

Abstract
Reelin, the protein defective in reeler mutant mice, plays a key role during brain development. Reelin is processed proteolytically at two sites, and the central fragment mimics function in vitro.
Here, we show that processing is functionally important in vivo, a question that could not be addressed in our previous study. New monoclonal antibodies directed against central Reelin block its binding to lipoprotein receptors and perturb cortical development in vitro, confirming the importance of the central fragment that is detected in tissue and body fluids. Processing occurs when Reelin is incubated with embryonic neurons in culture or with their supernatant, but inhibition of processing by a metalloproteinase blocker does not prevent Reelin signaling in neurons. Furthermore, neurons internalize similarly full-length or central Reelin. In contrast, inhibition of processing prevents signaling and perturbs cortical development in cultured embryonic brain slices. Moreover, in vivo, the concentration of central Reelin is dramatically and selectively increased in receptor-deficient tissue, suggesting its specific downregulation after binding to receptors and internalization. We propose that processing by end-migration neurons is required in tissue (where Reelin is likely anchored to the extracellular matrix) to release the central fragment that diffuses locally and signals to target cells, whereas, in vitro, all Reelin forms have indiscriminate access to cells, so that cleavage is not necessary for signaling.
Introduction
Reelin is a large extracellular protein that plays a pivotal role during brain development. Defective Reelin causes the reeler malformation in mice (D'Arcangelo et al., 1995; Tissir and Goffinet, 2003; Jossin, 2004) and lissencephaly in man (Hong et al., 2000). During cortical development, Reelin secreted by Cajal-Retzius cells in the marginal zone (MZ) acts on cells of the cortical plate (CP), by binding to two lipoprotein receptors, very low density lipoprotein receptor (VLDLR) and apolipoprotein-E receptor type 2 (ApoER2) (D'Arcangelo et al., 1999; Hiesberger et al., 1999; Trommsdorff et al., 1999). The signal is relayed by the Disabled-1 (Dab1) adaptor that docks to receptors (Howell et al., 1997; Sheldon et al., 1997; Ware et al., 1997; Bar et al., 2003) and is tyrosine phosphorylated by Src family kinases after Reelin binding (Arnaud et al., 2003a; Bock and Herz, 2003; Jossin et al., 2003; Kuo et al., 2005).
The Reelin sequence begins with a region with similarity to F-spondin, followed by eight repeats of 350–390 residues. Reelin is cleaved in vivo at two sites located C terminal to domains 2 and 6, respectively, generating a total of six predicted proteins (Lambert de Rouvroit et al., 1999; Ignatova et al., 2004). We and others showed that a protein composed of central repeats 3–6 recapitulates most functions of Reelin during cortical development (Jossin et al., 2004; Nogi et al., 2006). Thus far, however, central Reelin could not be detected directly in tissue, and the physiological significance of processing in vivo remains unclear. These are important issues, especially because results obtained with the function-blocking antibody CR50 point to the importance of the N-terminal region and studies of Reelin in human pathology have been performed using antibody 142 that probes the same region.
To further study the functional role of Reelin processing, we raised new antibodies against central Reelin repeats and showed that the predicted central fragment is indeed present in the embryonic brain and CSF. These antibodies perturbed the organization of the CP in cultured slices, by interfering with binding of Reelin to receptors and Dab1 phosphorylation, confirming the importance of the central region. Cultured cortical neurons secreted a metalloproteinase that cleaved exogenously added Reelin. However, blocking processing by a metalloproteinase inhibitor did not affect the ability of Reelin to stimulate Dab1 phosphorylation in neurons, in which full-length and central Reelin were internalized similarly. In contrast, inhibition of metalloproteinases in organotypic brain slices decreased Reelin cleavage and signaling and induced a reeler-like phenotype. Moreover, the concentration of central Reelin was specifically and dramatically increased in brains from double VLDLR and ApoER2 receptor-deficient mice, suggesting preferential downregulation by target cells in vivo.
These data suggest that cleavage does not influence function when target cells are directly accessible to Reelin but is nonetheless important in tissue. We hypothesize that, in vivo, Reelin is anchored to the extracellular matrix (ECM) and that a metalloproteinase secreted by migrating neurons releases central Reelin, which diffuses locally and binds to receptors on target cells.
Materials and Methods
Animals.
Animal procedures were performed in accordance with institutional guidelines and ratified by competent animal ethics committees. Normal CD1, RelnOrl (reeler), Dab1Scm (Scrambler), VLDLR, and ApoER2 mutant mice were bred in standard conditions. Embryonic development proceeds somewhat more rapidly in CD1 than in inbred mouse strains. Therefore, to obtain embryonic brains precisely at the preplate stage [embryonic day 13 (E13)], a CD1 colony was maintained on an inverted light cycle (lights “off” from 7:00 A.M. to 5:00 P.M.), and females were inspected for the presence of vaginal plugs between 5:00 and 7:00 P.M. Dab1−/− mutant brains were provided by J. Cooper (Fred Hutchinson Cancer Research Center, Seattle, WA).
Monoclonal antibodies.
