

Abstract
Multiple descending motor pathways likely contribute to the recovery of hand motor function following spinal cord injury (SCI). Reticulospinal neurons project to spinal motor neurons controlling hand muscles and extensively sprout into gray matter structures after SCI; therefore, it has been proposed that the reticulospinal tract is one of the descending motor pathways involved in recovery of hand function after injury. To test this hypothesis, we examined the StartReact response, an involuntary release of a planned movement via a startling stimulus that engages the reticulospinal tract, by measuring reaction times from electromyographic activity in an intrinsic finger muscle during three motor tasks requiring different degrees of hand dexterity: index finger abduction, a precision grip, and a power grip. Males and females with and without incomplete chronic cervical SCI were tested. We found that although SCI participants voluntarily responded to all tasks, reaction times were shorter during a startle cue while performing a power grip but not index finger abduction or precision grip. Control subjects had similarly shorter reaction times during a startle cue in all motor tasks. These results provide the first evidence for a contribution of the reticulospinal tract to hand control in humans with SCI during gross finger manipulations and suggest that this contribution is less pronounced during fine dexterous finger movements.
SIGNIFICANCE STATEMENT It has been long proposed that brainstem pathways contribute to the recovery of hand function in humans with spinal cord injury (SCI). Here, we show that individuals with anatomically incomplete chronic cervical SCI responded to a startle stimulus, a test that engages the reticulospinal tract, while performing a power grip but not during index finger abduction or precision grip. Control subjects responded to a startle stimulus similarly across tasks. These observations suggest that reticulospinal outputs after SCI contribute to hand motor tasks involving gross finger movements. Interestingly, this contribution is less pronounced during fine dexterous finger movements.
Introduction
An increasing number of anatomical and physiological studies in animals support the view that brainstem and propriospinal pathways play a role in the recovery of hand function following spinal cord injury (SCI; for reviews, see Isa et al., 2013; Isa and Nishimura, 2014; Baker et al., 2015). One of these pathways is the reticulospinal tract, a major descending motor pathway that projects directly and indirectly to spinal motor neurons controlling hand muscles (Riddle et al., 2009). A single reticulospinal neuron exhibits extensive collateralization projecting to multiple motor neuron pools (Peterson et al., 1979; Matsuyama et al., 1997), suggesting that this tract will be capable of activating groups of muscles required for finger manipulations. So far, the contribution of the reticulospinal tract to hand control following incomplete SCI remains unknown.
In humans, voluntary reaction time can be shortened by an acoustic startling cue, a process known as the StartReact response. StartReact is thought to reflect an involuntary release of a planned movement when an unexpected startle cue is presented (Brown et al., 1991; Valls-Sole et al., 1995, 1999). This is supported by the observation that a StartReact response is not present following a choice reaction time paradigm, but occurs when a single pre-prepared movement just needs to be released (Carlsen et al., 2004). The shortening in reaction time may involve subcortical structures such as the reticular formation (Davis and Gendelman, 1977; Davis et al., 1982; Valls-Sole et al., 1999; Carlsen et al., 2003, 2004, 2009). In agreement with this, patients with Parkinson's disease and associated dysfunction of the reticular formation and pedunculopontine nucleus do have a reduced StartReact response (Nonnekes et al., 2014). However, a contribution from cortical pathways cannot be excluded (Marinovic and Tresilian, 2016). StartReact responses in finger muscles have a similar degree of muscle fractionation to rapid responses without a startling cue, suggesting that the reticulospinal and corticospinal tract contribute in parallel to the StartReact paradigm (Dean and Baker, 2017), just as they do to voluntary movements to a non-startling cue (Soteropoulos et al., 2012). In patients with degeneration of the corticospinal tract without clinical weakness, the StartReact effect is greater (Fisher et al., 2013), which may suggest an enhanced reticulospinal outflow to compensate for the loss of corticospinal function. One unanswered question is whether, after SCI, the reticulospinal tract contributes to motor tasks involving different degrees of finger dexterity. On one side, the corticospinal tract contributes to manipulation requiring fine fractionated digit movements, such as a precision grip, in primates (Buys et al., 1986; Bennett and Lemon, 1996) and humans with and without SCI (Bunday et al., 2014; Perez and Rothwell, 2015), and lesion of the corticospinal tract impairs fine dexterous finger tasks (Lawrence and Kuypers, 1968; Lang and Schieber, 2003, 2004). On the other side, reticulospinal outputs are strengthened following a corticospinal lesion and these connections subserve some of the recovery of gross hand motor function (Zaaimi et al., 2012). Also after SCI, reticulospinal axons sprout extensively into gray matter structures contributing to performance of gross motor behaviors (Ballermann and Fouad, 2006; Filli et al., 2014). We therefore hypothesized a preferential contribution of the reticulospinal tract following SCI to gross hand function such as power grip rather than fine manipulations such as precision grip or independent finger movements.
