

Abstract
Acid-sensing ion channel-1a (ASIC1a) contributes to multiple fear behaviors, however the site of ASIC1a action in behavior is not known. To explore a specific location of ASIC1a action, we expressed ASIC1a in the basolateral amygdala of ASIC1a–/– mice using viral vector-mediated gene transfer. This rescued context-dependent fear memory, but not the freezing deficit during training or the unconditioned fear response to predator odor. These data pinpoint the basolateral amygdala as the site where ASIC1a contributes to fear memory. They also discriminate fear memory from fear expressed during training and from unconditioned fear. Furthermore, this work illustrates a strategy for identifying discrete brain regions where specific genes contribute to complex behaviors.
- ASIC1a
- region-restricted
- viral-vector-mediated gene transfer
- fear conditioning
- unconditioned fear
- amygdala
Introduction
Acid-sensing ion channel-1 (ASIC1) is an acid-activated cation channel (Waldmann and Lazdunski, 1998; Krishtal, 2003; Wemmie et al., 2006). In central neurons, it contributes to synaptic plasticity (Wemmie et al., 2002; Zha et al., 2006) and is distributed to dendritic spines where it increases Ca2+ concentration in response to low pH (Zha et al., 2006). Mice lacking ASIC1a exhibit deficits in multiple fear behaviors including unconditioned fear (Coryell et al., 2007) and the acquisition and retention of conditioned fear (Wemmie et al., 2003; Coryell et al., 2007). Conversely, overexpressing ASIC1a throughout the brain in a transgenic mouse increased context fear conditioning (Wemmie et al., 2004). Consistent with a role in fear behavior, endogenous ASIC1a protein is robustly expressed in the basolateral amygdala (BLA) (Wemmie et al., 2003; Coryell et al., 2007). However, ASIC1a is also expressed in other regions of the fear circuit, as well as elsewhere in central and peripheral nervous systems. Thus, the specific sites of ASIC1a action in fear behavior remain unknown.
The abundant ASIC1a expression in the basolateral amygdala (BLA) (Wemmie et al., 2003; Coryell et al., 2007) and the importance of the BLA in fear conditioning (Kim and Jung, 2006) led us to hypothesize that the BLA is a key site of ASIC1a action in fear. To test this hypothesis, we examined c-Fos expression in the BLA following fear conditioning. We also developed an adeno-associated virus vector that allowed us to drive ASIC1a expression in restricted brain regions. Upon finding that ASIC1a disruption reduced c-Fos in the BLA, we examined the behavioral effects of bilaterally restoring ASIC1a expression to the BLA in ASIC1a−/− mice. We hypothesized that restoring ASIC1a would rescue context fear conditioning but not the unconditioned fear response to the predator odor trimethylthiazoline (TMT).
Materials and Methods
Mice.
ASIC1a−/− mice have been previously described (Wemmie et al., 2002). Briefly, the neomycin resistance gene replaced the first translated exon in the ASIC1a-encoding gene (ACCN2), eliminating expression of the ASIC1a protein. ASIC1a−/− and ASIC1a+/+ mice were maintained in a congenic C57BL/6 background. Experimental groups were matched for age (ranging from 12 to 16 weeks) and gender. Mice were kept on a standard 12 h light/dark cycle and received standard chow (LM-485; Teklab) and water ad libitum. Animal care met National Institutes of Health standards and all procedures were approved by the University of Iowa Animal Care and Use Committee.
c-Fos immunohistochemistry.
One hour following single-shock (1 s, 0.5 mA) context conditioning, mice were killed with halothane and perfused pericardially with PBS (Invitrogen) followed by 4% paraformaldehyde. Brains were extracted and processed for c-Fos immunohistochemistry as described (Coryell et al., 2007). c-Fos-positive nuclei were quantified by an investigator blinded to genotype using NIH ImageJ software as described previously (Coryell et al., 2007). The anatomical boundaries for the structures tested were defined based on the Paxinos mouse brain atlas (Paxinos and Franklin, 2001). Sections were obtained between 1.6 and 1.7 mm caudal to bregma.