For antigen preparation, a cDNA sequence corresponding to nucleotides 5668–6627 from mouse reelin (GenBank accession number GI6755311) was amplified with primers, 5′-GCGGATCCGTTCCTGTTGTTCCTCTTCC-3′ (forward, BamHI site underlined) and 5′-GCAAGCTTCAGCAGGATAAAATCCCTGG-3′ (reverse, EcoRI site underlined), and cloned in pTrcHisA (Invitrogen, San Diego, CA). The encoded polypeptide, named 4B5A, contains residues 1796–2115 of Reelin (GenBank accession number GI 6755312). It contains subrepeats 4B and 5A but excludes epidermal growth factor motifs that were found to hamper expression in bacteria. After induction by isopropyl β-d-1-thiogalactopyranoside, the 4B5A protein was enriched from bacterial extracts using an Ni-NTA affinity column (Qiagen, Chatsworth, CA) (Ausubel, 2001) and further purified to homogeneity by SDS-PAGE and electroelution using a Model 422 apparatus (Bio-Rad, Hercules, CA). Protein 4B5A was used to immunize homozygous RelnOrl mice, from which monoclonal antibodies were generated by splenocyte fusion (de Bergeyck et al., 1998). Hybridomas were screened by ELISA and characterized using standard protocols (Ausubel, 2001), and isotypes were defined using the Isostrip kit (Roche, Basel, Switzerland).
Western blots and immunohistochemistry.
Samples of human CSF were obtained at the Laboratory of Neurochemistry, Université Catholique de Louvain. Before analysis by SDS-PAGE, samples were concentrated 5- to 10-fold using centrifugal filters (50K NMWL; Millipore, Bedford, MA) (Ignatova et al., 2004). Brain homogenates were prepared in ice-cold PBS with ETDA (5 mm) supplemented with a protease inhibitor mixture (Complete; Roche). Western blots were performed with anti-Reelin G10 or 142, directed against the N-terminal region, or a mix of antibodies 12 and 14 (12+14) that recognize epitopes in the C-terminal region (de Bergeyck et al., 1998). Signal was detected using HRP-coupled goat anti-mouse antibodies (Dako Cytomation, High Wycombe, UK) and the Super Signal West Pico kit (Pierce, Rockford, IL). For immunohistochemistry, brains were fixed in Bouin's fluid, and paraffin sections were incubated with antibodies and processed using the StreptABComplex/HRP Duet kit (Dako Cytomation). Primary antibodies were anti-bromodeoxyuridine (BrdU; Becton Dickinson, Mountain View, CA), polyclonal rabbit anti-Tbr1 (gift from Dr. R. Hevner, University of Washington, Seattle, WA), and anti-chondroitin sulfate CS-56 (Sigma, St. Louis, MO). For immunofluorescence, cells were fixed in paraformaldehyde (4% in PBS), permeabilized with 0.3% saponin, and incubated with antibody R4B or anti-Reelin E4 (de Bergeyck et al., 1998), followed by fluorescein-labeled antibodies against mouse IgG (Sigma).
Recombinant Reelin proteins, receptor pulldown, and Dab1 phosphorylation assays.
The production of recombinant Reelin proteins was described previously (Jossin et al., 2004). The binding to lipoprotein receptors was assessed in pull-down assays with soluble ectodomains of human LDLR, human VLDLR, and mouse ApoER2, produced as soluble fusion proteins with the Fc fragment or His tags, as described previously (Hiesberger et al., 1999). The ability of Reelin to stimulate Dab1 tyrosine phosphorylation in primary neuronal cultures was assessed by immunoprecipitation of Dab1 with a polyclonal antibody, followed by Western blotting and revelation with the 4G10 antiphosphotyrosine antibody (Upstate Biotechnology, Lake Placid, NY) or with an anti-Dab1 antibody (E1), as described previously (Howell et al., 1999; Jossin et al., 2004).
Primary neuronal and embryonic slice cultures.
For primary neuronal cultures, E16.5 fetal brains were dissected in cold calcium- and magnesium-free Hanks solution supplemented with 0.6% glucose (HBSS+G). Hemispheres were isolated, and ganglionic eminences, olfactory bulbs, and meninges were discarded to leave mostly the cerebral cortex. Cortices were dissociated with trypsin (Invitrogen) for 5 min at 37°C. After trituration with a fire-polished Pasteur pipette, samples were washed in HBSS+G and triturated in 1 ml of DMEM/F-12 (Cambrex BioWhittaker, Walkersville, MD) containing 0.025% DNase I and 0.4 mg/ml soybean trypsin inhibitor (Invitrogen). Cells were washed in DMEM/F-12 and plated in 12-well plates coated with ECL cell attachment matrix (entactin, collagen IV, and laminin; Upstate Biotechnology), at a density of 1 million cells per well, in DMEM/F-12 supplemented with B27 (Invitrogen). Metalloproteinases were inhibited by adding GM6001 (Calbiochem, La Jolla, CA) at a concentration of 80 μm, and the GM6601-negative control (Calbiochem) was used at the same concentration.