Materials and Methods
Subjects.
Seventeen individuals with SCI (mean age = 46.8 ± 15.6 years, 1 female; Table 1) and 22 age-matched controls (mean age = 40.1 ± 15.3 years, p = 0.2, 10 female) participated in the study. All subjects gave informed consent to the experimental procedures, which were approved by the local ethics committee at the University of Miami. SCI subjects had a chronic (≥1 year), cervical injury (C3–C8), an intact or impaired, but not absent innervation in dermatome C6 during light touch and pin-prick stimulus using the International Standards for Neurological Classification of Spinal Cord Injury sensory scores and residual hand motor function (Table 1).
SCI participants
Electromyography (EMG).
EMG was recorded from the first dorsal interosseous (FDI) muscle of the dominant arm in control subjects and from the less affected hand in individuals with SCI through surface electrodes secured to the skin over the belly of each muscle (Ag–AgCl, 10 mm diameter). EMG was also recorded from the sternocleidomastoid neck muscle (SCM; in 16 controls and 8 SCI subjects). The signals were amplified, filtered (20–1000 Hz), and sampled at 2 kHz for off-line analysis (CED 1401 with Spike2 software, Cambridge Electronic Design). During a maximal voluntary contraction (MVC), force exerted at the proximal interphalangeal joint of the index finger and thumb (Honeywell, range ± 498.1 N, voltage ± 5 V, high-sensitivity transducer 0.045 V/N) was measured by load cells. Force was sampled at 200 Hz and stored on a computer for off-line analysis.
Experimental paradigm.
During testing, subjects were seated with both arms flexed at the elbow by 90° and the StartReact response was examined when subjects performed index finger abduction, a precision grip, and a power grip in a randomized order (Fig. 1A). During index finger abduction, individuals were instructed to press with the index finger against a custom lever in the abduction direction with the forearm pronated and the wrist restrained by straps. During a precision grip, subjects were asked to pinch the index finger and thumb while the forearm was maintained in the neutral position and the wrist was restrained by straps. When performing a power grip, subjects were instructed to make a fist, requiring all fingers to flex at the metacarpophalangeal and proximal interphalangeal joints with the forearm maintained in the neutral position and the wrist restrained by straps. At the beginning of the experiment subjects performed two brief MVCs (3–5 s) with the index finger into abduction, separated by 30–60 s of rest. We examined the StartReact response using a previously tested paradigm (Fisher et al., 2013; Fig. 1B). Here, participants were asked to observe a red light-emitting diode (LED) located ∼1 m in front of them. When the LED was illuminated, individuals were asked to perform as fast as possible the current hand task. The LED remained illuminated for 20 ms. We measured the visual reaction time (VRT) as the time from cue to onset of the EMG burst in the FDI muscle after the LED presentation. In some trials, the LED was presented with either a quiet acoustic stimulus (80 dB; 500 Hz, 20 ms) or a startling acoustic stimulus (SAS; 115 dB; 500 Hz, 20 ms). These intensities are comparable to those which have been used in previous studies (Fisher et al., 2013; Nonnekes et al., 2014; Dean and Baker, 2017). The loud intensity evoked a clear startle in control subjects and in some individuals with SCI on initial presentation. The time delay between the presentation of the quiet acoustic stimulus and the onset of the EMG response was referred as the visual-auditory reaction time (VART), whereas the time between the SAS and the EMG onset was defined as the visual + startle reaction time (VSRT). Data were analyzed trial by trial using