Production of adeno-associated virus vectors.
Viral vectors were produced by the University of Iowa Gene Transfer Vector Core as described previously (Davidson et al., 2000). The vectors were adeno-associated virus 2/1 (AAV2/1) chimeric viruses with AAV1 capsids and AAV2 inverted terminal repeats and contained a CMV promoter driving either ASIC1a or enhanced green fluorescent protein (eGFP). The AAV-ASIC1a vector contained an internal ribosomal reentry site (IRES) followed by eGFP, downstream from ASIC1a.
Testing viral transduction and expression.
We transduced HT1080 cells (Stratagene) with AAV-ASIC1a or AAV-eGFP. We then lysed cells and performed Western blots to assess protein expression (supplemental Fig. S2, available at www.jneurosci.org as supplemental material). AAV-ASIC1a transduced cells showed ASIC1a expression in a dose-dependent manner. However, these cells expressed little if any eGFP from the IRES-eGFP cassette (data not shown). We also failed to detect any eGFP expression from brain regions injected with the AAV-ASIC1a vector. Therefore, to aid the mapping of AAV-ASIC1a transduced brain regions, we added AAV-eGFP to the AAV-ASIC1a injections. As a negative control, we also performed AAV-eGFP alone injections into the BLA.
Surgery and viral injection.
Mice were anesthetized and placed in a stereotaxic frame (Kopf). Viral vector injection was either a combination of 70% AAV-ASIC1a (titer 2.28 × 1013 viral genomes (vg)/ml) and 30% AAV-eGFP (titer 1.78 × 1014 vg/ml) or AAV-eGFP alone. Virus (0.5 μl) was infused using a 10 μl Hamilton syringe with a 33 gauge blunt tipped needle and a microinjector pump at a rate of 0.2 μl/min; the needle rested in position for 5 min postinjection. The injection coordinates were 1.5 mm posterior to bregma, 3.5 mm lateral to the midline, and 4.5 mm the from pia surface. Following injection, incisions were sutured and topical anesthetic was applied to the wound (Bupivicaine, 0.5%). Mice recovered for 14 d before behavioral analysis.
Tissue processing and target verification.
Following behavioral analysis, brains were freshly frozen on dry ice and embedded in OCT tissue freezing medium. Coronal slices (10 μm) were cut and mounted on slides using the CryoJane cryosectioning system (Electron Microscopy Systems). Slides were then either thawed for immediate eGFP detection and imaging or processed for immunohistochemical detection of ASIC1a as described previously (Coryell et al., 2007). Slides were coverslipped with Vectashield (Vector Laboratories) and visualized with a Zeiss epifluorescence microscope. Images were captured at 10× magnification and compiled using the tiling function to create composite images of coronal hemisections. Borders for the BLA were defined based on the anatomical landmarks in the Paxinos mouse brain atlas (Paxinos and Franklin, 2001). Correctly targeted injections (hits) were defined as having fluorescence above background within bilateral BLA. Off target injections (misses) were defined as having fluorescence above background in the temporal lobe but not in both BLA.
Context fear conditioning.
Mice were placed in a near-infrared video conditioning chamber (Med Associates) and allowed to habituate for 3 min. Five footshocks (1 s, 0.75 mA) were then delivered with an intershock interval of 1 min. Mice were returned to the home cage, and then, 24 h later, placed back into conditioning chamber for testing context-evoked freezing during a 5 min trial. Freezing was defined as an absence of movement other than respiration was assessed during training and testing sessions by automated video tracking software (Med Associates) which correlated closely with manual scoring.
TMT-evoked freezing.