The embryonic slice culture system was described previously (Jossin et al., 2003). Briefly, vibratome sections from E13 embryonic brains were cultured on collagen-coated polytetrafluoroethylene membranes (Transwell-COL, catalog #3494; Costar, Cambridge, MA) for 2 d in DMEM/F-12 supplemented with B27 and G5 (Invitrogen). Slices were cultured in a chamber (MIC-101; Billups-Rothenberg, Del Mar, CA) gassed with water-saturated 95%O2–5%CO2. Antibodies were drop-dialyzed (VSWP02500 membranes; Millipore) against culture medium and added at an IgG concentration between 0.2 and 1 mg/ml. For S-phase labeling, BrdU (Sigma) dissolved in 0.9% NaCl was administered intraperitoneally to pregnant mice at a dose of 20 μg/g body weight, at E10 for labeling of preplate cells or at E13 for analysis of cortical layering.
Data editing.
Figures prepared from morphological preparations and scanned gels were edited using Photoshop (Adobe Systems, San Jose, CA).
Results
Novel monoclonal antibodies R4B and R5A are directed against the central fragment of Reelin
After immunization of reeler mutant mice with protein 4B5A (see Materials and Methods), two IgG2b monoclonal antibodies were selected and named R4B and R5A because they are directed, respectively, against repeats 4B and 5A. Using Western blot analysis of supernatants from HEK293T cells transfected with different constructs, the R4B and R5A epitopes were mapped to sequences 1810–1825 and 1985–2058, respectively (data not shown). When applied to mouse brain sections, R4B and R5A detected immunopositive cells similar to those stained with validated anti-Reelin antibodies (Ogawa et al., 1995; de Bergeyck et al., 1998), or revealed by in situ hybridization with Reelin probes (Schiffmann et al., 1997; Alcantara et al., 1998). They reacted with Cajal-Retzius cells in the MZ of the neonatal cortex and in postnatal brains, with a subset of cortical interneurons, mitral and external granule cells in the olfactory bulb, neurons in the strata lacunosum moleculare and oriens of the hippocampus, and the hilus of the dentate gyrus (Fig. 1). Human CSF samples and mouse brain extracts were analyzed in Western blot with R4B and R5A and with antibodies 142 and 12+14 that react, respectively, with the N- and C-terminal regions (de Bergeyck et al., 1998; Ignatova et al., 2004) (Fig. 2A). Full-length and the following Reelin proteins were used as references: R3-6 (“repeats 3–6”, amino acids 1220–2664); R3-8 (“repeats 3–8”, amino acids 1220–3428); N-R6 (“N-terminal to end of repeat 6”, amino acids 1–2712); N-R2 (“N-terminal to end of repeat 2”, amino acids 1–1286); R7-8 (repeats 7 and 8, amino acids 2661–3428). In CSF, antibodies R4B and R5A reacted with Reelin proteins that contain the sequence 4B5A, particularly a protein of ∼190 kDa that comigrated with R3-6 (Fig. 2B).

Immunohistochemical validation of antibodies. Like the validated anti-Reelin G10, R4B and R5A react with Cajal-Retzius cells (CRc) in the cortical MZ of the newborn mouse [postnatal day 0 (P0)]. At P10, they stain some cortical interneurons (CI), mitral cells (Mc) and external granule cells (EGc) in the olfactory bulb, cells in the stratum oriens (SO) and stratum lacunosum moleculare (SLM) of the hippocampus, and in the hilus (H) of the dentate gyrus. Scale bar, 150 μm.

Pattern of Reelin processing. A, Reelin is cleaved at two sites (N-t and C-t), thus generating five different fragments in addition to the native polypeptide. Arrowheads point to the epitopes of the antibodies used in this study (142, G10, R4B, R5A, 12,14). F-Sp, Region with similarity to F-spondin. Repeats 1–8 are schematized as circles, with subrepeats A and B separated by epidermal growth factor motifs (vertical bar). B, Western blot of human CSF samples probed with antibodies R4B and R5A, 142, or 12+14. Supernatants from cells transfected with plasmid encoding different fragments (N-R2, R3-6, R3-8, N-R6) and full-length Reelin were loaded for reference. C, Sections in neocortex from wild-type (WT) and reeler mice at postnatal day 0, stained by immunofluorescence with the G10 (N-terminal), R4B (central), and 12,14 (C-terminal) antibodies. Cajal-Retzius cells are strongly labeled in all sections (arrows). Immunopositive dots are detected in WT but not in reeler sections, indicating that they correspond to extracellular Reelin (arrowheads). In the MZ, all three antibodies reveal extracellular Reelin, whereas only N-terminal and central epitopes are detected in the CP. Scale bar, 25 μm.
To assess whether processing might influence the diffusion of Reelin fragments in tissue, we examined postnatal day 0 wild-type cortex by immunofluorescence with antibodies against N-terminal (G10), central (R4B), and C-terminal (12,14) epitopes. As shown in Figure 2C, all three antibodies labeled intracellular Reelin in Cajal-Retzius cells as well as punctuate structures in the MZ. The latter are not detected in reeler mice, in which Reelin is not secreted (de Bergeyck et al., 1997), and correspond to extracellular Reelin, possibly Reelin oligomers or aggregates (Utsunomiya-Tate et al., 2000; Kubo et al., 2002). Intriguingly, extracellular Reelin was detected between CP cells with N-terminal and central, but not C-terminal, antibodies. This indicates that full-length Reelin is predominantly located in the MZ and that N-terminal and central fragments are both able to diffuse in the CP after cleavage.
