires rapid new transcription coupled to upregulation of the immediate early gene activity-regulated cytoskeleton-associated protein (Arc); and (4) BDNF-LTP is occluded during late phase but not early phase HFS-LTP. Together these results suggest a functional role for BDNF as a trigger of transcription-dependent, late phase LTP in vivo.
MATERIALS AND METHODS
The electrophysiological and infusion methods used are the same as those described previously with minor modifications (Messaoudi et al., 1998; Ying et al., 2002).
Surgery and electrode and cannula placement. Eighty-four Mol: SD rats (Møllegaards Avls-Laboratorium) weighing between 250 and 350 gm were anesthetized with urethane (1.5 gm/kg, i.p.) and placed in a stereotaxic apparatus. Rectal temperature was maintained at 37°C by a servo-heating pad. Electrophysiological methods for obtaining selective stimulation of the medial perforant path have been detailed previously (Bramham et al., 1991). Stereotaxic coordinates relative to Bregma were 7.9 mm posterior, 4.2 mm lateral for stimulation, 3.9 mm posterior, and 2.2 mm lateral for recording. The arrangement of the cannula–electrode assembly is depicted in Figure 1A. An outer (guide) cannula (24 gauge; Plastics One, Roanoke, VA) was beveled sharp at the tip to facilitate tissue penetration. A Teflon-coated stainless steel wire-recording electrode (coated diameter, 112 μm) was glued to the cannula shaft and cut so the tip extended 1 mm from the end of the cannula. A bipolar stimulating electrode was lowered into the dorsomedial aspect of the angular bundle for stimulation of the medial perforant path. After a small slit was made in the dura, the guide cannula and attached recording electrode were slowly lowered into the dorsal hippocampus until a positive-going field EPSP (fEPSP) of maximum slope was obtained in the dentate hilus. The final depth of the recording electrode ranged between 200 and 300 μm below the level of the maximum negative-going fEPSP sink recorded in the middle third of the dentate molecular layer. An inner infusion cannula (31 gauge) was then inserted so that it protruded 300 μm below the end of the guide. The tip of the infusion cannula was located in the deep stratum lacunosum-moleculare of field CA1, 700 μm above the hilar recording site and 300–400 μm above the medial perforant synapses.
The infusion cannula was connected via PE50 polyethylene tubing to a 5 μl Hamilton (Reno, NV) syringe. Solutions were delivered by an infusion pump at a rate of 80 nl/min. Two micrograms of BDNF were delivered in a volume of 2 μl over 25 min. Dose–response analysis showed that this was the lowest dose giving maximal BDNF-LTP (Messaoudi et al., 1998).
Electrophysiology. Biphasic rectangular pulses of 150 μsec duration were applied every 30 sec throughout the experiment. The stimulation intensity for test pulses was set to elicit a population spike of one-third of the maximal amplitude. After a 15 min period of stable responses, an input–output curve was collected, followed by baseline recording (20 min), BDNF infusion (25 min), and postinfusion recording (2–7 hr). Input–output curves were constructed from responses (mean of four sweeps) obtained at eight stimulus intensities. The stimulus intensity was increased in 100 μA increments starting 100 μA below the population spike threshold. In some experiments, HFS-LTP was induced using a paradigm that evokes LTP lasting 1–5 d and associated with BDNF and TrkB mRNA upregulation (Bramham et al., 1996). HFS consisted of eight pulses at 400 Hz, repeated four times, at 10 sec intervals. Three session of HFS were given at intervals of 5 min. The stimulus intensity for HFS was the same as for test pulses.
Tissue microdissection and sample preparation. At the end of electrophysiological recording, rats were decapitated, and the brain was rapidly removed and rinsed with oxygenated ice-cold artificial CSF (in mm: NaCl 124.0, NaHOC3 25.0,d-glucose 10.0, KCl 3.4, KH2PO4 1.2, MgSO4 1.0, and CaCL2 2.5, pH 7.4). The dentate gyrus and hippocampal CA1 and CA3 regions were rapidly dissected on a glass dish kept on ice. Tissues were hand-homogenized with 15 strokes in 300 μl of freshly made SDS sample buffer containing 10% glycerol, 2.3% SDS, 0.01% bromophenol blue, and 0.5% β-mercaptoethanol in 62.5 mm Tris-HCl, pH 6.8 at room temperature. Homogenates were boiled for 5 min, aliquoted into Eppendorf tubes, and stored at –80°C until use.