TMT-evoked freezing was measured as described previously (Coryell et al., 2007). Briefly, mice were placed in a chamber with a beaker containing TMT (30 μl) (PheroTech). These behavioral chambers were distinct from the fear conditioning apparatus to avoid contaminating the fear conditioning equipment with TMT. Freezing was defined the same as for fear conditioning above and was scored from videotapes by an experimenter blinded to treatment. The TMT-evoked freezing data for the nonvirus injected ASIC1a+/+ and ASIC1a−/− mice were reported previously (Coryell et al., 2007). These large samples of mice were maintained on the same genetic background (C57BL/6) as the mice in this study and provided useful contrasts for virus-injected groups. To address the possibility that the TMT response might have changed, or that there might be some batch-specific effects we examined the unconditioned response in six ASIC1a+/+ mice from four different home cages. The mean TMT response was 93.8 s, very similar to the mean TMT-evoked response (92 s) for the ASIC1a+/+ mice reported previously and shown in Figure 2G.
Statistical analysis.
Values are expressed as mean ± SEM. For c-Fos expression, two-way ANOVA was used to test for main and interaction effects of genotype and conditioning. For behavioral analyses, one-way ANOVA was used to test for overall group effects. Planned contrast testing was used to test hypothesized relationships between groups (SPSS). For contrasts specified a priori, p values (two-tailed) < 0.05 were considered significant.
Results
We tested whether ASIC1a disruption in mice reduced c-Fos expression in the BLA following exposure to single-shock context fear conditioning. We found that conditioning increased c-Fos expression in the BLA, and that ASIC1a disruption reduced the number of c-Fos-positive cells after conditioning (Fig. 1A,B). In the amygdala, the ASIC1a effect was specific to the BLA, because c-Fos in the lateral, medial and central nuclei was normal in ASIC1a-null mice (supplemental Fig. S1, available at www.jneurosci.org as supplemental material). These results support the hypothesized effect of ASIC1a in the BLA, and bolstered our decision to target the BLA in subsequent experiments.

ASIC1a disruption attenuates c-Fos expression in the BLA. A, Representative images of c-Fos immunohistochemistry at baseline and 1 h after fear conditioning in ASIC1a+/+ and ASIC1a−/− mice. B, Analysis of c-Fos-positive cell density in the BLA. Two-way ANOVA revealed significant main effects of conditioning (p < 0.001, F(1,20) = 46.4) and genotype (p < 0.05, F(1,20) = 4.79), but not an interaction (F < 1; ASIC1a+/+ conditioned, n = 8; ASIC1a−/− conditioned, n = 7; ASIC1a+/+ unconditioned, n = 4; ASIC1a−/− unconditioned, n = 4). Planned contrast testing identified a reduction in c-Fos levels after conditioning in the ASIC1a−/− mice relative to ASIC1a+/+ controls (*p = 0.03, t(19) = –2.33).
To test the hypothesis that ASIC1a in the BLA is key for normal fear behavior, we produced an AAV-encoding ASIC1a (AAV-ASIC1a) (see Materials and Methods) (supplemental Fig. S2, available at www.jneurosci.org as supplemental material). We targeted ASIC1a to the BLA in ASIC1a-null mice; mice expressing ASIC1a bilaterally in the BLA were defined as hits (AAV-ASIC1a-hit) (Fig. 1C). Mice lacking ASIC1a expression in the BLA bilaterally were considered misses (AAV-ASIC1a-miss) (Fig. 1D) and were included in our behavioral analysis as a control for anatomic specificity. Mice injected with a similar vector expressing eGFP (AAV-eGFP) served as an additional negative control.
Two weeks following injection, we used a five-shock, context-fear-conditioning paradigm to examine fear behavior. As previously reported (Wemmie et al., 2003), uninjected ASIC1a-null mice froze less than wild-type mice during training (Fig. 2A,B). This deficit was not rescued in the AAV-ASIC1a-hit mice compared with the AAV-ASIC1a-miss, AAV-eGFP, and uninjected ASIC1a−/− controls.