Antibodies R4B and R5A block Reelin functions in vitro
The effect of R4B and R5A on the binding of Reelin to lipoprotein receptors was assessed in pull-down reactions. Incubation of Reelin with R4B, but not with control antibodies such as G10 or 142 (N-terminal region) or 12+14 (C-terminal region), prevented binding to VLDLR-Fc and ApoER2-Fc recombinant proteins (Fig. 3A). Similar results were obtained with antibody R5A and by using VLDLR-His and ApoER2-His proteins (data not shown). Furthermore, the ability of Reelin to stimulate Dab1 phosphorylation in neuronal cultures was drastically decreased by preincubation with R4B (Fig. 3B) or R5A but not with control antibodies. Finally, when slices from wild-type embryonic brains were cultured for 2 d in the presence of R4B (Fig. 3C) or R5A, either separately or in combination, their development resembled that of slices prepared from Reelin-deficient embryos. The MZ was not sharply defined, and the CP was disorganized. In contrast, development proceeded normally in slices incubated with the G10 antibody. Staining with a chondroitin sulfate antibody that labels the cortical preplate showed that incubation with R4B or R5A prevented preplate splitting, another hallmark of reelin deficiency (Caviness, 1982). These results suggest R4B and R5A block Reelin function.

Antibody R4B inhibits Reelin–liporeceptor binding (A) and Reelin-induced Dab1 phosphorylation (B) and interferes with CP formation in brain slices (C). A, Ectodomains of VLDLR-Fc, ApoER2-Fc, and LDLR-Fc (negative control) were adsorbed to protein A–agarose beads. Beads were incubated with Reelin (control) or with Reelin preincubated with antibodies G10, 12+14, or R4B (comparable results were obtained with R5A). Coprecipitates were eluted, processed for Western blot, and probed with anti-Reelin G10. Whereas Reelin binds to its receptors in the presence of G10 or 12+14, the R4B antibody blocks this interaction. Similar results were obtained with receptor ectodomains fused to a His tag and adsorbed to Ni-NTA beads. B, Cultured cortical neurons were stimulated with Reelin or Reelin preincubated with antibodies G10, 12+14, or R4B (similar results were obtained with R5A). Dab1 was immunoprecipitated and assessed for tyrosine phosphorylation (P-Tyr). Reelin stimulates Dab1 phosphorylation in the presence of G10 or 12+14 but not R4B. WB, Western blot. C, Slices from 13-d-old embryonic brains were cultured for 2 d (2DIV) in the presence of R4B or the control antibody G10. Development proceeds normally in the control situation (a, d), with a radially organized, dense CP located between the clearly defined MZ and subplate (SP). In slices incubated with R4B (b, e), the developing CP is disorganized, and the MZ and SP are poorly defined, like in reeler mutant mice (comparable results were obtained with R5A and with the mix R4B and R5A). Incubation with G10 (c, f) does not perturb slice development. a–c, Hematoxylin and eosin staining; d–f, immunostaining of chondroitin sulfate proteoglycans, a preplate marker. Scale bar, 100 μm. VZ, Ventricular zone; SP, subplate; IZ, intermediate zone.
Cultured neurons secrete a Reelin-processing activity
To examine the effect of neurons on processing, full-length Reelin was incubated for 20 min at 37°C in the presence of embryonic cortical cells, their supernatant, or culture medium alone, after which the pattern of Reelin cleavage was examined. Preparations of full-length Reelin (supernatants from HEK293T cells transfected with the full-length Reelin cDNA) consistently displayed basal processing (Fig. 4A, lane 1). In the presence of cortical neurons, the concentrations of N-terminal Reelin and R3-8 increased dramatically, whereas that of the full-length and N-R6 proteins decreased and that of fragment R7-8 remained relatively unchanged, indicating increased cleavage at the N-terminal but not at the C-terminal site (Fig. 4A, compare lanes 2, 3; quantification in Fig. 4B). A similar pattern was observed when Reelin was incubated with the supernatant from cultured neurons (Fig. 4A, lane 4; quantification in Fig. 4B), but processing was abrogated in the presence of the metalloproteinase inhibitor GM6001 (Fig. 4A, lanes 5–7). These results indicate that cleavage at the N-terminal site is performed by a metalloproteinase secreted by cultured embryonic neurons and that cleavage at the N and C sites may occur independently.

Cleavage of Reelin by a metalloproteinase secreted by cultured neurons. A, Western blot analysis with antibodies G10, R4B,R5A, and 12,14, of full-length Reelin at time 0 (lane 1) and after incubation with cultured neurons (lanes 2, 5), with culture medium alone (lanes 3, 6), or with the supernatant of these neurons (lanes 4, 7). Incubation was performed without (lanes 2–4) and with (lanes 5–7) the metalloproteinase inhibitor GM6001 at 80 μm. B, Densitometric analysis of three independent experiments performed without GM6001. Signals are expressed relatively to those in lane 1, corresponding to the native Reelin preparation, as mean ± SEM. **p < 0.01 (Student's t test); ***p < 0.001, significantly different. SN, Supernatant.
