

Abstract
The temporal structuring of multiple events is essential for the purposeful regulation of behavior. We investigated the role of the lateral prefrontal cortex (LPFC) in transforming external signals of multiple sensory modalities into information suitable for monitoring successive events across behavioral phases until an intended action is prompted and then initiated. We trained monkeys to receive a succession of 1 s visual, auditory, or tactile sensory signals separated by variable intervals and to then release a key as soon as the fourth signal appeared. Thus, the animals had to monitor and update information about the progress of the task upon receiving each signal preceding the key release in response to the fourth signal. We found that the initial, short-latency responses of LPFC neurons reflected primarily the sensory modality, rather than the phase or progress of the task. However, a task phase-selective response developed within 500 ms of signal reception, and information about the task phase was maintained throughout the presentation of successive cues. The task phase-selective activity was updated with the appearance of each cue until the planned action was initiated. The phase-selective activity of individual neurons reflected not merely a particular phase of the task but also multiple successive phases. Furthermore, we found combined representations of task phase and sensory modality in the activity of individual LPFC neurons. These properties suggest how information representing multiple phases of behavioral events develops in the LPFC to provide a basis for the temporal regulation of behavior.
Introduction
The lateral prefrontal cortex (LPFC) possesses rich connections with posterior association areas (Barbas and Mesulam, 1985; Petrides and Pandya, 1999, 2002; Romanski et al., 1999b) and collects information about and controls motor behavior through interconnections with higher-order motor areas (Bates and Goldman-Rakic, 1993; Lu et al., 1994; Takada et al., 2004). This anatomical connectivity has been proposed as the means by which LPFC regulates the behavior according to diverse signals across sensory and motor domains (Wise and Murray, 2000; Miller and Cohen, 2001; Tanji and Hoshi, 2008).
Moreover, the LPFC is thought to be essential for temporal structuring of behavior. Patients with LPFC lesions experience difficulties in serially organizing multiple actions (Shallice and Burgess, 1991) and judging temporal relationships among multiple events (Milner, 1982; Shimamura et al., 1990; McAndrews and Milner, 1991). Monkeys with lesions in the mid-dorsal LPFC have shown deficits in performing tasks requiring the organization of the serial order of multiple items (Petrides, 1991a,b, 1995). Functional imaging studies in humans have shown LPFC activity in processing information about the temporal aspects of multiple events (Petrides, 1991a, 1995; Pochon et al., 2001; Konishi et al., 2002; Knutson et al., 2004; Amiez and Petrides, 2007). In line with these reports, LPFC neurons have been revealed to encode ordinal or rank position in a variety of visually guided motor behavior (Fujii and Graybiel, 2003; Hasegawa et al., 2004; Sigala et al., 2008; Berdyyeva and Olson, 2010). Furthermore, LPFC neurons are known to reflect specific combinations of spatial or object information and rank position (Barone and Joseph, 1989; Ninokura et al., 2004), information about forthcoming sequential movements (Averbeck et al., 2002, 2006), and multiple steps in future action plans (Mushiake et al., 2006; Shima et al., 2007).
However, it remained unclear how individual LPFC neurons participate in processing a series of sensory signals for the purpose of structuring behavioral phases. Accordingly, we conducted this study to explore the role of LPFC in monitoring the occurrences of sensory signals of multiple modalities and in generating temporal information necessary to construct behavioral phases structured to generate a motor behavior at an appropriate timing. We intended to ask the following specific questions. (1) Is the behavioral-phase information generated in individual neurons in a modality-specific manner or commonly, regardless of sensory modalities? (2) What is the time relation between the occurrences of modality-selective sensory responses and the behavioral phase-selective activity? (3) Do individual LPFC neurons represent a single task phase to be linked in a chain-like manner, or does each neuron represent multiple task phases in combination? (4) Are entire multiple phases continuously represented in single neurons, e.g., in a continuous fashion? We will show that LPFC neurons represent rich information related to the reception of sensory signals and the monitoring of task sequences consisting of multiple phases before prompting the initiation of action. LPFC neurons initially represent modality-selective sensory signals but soon come to reflect task phase, ultimately encoding multiple phases of behavior as a whole in a multidimensional manner.
Materials and Methods
Animals and experimental conditions.
We used two monkeys (Macaca fuscata; one male weighing 7.0 kg and one female weighing 5.0 kg) who were cared for in accordance with the National Institutes of Health guidelines and the guidelines of Tamagawa University. During the experimental sessions, each monkey sat in a chair with her or his head and left arm restrained. We installed a button in front of the chair at waist level; the monkey pushed and released the button with his or her right hand. A 19 inch color video monitor was placed in front of the monkey (25 cm from the eyes) to display a fixation point (white square, 1.4° visual angle) and a visual cue (white square 10° × 10° superimposed on the fixation point). Vibrotactile sinusoidal stimuli (20 Hz) were delivered to the surface of the palm of the left hand by a mechanical stimulator (DIA Medical System). Two speakers placed bilaterally in front of the monkey produced a pure tone of 500 Hz (60 dB), serving as an auditory cue. Eye positions were monitored at 120 Hz with an infrared eye-tracking system (resolution, 0.25° visual angle; RHS-M, Applied Science Laboratories). The TEMPONET system (Reflective Computing) was used to control the behavioral task and save data for offline analyses at 1000 Hz.
Behavioral task.
We devised a behavioral task that involved the monkeys' receiving a series of sensory signals in a single modality (visual, auditory, or tactile) and responding by releasing a button when a fourth signal appeared (Fig. 1A). After an intertrial interval of 2500–4500 ms, a trial commenced when the monkey pushed the button with the right hand and gazed at the fixation point that appeared at the center of the screen. When the monkey maintained fixation on the fixation point for ≥600 ms, the first cue was presented for 1000 ms and was followed by a randomly selected delay period, during which only the fixation point was visible. Subsequently, the second, third, and fourth cues were presented for 1000 ms each after a delay period. During the intertrial interval, the computer controlling the behavioral task randomly chose minimum durations of the first, second, and third delay periods in the next trial from values ranging from 700 to 2400 ms. The second, third, or fourth cue was presented if the monkey had already fulfilled the fixation requirement of ≥ 600 ms at the end of the minimum delay period. If not, the cue presentation was postponed until the fixation requirement was satisfied. Thus, the lower limit of the actual delay period was 700 ms, and its upper limit could take any value. The actual durations of the delay period during the task performance were widely distributed between 700 and 2400 ms in ≥ 88% cases in both monkeys, and the delay period was >2400 ms in the remaining cases. Thus, we designed the task to progress when the monkey gazed at the fixation point for ≥ 600 ms immediately before the onset of each cue, which ensured that the gaze was maintained on the fixation point when each cue was delivered. If the monkey released the button while the fourth cue was presented for 1000 ms, the cue terminated immediately, and a drop of liquid reward was delivered 500 ms later. A visual, auditory, or tactile stimulus was used as a cue in each trial. The sensory modality used as a cue was altered every five trials. If the monkey released the button before the fourth cue appeared (premature release error) or if the monkey failed to release the button while the fourth cue was presented (belated error), the trial was terminated as an error trial and an intertrial interval was initiated.
Surgery and physiological recording.
After completing the behavioral training, aseptic surgery was performed under pentobarbital sodium anesthesia (25 mg/kg i.v.) with atropine sulfate. Antibiotics and analgesics were used to prevent postsurgical infection and pain. Polycarbonate screws were implanted in the skull, and two plastic pipes, rigidly attached with acrylic resin, were used to securely fix the head during the daily recording session. A part of the skull over LPFC was removed, and a recording chamber was implanted. Neuronal activity was recorded with glass-insulated Elgiloy alloy microelectrodes (0.5–2.0 MΩ at 1 kHz) that were inserted through the dura mater using a hydraulic microdrive (MO-81-S, Narishige). Single-unit potentials were amplified with a multichannel processor and sorted using a multispike detector (MCP Plus 8 and ASD, Alpha Omega Engineering). We recorded neuronal activity from LPFC in the right hemisphere (Fig. 1B). Recording sites, first determined by observing the principal and arcuate sulci during craniotomy, were verified by examining magnetic resonance images (1.5 T; Sonata, Siemens) taken after the recording chamber was implanted (Fig. 1C–F). The recording sites were rostral to the frontal eye field, where saccadic eye movements were evoked by intracortical microstimulation applied through the tip of an inserted electrode (11 cathode pulses; 200 μs pulse width at 333 Hz; current, <50 μA; SS-203J, SEN-3401, Nihon Kohden).
Data analyses.
To detect task-related neuronal activity, we initially counted the number of spikes in 10 successive 200 ms bins before and after the onset of each of the four sensory cues (five bins before and five bins after the onset of each cue), yielding an array of firing rates consisting of 40 bins (i.e., 10 × 4 cues) for the trials in each modality. The array of the trials in each sensory modality was constructed separately. We defined a neuron as “task-related” if the distribution of discharge rates differed significantly in at least one of the three sensory modalities across the 40 bins (one-way ANOVA, p < 0.01, not corrected).
Two-way ANOVA.
To analyze the response selectivity of the task-related neurons, we used a two-way ANOVA (α = 0.01) with two main factors: the sensory “modality” (visual, auditory, and tactile) and the “phase” position (first, second, third, or fourth cue). We conducted separate analyses for the activity in two periods: 1) the cue period (from 50 to 550 ms after cue onset) and 2) the delay period (the 500 ms period before cue onset). Based on the results of the two-way ANOVA, we classified neuronal selectivity into four categories: selective for modality (modality < 0.01, phase > 0.01, modality*phase > 0.01), selective for phase (modality > 0.01, phase < 0.01, modality*phase > 0.01), selective for both (modality > 0.01 and phase > 0.01, or modality*phase < 0.01), and selective for neither (modality > 0.01, phase > 0.01, modality*phase > 0.01). We performed Tukey's HSD (p < 0.01) test on modality-selective neurons to examine responses to the three sensory modalities. If activity was greater in one modality than in the other two modalities, the neuron was classified as a unimodal neuron. Using Tukey's HSD (p < 0.01) test with four elements (first cue, second cue, third cue, and fourth cue) on phase-selective neurons, we classified neurons as phase selective for a single phase (activity was greater in one phase than in the remaining three), selective for two successive phases (activity was greater in two successive phases than in the remaining two), or selective for three successive phases (activity was greater in three successive phases than in the remaining one).
Early cue–response.
To detect the activity of neurons that responded to the onset of the cue (i.e., the early cue response), we initially counted the number of spikes in every 20 ms bin during the 200 ms period before cue onset (10 bins in total) and in the 220 ms period after cue onset (11 bins in total). We subsequently compared the distributions of firing rates for each 20 ms bin after cue onset with those of the 10 bins before cue onset using a Mann–Whitney U test. When the activity after cue onset initially became significantly greater than that during the precue period (p < 0.01) in two consecutive bins, response latency was determined by observing the central position of the first bin. This analysis was conducted separately for trials in each sensory modality.
Detection of activity latency.
To uniformly determine the latency of the activity modulation for population of neurons, as well as for each neuron, we applied an analysis different from that described above. We sought the initial point in time during the 20 ms period when the spike density function (smoothed with a Gaussian kernel, σ = 10 ms) exceeded a threshold, defined as the baseline activity (the 100 ms period before cue onset) plus half the maximum modulation above this value during cue presentation. We judged that this method would provide a more reliable measure, because the number of neurons in a group does not affect the sensitivity in detecting the activity latency.
Results
We trained two monkeys to release a button with their right hands when they received the fourth in a series of visual, auditory, or tactile sensory cues (Fig. 1A). Because each cue was presented for 1 s, the animals were required to release the button within 1 s after the onset of the fourth cue. The animals achieved success rates of 95.0% (Monkey 1) and 92.4% (Monkey 2). Analysis of the reaction times for the button release in successful trials revealed that the animals released the button promptly in response to the appearance of the fourth cue (Table 1). We subsequently investigated the extent to which the monkeys' performances were influenced by the total duration of the delays. First, we examined the effect of the interval between the first cue onset and the fourth cue onset on the reaction times in successful trials (Fig. 1G,H). We found that the effect, if any, was minimal because the slope of the regression line was <±10 ms per second. (If the monkeys had employed the timing strategy, the slope would have been close to −1000 ms per second.) Second, we asked whether the tendency for the animal to release the button prematurely in response to the third cue was related to the interval between the onsets of the first cue and the third cue. By examining the occurrences of the premature errors against the intervals, we found that the success rate was >90% at any interval in both monkeys. Altogether, these results indicated that the monkeys did not employ the timing strategy to perform the task, whereas they relied on the appearance of the four sensory signals.