SDS-PAGE and Arc immunoblotting. Protein levels in homogenate samples were determined using the Lowry method. Equal amounts of protein were loaded onto SDS-PAGE gels (10%) and run overnight at a constant current of 10 mA. Separated proteins were transferred to a nitrocellulose membrane using a constant voltage of 30 V. Membranes were blocked in 3% BSA on a gyro-rocker at 4°C overnight or for 1 hr at room temperature. Primary antibody recognizing the Arc N terminus (1:50; Santa Cruz Biotechnology, Santa Cruz, CA) was diluted in Tris-buffered saline containing 0.1% Tween 20 (TBST) and 3% BSA. Membranes were incubated in primary antibody for 2 hr at room temperature or at 4°C overnight with constant shaking, washed three times in TBST, and incubated for 1 hr with HRP-labeled anti-rabbit IgG (1:10,000 in TBST; Santa Cruz Biotechnology). Blots were washed in TBST and developed using enhanced chemiluminescence. Autoradiographs were scanned on a laser densitometer and quantitated using Phoretics 1D Plus software. Western blots were developed to be linear in the range used for densitometry. Optical density values obtained from the treated hippocampus were normalized relative to values in the nontreated hippocampus for each hippocampal subfield. Statistical analyses were based on paired and unpaired t tests for between- and within-group comparisons, respectively.
BDNF immunocytochemistry. BDNF immunocytochemistry was used to the study the distribution and clearance of exogenous BDNF during BDNF-LTP. Cytochrome c, which has a molecular weight and charge similar to those of BDNF, was used as a protein control. Cytochrome c has no effect on basal synaptic transmission or on several signal transduction pathways that have been studied (Messaoudi et al., 1998; Ying et al., 2002).
Animals were deeply anesthetized with an overdose of chloral hydrate and pentobarbital before killing. They were then transcardially exsanguinated with ∼50 ml of heparinized saline containing 1 mm sodium orthovanadate followed by fixation with 4% paraformaldehyde in first acetate, pH 6.5, and then borate, pH 9.5, buffers. All solutions were kept on wet ice throughout the perfusion procedure and were perfused at a rate of ∼1 l/hr. After perfusion fixation, brains were removed and placed in 30% sucrose in borate buffer at 4°C until they sank (∼5–7 d). Brains were frozen and sectioned coronally on a sliding microtome at 40 μm after sinking. Sections were stored at –20°C in cryoprotectant solution (Watson et al., 1986) until immunostained. Free-floating series of sections (1:12) were immunostained as described previously (Morse et al., 1993) for BDNF with the addition of a pretreatment in 0.1 mm sodium periodate in TBS to reduce endogenous peroxidase activity. Sections were blocked with goat serum and then placed in primary antibody solution (BDNF rabbit polyclonal; Amgen, Thousand Oaks, CA) overnight at 4°C. Staining with BDNF antibody at dilutions of >1:10,000 often reduces the endogenous staining, making it easier to see the distribution of exogenously administered protein. A titer of 1:20,000 was used for BDNF-infused brains, and 1:10,000 was used for cytochromec-infused brains. Sections were washed and incubated with biotinylated secondary antibody (goat anti-rabbit, 1:1500; Vector Laboratories, Burlingame, CA). After washing again, staining was completed using an avidin–biotin–peroxidase complex reaction (Vectastain Elite ABC kit; Vector Laboratories). The peroxidase was visualized using a diaminobenzidine chromagen and nickel sulfate intensification. The specificity of the antibody for BDNF relative to other neurotrophins has been documented previously (Morse et al., 1993).
Drugs. Human recombinant met-BDNF (a gift from Amgen-Regeneron Partners) was obtained as a concentrated stock solution in PBS (150 mm NaCl, 10 mm sodium phosphate buffer, pH 7.0, and 0.004% Tween 20). (RS)-3–2(2-Carboxypiperazin-4-yl)-propyl-1-phosphonic acid (CPP; Tocris Cookson Ltd., Bristol, UK) was dissolved in saline and injected intraperitoneally at a dose of 10 mg/kg 2 hr before the end of BDNF infusion. Actinomycin D (ACD) and recombinant cytochromec from yeast (Sigma, St. Louis, MO) were dissolved in PBS. All drugs were aliquoted in small volumes and stored at −80°C until use.
Response analysis and statistics. Signals from the dentate hilus were amplified, filtered (1 Hz to 3 kHz), digitized (25 kHz for field potentials), and stored to computer disk. Acquisition and analysis of field potentials were accomplished using DataWave Technologies (Longmont, CO) WorkBench software. The maximum slope of the fEPSP and the amplitude of the population spike measured from its negative-going apex to the tangent line joining the first two positive peaks were measured, and averages of four consecutive responses were obtained. ANOVA for repeated measures followed by a post hocScheffé test was used for statistical analysis of group effects, and a t test for dependent samples was used for analysis of individual effects (Statistica package; StatSoft Inc., Tulsa, OK). Statistics were based on values obtained during baseline and 2 hr after terminating infusion, unless otherwise indicated. p = 0.05 was chosen as the level of statistical significance.
RESULTS
BDNF tissue distribution and clearance
Exogenous BDNF diffuses relatively slowly in tissue, and the kinetics of BDNF tissue clearance is not well known (Biffo et al., 1995; Croll et al., 1998). If BDNF is retained in tissue for long periods, this might contribute to the increase in synaptic transmission seen in BDNF-LTP. We therefore sought to determine the spatial distribution and time course of BDNF clearance. Figure1A shows the cannula assembly with the infusion site located in stratum lacunosum-moleculare CA1, immediately above the hippocampal fissure, ∼300 μm above the medial perforant path. Rats were transcardially perfused with fixative solution at 15 min or 1, 3, 6, or 24 hr after BDNF (2 μg, 2 μl, 25 min) infusion, and coronal sections were immunostained for BDNF.

