

Abstract
Amyloid precursor protein (APP) belongs to a family of evolutionarily conserved transmembrane glycoproteins that has been proposed to regulate multiple aspects of cell motility in the nervous system. Although APP is best known as the source of β-amyloid fragments (Aβ) that accumulate in Alzheimer's disease, perturbations affecting normal APP signaling events may also contribute to disease progression. Previous in vitro studies showed that interactions between APP and the heterotrimeric G protein Goα-regulated Goα activity and Go-dependent apoptotic responses, independent of Aβ. However, evidence for authentic APP–Go interactions within the healthy nervous system has been lacking. To address this issue, we have used a combination of in vitro and in vivo strategies to show that endogenously expressed APP family proteins colocalize with Goα in both insect and mammalian nervous systems, including human brain. Using biochemical, pharmacological, and Bimolecular Fluorescence Complementation assays, we have shown that insect APP (APPL) directly interacts with Goα in cell culture and at synaptic terminals within the insect brain, and that this interaction is regulated by Goα activity. We have also adapted a well characterized assay of neuronal migration in the hawkmoth Manduca to show that perturbations affecting APPL and Goα signaling induce the same unique pattern of ectopic, inappropriate growth and migration, analogous to defective migration patterns seen in mice lacking all APP family proteins. These results support the model that APP and its orthologs regulate conserved aspects of neuronal migration and outgrowth in the nervous system by functioning as unconventional Goα-coupled receptors.
Introduction
Amyloid precursor protein (APP) is best known as the source of β-amyloid (Aβ) peptides that have been postulated to cause Alzheimer's disease (AD) (Hardy and Selkoe, 2002). However, therapeutic strategies targeting Aβ have been unsuccessful (Karran et al., 2011), suggesting that other APP-related processes may contribute to the disease (Mangialasche et al., 2010). APP is a member of an evolutionarily ancient family of type 1 glycoproteins that possess highly conserved extracellular and intracellular domains, indicating that they can participate in transmembrane signaling events (Turner et al., 2003; Gralle and Ferreira, 2007). Both full-length APP and its cleavage products have been ascribed multiple roles in neuronal motility (Perez et al., 1997; Sabo et al., 2003; Young-Pearse et al., 2008), including the control of neuronal migration in the developing brain (Herms et al., 2004; Young-Pearse et al., 2007; Rice et al., 2012). However, attempts to validate these functions in vivo have produced conflicting results, in part due to molecular redundancy with two closely related proteins (APLP1 and APLP2) and compensatory interactions by other guidance cues (Heber et al., 2000; Bergmans et al., 2010). Although APP may interact with a plethora of adapter and signaling proteins (Reinhard et al., 2005), the mechanisms by which APP and its orthologs regulate neuronal motility in the nervous system have remained elusive.
Intriguing studies have shown that APP interacts with the heterotrimeric G protein Goα, at least under some conditions. In artificial liposomes and extracted membranes, APP can regulate Goα activity (Nishimoto et al., 1993; Okamoto et al., 1995), while cells transfected with APP isoforms associated with familial AD (FAD) exhibit constitutive Goα activation and accelerated apoptosis (Okamoto et al., 1996; Yamatsuji et al., 1996). Notably, these effects were prevented by the Gi/o inhibitor pertussis toxin or by expressing APP isoforms lacking their putative Go-binding domain (Yamatsuji et al., 1996). Elevated G protein activity and decreased APP–Goα interactions have also been detected in brain samples from AD patients (Reis et al., 2007; Shaked et al., 2009), while cell culture studies suggest that Aβ peptides induce neurotoxic effects via the dysregulation of APP-Goα signaling (Sola Vigo et al., 2009). These results support the model that APP might function as an atypical Go-coupled receptor whose normal functions are disrupted in AD. However, a viable assay for investigating endogenous APP–Goα interactions in neurons has been lacking.