The temporal sequence of behavioral events, the recording site, and the behavior of monkeys. A, The top, middle, and bottom rows indicate the sequence of visual, auditory, and tactile trials, respectively. Signals for a single modality were presented in each trial, and the sensory modality was altered after a block of five trials. A trial commenced when the monkey pressed the button with the right hand and gazed at the fixation point for >0.6 s. Subsequently, a series of sensory signals of 1 s duration were delivered, separated by interstimulus intervals of 0.7–2.4 s. The animals were required to gaze at the fixation point for >0.6 s before the onset of the second, third, and fourth cues. The animals were rewarded if they released the button within 1.0 s after the appearance of the fourth cue. B, Lateral view of the frontal cortex. Electrode penetrations covered the cortical area displayed with each shaded area (the lateral prefrontal cortex, LPFC, of the right hemisphere). AS, Arculate sulcus; CS, central sulcus; PS, principal sulcus. C–F, Magnetic resonance images. Scale bars, 10 mm. C, D, Images showing coronal slices at the levels of the rostral (C) and caudal LPFC (D). E, F, Images showing the lateral surface of the cerebral cortex of Monkey 1 (E) and Monkey 2 (F). G, H, Relationship of the response time to the interval between the first cue onset and the fourth cue onset for Monkey 1 (G) and Monkey 2 (H). From left to right, data of visual, auditory, and tactile trials in randomly sampled 1000 successful trials are shown. At the bottom of each panel, the slope of the regression line is shown.
Reaction times for button release after the onset of the fourth cue
While the monkeys performed the behavioral task, we recorded the neuronal activity in the LPFC of the right hemisphere (Fig. 1B–F) opposite to the left hand, to which the vibrotactile stimuli were delivered, and ipsilateral to the right hand, with which the animal pressed and released the button. We found task-related activity in 1659 LPFC neurons (n = 1110 in Monkey 1 and 549 in Monkey 2). The following three sections will describe the response properties of neurons in the LPFC. First, we will focus on the retrieval and maintenance of sensory signals in each modality and show that the distribution of unimodal neurons after cue onset was consistent with the anatomical organization of the prefrontal connections with the sensory association areas. Second, we will show three types of task phase representations in the LPFC that demonstrated robust and continuous representations of the whole task. Third, we will show that the arrival of the sensory signal preceded the rapid development of the task phase coding that was used for the timely initiation of the behavioral response.
Retrieval and maintenance of sensory signals
To investigate the retrieval and maintenance of sensory signals in each modality, we applied two analyses in parallel. The first analysis examined the timing when LPFC neurons began to reflect the onset of sensory signals. We focused on the activity in the short period after the first cue onset (≤ 200 ms). The activity that began in this period was called “early cue responses” and was considered to reflect the initial retrieval of sensory signals by LPFC neurons. We analyzed the activity with the temporal resolution of 20 ms (Yamagata et al., 2009). The 20 ms bins were used because the resolution was high enough while it was possible to reliably detect the activity onset even in neurons with responses of fairly short duration. The second analysis examined the representations of sensory signals and task phases during the cue and delay periods. Because the monkeys gazed at the fixation point for ≥600 ms immediately before the onset of each cue, we chose the last 500 ms of each delay period as an analysis window of the “delay period” activity. To directly compare the selectivity between the cue period and the delay period in each neuron, another 500 ms window (50–550 ms after the onset of each cue) was used for the analysis of the cue period activity. We will describe results of the two analyses in the following sections.
Selectivity for modality versus task phase in early cue responses
We first addressed neuronal activity that responded with short latency to the cue onsets. Because we noticed that the duration of the auditory sensory response is fairly short, here we used an analysis suitable for detecting the onset of neuronal activity (see Materials and Methods for the definition of the early cue response). We found that 291 neurons exhibited early responses to the appearance of the first cue. Three examples of early cue responses are illustrated in Figure 2A–C. This figure shows that a majority (84%, n = 243) of early cue responses were unimodal, responding selectively to the appearance of a visual (n = 130, Fig. 2A), auditory (n = 44, Fig. 2B), or tactile (n = 69, Fig. 2C) cue. The population-level activity of each group of unimodal neurons is illustrated in Figure 2D–F. Thirty-nine of the remaining neurons responded to two sensory modalities, and nine responded to three sensory modalities.