MATLAB (MathWorks). An automated program identified the reaction time, defined as the time point where mean rectified EMG signals exceeded 3 SD of the mean EMG measured 100 ms before each stimulus presentation; every trial was inspected visually, and erroneous activity onset times (caused, for example, by electrical noise artifacts) were manually corrected. The reaction time was used to calculate the mean of the rectified EMG activity in the FDI muscle starting at burst onset for 100 ms. EMG activity in the SCM was also defined as present when mean rectified EMG signals exceeded 3 SD of the mean EMG measured 100 ms before each stimulus presentation. As in previous studies (Carlsen et al., 2004, 2009), we calculated the number of trials in which SCM activity was present as a percentage of the total number of trials tested in each condition. A familiarization trial consisting of three to four repetitions of each task responding to the LED was completed at the beginning of each experiment to ensure that control and SCI subjects were able to complete all tasks. After that subjects were presented with five consecutive SAS, without performing the task, to get them familiarized with the startling cue (Fisher et al., 2013). In each task, 20 responses were recorded in each condition (VRT, VART, and VSRT) in a randomized order with an interval between trials of 5 s. The VART and VSRT are both mediated via the cochlear nuclei, but only the high intensity sound of the VSRT activates the reticulospinal tract. Therefore, to estimate changes in the gain of reticulospinal outputs we normalized the data as follows:
 6.1 ± 3.8% of the total of all trials, in which mean rectified EMG activity exceeded ±2 SD of the mean rectified EMG measured 100 ms before the presentation of an LED with and without a quiet or a startling acoustic stimulus, were excluded from further analysis to ensure that subjects were at rest across tasks at the presentation of the cue onset.
6.1 ± 3.8% of the total of all trials, in which mean rectified EMG activity exceeded ±2 SD of the mean rectified EMG measured 100 ms before the presentation of an LED with and without a quiet or a startling acoustic stimulus, were excluded from further analysis to ensure that subjects were at rest across tasks at the presentation of the cue onset.

Experimental setup. A, Schematic of the hand postures tested including index finger abduction, a precision grip, and a power grip. B, Representation of the paradigm used to examine the StartReact response in both groups. A red LED was presented alone or in combination with a quiet acoustic stimulus (80 dB; 500 Hz, 20 ms) or a SAS (115 dB; 500 Hz, 20 ms).
Data analysis.
Normal distribution was tested by the Shapiro–Wilk test and homogeneity of variances by the Levene's test of equality and Mauchly's test of sphericity. When sphericity could not be assumed the Greenhouse–Geisser correction statistic was used. Repeated-measures ANOVAs were performed to determine the effect of Group (controls, SCI), Task (index finger abduction, precision grip, power grip), and Condition (VRT, VART, VSRT) on reaction time, and EMG activity measured before and after the reaction time. Repeated-measures ANOVAs were also performed to determine the effect of Group and Task on VRT, VART, VSRT, and ΔTSR/ΔTAR ratio. Additional repeated ANOVAs were performed on each group separately as needed. Bonferroni post hoc tests were used to test for significant comparisons. Pairwise comparisons were used to test for significant comparisons between MVCs and age. Paired t tests were used to compare reaction times in trials in which EMG activity in the SCM muscle was present versus absent. A Pearson correlation analysis was used as needed. Significance was set at p < 0.05. Group data are presented as the mean ± SD in the text.