To address this issue, we have established the embryonic nervous system of Manduca sexta (hawkmoth) as a novel preparation for testing how APP family proteins control neuronal migration. As in other invertebrate models, Manduca express only one APP ortholog (APP-Like; APPL), and previous studies have shown that both APPL and Goα are robustly expressed by migratory neurons in this system (Horgan et al., 1995; Swanson et al., 2005). We have now used a combination of in vitro and in vivo assays to determine whether endogenously expressed APP family proteins interact with Goα in neurons from multiple species, whether this interaction is direct, and whether APPL-Goα signaling regulates neuronal migration within the developing nervous system.
Materials and Methods
Whole-mount immunostaining of staged embryos.
Synchronous groups of embryos of either sex were collected from an in-house colony of M. sexta and staged according to a panel of external and internal developmental markers (Copenhaver and Taghert, 1989a,b). When reared at 25°C, embryogenesis is complete in 100 h, whereby 1% of development is equivalent to 1 h postfertilization (hpf). Embryos were collected at 55, 58, and 65 hpf (before, during, and after enteric plexus (EP) cell migration) and dissected in defined saline (140 mm NaCl, 5 mm KCl, 28 mm glucose, 40 mm CaCl2, and 5 mm HEPES, pH 7.4) to expose the enteric nervous system (ENS), as described previously (Horgan et al., 1995; Coate et al., 2007). For immunohistochemical analysis, embryos were filleted dorsally to expose the developing ENS and subsequently fixed in 4% paraformaldehyde (PFA; Sigma-Aldrich) in PBS for 1 h at room temperature. After extensive rinsing in PBS plus 0.1% Triton X-100 (PBST), embryos were pre-incubated for 1 h in blocking solution (10% normal horse serum plus 0.1% sodium azide in PBST). Embryos were then incubated in antibodies diluted in blocking solution with constant agitation for either 1 h at room temperature or overnight at 4°C.
For immunostaining, antibodies were used at the following concentrations: anti-mouse Fasciclin II (Fas II; C3 monoclonal), 1:20,000 (Wright et al., 1999); affinity-purified anti-Goα, 1:100 (generated against amino acids 343–355 of Manduca Goα; Horgan et al., 1995); and anti-cAPPL, 1:2500 (previously referred to as anti-msAPPL-cyt), generated against the sequence YENPTYKYFEVKE within the cytoplasmic domain of Manduca APPL (Swanson et al., 2005). We also generated an additional polyclonal antiserum (anti-nAPPL, #21506; 1:5000) against a fusion protein derived from the E1 ectodomain region of Manduca APPL (amino acids 1–197). The specificity of this antiserum for APPL was validated in Western blots of Manduca and Drosophila lysates; by coimmunohistochemical staining with our other APPL antibodies; by pre-adsorption against the fusion protein (vs control fusion proteins); and by cross-immunoprecipitation with other anti-APPL antibodies (data not shown). Primary antibodies against Fas II and APPL were detected with fluorochrome-conjugated secondary antibodies diluted in blocking solution. Antibodies conjugated to Alexa Fluor 488, 568, or 647 (Invitrogen/Life Technologies) were used at a final concentration of 1:1000; antibodies conjugated to Cy3 and DyLight 549 (Jackson ImmunoResearch) were used at 1:400. Anti-Goα was detected with anti-guinea pig secondary antibodies coupled to horseradish peroxidase (HRP) and visualized using the tyramide signal amplification system (TSA Plus Fluorescence Kit; PerkinElmer), following the manufacturer's instructions. Whole-mount immunostained preparations were stored in Elvanol (Banker and Goslin, 1998) and imaged with an Olympus FluoView 300 laser scanning confocal head mounted on an Olympus BX51 microscope (located in the Live Cell Imaging Facility, Center for Research on Occupational and Environmental Toxicology, OHSU), or with an inverted Zeiss LSM710 confocal microscope (located in the Advanced Imaging Center of the Jungers Institute, OHSU). Maximum intensity projections of flattened Z-stack confocal images were generated using MetaMorph software. To illustrate the relationships between the EP cells and their muscle band pathways (Fig. 1), Fas II immunostaining in each panel was cropped at either the foregut (FG)/midgut (MG) boundary (55 hpf) or at positions adjacent to the most posterior neurons on each pathway (58–65 hpf). The cropped Fas II images were then montaged with the flattened Z-stack images of APPL immunostaining in the same preparations.