Processing does not affect signaling and internalization in dissociated neurons in culture
Using antibodies R4B and R5A to adjust concentrations, we tested whether processing of Reelin modifies its capacity to stimulate Dab1 phosphorylation in cultured dissociated neurons, and we compared the internalization of recombinant Reelin proteins in neurons. Similar concentrations of full-length Reelin, R3-8, R3-6, and N-R6 were equally able to stimulate Dab1 phosphorylation in neurons (Jossin et al., 2004). Because full-length Reelin was rapidly cleaved at the N-terminal site when added to neurons, Dab1 phosphorylation assays were repeated in the presence of GM6001, with similar results (Fig. 5A). Thus, inhibition of Reelin processing did not affect biological activity when tested on neurons. To study ligand internalization, cortical neurons were incubated with full-length, R3-6, or N-R2 recombinant Reelins, for 10 min at 37°C, conditions that allow endocytosis of Reelin (D'Arcangelo et al., 1999; Morimura et al., 2005), and at 4°C as control. In contrast to the N-terminal fragment N-R2, which did not enter cells, full-length Reelin and recombinant R3-6 were internalized similarly, indicating that both proteins follow similar intracellular routes when presented to dissociated cultured neurons in vitro. No Reelin internalization was detected in neurons from VLDLR−/−;ApoER2−/− mutant cortices, confirming that internalization is receptor mediated (Fig. 5B).

Processing does not affect signaling and internalization in dissociated neurons in culture. A, Comparison of Dab1 tyrosine phosphorylation (P-Dab1) induced in primary neurons by Reelin and various recombinant proteins containing the active central fragment. Similar results are obtained in the absence or presence of 80 μm GM6001. The graph shows the densitometric analysis of three independent experiments. Signals are expressed relative to the control experiments, performed by the addition of mock supernatant, as mean ± SEM. ***p < 0.001, significantly different (Student's t test). B, E16.5 cortical reeler neurons were cultured for 3 d before the addition of Reelin (a, d), R3-6 (b, e), or N-R2 (c, f), for 10 min at 37°C (a–c) or at 4°C for control experiments (d–f). Internalized proteins are detected in endocytic vesicles with the N-terminal anti-Reelin E4 (a, c, d, f) or with antibody R4B (b, e). This internalization is not detected when Reelin is added to neurons from VLDLR−/− and ApoER2−/− mutant mice (g–i).
The generation of central Reelin is required for signaling in tissue
When E13 embryonic brain slices were cultured in the presence of the metalloproteinase inhibitor GM6001, a disorganized CP developed (Fig. 6A). In slices from wild-type embryos pulsed with BrdU at E13, 2 h before culture, several BrdU-positive cells migrated beyond the Tbr1-positive neurons that were generated earlier. In contrast, in slices cultured in the presence of GM6001, like in reeler slices, no BrdU-labeled cells migrated beyond Tbr1-positive cells (Fig. 6A). Moreover, when preplate cells were labeled with BrdU in vivo at E10, the splitting of the preplate was observed in slices prepared from wild-type brains, but not in wild-type slices treated with GM6001, that developed like reeler slices (Fig. 6B). Slices cultured in the presence of the GM6001-negative control developed normally, confirming that the effects of GM6001 are attributable to inhibition of metalloproteinase activity (data not shown). Although inhibition of metalloproteinases in brain slices decreased strongly Reelin processing, the presence of some Reelin fragments in the supernatant from GM6001-treated slices indicated that processing was not completely blocked (Fig. 6C). Presumably, GM6001-resistant enzymes expressed in the organotypic culture may be responsible for residual processing activity. Yet, even the partial inhibition of cleavage downregulated Reelin signaling significantly, as indicated by decreased Dab1 phosphorylation after 4 h and upregulation of Dab1 protein concentration after 24 h (Fig. 6D) (Rice and Curran, 2001; Jossin et al., 2004). Thus, in brain slice culture, blocking Reelin processing perturbs CP development and impairs Reelin signaling.