Tissue distribution and clearance of infused BDNF. BDNF (2 μg/2 μl, 25 min) was infused into stratum lacunosum-moleculare CA1, immediately above the hippocampal fissure, ∼300 μm above the medial perforant path→granule cell synapses. Cytochrome c (Cyt C) was infused as a protein control. Brains were obtained at 15 min or 1, 3, 6, or 24 hr after infusion, and coronal sections were immunostained for BDNF.A, Schematic depiction of cannula–electrode assembly in the hippocampus. B, C, BDNF immunostaining obtained at 15 min and 1 hr after BDNF infusion, respectively. BDNF was rapidly delivered and cleared from the dentate gyrus. D, Cyt C 15 min control. B–D were taken through the area of the cannula tract. The BDNF antibody titer for the BDNF-infused brains was 1:20,000 (minimizing endogenous staining and facilitating detection of exogenous protein). The antibody titer for Cyt C-infused brains was 1:10,000 (therefore darkerstaining of mossy fibers). Scale bar, 1 mm.
At 15 min, BDNF immunostaining was observed in the dentate gyrus in six of six experiments, indicating a rapid delivery of BDNF (Fig.1B). BDNF spread radially from the site of infusion and along the cannula tract in the CA1 region, with variable spread to CA3. Typically, the borders of the BDNF stain were sharply demarcated, indicating an abrupt arrest of BDNF diffusion or detection. At 1 hr, BDNF was cleared from the dentate gyrus in two of two experiments (Fig.1C). At 3 and 6 hr, no BDNF staining could be detected in the dentate gyrus in two of three brains at each time point; the remaining brain showed moderate staining in the dentate gyrus. No staining was observed 24 hr (n = 2) after BDNF infusion. Staining of endogenous BDNF, for instance, in mossy fibers, appeared to be identical in ipsilateral (infused) and contralateral hippocampus, although no attempt was made to quantify this relationship. BDNF-LTP lasts for at least 15 hr in anesthetized rats, and induction of the potentiation requires rapid activation of extracellular signal-regulated protein kinase (ERK) (Messaoudi et al., 1998; Ying et al., 2002). The rapid delivery (<15 min) and clearance (<1 hr) of BDNF are consistent with the electrophysiological and biochemical effects and demonstrate that BDNF-LTP does not involve the continuous presence of exogenous BDNF.
BDNF enhances both synaptic transmission and E–S coupling
HFS-LTP is associated with two changes: (1) an increase in synaptic strength and (2) an increased excitability of the postsynaptic neuron for a given excitatory input (Bliss and Lomo, 1973; Abraham et al., 1987; Lu et al., 2000). The latter phenomenon, termed E–S coupling, has not been examined in BDNF-LTP. BDNF infusion resulted in a rapid increase in the fEPSP slope and population spike amplitude (Fig. 2A;n = 6), corroborating earlier work (Messaoudi et al., 1998). Input–output curves were collected immediately before baseline recording and 2 hr after BDNF infusion (Fig. 2B). E–S plots derived from the input–output curves show a marked leftward shift, indicating an increase in granule cell excitability to medial perforant path input (Fig. 2C). Thus BDNF-LTP, like HFS-LTP, is associated with a parallel increase in fEPSP strength and granule cell excitability.

BDNF infusion elicits enhanced synaptic transmission and enhanced E–S coupling at medial perforant path→granule cell synapses. A, Time course plots showing BDNF-LTP of the evoked fEPSP and population spike (Pop Spike). BDNF was infused during the period indicated by the hatched bar. Values are group means ± SEM) expressed as percentage of baseline (n = 6).B, Representative input–output curves obtained during baseline and 2 hr after BDNF infusion. Values are means of four responses. C, E–S plot based on values shown inB. The leftward shift in the E–S curve indicates an increase in granule cell excitability to synaptic input. The regression coefficient was 0.95 in both plots.
Next we asked whether BDNF-LTP is associated with an increase in the excitability of presynaptic fibers. Although presynaptic fiber excitability is not affected during HFS-LTP, this has not been investigated in BDNF-LTP (Bliss and Lomo, 1973). BDNF regulation of presynaptic ion channel function is clearly a possibility, because BDNF has been shown to modulate a voltage-dependent sodium channel (Kafitz et al., 1999) and the potassium-channel Kir3 (Rogalski et al., 2000). We therefore determined the minimum stimulus current needed to consistently (in >90% of trials) evoke an fEPSP. There was no difference in the mean current threshold based on eight rats recorded before (40 ± 12 μA) and 2 hr after (46 ± 13 μA) BDNF infusion. We conclude that BDNF-LTP, like HFS-LTP, does not involve a change in presynaptic excitability.