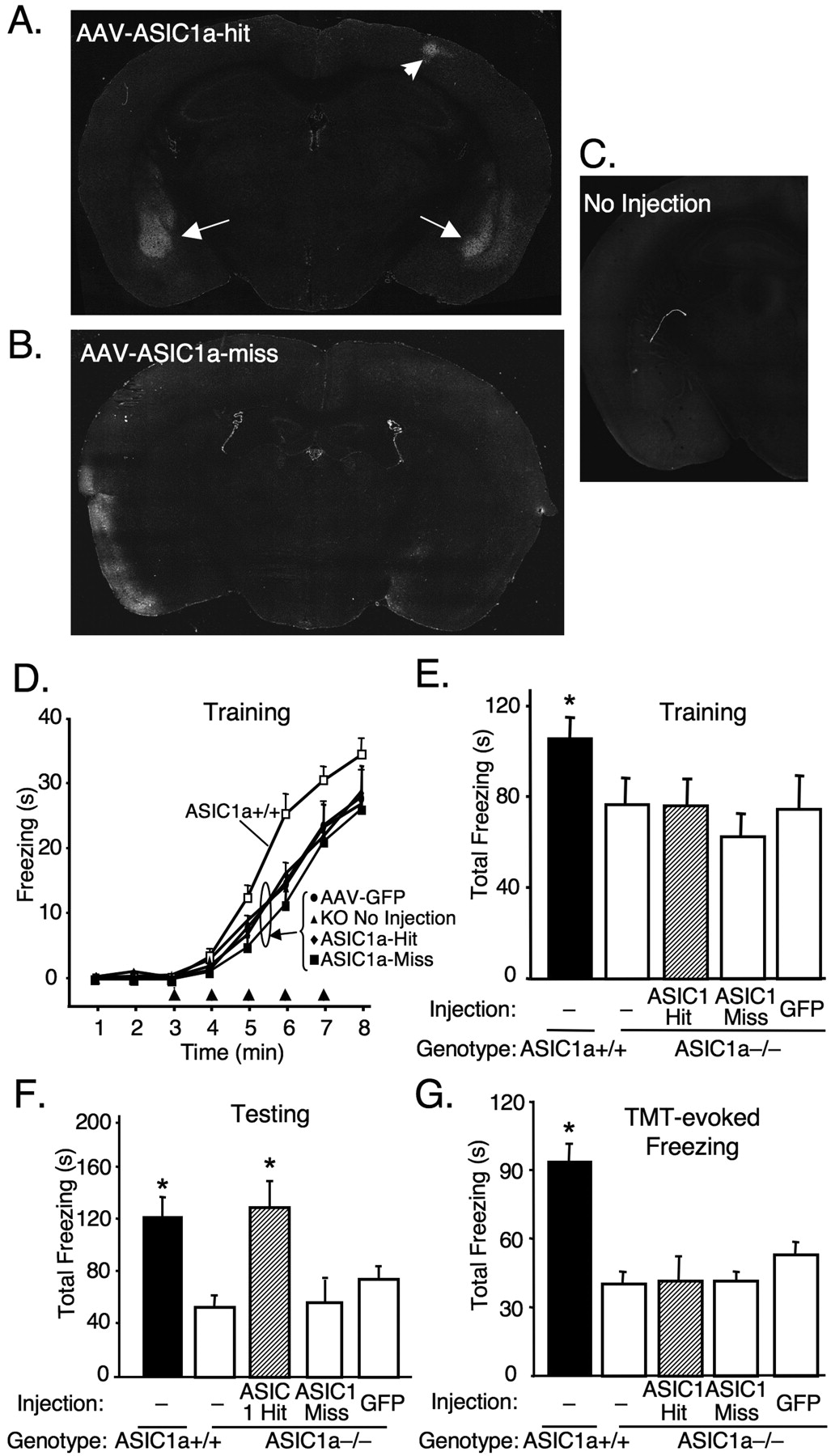
Restoring ASIC1a expression in the BLA of ASIC1a-null mice rescues deficits in conditioned freezing but not unconditioned freezing during training or in response to TMT. A, B, Representative examples of ASIC1a immunohistochemistry in ASIC1a−/− mouse with an AAV-ASIC1a injection that targeted ASIC1a expression to the BLA (arrows) bilaterally (AAV-ASIC1a-hit) and an injection that missed the BLA bilaterally (AAV-ASIC1a-miss). Small area of ASIC1a antibody labeling in right sensory/motor cortex (AAV-ASIC1a-hit) (arrowhead) is at the site of needle entry. C, Example of ASIC1a immunohistochemistry in uninjected ASIC1a−/− mouse. D, E, Freezing during context fear training with five shocks (arrowheads). One-way ANOVA revealed a significant effect of group (p < 0.05, F(4,65) = 2.55; ASIC1a+/+, n = 16; ASIC1a−/− no injection, n = 21, AAV-ASIC1a-hit, n = 12; AAV-ASIC1a-miss, n = 7; AAV-eGFP, n = 14). In addition, contrast testing revealed more freezing in the ASIC1a+/+ group versus the ASIC1a−/− groups (*p = 0.003, t(65) = 3.04). However, bilateral ASIC1a expression produced no effect in the ASIC1a−/− mice (ASIC1a-hit) relative to the other ASIC1a−/− groups (p = 0.345, t(65) = –0.95). F, Conditioned freezing assessed 24 h after training during a 5 min testing session. One-way ANOVA revealed a significant effect of group (p < 0.01, F(4,65) = 4.9). In striking contrast to the training results, restoring ASIC1a to the amygdala (ASIC1a-hit) increased conditioned freezing relative to the other ASIC1a−/− groups (*p = 0.005, t(65) = 3.58), and to levels that did not differ significantly from the ASIC1a+/+ controls (p = 0.66, t(65) = 0.441). As with previous results (Wemmie et al., 2003), conditioned freezing was significantly greater in wild-type mice relative to uninjected ASIC1a−/− mice (*p = 0.004, t(65) = 2.99). G, ASIC1a expression in the BLA did not rescue the deficit in unconditioned freezing to the predator odor