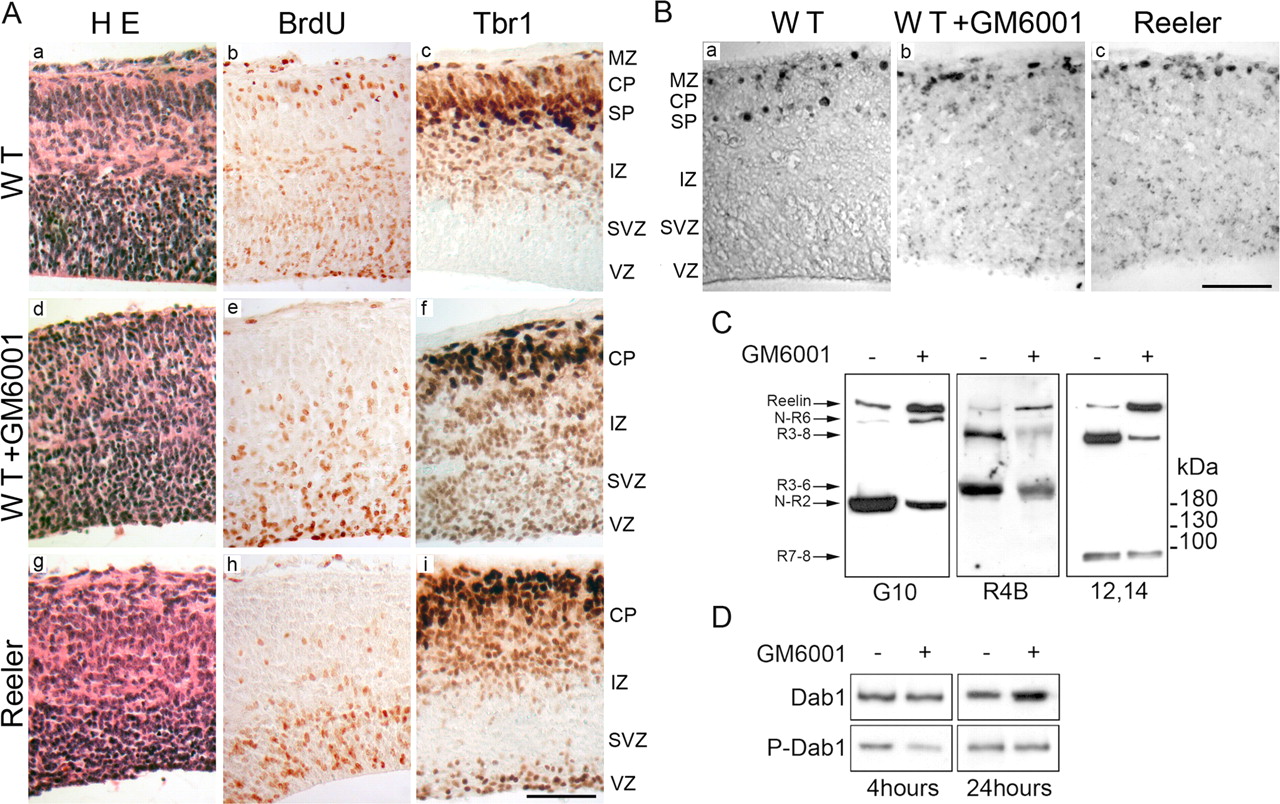
Metalloproteinases are important for CP formation (A), preplate splitting (B), and Reelin signaling (C, D). A, Slices from 13-d-old wild-type embryonic brains were cultured for 2 d with or without 80 μm GM6001. a, Development proceeds normally in the absence of the inhibitor, with a radially organized, dense CP bracketed by a clearly defined MZ and subplate. b, c, Several neurons labeled by a BrdU pulse 2 h before slice preparation (b) are able to migrate past older Tbr1-positive neurons (c), reflecting a normal inside-to-outside layering. d–i, In slices incubated with GM6001, the developing CP is disorganized, the cell-poor MZ is absent (d), and layering is inverted (e, f), a phenotype reminiscent of that in slices prepared from reeler mutant embryos (g–i). Scale bar, 100 μm. B, BrdU was administered in vivo at E10, and brain slices were prepared at E13 and cultured for 2 d. In wild-type slices, BrdU-positive cells are located in the MZ and the subplate, illustrating the splitting of the preplate (a), whereas in wild-type slices incubated in the presence of GM6001, BrdU-positive cells migrate in the subpial region (b) like in reeler slices (c), demonstrating defective preplate splitting. Scale bar, 100 μm. C, Supernatants from E13 brain slices cultured for 2 d with or without GM6001 were analyzed by Western blot with anti-Reelin antibodies G10, R4B, or 12+14. There is clear, albeit incomplete, inhibition of Reelin processing in supernatant from slices cultured in the presence of the metalloproteinase inhibitor. D, Dab1, immunoprecipitated from E13 slices cultured for variable times with and without GM6001, was analyzed by Western blot using anti-Dab1 and anti-phosphotyrosine antibodies. The presence of GM6001 decreased the phosphorylation of Dab1 after 4 h of culture. After 24 h, Dab1 phosphorylation was similar in slices incubated with or without GM6001, but the concentration of the Dab1 protein was increased in the presence of the inhibitor. WT, Wild type; SP, subplate; H E, hematoxylin and eosin staining; IZ, intermediate zone; SVZ, subventricular zone; VZ, ventricular zone.
To further investigate whether central Reelin is important for signaling in tissue, we analyzed brain extracts from wild-type and mutant mice defective in Dab1, VLDL, and ApoER2 receptors. The concentration of central Reelin was similarly low in extracts from wild-type Dab1 mutant (both null and scrambler) brains, as well as from VLDLR−/−;ApoER2+/− tissue. In contrast, it was dramatically increased in tissues from double receptor (VLDLR−/−;ApoER2−/−) mutant mice. The relative concentrations of all other Reelin forms were similar in all genotypes (Fig. 7). This result provides strong evidence that, in vivo, central Reelin binds preferentially to receptors and is selectively degraded.