Early cue-responses. A–C, Three examples of neurons with early cue responses shown in A (visual neuron), B (auditory neuron), and C (tactile neuron). Rasters and spike–density functions (smoothed using a Gaussian kernel; σ = 10 ms) indicate activity sorted according to the onset of the first, second, third, and fourth cues. The red-, green-, and blue-colored raster displays and spike–density functions represent the activities in the visual, auditory, and tactile trials, respectively. The ordinate represents the instantaneous firing rate (in spikes/s). The gray area indicates the 1 s period during which each cue was presented. Because the fourth cue was terminated immediately after the animal released the button, the gray area of the fourth cue was drawn based on the average reaction time. The reward was delivered for about 100 ms at 500 ms after button release. The tick marks on the horizontal axis are placed at 1 s intervals. D–F, Population-level activities of neurons with early cue responses after the appearance of cues. Red line, Responses in visual trials; green line, responses in auditory trials; blue line, responses in tactile trials. D, Activity of visual neurons (n = 130). E, Activity of auditory neurons (n = 44). F, Activity of tactile neurons (n = 69). D′–F′, Normalized population-level responses to the second, third, and fourth cues were calculated with reference to the activity after the first cue in each group of neurons (mean ± SEM). D′, Visual neurons (n = 130). E′, Auditory neurons (n = 44). F′, Tactile neurons (n = 69). Asterisk (*) indicates activity significantly differed from that after the onset of the first cue (paired t test, p < 0.01). G–I, The spatial distribution of visual (G, n = 130), auditory (H, n = 44), and tactile neurons (I, n = 69) are displayed by overlaying the data from the two animals with reference to the principal sulcus and its caudal end. Each dot indicates a neuron. Electrode penetrations covered the cortical area displayed with each shaded area. The asterisk (*) indicates that the tactile neurons were clustered differently from the visual and auditory neurons (MANOVA, p < 0.01).
We examined the task phase selectivity of these early cue responses. For this purpose, we first calculated the population-level activity of each group of unimodal neurons. The magnitude of the responses was quantified by calculating each neuron's normalized responses to the second, third, and fourth cues relative to their responses to the first cue using a measure of the maximum discharge rate during the 200 ms period after onset of the cues. We found that the magnitude of activity after the onset of the second to fourth cues did not differ from that after the onset of the first cue in visual (Fig. 2D′), auditory (Fig. 2E′), and tactile neurons (Fig. 2F′) (paired t test, p ≥ 0.01), with the exception of the responses of the tactile neurons to the third cue (Fig. 2F′). We then studied the task phase selectivity of bimodal and trimodal neurons by applying the same analysis to the early cue response to the appearance of each sensory modality. In both bimodal (n = 39) and trimodal neurons (n = 9), the magnitude of the activity after the onset of the second to fourth cues did not differ from that of the activity after the onset of the first cue (paired t test, p > 0.3). These findings indicated that the short latency responses to sensory cues were, in general, not selective for the task phase.
The spatial distributions of neurons with visual, auditory, and tactile early cue responses are illustrated in Figure 2G–I. The visual (Fig. 2G) and auditory neurons (Fig. 2H) were widely distributed in the caudal sector of the LPFC. By contrast, the tactile neurons (Fig. 2I) were more often found in the ventrorostral region. We found that the tactile neurons (Fig. 2I) were clustered differently from the visual and auditory neurons [The multiple ANOVA (MANOVA) with two dependent variables of anterior–posterior and medial–lateral positions of each neuron and a single independent variable of the selectivity, p < 0.01]. In the area ventral to the principal sulcus, the tactile neurons were located more rostral than were the visual neurons (Kolmogorov–Smirnov Test applied to the distribution function about the anterior–posterior position of the neurons, p < 0.01).
Modality-selective responses during the cue and delay periods
In the previous section, we focused on the onset of the neuronal activity after the appearance of the sensory cue. Separately from that analysis, here we investigated the representation of the sensory–modality selectivity and the task phase selectivity. For this purpose, the two-way ANOVA was used to identify activity selective for the task phase and/or the sensory modality (see Materials and Methods). This analysis allowed us to systematically analyze the effects of multiple behavioral factors on the neuronal activity. The activity was separately analyzed during the cue period (50–550 ms after cue onset) and the delay-period (500 ms before cue onset). The time window of the delay period was chosen because we wanted to detect the sustained representation of the task phase between successive cues. Although the activity before the appearance of the first cue corresponds to the “anticipatory” activity (Mauritz and Wise, 1986), here we used the term “pre-first cue delay” because the activity may encode the task phase (i.e., the period preceding the first cue appearance), given that the monkeys gazed at the fixation point waiting for the cue appearance as in other delay periods.
During the cue period, 18% (n = 303) of the neurons showed selectivity only for the sensory modality, whereas 14% (n = 224) showed selectivity for this modality only during the delay period (Table 2). Fewer neurons were selective for modality during the delay than during the cue period (χ2 test, p < 0.0003). Of the 303 neurons selective for the sensory modality only during the cue period, 94, 9, and 19 were found to be significantly more active in the visual, auditory, and tactile trials, respectively, than in trials with the remaining two modalities (Tukey's HSD test, p < 0.01). The population-level activity of each group of neurons, shown in Figure 3A (visual neurons) and Figure 3C (tactile neurons), revealed that these neurons responded to the appearance of a specific sensory modality. By contrast, the auditory neurons (Fig. 3B) seemed not to show phasic responses. However, this apparent unresponsiveness was attributable to our inability to detect the activity within the time window of 500 ms given that the duration of the sensory responses of the auditory neurons was generally very short, as exemplified in Figure 2, B and E. This contrast in response duration suggests that the mode of information encoding used by auditory versus visual or tactile neurons differs.
Distribution of response selectivity classified by ANOVA