TMT during a 5 min exposure. One-way ANOVA revealed a significant group effect (p < 0.001, F(4,76) = 11.78; ASIC1a+/+, n = 33; ASIC1a−/− no injection, n = 31; AAV-ASIC1a-hit, n = 5; AAV-ASIC1a-miss, n = 4; AAV-eGFP, n = 8). ASIC1a+/+ mice froze significantly more than the ASIC1a−/− groups (*p < 0.001, t(76) = –5.98). However, restoring ASIC1a to the amygdala (ASIC1a-hit) did not affect the TMT response relative to the other ASIC1a−/− groups (p = 0.860, t(76) = 0.178). TMT-evoked freezing data for the uninjected ASIC1a+/+ mice and ASIC1a−/− mice were reported previously (Coryell et al., 2007) (see Materials and Methods).
The following day, we assessed the context-dependent fear response by returning the mice to the conditioning chamber for 5 min (minus footshocks). Strikingly, injection of AAV-ASIC1a in the BLA increased freezing relative to the other ASIC1a−/− groups and restored freezing to levels observed in wild-type mice (Fig. 2C). Together these data suggest that ASIC1a expression in the BLA is sufficient to rescue the deficit in conditioned fear memory in ASIC1a-null mice, but not the normal freezing response to footshock during training.
The failure of BLA ASIC1a to rescue the ASIC1a−/− freezing deficit during training raised the question of whether it would rescue unconditioned fear (Rosen, 2004). ASIC1a-null mice exhibit deficits in unconditioned fear responses to the predator odor trimethylthiazoline (TMT) (Coryell et al., 2007),. Unlike conditioned fear, such unconditioned fear behaviors are thought to depend more on the bed nucleus of the stria terminalis than the BLA (Walker et al., 2003; Rosen, 2004; Fendt et al., 2005), although the BLA may play a role (Müller and Fendt, 2006). Therefore, we hypothesized that expressing ASIC1a in the BLA would have little effect on TMT-evoked freezing in the ASIC1a-null mice. Indeed, the ASIC1a-null mice with ASIC1a targeted to the BLA did not differ from the other ASIC1a-null groups (Fig. 2D).