Results
EMG
EMG activity during index finger abduction MVC was larger in controls than in SCI subjects (controls = 0.59 ± 0.3 mV; SCI = 0.13 ± 0.1 mV; p < 0.001). Repeated-measures ANOVA showed no effect of Group (F(1,37) = 1.0, p = 0.3, ηp2 = 0.03), Condition (F(1.3,74) = 2.3, p = 0.13, ηp2 = 0.05), Task (F(1.3,74) = 0.21, p = 0.7, ηp2 = 0.006) nor in their interaction (F(2.1,148) = 0.23, p = 0.8, ηp2 = 0.006) on mean rectified EMG activity in the FDI muscle measured 100 ms before stimulus presentation. However, we found an effect of Group (F(1,37) = 14.8, p < 0.001, ηp2 = 0.28), Condition (F(1.4,74) = 17.0, p < 0.001, ηp2 = 0.31), Task (F(1.4,74) = 26.8, p < 0.001 ηp2 = 0.42), and in their interaction (F(2.4,148) = 3.4, p = 0.03, ηp2 = 0.09) on mean rectified background EMG activity in the FDI muscle measured 100 ms after the reaction time. Pairwise comparison revealed that in control subjects the mean EMG activity measured 100 ms after the reaction time was larger during VRST compared with VRT (index finger abduction, p < 0.01; precision grip, p < 0.01; power grip, p = 0.01) and VART (index finger abduction, p = 0.01; precision grip, p = 0.02; power grip, p < 0.01) trials in all tasks. Also, mean EMG activity was larger during VART compared with VRT trials in all tasks (index finger abduction, p = 0.04; precision grip, p = 0.01; power grip, p = 0.03). In SCI participants, mean EMG activity was larger during VRST compared with VRT (p < 0.01) and VART (p = 0.02) trials only during a power grip. Mean EMG activity was also larger during VART compared with VRT trials (p = 0.01). During index finger abduction (p = 0.31) and precision grip (p = 0.17) no differences were found between VSRT and VART trials.
EMG activity in the SCM muscle was not observed in VRT trials in both groups but it was sometimes present in VART (controls: 2/16 subjects in 12.5% of trials; SCI: 4/8 subjects in 18.9% of trials) and VSRT (controls: 5/16 subjects in 20% of trials; SCI: 5/8 subjects in 49% of trials). No differences were found in reaction time in VSRT trials with and without the presence of SCM muscle activity in controls (p = 0.4) and SCI (p = 0.8) subjects.
VRT, VART, and VSRT
Repeated-measures ANOVA showed a significant effect of Group (F(1,37) = 12.6, p < 0.01, ηp2 = 0.25), Condition (F(1.4,74) = 324.4, p < 0.001, ηp2 = 0.89), not Task (F(2,74) = 1.1, p = 0.32, ηp2 = 0.03) but in their interaction (F(2.7,148) = 3.5, p = 0.02, ηp2 = 0.09) on reaction time. Post hoc testing showed that VRTs were prolonged across tasks in SCI (index finger abduction, 236 ± 53 ms; precision grip, 223 ± 52 ms; power grip, 241 ± 56 ms) compared with controls (index finger abduction, 182 ± 32 ms; precision grip, 197 ± 34 ms; power grip, 196 ± 34 ms) subjects (p < 0.01; Fig. 2A). We also found that VARTs were prolonged across tasks in SCI (index finger abduction, 188 ± 55 ms; precision grip, 170 ± 38 ms; power grip, 195 ± 32 ms) compared with controls (index finger abduction, 136 ± 31 ms; precision grip, 157 ± 35 ms; power grip, 151 ± 31 ms) subjects (p = 0.01; Fig. 2B). VSRTs were prolonged across tasks in SCI (index finger abduction, 180 ± 54 ms; precision grip, 163 ± 41 ms; power grip, 168 ± 37 ms) compared with controls (index finger abduction, 115 ± 35 ms; precision grip, 135 ± 40 ms; power grip, 126 ± 31 ms) subjects (p < 0.01; Fig. 2C).