Population-level activities and normalized responses of unimodal neurons. A–C, Population-level activities of neurons with unimodal responses in the cue period. Activities are presented separately for visual neurons (A, n = 94), auditory neurons (B, n = 9), and tactile neurons (C, n = 19). The red-, green-, and blue-colored spike–density functions (smoothed using a Gaussian kernel; σ = 10 ms) represent activities in the visual, auditory, and tactile trials, respectively. The ordinate represents the instantaneous firing rate (in spikes/s). The gray area of the first to third cues indicates the 1 s period during which each cue was presented. That of the fourth cue was trimmed at the averaged response time of the animals. The reward was delivered 500 ms after button release. The tick marks on the horizontal axis are placed at 1 s intervals. The horizontal bars at the bottom of each panel indicate the cue periods (50–550 ms after cue onset). D–F, Population-level activities of neurons with unimodal responses in the delay period. The figures are displayed as in (A–C). Activities are presented separately for visual neurons (D, n = 26), auditory neurons (E, n = 10), and tactile neurons (F, n = 11). The horizontal bars at the bottom of each panel indicate the delay periods (500 ms before cue onset). A′-C′, Normalized activity (± SEM) of the unimodal neurons in the cue period. Activities of neurons preferring the visual cue (A′), the auditory cue (B′), and the tactile cue (C′) during the cue period are displayed separately. Red, Visual trials; green, auditory trials; blue, tactile trials. D′-F′, Normalized activity (± SEM) of the unimodal neurons in the delay period. Activity of neurons preferring the visual cue (D′), the auditory cue (E′), and the tactile cue (F′) in the delay period are displayed separately. Red, Visual trials; green, auditory trials; blue, tactile trials.
To examine the magnitude of selectivity at the population level, the activity of each unimodal neuron was normalized with respect to the activity in the modality and phase with the highest discharge rates, which were subsequently averaged across neurons in each group. The results of these analyses are shown in Figure 3A′ (visual neurons), Figure 3B′ (auditory neurons), and Figure 3C′ (tactile neurons), which show that activity in the preferred sensory modality was twofold greater than that in the other modalities.
During the delay period, 26, 10, and 11 neurons of the 224 modality-selective neurons were more active in the visual, auditory, and tactile trials, respectively (Tukey's HSD test, p < 0.01). The population-level activity of each group of neurons is shown in Figure 3D (visual neurons), Figure 3E (auditory neurons), and Figure 3F (tactile neurons), and the degree of population-level activity is shown in Figure 3D′ (visual neurons), Figure 3E′ (auditory neurons), and Figure 3F′ (tactile neurons), which again reveal that activity was 1.5- to 2-fold greater in the preferred sensory modality.
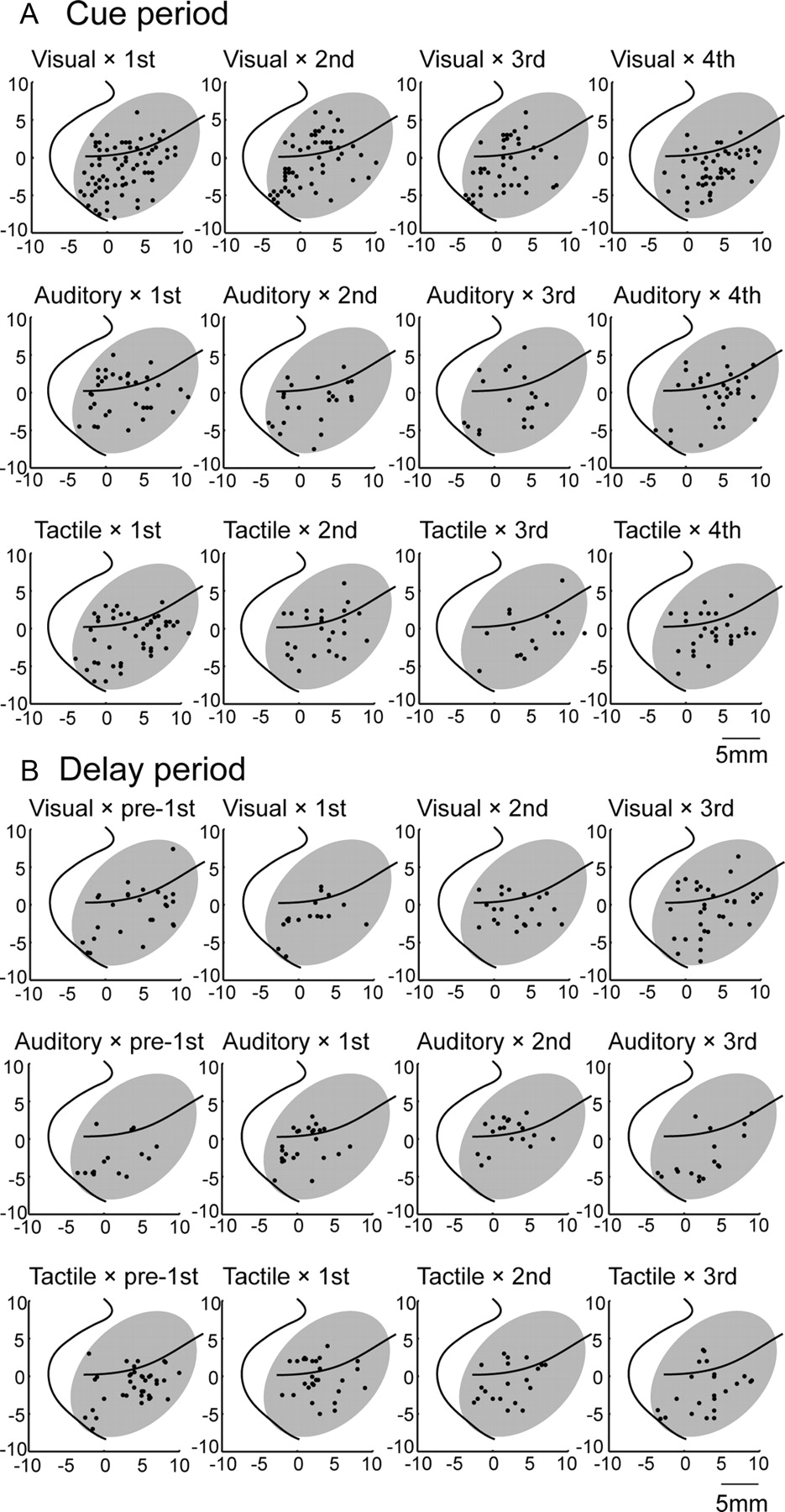
Neurons that were modality selective in the cue (Fig. 4A) or delay (Fig. 4B) period were distributed widely in the LPFC, with a tendency for tactile-selective neurons to be located more anterior in the area ventral to the principal sulcus. In the cue period (Fig. 4A), the tactile neurons were clustered differently from the visual and auditory neurons (MANOVA, p < 0.01), with the tactile neurons located more rostral than the visual neurons in the area ventral to the principal sulcus (Kolmogorov–Smirnov test, p < 0.01).