The concentration of central Reelin is selectively increased in brains extracts from VLDLR−/−;ApoER2−/− mutant mice. A, Western blot analysis of brain extracts from wild-type (WT), VLDLR−/−, and ApoER2−/− mutant, Scrambler (Dab1Scm/Scm), and reeler (RelnOrl) probed with Reelin antibodies G10, R4B, and 12+14. The specific upregulation of central Reelin in double receptor-defective tissue is evident (arrow). B, Western blot analysis of brain lysates from five littermates with different doses of ApoER2 on a VLDLR mutant background, revealed with the R4B antibody. Only in double receptor mutant tissue is the concentration of central Reelin increased (arrows). C, Western blot analysis of brain lysates from six littermates with different Dab1 genotypes, revealed with the R4B antibody. The signal related to central Reelin (R3-6) is unaffected by genotype. The Dab1 mutation used in this experiment is the null allele (gift from J. Cooper).
Discussion
Antibodies R4B and R5A, directed against epitopes in the central region of Reelin, allow detection of the predicted central processing fragment in tissue extracts and CSF and block signaling, confirming the pivotal role of central Reelin (Jossin et al., 2004). We also provide evidence that processing of Reelin is important for function in vivo. Embryonic neurons in culture secrete a metalloproteinase activity that processes Reelin. Inhibition of cleavage does not affect Reelin signaling in cultured dissociated neurons, which are able to internalize full-length and central Reelin similarly. In contrast, inhibition of metalloproteinases in organotypic brain slice cultures inhibits Reelin cleavage and signaling and disturbs the formation of the CP. Furthermore, the relative concentration of central Reelin is dramatically increased in brains from mice deficient in VLDL and ApoER2 receptors, indicating that this fragment is preferentially internalized by target cells in vivo. To resolve the apparent discrepancy between observations in tissue and in cultured neurons, we propose that, in tissue, full-length Reelin cannot diffuse freely because it is anchored to the ECM and that metalloproteinases secreted by migrating neurons liberate central Reelin that diffuses toward and activates signaling in target cells (Fig. 8). Below, we discuss briefly the role of central versus N-terminal Reelin, Reelin endocytosis, and some implications of the proposed model.

Model for the production and action of central Reelin in tissue. Reelin secreted by Cajal-Retzius cells (CRc) in the MZ is anchored to the ECM, probably by its N-terminal (black) and possibly C-terminal (red) regions. Processing at an N-terminal site by a metalloproteinase secreted by migrating neurons (black scissors) and at a C-terminal site by an undefined enzyme (red scissors) generates N-terminal, C-terminal, and central (yellow) fragments. The central fragment is then free to bind to receptors at the leading edge of CP neurons and undergoes endocytosis.
Previous studies with antibodies against the N- and C-terminal regions of Reelin showed that several fragments are present in brain extracts, CSF, and plasma (Lambert de Rouvroit et al., 1999; Smalheiser et al., 2000; Ignatova et al., 2004). The pattern observed indicated that Reelin is cleaved at two sites located, respectively, C-terminal to repeats 2 and 6. However, available reagents did not allow detection of the central predicted fragment that we now characterize directly. Central Reelin is necessary and sufficient to bind to VLDL and ApoER2 receptors, to induce phosphorylation of Dab1, and to rescue the organization of the CP in organotypic brain slices from reeler mutant embryos (Jossin et al., 2004). Here, we also demonstrate that, by preventing binding of Reelin to VLDLR and ApoER2, antibodies R4B and R5A block all known functions of Reelin during early cortical development. This provides additional argument for the crucial role of central Reelin, also underscored in a recent structural study (Nogi et al., 2006). On the other hand, numerous studies using the CR50 antibody (Ogawa et al., 1995) point to the importance of the N-terminal region. Although CR50 is directed against an epitope in the N-terminal region that is not engaged in binding to receptors (Hiesberger et al., 1999; Jossin et al., 2004), it counteracts the action of Reelin in various assays (Ogawa et al., 1995; Nakajima et al., 1997; D'Arcangelo et al., 1999; Utsunomiya-Tate et al., 2000; Hack et al., 2002; Kubo et al., 2002; Niu et al., 2004; Cariboni et al., 2005). At first sight, the blocking action of CR50 would predict that fragments of Reelin that do not contain the N-terminal region should have reduced biological activity, which is not what we observe. Several explanations can be proposed. Because CR50 cannot be used in Western blot and probably recognizes a configuration epitope, a cross-reaction with central Reelin is not formally excluded. Alternatively, fragments of Reelin could remain physically associated after cleavage, and CR50 might affect indirectly the structure of the central region in such complexes, or perturb receptor binding by steric hindrance. Finally, incubation with CR50 may hamper N-terminal processing and prevent the formation of the central fragment or its access to receptors on target cells. The functional role of central Reelin is also relevant to studies of Reelin in human pathology. Thus far, most analyses were performed using antibody 142 that detects an epitope in N-terminal Reelin. Because extracellular Reelin is almost completely cleaved in tissue, conclusions in terms of function should be considered carefully and evaluated further by studies of central Reelin.