To examine the colocalization of mammalian APP with Goα, primary cultures of embryonic rat hippocampal neurons (grown on coverslips) were provided by Dr. Gary Banker (OHSU; Dotti et al., 1988). Neurons were fixed with 4% PFA in PBS for 15 min at room temperature, permeabilized for 10 min with 0.1% PBST, and blocked for 15 min with 5% fish skin gelatin in PBST. The following primary antibodies were applied for 60 min at room temperature: anti-22C11 targeting amino acids 66–81 of human APP (Millipore #MAB348, 1:100); anti-pAPP, specific to pThr668 within amino acids 666–670 of human APP (Sigma-Aldrich #SAB4300153; 1:200); and affinity-purified anti-Goα, 1:300 (Horgan et al., 1995). Secondary antibodies conjugated to Alexa Fluor 488, 568, or 647 (Invitrogen/Life Technologies) were used at a final dilution of 1:1000. Coverslips were mounted in Elvanol and imaged as described above.
Coimmunoprecipitation and immunoblotting.
Staged Manduca embryos (20 per reaction; dissected at 65 hpf) and adult Drosophila heads (20 per reaction) were collected on dry ice and homogenized in 1% Triton lysis buffer (1% Triton X-100, 150 mm NaCl, 50 mm Tris, pH 8) or 1% NP40 lysis buffer (150 mm NaCl, 50 mm Tris, pH 8). Aliquots of each lysate were centrifuged at 16,000 rpm for 10 min, and the supernatants were precleared with Protein A/G beads (Santa Cruz Biotechnology). The supernatants were then incubated with primary antibodies (described below) for 1–3 h at room temperature or overnight at 4°C and incubated with prewashed beads for 1 h. The bead-bound antibody complexes were then pelleted by centrifugation. After washing in chilled lysis buffer, immunoprecipitated protein complexes were eluted by boiling the beads in SDS sample buffer for 1 min (Swanson et al., 2005). The samples were then separated on 10% or 4–12% Criterion polyacrylamide gels (Bio-Rad), transferred to nitrocellulose, and immunoblotted with antibodies diluted in Tris-buffered saline plus 0.1% Tween 20 (polysorbate) and 5% Carnation dry milk.
For coimmunoprecipitations using mouse brain, frozen tissue samples were provided by Dr. Joseph Quinn and Christopher Harris (Department of Neurology, Portland Veterans Administration Medical Center, and Layton Center for Aging and Alzheimer's Disease Research, OHSU). Brains were divided in half under liquid nitrogen, extracted in NP40 lysis buffer, and immunoprecipitated by the methods described above. Alternatively, membranes were prepared from lysed mouse brain samples following the protocol described in (Hortsch, 1994). Briefly, tissue was homogenized in hypotonic saline (100 mm Tris-HCl, pH 8.0; 1 mm EDTA) and centrifuged at 100,000 × g for 1 h at 4°C. The pellets were then resuspended in NP40 lysis buffer, incubated with primary antibodies for 3 h at room temperature or overnight at 4°C, and immunoprecipitated as described above. For coimmunoprecipitations using human brain tissue, frozen samples were made available from the Oregon Brain Bank (collected and distributed following established protocols), with the assistance of Dr. Randall L. Woltjer (Department of Pathology, OHSU). Approximately 90 mg of tissue per reaction was homogenized with a Dounce homogenizer in 1 mm EDTA, 100 mm NaCl, 50 mm Tris-HCl, pH 8), plus 1% Triton X-100 (Roche) and 0.5% sodium cholate (Sigma-Aldrich). Undigested protein was then pelleted in a Beckman L7–55 ultracentrifuge for 1 h at 100,000 × g at 4°C, and the supernatants used for subsequent immunoprecipitation reactions as described by (Shaked et al., 2009). Samples were incubated with antibodies overnight at 4°C with continuous rocking and then immunoprecipitated, as described above.