A, B, Cortical surface map displaying the distribution of neurons with unimodal responses. The top row (A) shows the distribution of unimodal neurons in the cue period. The asterisk indicates that the tactile neurons were clustered differently from the visual and auditory neurons (MANOVA, p < 0.01). The bottom row (B) shows the distribution of unimodal neurons in the delay period. Each panel shows the distributions of visual, auditory, and tactile neurons, respectively, from left to right. Data from the two animals were overlaid according to the principal sulcus and its caudal end. Electrode penetrations covered the cortical area displayed with each shaded area.
The analyses described above revealed that subsets of LPFC neurons promptly retrieved the sensory signals after cue onset and then maintained the sensory information during the delay period. In this study, we further found that LPFC neurons encoded much information concerning the task phase in addition to that concerning the sensory signals themselves. In the following section, we will describe three types of task phase representations in the LPFC.
Development and maintenance of task phase selectivity during the cue and delay periods
We examined the task phase selectivity of neuronal responses during the cue and delay periods by conducting a two-way ANOVA on cue period (50–550 ms after cue onset) and delay-period (500 ms before cue onset) activity. During the cue period, 30% (n = 498) of the responsive neurons were selective for only the task phase, and 33% (n = 553) were selective for both the task phase and the sensory modality (Table 2). During the delay period, 33% (n = 550) of responsive neurons were selective for only the task phase, and 17% (n = 279) were selective for both the task phase and the sensory modality (Table 2). Fewer neurons were selective for combinations of modality and phase during the delay period than during the cue period (p < 0.0001), and the number of neurons selective for phase did not differ according to period (p = 0.0568). Examination of the proportion of neurons with either phase or modality selectivity during the two task periods (Table 3) revealed the following. (1) Of the modality-selective neurons in the cue period (n = 303, the top row in Table 3), 53% (n = 161) turned into nonselective neurons in the delay period; the fraction of the nonselective neurons was greater than that of neurons remaining in the same category (i.e., modality-selective, n = 79; Binomial test, p < 0.0001). (2) Of the phase-selective neurons in the cue period (n = 498, the second row in Table 3), 42% (n = 211) remained in the same class during the delay period. This fraction was greater than the fraction of neurons remaining in the same category in the other three groups (χ2 test, p < 0.0001). (3) Of the 553 combination-selective neurons in the cue period (the third row in Table 3), 32% (n = 179) turned into phase-selective neurons and 29% (n = 161) remained in the same category. These fractions were greater than the fraction of neurons turning into the modality-selective neurons (n = 57; binomial test, p < 0.0001). (4) Nonselective neurons in the cue period (n = 305, the fourth row in Table 3) tended to become phase-selective neurons in the delay period (n = 124, 41%; χ2 test, p < 0.0001). These four observations revealed that the information reflecting task phase tended to increase from the cue to the delay period, indicating that task phase coding was developing into the dominant representation among LPFC neurons. In the following sections, we will describe three classes of phase coding found in the LPFC: single-phase coding, two- or three-phase coding, and modality and phase coding.
Comparison of response selectivity between cue and delay periods
Single-phase coding
During the cue period, 498 neurons exhibited selectivity for the task phase but not the modality; of these, 118 (24%) preferred a single phase (Tukey's HSD test, p < 0.01). Figure 5 shows four examples of neurons that were preferentially active in response to the first (Fig. 5A), second (Fig. 5B), third (Fig. 5C), or fourth (Fig. 5D) cue, regardless of sensory modality. Preferences were distributed across the four cue periods (39 during the first cue, 16 during the second, 20 during the third, and 43 during the fourth). The population-level activity of each group of single-phase selective neurons is illustrated in Figure 6A–D. The prompt development of the task phase-selective activity in response to one of the individual cues with a sharp preference for one of the four phases indicates the potency of LPFC neurons in providing phase-specific information.

Four examples of the phase-selective neuronal activity in the cue period. Rasters and spike-density functions (smoothed using a Gaussian kernel; σ = 10 ms) indicate activity sorted according to the onset of each cue. The red-, green-, and blue-colored raster displays and spike–density functions represent the activities in the visual, auditory, and tactile trials, respectively. The ordinate represents the instantaneous firing rate (in spikes/s). The gray area indicates the 1 s period during which each cue was presented. The gray area for the fourth cue was trimmed at the average response time of animals. The reward was delivered 500 ms after button release. The tick marks on the horizontal axis were placed at 1 s intervals. A, Activity of a first cue-selective neuron. B, Activity of a second cue-selective neuron. C, Activity of a third cue-selective neuron. D, Activity of a fourth cue-selective neuron.

Population-level activities and normalized responses of phase-selective neurons. A–D, Population-level activities of neurons with phase-selective responses in the cue period. The format is the same as that used in Figure 3. Red, Visual trials; green, auditory trials; blue, tactile trials. Activity of neurons preferring the first cue (A, n = 39), the second cue (B, n = 16), the third cue (C, n = 20), and the fourth cue (D, n = 43) are shown separately. E–H, Population-level activities of phase-selective neurons in the delay period. Activity of neurons preferring the pre-first delay (E, n = 73), the first delay (F, n = 9), the second delay (G, n = 17), and the third delay (H, n = 20) are shown separately. A′–H′, Normalized activity (± SEM) of phase-selective neurons in the cue period. Activity preferring the first cue (A′), the second cue (B′), the third cue (C′), and the fourth cue (D′) are displayed separately. E′–H′. Normalized activity of phase-selective neurons preferring the pre-first cue (E′), the first delay (F′), the second delay (G′), and the third delay (H′) in the delay period.
During the delay period, single-phase selective activity was observed in 119 of the 550 phase-selective neurons. Figure 7 shows four examples of neurons that were preferentially active in the pre-first cue delay (Fig. 7A), the first delay (Fig. 7B), the second delay (Fig. 7C), and the third delay (Fig. 7D) regardless of sensory modality. The preferences of neurons were distributed across the four delay periods (73 during the pre-first cue delay, nine during the first delay, 17 during the second delay, and 20 during the third delay; Tukey's HSD test, p < 0.01). The population-level activity of single-phase selective neurons during the delay period is displayed in Figure 6E–H. The phase-selective activity during the delay period was characterized by its long duration, which prompted us to examine the duration of activity and to identify its beginning and end according to each cue. The activity onset was determined by identifying when the population-level activity exceeded a threshold, defined as baseline activity (the 100 ms period before cue onset) plus half the maximum modulation above this value during the last 100 ms of the subsequent delay period. The activity offset was determined by identifying when the population-level activity fell below a threshold, defined as baseline activity (the 100 ms period before cue onset) minus half the maximum modulation below this value during the last 100 ms of the subsequent delay period. The onset latency, measured from the onset of the cue preceding the delay, ranged from 243 to 618 ms (Table 4). The offset latency ranged from 283 to 748 ms after the onset of the next cue that appeared during the later part of the delay (Table 4). This analysis indicated that the single-phase-selective delay-period activity encompassed a time period sufficiently long enough to link the two successive cue periods.

Four examples of phase-selective neuronal activity in the delay period. The display format is the same as that used in Figure 5. A, Activity of a pre-first cue-selective neuron. B, Activity of a first delay-selective neuron. C, Activity of a second delay-selective neuron. D, Activity of a third delay-selective neuron.
Onset and offset latency of single-phase-selective delay responses
To examine the magnitudes of the selectivity during the cue and delay periods, we normalized the activity of each single-phase neuron relative to its activity in the phase with the highest discharge rates. The normalized activity of each neuron was subsequently averaged across neurons in each group. The normalized population-level responses during the cue (Fig. 6A′–D′) and delay (Fig. 6E′–H′) periods revealed that the responses representing the preferred phase were twofold greater than those during other phases. Neurons displaying single-phase selectivity were distributed widely in both the dorsal and ventral aspects of the LPFC (Fig. 8A, top row, for the cue period; Fig. 8B, top row, for the delay period). We did not find any clustering of neurons classified in each group (MANOVA, p > 0.05).