BDNF-LTP is NMDA receptor-independent
HFS-LTP at medial perforant path→granule cell synapses requires NMDAR activation (Errington et al., 1987; Bramham et al., 1991). BDNF has been shown to facilitate presynaptic transmission in cultured hippocampal neurons (Lessmann et al., 1994) and to increase potassium-evoked glutamate release from synaptosomes prepared from adult dentate gyrus (Gooney and Lynch, 2001) and neocortex (Jovanovic et al., 2000). This raises the possibility that BDNF may induce LTP indirectly by acutely releasing glutamate and activating NMDARs. We therefore examined the induction and expression of BDNF-LTP under conditions of NMDAR blockade. The competitive NMDAR antagonist CPP was injected intraperitoneally at a dose of 10 mg/kg. Under these conditions, NMDAR activation and LTP induction are blocked for at least 8 hr (Abraham and Mason, 1988; Villarreal et al., 2002). As shown in Figure 3A, BDNF induced a robust potentiation of the fEPSP slope and population spike amplitude in the CPP-treated animals (p < 0.05;n = 6). The magnitude and time course of potentiation were not significantly different from those of controls receiving BDNF alone (Fig. 3D) (p < 0.05). In another series of experiments (n = 4), the effect of CPP on HFS- and BDNF-induced LTP was determined in the same animal. Although HFS-LTP was completed abolished, infusion of BDNF 20 min after HFS resulted in full BDNF-LTP (Fig. 3B,C). We conclude that NMDAR activation is not required for the induction or expression of BDNF-LTP.

BDNF-LTP does not require NMDAR activation.A, Group mean changes in the fEPSP slope and population spike (Pop Spike) amplitude. CPP was injected intraperitoneally (10 mg/kg) 2 hr before BDNF infusion (hatched bar; n = 6). B, Representative plot showing the effect of HFS (arrow) plus BDNF (bar) in the presence of CPP. NMDAR blockade abolished HFS-LTP, but had no effect on BDNF-LTP. C, Field potentials (average of four sweeps) obtained at the times indicated in B. Calibration: 3 mV, 2 msec. D, Mean fEPSP slope obtained 2 hr after BDNF infusion in the CPP-treated specimens and nontreated controls.
BDNF-LTP does not require low-frequency test stimulation
The protocol for BDNF-LTP includes delivery of low-frequency test stimuli (one per 30 sec) throughout the experiment. We therefore asked whether this test stimulation is required. Low-frequency stimulation may provide an activity-dependent signal (i.e., intracellular calcium rise) acting in concert with stimulation of TrkB receptors. To investigate this issue, test stimulation was omitted during the period of BDNF infusion and for either 2 hr (n = 5) or 6 hr (n = 4) thereafter, whereupon six test responses were collected. Significant increases in the fEPSP and population spike were seen at both time points, indicating that BDNF-LTP is not dependent on test stimulation (Fig. 4).

BDNF-LTP does not require low-frequency test stimulation. The normal paradigm for monitoring responses involves delivery of low-frequency test responses throughout the experiment at a rate of one per 30 sec. Here, test stimulation was omitted during BDNF infusion (hatched bar) and for 6 hr after infusion. At the end of this time, six responses were collected. Changes in the fEPSP and population spike (Pop Spike) are shown. Values are means ± SEM of four experiments expressed as percentage of baseline. Note that potentiation is seen in response to the first stimulus applied after infusion. The magnitude of the fEPSP increase (41.6 ± 4.1%) was not significantly different from that of controls (47.6 ± 5%; n = 5) receiving continuous test stimulation.
BDNF-LTP is transcription-dependent
The development of late HFS-LTP requires new gene transcription. A possible role for transcription in BDNF-LTP was investigated using local infusion of the RNA synthesis inhibitor ACD. Figure5 shows the effect of ACD application at various time points relative to BDNF infusion. Infusion of BDNF alone induced stable potentiation of the fEPSP and population spike (Fig. 5A; only the fEPSP is shown). Infusion of ACD (4 μg in 1 μl; n = 6) 1 hr before BDNF completely abolished the BDNF-LTP. ACD alone had no effect on baseline synaptic transmission in the interval before BDNF infusion (Fig. 5C) or in a separate group of animals receiving ACD alone and recorded for >7 hr (Fig. 5B) (n = 5).

BDNF-LTP requires rapid transcription. The RNA synthesis inhibitor ACD was applied at various time points relative to infusion of BDNF. A–E, Group time course plots. A, BDNF alone (n = 5).B, ACD alone (n = 5). ACD was given 1 hr before BDNF (C; n = 6), immediately before BDNF (D; n = 6), or 2 hr after BDNF (E; n = 5). The periods of ACD infusion (4 μg, 1 μl; black bar) and BDNF infusion (hatched bar) are indicated.F, Summary bar graph of fEPSP changes. All values are group means ± SEM expressed as percentage of baseline. Values for the bar graph were obtained 2 hr after BDNF infusion in the ACD pretreatment group and 4 hr after BDNF (or ACD alone) infusion in the other groups. *Significantly different from BDNF group. The residual potentiation in the ACD + BDNF group was significantly elevated above baseline. Pop Spike, Population spike.