The following antibodies were used to immunoprecipitate APP-associated proteins: for Manduca lysates, we used anti-cAPPL and anti-nAPPL-EX, previously referred to as anti-msAPPL-ect (generated against the sequence EDDDYTDADDSAWPRPES within the extracellular domain of Manduca APPL; Swanson et al., 2005). For Drosophila lysates, we used anti-cAPPL. For mouse lysates, we used anti-nAPP (Sigma-Aldrich #8967, targeting amino acids 46–60 of human APP695); anti-cAPP (Sigma-Aldrich A8717, targeting amino acids 676–695 of human APP695); and anti-APP668 (Sigma-Aldrich #SAB4300464, targeting amino acids 666–670 of human APP695). For human brain lysates, we used anti-nAPP (Sigma-Aldrich #8967). For each experiment, replicate lysates were immunoprecipitated with matched control immunoglobulins: purified IgY (7.5 μg, Aves #N-1010) was used in experiments using chicken-derived antibodies; purified IgG (10–12 μg; The Jackson Laboratory #1-000-003) or normal rabbit serum (1–3 μl) was used in experiments using rabbit-derived antibodies. The following antibodies were used to detect G proteins that were coimmunoprecipitated with APP or APPL: anti-Goα (1:100–1:250; Horgan et al., 1995); anti-Gsα (1:1K) against a conserved sequence shared by Drosophila and mammalian Gsα (Santa Cruz Biotechnology #sc-383); anti-Giα (1:1000), targeting the conserved C-terminal sequence 327–355 (a gift from Dr. Michael Forte, OHSU; as described by Copenhaver et al., 1995); and anti-Gβγ (1:75; BD Transduction Laboratories). Secondary antibodies coupled to HRP were purchased from Jackson ImmunoResearch and used at 1:10K, and detected using standard chemiluminescent protocols (with either West Pico or West Femto substrates; ThermoFisher).
For coimmunoprecipitation experiments to test whether the Go-binding domain in APPL was required for APPL-Goα interactions in Drosophila, we used the following transgenic lines (provided by Dr. Vivian Budnik and Michael Gorczyca, University of Massachusetts Medical School, Worcester, MA): Appld, a null mutation of the Appl gene (Luo et al., 1992); UAS-Applsd, encoding an in-frame mutation that lacks amino acids 758–791 (and has the point mutations Arg795Arg796-Leu795Ser796), which is insensitive to secretase cleavage (secretase-deficient APPL; Luo et al., 1992); and UAS-ApplsdΔCg, secretase-deficient APPL that lacks amino acids 845–855 within its Go domain (APPLsdΔCg; Torroja et al., 1999b). The UAS transgenes were crossed into Appld flies to test the interactions of each APPL variant in the absence of wild-type protein; Appld flies (lacking all APPL) served as a negative control. Expression of the transgenes was accomplished with the GAL4/UAS system (Brand and Perrimon, 1993), using the eye-specific GMR-GAL4 driver line (Bloomington Stock Center). For these assays, Drosophila lysates were immunoprecipitated with anti-Goα (EMD/Calbiochem #371726) and immunoblotted with either anti-cAPPL (1:2500) or anti-Gβγ (1:75), as described above.
Analysis of direct Goα-APPL interactions by Bimolecular Fluorescence Complementation.