Cortical surface map displaying the distribution of neurons with phase-selective responses in the cue period (A) and delay period (B). A, B, The top row shows the distribution of neurons with preference for a single phase. The middle row shows the distribution of neurons with preference for two successive phases. The bottom row shows the distribution of neurons with preference for the three successive phases. The preference is indicated on top of each panel.
Two-phase and three-phase coding
We identified the second group of task phase-selective neurons whose activity signaled the occurrence of two or three successive phases in combination. This property was thought to be useful for binding multiple task phases. Of the 498 neurons showing phase selectivity in the cue period, 25 also showed elevated activity during two successive cue periods (two-phase coding in the cue period; Tukey's HSD test, p < 0.01) and 30 showed elevated activity during three successive cue periods (three-phase coding in the cue period; Tukey's HSD test, p < 0.01). Of the 25 two-phase coding neurons, seven responded to the first and the second cues (Fig. 9A,A′), 11 responded to the second and third cues (Fig. 9B,B′), and seven responded to the third and fourth cues (Fig. 9C,C′). Of the 30 three-phase coding neurons, 24 were active from the first to the third cues (Fig. 9D,D′, n = 24), and six were active from the second to the fourth cues (Fig. 9E,E′).

Population-level activities and normalized responses of phase-selective neurons with elevated activity in successive task phases. The display format is the same as that used in Figure 6. A–C′, Population-level activities (A–C) and normalized responses (A′–C′) of neurons with two successive phase-selective responses in the cue period. A, A′, First and second cue-selective responses (n = 7). B, B′, Second and third cue-selective responses (n = 11). C, C′, Third and fourth cue-selective responses (n = 7). D–E′, Population-level activities (D, E) and normalized responses (D′, E′) of neurons with three successive phase-selective responses in the cue period. D, D′, First to third cue-selective responses (n = 24). E, E′, Second to fourth cue-selective responses (n = 6). F–H′, Population-level activities (F–H) and normalized responses (F′–H′) of neurons with two successive phase-selective responses in the delay period. F, F′, Pre-first cue and first delay selective responses (n = 5). G, G′, First and second delay-selective responses (n = 11). H, H′, Second and third delay-selective responses (n = 8). I–J′, Population-level activities (I, J) and normalized responses (I′, J′) of neurons with three successive phase-selective responses in the delay period. I, I′, Pre-first cue to second delay-selective responses (n = 20). J, J′, First to third delay-selective responses (n = 32).
Of the 550 neurons with phase selectivity during the delay period, 24 showed elevated activity during two successive delay periods (two-phase coding in the delay period; Tukey's HSD test, p < 0.01) and 52 showed elevated activity during three successive periods (three-phase coding in the delay period; Tukey's HSD test, p < 0.01). Of the 24 two-phase coding neurons, five showed elevated activity during the pre-first cue and first delay periods (Fig. 9F,F′), 11 showed elevated activity during the first and second delay periods (Fig. 9G,G′), and eight showed elevated activity during the second and third delay periods (Fig. 9H,H′). Of the 52 three-phase coding neurons, 20 were active from the pre-first delay to the second delay periods (Fig. 9I,I′), and 32 were active from the first to the third delay periods (Fig. 9J,J′). These neurons were distributed widely in both the dorsal and ventral aspects of the LPFC (Fig. 8A presents the cue-period-selective, and Fig. 8B presents the delay-period-selective neurons; the middle row shows two-phase neurons, and the bottom row shows three-phase neurons). We did not find any clustering of neurons classified in each group (MANOVA, p > 0.05).
Modality and phase coding
In addition to the single-phase and the two- and three-phase coding, a third type of task phase coding, in which the representation of the task phase was combined with that of the sensory modality, was also observed.
Analysis using a two-way ANOVA revealed that 553 neurons were selective for a specific combination of modality and phase during the cue period, and 279 neurons exhibited this selectivity during the delay period (Table 2). Three examples of neuronal activity of this sort are shown in Figure 10A–C. The neuron in Figure 10A was selectively active during the third delay period in the visual trials. The activity of the neuron in Figure 10B was greater in the first, second, and third delay periods during the auditory trials and also responded most strongly to the fourth auditory cue. The neuron displayed in Figure 10C showed greater discharge rates from the first to the second cues of the tactile trials.
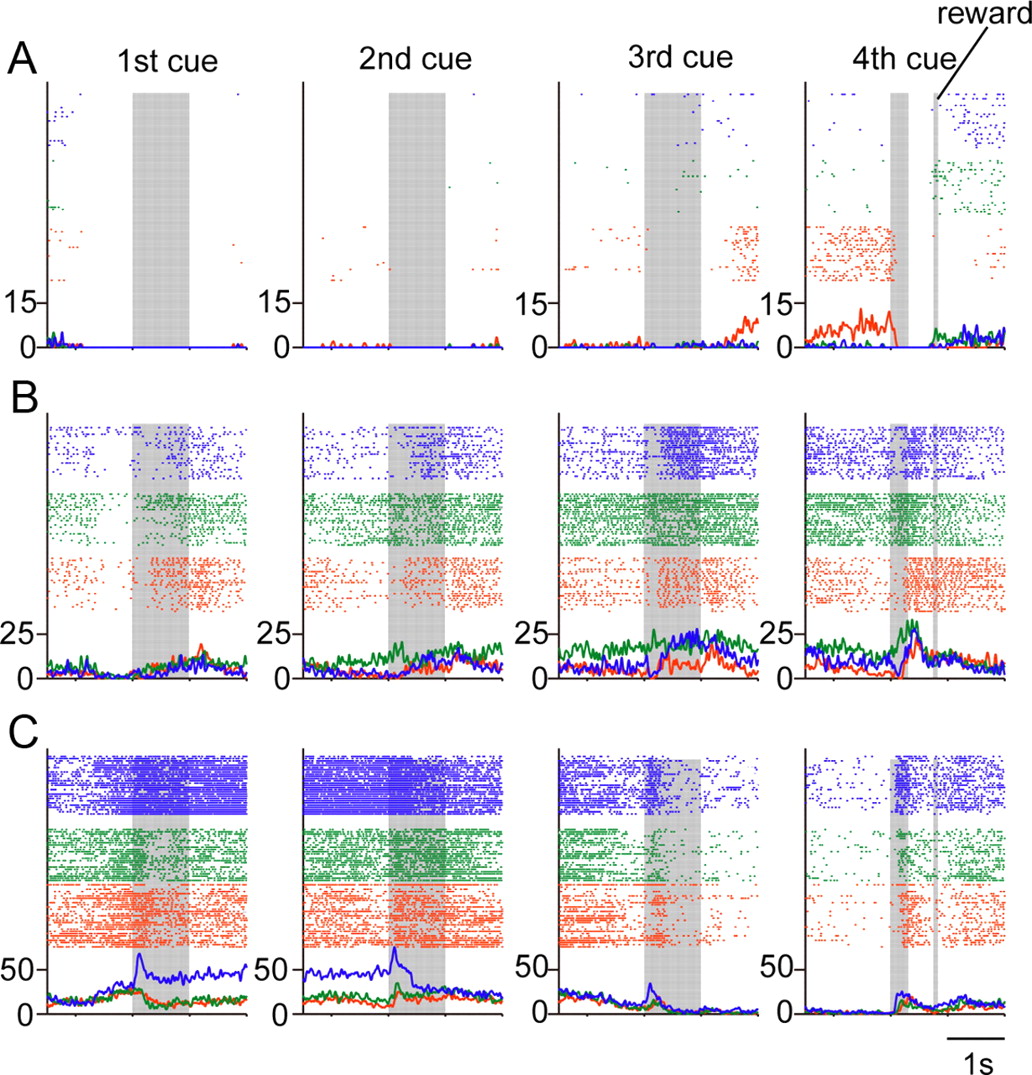
Three examples of neuronal activity selective for specific combinations of modality and phase. The display format is the same as that in Figure 5. A, Activity selective for the visual modality and the phase position (the third delay). B, Activity selective for the auditory modality and the phase position (the second delay to the fourth cue). C, Activity selective for the tactile modality and the phase position (the first to the second cue).
To examine the response profiles of neurons selective for combinations, we classified neurons according to the combination of the modality and phase that led to the highest discharge rate (Table 5). The population-level activity and the quantification of the activity (normalized responses) for each group of neurons during the cue period are shown in Figure 11, and those during the delay period are presented in Figure 12. These analyses indicated that every combination of phase and sensory modality was represented by LPFC neurons during both cue and delay periods. Modality- and-phase-coding neurons appeared widely in both the dorsal and ventral aspects of the LPFC when we examined both the cue (Fig. 13A) and delay (Fig. 13B) periods. We did not find any clustering of neurons classified in each group (MANOVA, p > 0.05). These results revealed that LPFC neurons were involved in encoding the phase in conjunction with the sensory modality.
Distribution of the selectivity of neurons showing combination selectivity