However, it is possible that ACD blocks an ongoing transcriptional process (housekeeping function) that, although not critical for basal synaptic transmission, allows synapses to respond to BDNF. To obviate this issue, ACD was applied immediately before BDNF. A rapid potentiation was observed during the first hour after BDNF infusion, followed by a sharp decline to a stable plateau level (Fig.5D) (n = 6). In this paradigm, the initial response to BDNF is intact, but the sustained response is strongly inhibited. The mean increases of the fEPSP slope and population spike amplitude measured 2 hr after BDNF infusion were 12.6 and 17.27%, respectively. Although inhibited relative to control, the residual potentiation remained significantly elevated above baseline (p < 0.05). Finally, ACD was applied 2 hr after BDNF infusion during established BDNF-LTP. As shown in Figure5E, the increase in the fEPSP slope and population spike amplitude observed 2 hr after ACD infusion was not significantly different from those of time-matched controls receiving BDNF alone (n = 5) (p < 0.05). Together the results suggest that the induction but not the maintenance of BDNF-LTP requires new transcription. The effects are summarized in Figure 5F.
The immediately early gene Arc is required for generation of late HFS-LTP and long-term memory (Guzowski et al., 2000). We have shown recently that BDNF-LTP is accompanied by upregulation of ArcmRNA and protein in the dentate gyrus (Ying et al., 2002). Here we assessed the effect of ACD on Arc upregulation (Fig.6) (n = 5). Arc protein levels were measured by Western blot analysis of homogenates obtained from microdissected dentate gyrus and CA1 and CA3 regions. ACD blocked a threefold upregulation of Arc protein expression in dentate gyrus in parallel with blockade of BDNF-LTP. ACD had no effect on basal levels of Arc expression in CA1 and CA3. These results strengthen the correlation between Arc and BDNF-LTP and demonstrate that Arc protein derives predominantly from transcription of new Arc mRNA rather than from translation of preexisting transcripts.

ACD blocks Arc upregulation associated with BDNF-LTP. Western blot assays of Arc were run on aliquoted samples from microdissected dentate gyrus (DG) and hippocampal regions CA1 and CA3 after BDNF-LTP in vivo.A, Group mean + SEM changes in Arc immunoreactivity levels based on densitometric analysis. Optical density values are expressed as a ratio between the treated and nontreated (control) side for each region. BDNF-LTP is associated with enhanced Arc expression at 3 hr (n = 7) but not 15 min (n= 8; data not shown). ACD infusion 1 hr before BDNF blocked BDNF-LTP and the associated increase in Arc expression (n = 5). No changes in Arc expression were detected in the CA1 or CA3 regions. B, Representative immunoblot from the ACD-pretreated group. Infusions were made into the left hippocampus.
BDNF-LTP is occluded during late phase but not early phase HFS-LTP
A major outstanding issue is whether BDNF-LTP is physiologically relevant. Does exogenous BDNF reflect the actions of endogenous BDNF? If BDNF-LTP and HFS-LTP have a common mechanism, the expression of one should occlude expression of the other. To be able to draw conclusions from occlusion experiments, it is critical to establish saturation of the phenomenon on which the occlusion test is based. HFS-LTP is suitable for this purpose, because saturation is rapidly achieved and easily assessed, whereas BDNF-LTP is unsuitable in our view because of the slow time course of the potentiation.
HFS-LTP was induced using parameters (three sessions of 400 Hz stimulation) that generate LTP lasting days (Bramham et al., 1996). After 30 min of recording, the stimulus intensity was lowered to reset the fEPSP slope to baseline, keeping the response on the rising phase of the input–output curve. As shown in Figure7A (n = 6), a second round of HFS produced no further increase, demonstrating saturation of HFS-LTP. By contrast, infusion of BDNF-LTP at this time (1 hr after the first HFS) led to an increase in the fEPSP slope and population spike. The kinetics of the increase and the magnitude of the increase were not significantly different from those of the control (Fig. 7C). The lack of occlusion indicates that BDNF-LTP does not contribute to expression of early HFS-LTP.