A main finding of the present work is that the concentration of central Reelin is dramatically increased in brains from double receptor mutant mice. This suggests strongly that production of central Reelin is required in tissue and that this fragment is downregulated in a receptor-dependent manner, presumably by endocytosis. In line with this, the central fragment is rapidly internalized by neurons in a receptor-dependent manner, whereas the N-terminal fragment is not observed in intracellular vesicles. Full-length Reelin is also internalized, indicating that processing is not required for endocytosis. Thus, the selective clearance of the central fragment in vivo likely reflects preferential access to cells, rather than high affinity for receptors. The observation that the concentration of central Reelin is normal in Dab1 mutant tissue indicates that downregulation occurs in parallel to signaling. Because Dab1 regulates receptor trafficking (Morimura et al., 2005), this finding was somewhat unexpected. However, the reported effect of Dab1 on receptor expression and decreased Reelin uptake is moderate (Morimura et al., 2005), whereas a complete inactivation of both VLDLR and ApoER2 is absolutely required to generate a reeler-like phenotype (Trommsdorff et al., 1999) and to upregulate the concentration of central Reelin in vivo (Fig. 7). Presumably, compensation or adaptation of the trafficking process occurs during development, dampening subtle events that are demonstrated better in vitro. Like Dab1, adaptors implicated in endocytosis, such as ARH and Dab2, bind to the NPXY motif in the tail of Reelin receptors (Morris and Cooper, 2001; Jones et al., 2003; Cuitino et al., 2005; Maurer and Cooper, 2005; Stolt and Bock, 2006). The prompt degradation of Dab1 by the proteasome after Reelin signaling and Dab1 phosphorylation (Arnaud et al., 2003b; Bock et al., 2004) may facilitate their docking to receptors, followed by clathrin-mediated clustering in coated pits and endocytosis.
The model schematized in Figure 8 posits that, after secretion, Reelin is anchored to the ECM and that the central fragment is released by metalloproteinase activities secreted by migrating neurons. ECM anchoring could occur via formation of Reelin oligomers or aggregates (Utsunomiya-Tate et al., 2000; Kubo et al., 2002), or by interaction with other ECM proteins. The observation that extracellular Reelin is detected in the CP with N-terminal and central antibodies but not with C-terminal antibodies suggests that both the N-terminal and central Reelin can diffuse after cleavage and that the C terminus might also contribute to ECM attachment. We have preliminary evidence that a high molecular weight component coimmunoprecipitates with Reelin in embryonic brain explants, but thus far no Reelin-binding ECM molecule has been identified. The concept that active fragments from proteins attached to the ECM are released by proteolytic processing is not new and was proposed, among others, for F-spondin (Tzarfaty-Majar et al., 2001), hepatocyte growth factor (Matsuoka et al., 2006), or transforming growth factor-β (Dallas et al., 2002).
Although the implication of metalloproteinases in Reelin processing is established beyond reasonable doubt, several questions remain. The metalloproteinase responsible for the cleavage of Reelin at the N-terminal site is secreted by cortical cells in culture, but its identity and the sequence at the cleavage site remain unknown. In addition, processing is never fully prevented by metalloproteinase inhibitors in slice cultures, suggesting that other enzymes are able to process Reelin in some conditions. Cleavage at the C-terminal site may be performed by a different enzymatic activity (Lugli et al., 2003) that is even less characterized. Although our observations provide strong argument that processing is required for Reelin function in tissue, definitive proof awaits demonstration in vivo. As far as we know, no mouse with genetic inactivation of metalloproteinases has a reeler-like phenotype, but there is obvious redundancy among proteolytic enzymes, and multiple knock-outs are often embryonic lethal, preventing manifestation of the reeler trait. A partial rescue of the reeler phenotype was achieved in transgenic mice that express full-length Reelin under control of the nestin promoter (Magdaleno et al., 2002). Our model would predict that such rescue will not happen in transgenic mice that express a cleavage-resistant Reelin, but the production of such mice also requires identification of the cleavage sites.
Footnotes
-
This work was supported by Actions de Recherches Concertées Grant ARC 02/07-276, Fonds de la Recherche Fondamentale et Collective Grant 2.4504.01, and by the Fondation Médicale Reine Elisabeth, all from Belgium. Y.J. is a postdoctoral researcher with the Fonds National de la Recherche Scientifique, Belgium, and L.G. was on a fellowship from the University of Louvain. We thank Fadel Tissir and Jacques Van Snick for discussion, Isabelle Lambermont and Esther Paitre for technical assistance, Vincent Strooband for advice on epitope mapping, Christian Sindic for human CSF samples, Joachim Herz for lipoprotein receptor constructs, Jon Cooper for Dab1 mutant brain tissue, and Tom Curran for the Reelin cDNA.
- Correspondence should be addressed to Dr. André M. Goffinet, Developmental Neurobiology Unit, Université Catholique de Louvain, Avenue E. Mounier, 73, Box DENE 7382, B1200 Brussels, Belgium. Andre.Goffinet{at}dene.ucl.ac.be





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}