VRT, VART, and SVRT. We measured the VRT (A), VART (B), and VSRT (C) during index finger abduction, precision grip, and power grip in controls and SCI participants. Group data are shown in bar graphs (controls, n = 22; SCI, n = 17). The abscissa shows the hand postures tested in both groups (index finger abduction, white bars; precision grip, light gray bars; power grip, dark gray bars). The ordinate shows the reaction time (in milliseconds). Note that overall, VRT, VART, and VSRT values were larger in SCI compared with control participants. Error bars indicate SE. *p < 0.05.
To ensure that any changes in reaction times were related to changes in reticulospinal gain, we compared the ΔTSR/ΔTAR ratios across groups and tasks. Figure 3A illustrates rectified EMG traces in the FDI muscle during all tasks in a representative control and SCI participant. Note that reaction time shortened in the control subject to a similar extent during the VSRT compared with the VART in all hand motor tasks. By contrast, in the SCI participant reaction times further decreased during VSRT compared with the VART only during a power grip. Repeated-measures ANOVA showed a significant effect of Group (F(1,37) = 5.2, p = 0.02, ηp2 = 0.12), not Task (F(2,74) = 2.4, p = 0.09, ηp2 = 0.06) but in their interaction (F(2,74) = 6.9, p < 0.01, ηp2 = 0.16) on ΔTSR/ΔTAR ratios. Post hoc testing showed that ratios were similar in controls subjects across tasks (p = 0.33; Fig. 3B). However, in SCI subjects ratios were larger during power grip compared with index finger abduction (p < 0.01) and precision grip (p < 0.01; Fig. 3B). Also, in SCI participants, ratios were lower during index finger abduction (p = 0.01) and precision grip (p < 0.01) compared with control subjects but similar during power grip (p = 0.14).
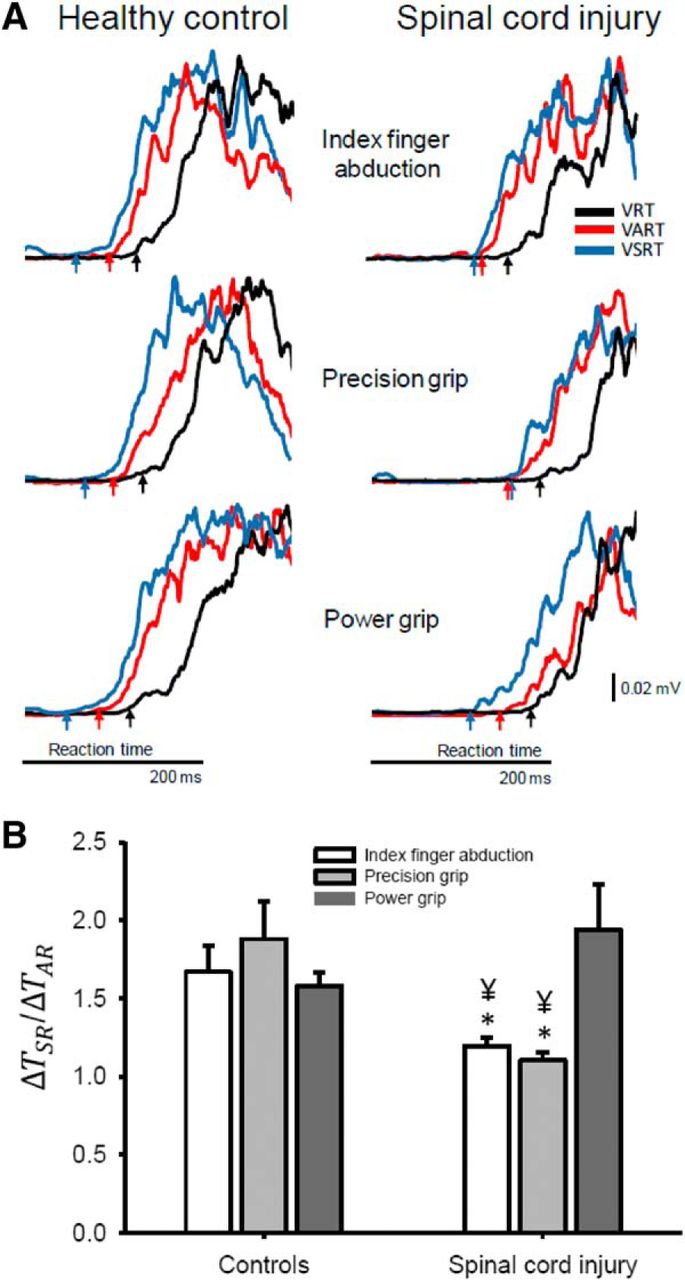
ΔTSR/ΔTAR ratios. A, Raw EMG data in a representative control (left) and SCI (right) subject showing rectified traces during VRT (black traces), VART (red traces), and VSRT (blue traces). Reaction times were measured during all conditions and arrows below each of the traces indicates the reaction time in each of the conditions tested. The EMG traces were aligned to the cue onset. The cue onset is not shown to zoom in on the reaction time across conditions (see arrows). Note that VSRT was always shorter in all tasks in the control subject, whereas in the SCI subject VSRT was only shorter during the power grip task. B, Group data are shown in bar graphs (controls, n = 22; SCI, n = 17). The abscissa shows the hand postures tested in both groups (index finger abduction, white bars; precision grip, light gray bars; power grip, dark gray bars). The ordinate shows the ΔTSR/ΔTAR ratios (see Materials and Methods for a detailed description). Note that in SCI subjects ratios were maintained to a similar level as control subjects during power grip but not during index finger abduction and precision grip. Error bars indicate SE. *p < 0.05, comparison between groups; ¥p < 0.05, comparison between tasks.