Population-level activities and normalized responses of neurons with selectivity for the combination of modality and phase in the cue period. The display format is the same as that used in Figure 6. A–D′, Population-level activities of visual neurons selective for the first cue (A, A′; n = 96), the second cue (B, B′; n = 73), the third cue (C, C′; n = 58), and the fourth cue (D, D′; n = 58). E–H′, Population-level activities of auditory neurons selective for the first cue (E, E′; n = 46), the second cue (F, F′; n = 27), the third cue (G, G′; n = 20), and the fourth cue (H, H′; n = 37). I–L′, Population-level activities of tactile neurons selective for the first cue (I, I′; n = 59), the second cue (J, J′; n = 28), the third cue (K, K′; n = 18), and the fourth cue (L, L′; n = 33).

Population-level activities and normalized responses of neurons with selectivity for the combination of modality and phase in the delay period. The display format is the same as that used in Figure 6. A–D′, Population-level activity of visual neurons selective for the pre-first delay (A, A′; n = 26), the first delay (B, B′; n = 17), the second delay (C, C′; n = 21), and the third delay (D, D′; n = 34). E–H′, Population-level activities of auditory neurons selective for the pre-first delay (E, E′; n = 15), the first delay (F, F′; n = 24), the second delay (G, G′; n = 19), and the third delay (H, H′; n = 17). I–L′, Population-level activities of tactile neurons selective for the pre-first delay (I, I′; n = 35), the first delay (J, J′; n = 28), the second delay (K, K′; n = 20), and the third delay (L, L′; n = 23).

Cortical surface map displaying the distribution of neurons with combination selectivity in the cue period (A) and delay period (B). A, B, The top row shows the distribution of neurons with a preference for a specific combination of the visual cue and the phase position. The middle row shows the distribution of neurons with a preference for a specific combination of the auditory cue and the phase position. The bottom row shows the distribution of neurons with a preference for a specific combination of the tactile cue and the phase position. The activity preference is indicated at the top of each panel.
Development of task phase selectivity after the arrival of the sensory signal
To investigate the temporal relationship between activity reflecting the modality of the sensory signals and that reflecting the task phase, we analyzed the onset activity of neurons in three groups of neurons possessing the following: (1) early cue responses with unimodal selectivity, (2) single phase-selective responses during the cue period; and (3) responses selective for combinations of modality and phase during the cue period (see Materials and Methods, Detection of activity latency). We compared the cumulative distributions of the neuron latency in the modality-selective, phase-selective, and combination-selective categories (Fig. 14). The median latencies are summarized in Table 6. In the visual (Fig. 14A), auditory (Fig. 14B), and tactile (Fig. 14C) trials, neurons with modality-selective early cue responses began to discharge earlier than did phase-selective neurons after the appearance of every cue (Kolmogorov–Smirnov Test, p < 0.05, except for the responses to the third cues in the auditory trials), revealing that activity reflecting the modality of sensory signals began earlier than did task phase-selective activity. We then compared the onset latency of the combination-selective neurons with that of the modality- and phase-selective neurons. Although the combination-selective neurons tended to discharge after the neurons with the early cue responses (Kolmogorov–Smirnov Test, p < 0.05, except for the responses to the third and fourth cues in the auditory trials and the third cues in the tactile trials), the activity of the combination-selective neurons did not generally differ from those of the phase-selective neurons (Kolmogorov–Smirnov test, p ≥ 0.05, with the exception of the responses to the first and second cues in the visual trials), suggesting that combination-selective activity was similar to phase-selective activity in terms of time of appearance.