Plasmids containing the coding domains for complementary portions of Venus Fluorescent Protein (Vn1 and Vn2) were provided by Dr. Stephen Michnick (University of Montreal, Montreal, Quebec, Canada) and used for a modified version of the Protein-fragment Complementation Assay (Remy et al., 2004). The APPL-Vn1 plasmid construct was generated in pcDNA3 (Invitrogen) by ligating the full-length coding domain of APPL in-frame with Vn1, consisting of amino acids 1–158 of Venus Fluorescent Protein plus a 10 aa glycine-rich linker domain (Benton et al., 2006; Mervine et al., 2006). The APPLΔGo-Vn1 construct was generated using PCR primers designed to omit the full Go-binding domain of Manduca APPL (amino acids 762–791; HAQGEVQVEQTGVVAPTPEERHVANMQING), identified by alignment with the Go-binding domain of human APP695 (His657-Lys676; as described by Nishimoto et al., 1993). Fusion constructs containing the coding domains of insect Goα, Giα, and Gsα were ligated in-frame at their N termini with Vn2, consisting of amino acids 159–239 of Venus Fluorescent Protein plus a 10 aa glycine-rich linker region. For an in vitro analysis of Bimolecular Fluorescence Complementation (BiFC) induced by reassembly of Vn1- and Vn2-tagged fusion proteins, COS7 cells were plated on poly-lysine-coated coverslips (at 20% density) and transiently transfected the following day with either 1000 ng total plasmid DNA or 20 ng plasmid DNA plus 980 ng pGEM-T helper plasmid, using TransIT-LT1 (Mirus Bio). After 18 h, 24 h, or 48 h, the cells were fixed for 10 min in 4% PFA, followed by incubation in blocking solution (PBS plus 10% normal horse serum) for 30 min. Primary antibodies were diluted in blocking solution and applied to the cells for 1 h at room temperature. For this analysis, we labeled the cells with anti-nAPPL (1:1000), anti-Goα (1:100), and mouse anti-GFP (Invitrogen, A-11120), which recognizes the Vn2 fragment of the holoprotein. After rinsing with PBS, the cells were incubated with Alexa Fluor 568- and Alexa Fluor 647-conjugated secondary antibodies (diluted 1:1000) for 30 min, rinsed again, and mounted on glass slides with Elvanol for imaging with confocal microscopy. Z-stack images of each cell (five optical sections per stack) were acquired under linear parameters, using identical laser and acquisition settings. The stacked images were flattened in MetaMorph and adjusted for brightness and contrast, using identical settings for all cells imaged in an experiment. To quantify membrane-associated BiFC levels generated by different combinations of the Vn1 and Vn2 constructs, maximum intensity projections of each stacked image were generated in MetaMorph, and intensity values were determined for equivalent perimeter segments for each cell using Fiji software. Values were obtained independently for anti-APPL, anti-Goα, and BiFC (visualized in different wavelengths). The mean values of average pixel intensities were then derived for replicate sets of cells (at least 20 cells per group), and used to evaluate statistical differences between the BiFC signals generated by the Venus-tagged proteins when expressed individually or in combination.
For BiFC analysis of APPL-Goα interactions in vivo, constructs containing the coding domains for Manduca APPL, APPLΔGo, and Goα were cloned in-frame with Vn1 and Vn2, respectively. The constructs were then ligated into the pUASg (Appl) or pUAST (Goα) vector and used to transform Drosophila (BestGene). Transformant flies homozygous for UAS-APPL-Vn1, UAS-APPLΔGo-Vn1, UAS-Vn2-Goα, or recombined to express combinations of the constructs (UAS-APPL-Vn1 + Vn2-Goα or UAS-APPLΔGo-Vn1 + Vn2-Goα) were crossed with flies carrying the eye-specific GMR-GAL4 promoter construct (Brand and Perrimon, 1993). Mated flies were maintained at 28°C, and third instar larvae were collected at various stages of development for imaging and analysis. Larval eye discs were isolated in PBS and immediately placed on ice. Discs were fixed for 10 min in PBS plus 4% PFA at 4°C, then blocked for 30 min with 10% normal horse serum in PBS plus 0.1% Triton X-100. To detect the expression of either Vn1 or Vn2 fusion constructs, discs were immunostained with polyclonal GFP antibodies (Aves #GFP1020 or Invitrogen #A11122; 1:1000), which recognize both fragments of Venus fluorescent protein. To amplify BiFC signals specifically induced by the reassembly of Vn1 and Vn2, we used anti-GFP (Sigma-Aldrich #G6539; 1:1000), which only labels the recombined protein. Discs were incubated in primary antibodies for 1.5 h, then rinsed with PBS-0.1% Triton X-100, and incubated with secondary antibodies (diluted in blocking solution) for 30 min at room temperature. DyLight 549 goat-anti-chick antibodies (Jackson ImmunoResearch) were used at 1:200; Alexa Fluor-conjugated secondary antibodies (Invitrogen) were used at 1:1000. Whole discs were mounted in Elvanol, and flattened Z-stack images were obtained by confocal microscopy and ImageJ processing, using the same number of optical sections and laser settings for each disc.