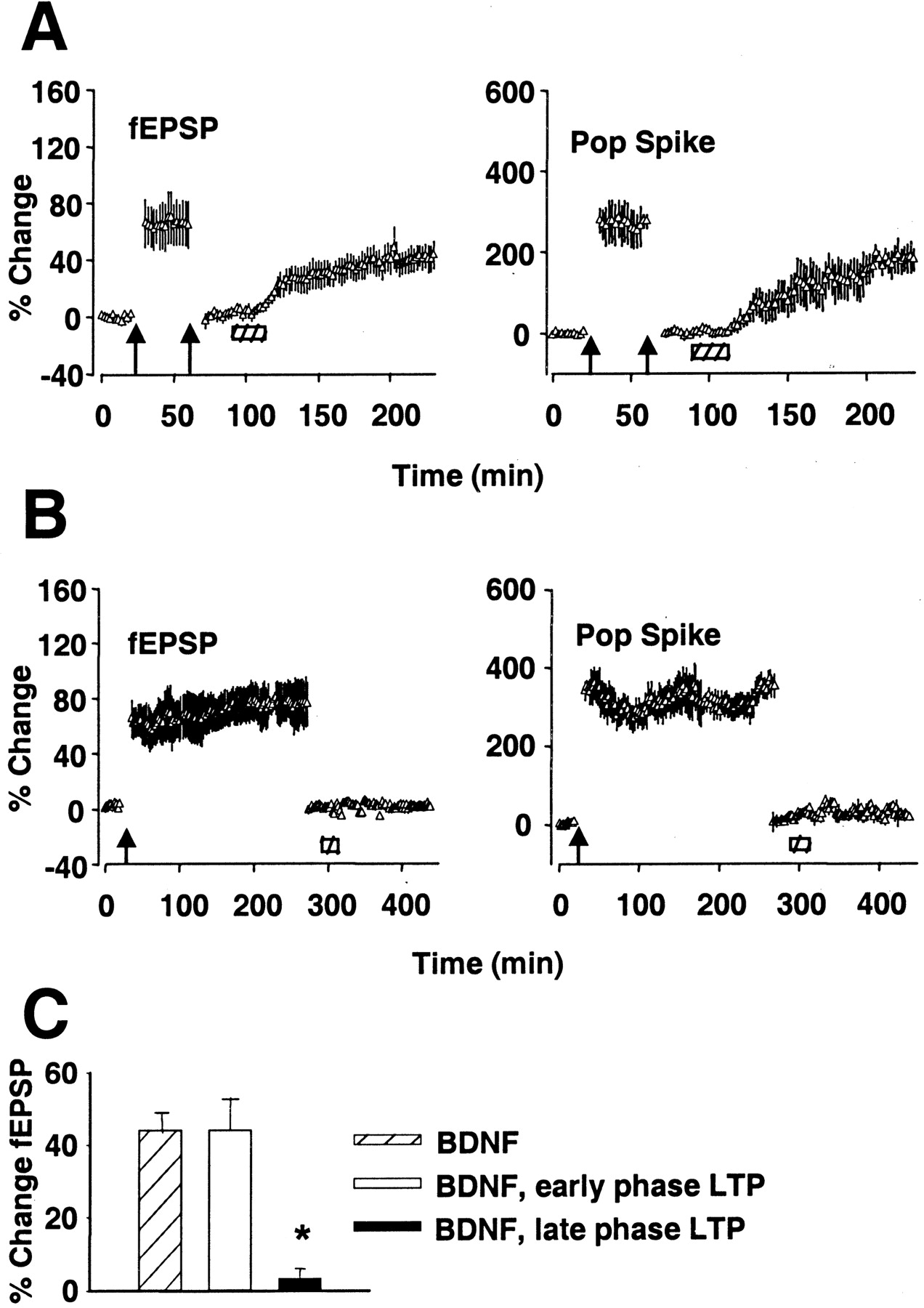
BDNF-LTP is occluded by late phase but not early phase HFS-LTP. A, LTP was induced by three sessions of HFS (400 Hz) and recorded for 30 min. The stimulus intensity was then lowered to reset the fEPSP slope to baseline. A second session of HFS produced no further increase, demonstrating saturation of HFS-LTP. BDNF infusion (hatched bar) 60 min after the first HFS led to increased synaptic transmission (n = 6).B, HFS-LTP was induced and recorded for 240 min and then reset to baseline as in A. BDNF infusion 260 min after HFS had no effect on fEPSP slope or population spike (Pop Spike) amplitude for the duration of recording (n = 6). Values are group means ± SEM expressed as percentage of baseline. C, Summary of fEPSP slope increases obtained in group receiving BDNF after baseline recording (BDNF), 60 min after HFS (BDNF, early phase LTP), or 260 min after HFS (BDNF, late phase LTP). *Significantly different from BDNF group. The magnitude of LTP in the BDNF group and BDNF, early phase LTP group was not statistically different (p < 0.05).
BDNF may nonetheless take part in late LTP based on a transcription-dependent mechanism. Occlusion experiments were performed 260 min after HFS, which by all accounts falls within the period of mRNA synthesis-dependent LTP. As shown in Figure 7B(n = 6), BDNF-LTP was completely occluded under these conditions. These results show that exogenous BDNF-LTP selectively occludes with late HFS-LTP.