Latencies of individual neurons. A–C, Cumulative distributions of latencies at which individual neurons began to respond to the appearance of the cue. A, Visual trials. B, Auditory trials. C, Tactile trials. In A–C, the four panels from left to right indicate data after the onset of the first, second, third, and fourth cue, respectively. “M” shows the onset latencies of the unimodal neurons with early cue responses and with a preference for the visual cue (A, the red line labeled “Visual”), the auditory cue (B, the green line labeled “Auditory”), and the tactile cue (C, the blue line labeled “Tactile”). “P” (black line labeled “Phase”) shows the onset latencies of the single-phase selective neurons in the cue period. The latencies were calculated according to the preference for the phase position (first to fourth cue) and for the modality [visual trials (A), auditory trials (B), and tactile trials (C)]. “C” shows the onset latencies of neurons selective for the combination of phase and modality in the cue period (indicated with “Combination”). The latencies were calculated separately for neurons preferring a specific combination of phase and modality (Table 5). The p values of the statistical analysis (the two-sample Kolmogorov–Smirnov test) are displayed for each comparison of the onset latencies. M-P, Between unimodal neurons with early cue responses and neurons with single-phase selectivity; M-C, between unimodal neurons and combination neurons; P-C, between phase neurons and combination neurons.
Median onset latency of selective responses
Discussion
By analyzing PFC neuronal activity while monkeys performed a task requiring the continuous monitoring of four consecutive signals to correctly initiate an action, we were able to identify three types of neuronal selectivity: sensitivity to sensory modality, sensitivity to task phase, and sensitivity to the combination of sensory modality and task phase. Most importantly, we found that LPFC neurons encoded the task phase indicated by the discrete sensory events in multiple ways: with single-phase selectivity, successive-phase selectivity, and a combination of phase and modality selectivity. Moreover, these phase representations, found continuously arranged throughout the task sequence, were updated promptly by the reception of each sensory signal, which likely arrived via sensory association areas, in time for the initiation of the behavioral response. We will discuss these findings in the context of previous results and in terms of functional significance in what follows.
Encoding of sensory signals by LPFC neurons
Of the neurons exhibiting early cue responses, 84% (15% of the total population) were responsive to only one sensory modality. When all the neurons responsive during the cue period (50–550 ms after onset) were considered, 18% were selective for only sensory modality. Thus, 15–18% of task-related LPFC neurons are thought to be involved in the initial reception of each sensory signal. Visual neurons were prevalent in the caudal region of the LPFC, which receives inputs from visual areas in the posterior association areas (Barbas and Mesulam, 1985, Webster et al., 1994, Schall et al., 1995). In contrast, tactile-responsive neurons were primarily located ventral to the principal sulcus at the rostrocaudal middle level that corresponds to the region having anatomical connectivity with the secondary somatosensory area (Preuss and Goldman-Rakic, 1989). A somatosensory region in the ventral LPFC of humans (Kostopoulos et al., 2007) in which neurons reflect the frequency of vibrotactile stimuli has been identified (Romo et al., 1999). Wide regions in the LPFC receive specific inputs from auditory areas in the superior temporal cortex (Hackett et al., 1999; Romanski et al., 1999a,b), and functional specialization has been demonstrated between the dorsal and the ventral LPFCs (Vaadia et al., 1986; Romanski et al., 1999b; Kikuchi-Yorioka and Sawaguchi, 2000; Romanski and Goldman-Rakic, 2002; Cohen et al., 2004; Sugihara et al., 2006; Russ et al., 2008; Cohen et al., 2009). In the current study, we observed a widespread distribution of auditory neurons without a clear functional segregation, which was reasonable because we asked the animals to detect the onset of a simple auditory signal. In general, the distribution of unimodal-responsive neurons during the cue period was consistent with the anatomical organization of prefrontal connections with sensory association areas, suggesting that the retrieval of each sensory signal was, at least in part, achieved through these pathways.
Continuous and multidimensional representation of the behavioral phase
Of the task-related neurons, 30 and 33% reflected only the task phase position during the cue and delay periods, respectively. A subset of these neurons exhibited preferential activity for a single phase. The timing of the onset and offset of the activity of these neurons tended to be later than the onset of unimodal-selective activity, suggesting that phase selectivity was initiated after a certain amount of information was processed following the arrival of the sensory signal. We further found that the activity of neurons with single-phase selectivity during the delay period had already begun while the preceding cue was present and continued until well after the subsequent cue was presented. This finding indicated that the task phase was continuously represented throughout the entire task period by the combination of cue-responsive and delay-responsive neurons. Additionally, the fourth cue-selective responses began in advance of the monkeys' key release, which was early enough to trigger the motor task. These observations were construed as indicating that the task phase was continuously monitored by individual types of neuronal activity with selectivity for each phase until the initiation of an action, which occurred when the fourth signal was detected.
We found neuronal activity reflecting specific combinations of sensory modality and task phase. Every combination of phase and modality preference was represented during both cue and delay periods. The onset latency of the combination-selective neurons was closer to that of the phase-selective neurons, and both were longer than that of the unimodal-selective neurons. This finding suggests that combination-selective activity may be involved in encoding the task phase in conjunction with the sensory modality. Alternatively, the combination selectivity may be used for monitoring the context of the behavioral sequence, encompassing a wide range of sensory and motor events. We also identified the phase-selective neurons that were active in two or three successive phases. The activity of these neurons seemed to be useful in binding multiple phases of the task to render the task phase coding more relational and robust. In relation to the last point, Bernacchia et al. (2011) applied a theoretical analysis to neuronal activity recorded from the dorsolateral prefrontal cortex and related areas of monkeys performing a matching penny task. They found that neuronal activity reflects multiple time scales of reward memory ranging from hundreds of milliseconds to tens of seconds, suggesting that the time and phase are represented with multiple spans in the association areas.
Together, these results led to the view that LPFC neurons encode multiple task phases not only in a chain-like fashion by connecting activity with single-phase selectivity but also in a multidimensional manner by binding information representing multiple successive phases and by attaching information about the modality of sensory signals to task phase information at the level of individual neurons.
It was reported previously that patients with excision of their frontal lobes experienced difficulty with counting tactile or auditory stimuli presented at 1 s intervals, although they performed normally when these stimuli were presented at shorter intervals, indicating that the frontal lobe is involved in sustaining attention for counting sensory events (Wilkins et al., 1987). The continuous and sustained representation of the task phase found in our study is viewed as one of the crucial neural correlates of frontal lobe functions that could explain the deficits found in that report.
Lashley (1951) commented on the neural mechanisms for temporal control by arguing as follows: “The signal input is never into a quiescent or static system, but always into a system that is already actively excited and organized. In the intact organism, behavior is the result of interaction of this background excitation with input from any designated stimulus” (p. 112). We suggest that the variety of LPFC activity found in the delay period corresponded to the background excitation and that the prompt development of the subsequent task phase and the prompt termination of the preceding task phase during the cue period corresponded to the interaction of this background excitation with input from the designated stimulus. Recent studies (Genovesio et al. 2006, 2011) showed that LPFC neurons of monkeys encoded whether the first or second stimulus had lasted longer or appeared farther as a part of neural processes, extending Lashley's concept by revealing that phase coding is used as an intermediate representation in a variety of cognitive operations.
Interaction between the LPFC and other cortical areas in phase coding
The multimodal nature of information representation has been shown in the parietal association cortex (Andersen et al., 1997; Duhamel et al., 1998), the temporal cortex (Ghazanfar et al., 2005; Kayser et al., 2005, 2007), and the polysensory region of the superior temporal sulcus (Bruce et al., 1981; Hikosaka et al., 1988). Because the LPFC is connected with all these regions, it is probable that the variety of activity found in the current study developed through interactions with these areas. On the other hand, a robust rank or phase representation has been shown in multiple areas (Nieder and Dehaene, 2009), including the anterior part of the cingulate sulcus (Procyk et al., 2000; Hoshi et al., 2005), the supplementary eye field (Berdyyeva and Olson, 2009; Campos et al., 2010), the presupplementary motor area (Tanji, 2001; Sohn and Lee, 2007), and the parietal association cortex (Sawamura et al., 2002; Nieder et al., 2006), leading to the possibility that the interaction of the LPFC with these areas plays an important role. Barbas and colleagues have provided structural evidence that the anterior cingulate cortex (area 32) exerts an inhibitory effect on the neuronal activity of the LPFC (area 46) by preferentially innervating the inhibitory interneurons (Medalla and Barbas, 2009, 2010). Thus, the anterior cingulate may play a major role in terminating sustained delay period activity. We need to examine how the interaction of the LPFC with other cortical and subcortical areas leads to higher cognitive functions, such as task phase coding, which is well developed in human and nonhuman primates.
Conclusions
Neurons in the LPFC provide continuous and multidimensional representations of behavioral task phases that are updated promptly after the reception of sensory signals from multiple modalities. These properties of neuronal activity are viewed as essential for establishing the temporal contingencies among multiple behavioral events, leading to the purposeful organization of motor behavior.
Footnotes
Acknowledgments: This work was supported by Grant-in-Aid for Scientific Research (19300110, J.T.), Grant-in-Aid for Young Scientists (19670004, E.H.), and Takeda Science Foundation (E.H.). We thank members of the Tanji Laboratory for assistance and discussions and S. Hoffman for technical assistance.
The authors declare no competing financial interests.
- Correspondence should be addressed to Dr. Eiji Hoshi, Frontal Lobe Project, Tokyo Metropolitan Institute of Medical Science, Kamikitazawa 2-1-6, Setagaya-ku, Tokyo 156-8506, Japan. hoshi-ej{at}igakuken.or.jp





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}