Manipulations of endogenous G protein activity.
To test the role of G protein activity in regulating Goα-APPL interactions, lysates were prepared from staged Manduca embryos or from GV1 cells (Hiruma and Riddiford, 2004), a Manduca cell line of ectodermal origin that endogenously expresses APPL and Goα (as well as other neuronal proteins; Coate et al., 2009). Cells and tissues were homogenized in either chilled NP40 buffer (1%) or radioimmunoprecipitation assay buffer, and clarified by centrifugation at 13,000 rpm for 10 min. The following reagents were then added to replicate aliquots of the lysates at the following final concentrations: GTPγS (Sigma), 0.05–10 μm; GDPβS (Sigma), 1–20 μm; Mas7 (a mastoparan analog with enhanced activity; Enzo Life Sciences), 50–400 μm; Mas17 (an inactive mastoparan analog; Enzo Life Sciences), 30 μm; and Pertussis Toxin A protomer (Enzo Life Sciences), 1–4 μg/ml plus 1 mm NAD+. After incubation at room temperature for 30 min, the lysates were briefly centrifuged to remove cellular debris and analyzed by coimmunoprecipitation and immunoblotting, as described above. For analysis using the Odyssey Infrared Imaging System (LI-COR), immunoblots were labeled with IRDye 680- and 800-conjugated secondary antibodies (from Rockland Immunochemicals and Invitrogen). Quantification of band intensities was performed using Odyssey software, and each experimental condition was normalized with respect to untreated control immunoprecipitations (run in parallel on the same immunoblots). Alternatively, immunoblots were processed using the chemiluminescent protocols described above; the blots were then imaged on x-ray film with X-OMAT processing, scanned into TIFF files with Photoshop (Adobe), and relative pixel intensities were calculated for bands of interest using ImageJ (National Institutes of Health, Bethesda, MD). For each treatment condition, relative levels of Goα and APPL were normalized to levels detected in matched control samples (run on the same immunoblots) and imaged with identical parameters. Goα levels were then calculated as a ratio of Goα/APPL levels; each experiment was repeated at least three times. For statistical analysis, levels of immunoprecipitated Goα for each experimental condition were normalized to their respective untreated controls. Mean Goα levels for each treatment condition were then compared with matched untreated controls, using Student's two-tailed t tests. Each experiment was repeated at least three times.
Embryonic culture and migration assays.