DISCUSSION
The present in vivo study demonstrates that exogenous BDNF triggers a long-term increase in synaptic strength (BDNF-LTP), which requires rapid transcriptional activation and which occludes with late but not early HFS-LTP. Endogenous BDNF has multiple actions in the regulation of synapse function and plasticity (Schinder and Poo, 2000;Bramham et al., 2002). Endogenous BDNF has been shown to contribute to the generation of stable HFS-LTP (Kang et al., 1997; Korte et al., 1998; Chen et al., 1999; Minichiello et al., 1999; Xu et al., 2000;Patterson et al., 2001). Late HFS-LTP and long-term memory both depend on activation of ERK and induction of the immediate early gene Arc (Atkins et al., 1998; Impey et al., 1998; Davis et al., 2000; Guzowski et al., 2000; Rosenblum et al., 2002). We have shown recently that BDNF-LTP similarly requires ERK activation coupled to ERK-dependent activation of the nuclear transcription factor calcium/cAMP responsive-element binding protein and upregulation of Arc mRNA and protein (Ying et al., 2002). As seen with HFS-LTP (Lyford et al., 1995;Link et al., 1995; Valentine et al., 2000), Arc mRNA is selectively induced in dentate granule cells and delivered to dendritic processes. In the present work, local inhibition of transcription blocked BDNF-LTP and the attendant increase in Arc protein expression. By applying ACD at different time points relative to BDNF infusion, we have identified a window of critical transcription, which parallels the window of ERK activation. Taken together, these data strongly support a role for BDNF as a trigger for transcription-dependent synaptic strengthening. Signal-transducing TrkB receptors are located on axon terminals and in the postsynaptic density of glutamatergic synapses (Drake et al., 1999; Aoki et al., 2000). Without excluding presynaptic effects, these data clearly point to a role for postsynaptic modifications in dentate granule cells.
In these experiments, BDNF is infused immediately above the dentate gyrus, some 300 μm above the medial perforant path synapses. BDNF infusion at this site results in selective activation of signal transduction pathways in the dentate gyrus; no biochemical effects have so far been observed in microdissected CA1 or CA3 tissue (Kanhema et al., 2001; Ying et al., 2002). Using BDNF immunostaining, we show that exogenous BDNF is rapidly delivered and cleared from the dentate gyrus. BDNF is clearly being delivered to the CA1 region, and the lack of effect on signal transduction is interesting in this regard. It could be related to differences in activity-dependent gene expression between the CA1 region and dentate gyrus (French et al., 2001). However, there are salient methodological points to this issue. Because the cannula is placed into deep-field CA1 (stratum lacunosum-moleculare), differences can be expected with regard to the concentration and kinetics of BDNF as it travels upward along the cannula tract as well as possible trauma and pressure effects. Experiments combining recording of Schaffer collateral–CA1 responses with infusion of BDNF above the synaptic region are needed to resolve this issue.
The physiological relevance of exogenous BDNF application may be questioned. The fact that BDNF-LTP occludes selectively with late phase LTP and activates a common set of critical intracellular events in the intact adult brain attests to the physiological relevance of the phenomenon. The similarity between HFS-LTP and BDNF-LTP is further evidenced by the fact that both are associated with a parallel increase in synaptic strength and E–S coupling.
ACD has commonly been used to draw the distinction between transcription-dependent (late) and transcription-independent (early) LTP. Previous work has shown that ACD blocks development of late HFS-LTP, leaving early LTP intact (Frey et al., 1996; Nguyen and Kandel, 1996). In contrast, BDNF-LTP is almost completely transcription-dependent, consistent with a selective role for BDNF in late phase LTP. When ACD is applied immediately before BDNF, a normal initial response to BDNF is followed by a drop to a small but stable plateau level. There are two plausible explanations for the residual potentiation. First, it could reflect an mRNA synthesis-independent process, such as post-transcriptional regulation of protein synthesis or protein phosphorylation, and evidence for both actions exists (discussed below). Alternatively, the residual potentiation could reflect incomplete inhibition of transcription by ACD. For example, if ACD takes effect after BDNF, this would allow time for initiation of a small, transcription-dependent potentiation. We cannot rule out this scenario, although ACD was shown to act rapidly and potently (1 hr pretreatment blocked BDNF-LTP).
In the CA1 region of the hippocampus, BDNF-LTP appears to involve protein synthesis from dendritically localized mRNA (Kang and Schuman, 1996). Using slices in which the synaptic neuropil was isolated (connections with the CA3 and CA1 cell bodies were severed), Kang and Schuman (1996) showed that BDNF induces LTP that was blocked by protein synthesis inhibitors. However, a role for transcription has not been investigated in CA1. In this context, it should be pointed out that activation of the translation machinery in dendrites may go hand in hand with the arrival of Arc transcripts, effectively driving Arc protein synthesis at synaptic sites (Ying et al., 2002). Supporting this view, BDNF has been shown recently to stimulate post-transcriptional synthesis of Arc in isolated synaptoneurosomes (Yin et al., 2002). This mechanism may well be accentuated in the context of elevated Arc mRNA. The present work using the transcription inhibitor ACD shows that Arc protein expression in BDNF-LTP derives predominantly from new mRNA synthesis rather than from translation of preexisting mRNA. BDNF can modulate translation through multiple signaling cascades, and the contribution of these pathways to dendritic protein synthesis and synaptic plasticity are only beginning to be resolved (Steward and Schuman, 2001; Takei et al., 2001). BDNF-LTP and late HFS-LTP in the CA1 field are both blocked by inhibition of mammalian target of rapamycin, a key regulator of translation initiation (Tang et al., 2002). In the dentate gyrus, BDNF-LTP is coupled to ERK-dependent activation of elongation factor-2, an important regulator of peptide chain elongation (Kanhema et al., 2001).