Discussion
Together these data identify the BLA as a key site of ASIC1a action in fear memory. In contrast, ASIC1a in the BLA failed to rescue the freezing deficit during training and the unconditioned freezing responses to TMT. Thus, these two latter fear responses in wild-type mice likely depend on ASIC1a in other brain regions. The literature suggests several sites where ASIC1a might mediate its effects on unconditioned fear of predator odor. The bed nucleus of the stria terminalis (BNST) is an example; BNST manipulations disrupt TMT-evoked fear (Fendt et al., 2003) and ASIC1a is abundant in the BNST (Coryell et al., 2007). The medial amygdala and dorsal periaqueductal gray are also potential sites of ASIC1a action in predator odor evoked fear; disrupting ASIC1a altered c-Fos induction by TMT at these locations (Coryell et al., 2007). It will be interesting to learn whether ASIC1a at these sites affects unconditioned fear responses to predator odor and also whether it affects fear conditioning.
Although linked, the relationship between the freezing during training and the development and expression of long-lasting fear memory is not clear. Previous studies indicate that inactivating or lesioning the BLA blocks both the freezing response that follows footshocks during training and also the expression of context fear memory (Rosen, 2004; Kim and Jung, 2006). Moreover, the freezing during training has been convincingly argued to be learning-dependent (Fanselow, 2000; Rudy et al., 2004). Therefore, we were surprised when the ASIC1a-null mice with ASIC1a in the BLA showed normal fear memory despite an apparent learning deficit. One potential explanation is that ASIC1a rescued memory consolidation, a plausible speculation given the role of ASIC1a in synaptic plasticity (Wemmie et al., 2002; Zha et al., 2006). However, an effect on the recollection and expression of fear memory cannot be excluded from these data. Injecting AAV-ASIC1a or ASIC1a antagonists into the BLA after memory consolidation may help discriminate between consolidation and recollection. Even so, the present data support the idea that the response to footshock during training and the resulting memory are separable. Moreover, they indicate that different ASIC1a-dependent pathways mediate the two processes.
Previous studies suggest that different cell populations in the lateral amygdala may mediate short versus long-lived responses to fear conditioning (Repa et al., 2001). Thus, delineating the effects of ASICs on different amygdala neuron types could be valuable for clarifying ASIC's role in fear during training and long-lasting fear memory. Nevertheless, because the responses are separable, these data suggest that the strength of a traumatic memory could be more severe than what might be predicted from the initial fearful experience. This result could have implications for anxiety disorders such as posttraumatic stress disorder, where only a subset of individuals who experience a similar trauma go on to develop the disorder.
Region-specific gene disruption and expression have been powerful tools for probing the relationship between genes, brain function, and behavior. Yet in many cases, the genetic tools required are either not available or can take substantial time to generate. Our data illustrate the value of an alternate approach using localized viral vector-mediated gene expression in a mouse with a global gene disruption. This strategy allowed us to pinpoint a site of ASIC1a action in context-dependent fear memory. We speculate that this approach may also be of value in exploring the role that other genes and discrete brain regions play in complex neuronal processes.
Footnotes
-
This work was supported by National Institutes of Health Grant F31MH076414-03 (M.W.C.), and National Alliance for Research on Schizophrenia and Depression Young Investigator Award and Department of Veterans Affairs Merit Review Award (J.A.W.). We acknowledge Maria Scheel of the University of Iowa Gene Transfer Vector Core for production of AAV vectors. We thank Dr. Douglas Langbehn for statistical assistance.
- Correspondence should be addressed to Dr. John A. Wemmie, Department of Psychiatry, University of Iowa College of Medicine, 3139 Medical Laboratories, Iowa City, IA 52242. john-wemmie{at}uiowa.edu





{kind=link}
{kind=link}