Staged Manduca embryos were isolated at 52 hpf, shortly before the onset of EP cell migration. Embryos were dissected from their egg shells and extraembryonic membranes and placed in Sylgard-coated chambers filled with either culture medium (50% Schneider's Drosophila medium, 40% MEM with Hank's salts, 9.7% heat-inactivated normal horse serum, 0.2% 20-hydroxyecdysone, 0.1% insulin, and 0.01% penicillin-streptomycin (pen-strep), pH 7.4); or defined saline (140 mm NaCl, 5 mm KCl, 28 mm glucose, 5 mm HEPES, 4 mm CaCl2, 0.2% 20-hydroxyecdysone, 0.1% insulin, and 0.01% pen-strep, pH 7.4) (Horgan and Copenhaver, 1998). A small incision in the dorsal body wall between segments T2 and T3 (a position just anterior to the FG/MG boundary) was made to expose the EP on the developing gut. The premigratory EP cells were then directly treated with experimental reagents targeting APPL or Goα. The incision was then gently closed with glass electrodes, and the embryos were allowed to develop in culture for another 12–24 h at 28°C, spanning the time of normal EP cell migration and axon outgrowth. In experiments targeting subsequent aspects of axon elongation and terminal branch formation, embryos were opened in culture at 65 hpf (at the end of the migratory period), treated with experimental reagents, and incubated for an additional 24 h, spanning the normal period of synaptogenesis on the midgut musculature (Wright et al., 1998). Aluminum fluoride (AlF4−) was prepared by diluting premixed solutions of 0.6 m NaF and 0.5 mm AlCl3 (1:40) in culture medium to give a final concentration of 12.5 μm AlF4-; equivalent concentrations of NaCl premixed with AlCl3 served as controls (Horgan and Copenhaver, 1998). Pertussis toxin (PTX; intact molecule; Invitrogen) was pre-activated with 1 m dithiothreitol and diluted in serum-free culture medium (Horgan et al., 1995). Treatment of the EP cells with anti-nAPPL antibodies (#21506) was performed by diluting a purified IgG fraction of the antibody in culture medium; equivalent concentrations of control rabbit IgG (Jackson ImmunoResearch) were used as controls.
At the end of each experiment, embryos were dissected completely to expose the ENS, then fixed and immunostained with anti-Fas II to reveal the full extent of EP cell migration and outgrowth (Horgan et al., 1995; Wright et al., 1999). Anti-Fas II immunoreactivity was detected with biotinylated anti-mouse secondary antibodies (1:200) and the avidin-biotin-HRP protocol from Vector Laboratories (ABC kit). The distributions of postmigratory EP cells and their axons were analyzed using photomicrographic and camera lucida techniques. Ectopic outgrowth was quantified using a grid-based analysis of EP cell processes that had extended into each interband region of the midgut (normally inhibitory to the neurons); ectopic neurons were counted manually. To account for day-to-day variability in culture conditions, values obtained from each experimental preparation were normalized to mean values calculated from replicate sets of matched control preparations included with each experiment. Pairwise statistical differences between control and experimental groups were then calculated using Student's two-tailed t tests.
Morpholino antisense oligonucleotides (morpholinos; Gene Tools) were designed against several regions of the mRNA sequence encoding Manduca APPL, and initially tested for their effectiveness and specificity in GV1 cells. Morpholinos were applied to cultured GV1 cells for 48 h (grown in GV1 medium: 3.32 g of lactalbumin hydrolysate and 20 ml 50× Yeastolate per liter of Grace's medium, plus 10% heat inactivated fetal bovine serum and 1% pen-strep). The cells were then lysed in chilled NP40 lysis buffer, and residual APPL levels were analyzed by Western blotting methods; the expression of several control proteins (including Fas II and tubulin) were visualized in the same samples to monitor for off-target effects of the morpholinos. The most effective APPL-specific morpholino (directed against a sequence spanning the initiation codon; CCGCGTTGCTTCCCACCAGCCC) was subsequently used to knock down APPL expression in cultured embryos. Morpholinos were diluted in defined saline (1–50 μm) supplemented with 10% heat-inactivated horse serum, 0.5% pen-strep, 0.2% 20-hydroxyecdysone, 0.1% insulin, and 0.2 m l-glutamine. Embryos were opened in culture at 48 hpf, and the morpholinos were delivered into the EP cells with 0.6% Endo-Porter (Coate et al., 2008). Matched sets of embryos were treated with standard control morpholinos (Gene Tools) or Endo-Porter alone. The embryos were then allowed to develop for an additional 24–48 h at 28°C before fixation and analysis, as described above. Matched sets of cultured preparations were also immunostained with anti-nAPPL to monitor the effectiveness of the morpholinos in inhibiting APPL expression.