Discussion
Our novel results indicate that the reticulospinal tract contributes to hand motor tasks involving gross finger manipulations but to a lesser extent to fine dexterous finger movements in humans with anatomically incomplete chronic cervical SCI. We found that a startling cue shortened reaction times to a similar extent in control subjects regardless of what hand task they performed. However, although participants with SCI voluntarily responded in all tasks, their reaction times relative to the quiet acoustic cue were only shortened by a startling cue while performing a power grip but not index finger abduction or precision grip. We hypothesize that differences in functional contributions from reticulospinal and corticospinal tracts to hand control may explain our results in SCI participants.
Contribution of the reticulospinal tract to hand control in uninjured humans
So far, the StartReact paradigm has been used as a noninvasive tool to assess the influence of the reticulospinal tract on human movement. During StartReact responses voluntary reaction times can be shortened by ∼70 ms if a quiet acoustic stimulus is replaced by a loud acoustic stimulus, likely due to the engagement of subcortical structures such as the reticular formation (Brown et al., 1991; Valls-Sole et al., 1995, 1999; Carlsen et al., 2003, 2004, 2009) although the contribution from cortical pathways cannot be excluded (Marinovic and Tresilian, 2016). Anatomical data from microstimulation and spike-triggered averaging studies show that a single corticospinal cell facilitates hand and forearm muscles with close functional relations (Buys et al., 1986) making it more suitable for small fractionated movements. The corticospinal tract does indeed seem to be heavily involved in manipulations requiring fine fractionated digit movements, such as a precision grip, in primates (Buys et al., 1986; Bennett and Lemon, 1996) and humans (Bunday et al., 2014; Perez and Rothwell, 2015). Corticomotoneuronal cells fire preferentially during a precision grip compared with a power grip (Muir and Lemon, 1983), and in humans the motor cortex is less involved during power grip compared with index finger abduction and precision grip (Tazoe and Perez, 2017). A single reticulospinal neuron by contrast has extensive collateralizations projecting to multiple motoneuron pools (Peterson et al., 1979; Matsuyama et al., 1997). This means that, on its own, the reticulospinal tract can only generate gross, and not fine finger manipulations. However, cells in the reticular formation do modulate their discharge strongly during isolated independent finger movements (Soteropoulos et al., 2012). In the uninjured state, therefore, previous data support the idea that fine finger movements are controlled cooperatively by both corticospinal and reticulospinal pathways (as well as other subcortical circuits, such as the C3–C4 propriospinal system; Baker, 2011), whereas power grip has a much reduced corticospinal tract contribution.