The selective occlusion of the BDNF response during late LTP suggests a rapid transition (switch) in the mechanism of LTP expression taking place between 1 and 4 hr after HFS. Frey et al. (1995) reached a similar conclusion based on occlusion experiments using two sessions of HFS. They found that early LTP was occluded at 1 hr after HFS (as would be expected) but could again be induced at 4 hr. Our results are consistent with these findings and implicate BDNF as a trigger for the conversion between early and late LTP.
The mechanism of occlusion remains to be identified. Although Arc synthesis is required in late HFS-LTP, there is no evidence that it is sufficient. Work by Chowdhury et al. (1998) suggests that Arc acts as a physical tether for calcium/calmodulin-dependent protein kinase II (CaMKII), anchoring it to the cytoskeleton within or in close proximity to the postsynaptic density. Arc and CaMKII are both synthesized from dendritically localized mRNA (Lyford et al., 1995; Steward and Schuman, 2001), and recent work shows that BDNF-LTP is associated with CaMKII activation (Kanhema et al., 2001). The exact functional relationship between Arc and CaMKII will be important to define. Taken together, the data suggest that BDNF stimulates synaptic consolidation through transcription-dependent and -independent mechanisms in which Arc may play a key role.
The critical period of BDNF release and TrkB receptor activation in LTP remains to be defined. Hartmann et al. (2001) found that HFS triggers immediate release of BDNF–green fluorescent protein from postsynaptic sites in hippocampal cell cultures. Evidence for rapid release has also been obtained after LTP-inducing stimuli in hippocampal slices (Kossel et al., 2001). Other evidence suggests an important function for more sustained or delayed events. TrkB activation in the dentate gyrus is enhanced 40 min after HFS (Gooney and Lynch, 2001), and late LTP is impaired by application of a BDNF scavenger, TrkB-Fc, 30–60 min after HFS (Kang et al., 1997). A sustained or delayed activation of TrkB by endogenous BDNF is most consistent with our data, because BDNF-LTP was not occluded 1 hr after HFS.
BDNF has been shown to act presynaptically to enhance glutamate release, raising the possibility that BDNF might, by releasing glutamate, trigger classic NMDAR-dependent LTP (Jovanovic et al., 2000;Gooney and Lynch, 2001). This is not the case, however, because CPP abolished HFS-LTP but had no effect on BDNF-LTP. Our results confirm a previous study by Kang and Schuman (1995) in the CA1 region in vitro and extend it to the dentate gyrus in vivo. This result does not rule out a contribution of other glutamate receptor types in BDNF-LTP induction. The role of glutamatergic transmission in BDNF release is another issue. Release of endogenous BDNF during HFS depends on activation of postsynaptic ionotropic glutamate receptors (Hartmann et al., 2001). Using exogenous application, we are bypassing this initial release event.
In cultured hippocampal neurons, BDNF elicits a transient potentiation of excitatory synaptic transmission (Gottschalk et al., 1998; Lessmann and Heumann, 1998; Li et al., 1998; Crozier et al., 1999). This effect is inhibited by NMDAR antagonists and is mediated at least in part by enhanced conductance of postsynaptic NMDAR-2B-containing receptors (Levine et al., 1998; Crozier et al., 1999). Although the NMDAR-2B mechanism is important in the developing hippocampus, the fact that BDNF-LTP is undiminished during NMDAR blockade argues against this mechanism occurring in the adult dentate gyrus. On the other hand, there is evidence that NMDAR-2B is upregulated later (2 d) in the process of HFS-LTP (Williams et al., 1998).
BDNF is a versatile molecule acting in the short and long term to regulate a diverse range of functions in the developing and adult nervous systems. In the context of synaptic plasticity, the question has been asked of whether BDNF serves a housekeeping function, maintaining the machinery that makes synapses modifiable, or an active, instructive role in triggering synaptic change (Schinder and Poo, 2000). The present study supports a direct, instructive role for BDNF in transcription-dependent, late phase LTP. Furthermore, the phenomenon of exogenous BDNF-LTP provides a valuable tool for elucidating the molecular basis of BDNF action specific to the consolidation process.
Footnotes
This work was supported by the Norwegian Foundation for Public Health and European Union Biotechnology Program Grant BIO4-CT98-0333.
Correspondence should be addressed to Dr. Clive Bramham, Department of Physiology, University of Bergen, Årstadveien 19, N-5009 Bergen, Norway. E-mail: clive.bramham{at}fys.uib.no.
Dr. Ying's present address: Department of Anesthesiology, A-1050, Weill Medical College of Cornell University, 525 East 68th Street, New York, NY 10021.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}