Consistent with the concept that the reticulospinal tract contributes to all types of hand movement, we found in control subjects that reaction times shortened to a similar extent after a startling cue during index finger abduction, precision, and power grip. Dean and Baker (2017) likewise demonstrated a StartReact effect on an even wider range of different hand and arm tasks. Recent electrophysiological data show that a startling cue modulates the size of responses following stimulation of corticospinal axons to the same extent during all of the motor tasks described here (Tazoe and Perez, 2017). As similar results were seen for electrical stimulation at the cortex and in the cervico-medullary junction, this likely reflects subcortical contributions (Taylor and Gandevia, 2004). Although we did not record reticular activity directly, these observations suggest that in control subjects the interactions between the reticulospinal and corticospinal pathway, as well as other spinal and/or descending motor pathways involved in these hand manipulations, contributed to the similar StartReact response across tasks.
Contribution of the reticulospinal tract to hand control after SCI
An intriguing question is why a startling cue decreased reaction time during a power grip but not index finger abduction or precision grip in humans with SCI? We favor the hypothesis that these results reflect differences in the relative strength of functional contributions from the reticulospinal and corticospinal tract to these different hand manipulations, which become more apparent when these tracts are affected by an injury.
Most SCIs in humans are associated with high energy fracture-dislocations and burst fractures (Oxland et al., 2010), which impact the ventral region of the spinal cord. Although the corticospinal tract is more dorsally located in the lateral funiculus than the reticulospinal tract, in many cases both will suffer injury. Both injured corticospinal and reticulospinal axons can form connections with nearby target neurons of other undamaged tracts (Oudega and Perez, 2012). Our SCI participants had prolonged reaction times compared with control subjects for all conditions tested, agreeing with a generalized impairment. The reduced number and/or strength of descending connections may force SCI subjects to use the strongest available contributions in each task. The selective shortening in reaction time with a startling cue during power grip, but not during the other motor tasks, in SCI participants suggests that reticulospinal connections are available after injury and that these connections are especially associated with the power grip task. The reticulospinal tract may have a greater capacity for regeneration than the corticospinal tract (Vavrek et al., 2007; Zörner et al., 2014). In monkeys, reticulospinal outputs are strengthened following a corticospinal lesion and these connections subserve some of the recovery of gross hand motor function (Zaaimi et al., 2012). After SCI, reticulospinal axons spontaneously arborize (Ballermann and Fouad, 2006; Weishaupt et al., 2013) and form contacts with relay neurons (Filli et al., 2014), which results in improvement of gross motor functions. Thus, the enhanced ability to strengthen reticulospinal compared with corticospinal connections may mean that in SCI subjects with some recovery of hand function, there is an even greater dependence on reticulospinal contributions to gross compared with fine dexterous finger manipulations. The lesser contribution of the reticulospinal tract to fine finger manipulations may reflect the relative importance of the corticospinal tract in this skilled grasping behavior (Lemon and Griffiths, 2005). This also agrees with evidence showing that StartReact responses are larger in patients with degeneration of the corticospinal tract without clinical weakness, consistent with the view that strengthened reticulospinal connections contribute to compensate for the loss of corticospinal function (Fisher et al., 2013).
It is also important to consider whether some methodological factors might have contributed to our results. Previous work on the StartReact response recorded from the SCM muscle as a marker of an overt startle response, and found activation in the SCM on trials with a loud cue (Valls-Sole et al., 1999). Later studies found that SCM activation following loud cues was more variable and not present at all times when a startling cue was presented during different hand motor tasks (Honeycutt et al., 2013; Dean and Baker, 2017). Consistent with these results, we found that SCM activation during startling cues was unreliable and it was present both in trials with a loud and a quite cue. This might reflect that the StartReact response has important differences from an overt startle response. For example, StartReact responses do not habituate and do not show pre-pulse inhibition (Valls-Sole et al., 2005). Therefore, we selected trials following a loud cue based on their reaction time rather than using SCM recordings. Note that the shortening in reaction time that we observed during the startling cue was comparable to what has been presented in previous studies during hand motor tasks (Fisher et al., 2013; Honeycutt et al., 2013; Dean and Baker, 2017), supporting the view that a StartReact response was evoked. In addition, all control subjects showed an overt startle response to the first presentation of the loud stimulus, indicating that this did recruit startle circuits. Clear startle responses were also seen in some patients, although it was difficult to assess this reliably by eye given the constraints of sitting in a wheelchair and the level of overall weakness, especially in the trunk. The StartReact results are also based on changes in reaction time across conditions and differences in reaction time exist across groups. We found that reaction times were prolonged in SCI subjects compared with controls in all tasks and conditions, which is consistent with previous results (Federico and Perez, 2017). This might reflect the longer time needed to raise spinal motoneurons to threshold because of the reduced descending drive after the injury. In chronic SCI, spinal motoneurons are activated by longer periods of depolarization compared with controls (Norton et al., 2008). Prolonged reaction times in humans with SCI have also been related to an altered ability to synchronize corticospinal descending volleys at the spinal cord level (Cirillo et al., 2016). Importantly, SCI subjects were able to shorten their reaction times during a quiet and a loud stimulus compared with a visual stimulus alone across tasks, in a proportional manner to controls. This indicates that the injury did not limit their ability to perform all tasks and became faster as needed and suggests that it is less likely that this factor affected our results.
As noted above, without direct recordings from the reticular formation our conclusions about reticulospinal activity are necessarily indirect, and there may also be a corticospinal contribution to the StartReact effect (Marinovic and Tresilian, 2016). However, it is difficult to explain why reaction time shortening in SCI subjects following a loud stimulus should occur preferentially for the power grip on the basis of corticospinal involvement alone, as it is known that corticospinal neurons show little involvement in this task (Muir and Lemon, 1983).
Functional considerations
More than 50% of humans with SCI have cervical damage and consequently show impairments in hand function. Our results indicate that the reticulospinal tract contributes to hand motor tasks involving gross finger manipulations but to a lesser extent to more dexterous finger movements in humans with cervical SCI, which may open new routes for protocols aiming to enhance the recovery of hand function after SCI (Bunday and Perez, 2012). This may be particularly relevant for individuals with severe paralysis and weak residual control of hand muscles in whom more EMG activity could be engaged by rapidly responding to an acoustic startle stimulus during power grip compared with other more precise dexterous hand manipulations. Thus, our present results extend previous observations by showing that the reticulospinal tract in intact humans contributes to the control of hand muscles during tasks involving different degrees of hand dexterity (Carlsen et al., 2009; Honeycutt et al., 2013; Dean and Baker, 2017; Tazoe and Perez, 2017) and suggest reticulospinal outputs are linked to gross finger manipulations after SCI.
Footnotes
The authors declare no competing financial interests.
- Correspondence should be addressed to either of the following: Dr. Monica A. Perez, Department of Neurological Surgery, University of Miami, Miami, FL 33136, perezmo{at}miami.edu; or Dr. Stuart N. Baker, Institute of Neuroscience, Newcastle University, Newcastle upon Tyne NE2 4HH, UK, stuart.baker{at}newcastle.ac.uk





{kind=link}
{kind=link}
{kind=link}