Results
Goα colocalizes with endogenous APPL and APP in motile neurons
During formation of the ENS in Manduca, a population of ∼300 neurons (EP cells) undergoes a stereotyped sequence of migration along preformed pathways to form a branching nerve plexus (the EP), which spans the FG/MG boundary (Fig. 1A–C). After delaminating from a neurogenic placode in the FG epithelium (Copenhaver and Taghert, 1990), the EP cells first spread bilaterally around the FG (from 40 to 55 hpf). During this phase of migration (Fig. 1A; 52 hpf) approximately equal subsets of neurons align with one of the eight longitudinal muscle bands that have recently differentiated on the MG surface (Fig. 1A, b; only the four dorsal bands are shown). From 55 to 65 hpf (Fig. 1B; 58 hpf), the neurons migrate in a chain-like manner along each of the muscle bands while avoiding adjacent interband regions (Fig. 1A, ib). From 65 to 75 hpf (Fig. 1C; 65 hpf), the neurons continue to elaborate long axons posteriorly along the MG (Copenhaver and Taghert, 1989a,b). During this process, EP cells actively extend and retract filopodia onto the band and interband muscles but remain aligned with their selected band pathways. Only once migration and axon outgrowth are complete do the neurons eventually sprout lateral branches that innervate the adjacent interband musculature (from 80 to 100 hpf). However, a small number of EP cells also normally extend processes onto the interband regions near the FG/MG boundary (Fig. 1F, asterisks; Copenhaver and Taghert, 1989b).

APPL is expressed by migrating EP cells and their motile processes. A–C, Schematic representation of the developmental sequence of EP cell migration (green) along the eight muscle band pathways (magenta) to form the enteric plexus of the ENS in Manduca (only the dorsal four muscle bands are shown). D–F, Montages of whole-mount preparations of filleted embryos that show the developing ENS from corresponding embryonic stages, immunostained for APPL (green) and Fas II (magenta; Fas II). (For these montaged images, Fas II staining was omitted from the EP cells to clearly show their alignment with the muscle bands; see Materials and Methods). A, D, By 55 hpf, the EP cells have spread bilaterally around the surface of the foregut, adjacent to the foregut/midgut (FG/MG) boundary; all of the neurons express APPL as they extend filopodia (arrows) preferentially onto the Fas II-positive muscle band pathways (b). B, E, By 58 hpf, subsets of EP cells have begun migrating along each muscle band pathway, while avoiding the adjacent interband regions (ib). APPL is strongly expressed throughout the motile cell bodies and within their leading processes that have extended along the bands (arrows). C, F, By 65 hpf, the EP cells have completed their migration (arrows) but will continue to grow axonal processes posteriorly along the band pathways for another 15 h, before eventually innervating the lateral visceral musculature. Throughout this developmental sequence, the EP cells maintain robust levels of APPL expression (particularly in their most motile regions) and remain confined to the muscle band pathways, although a small number occasionally extend processes from the foregut-midgut boundary onto the interband regions (asterisks). EN, esophageal nerve of the FG. White scale bar, 50 μm.
In previous work, we showed that a combination of permissive and repulsive guidance factors regulate the guidance of the EP cells along the muscle bands. The Ig-CAM Fas II (the insect ortholog of NCAM and OCAM) is expressed by both the EP cells and the muscle band cells, and homophilic signaling interactions mediated by Fas II play an essential role in promoting migration along these pathways (Copenhaver et al., 1996; Wright et al., 1999). (For clarity, the montages in Fig. 1, D–F, include Fas II immunostaining only in the muscle bands; see Materials and Methods). In addition, we recently showed that the migratory EP cells express APPL (Fig. 1D–F; green), coincident with their expression of Goα (Horgan et al., 1995; Swanson et al., 2005). Given the in vitro evidence that APP can interact with Goα (Nishimoto et al., 1993) and our previous data showing that Goα-dependent signaling restricts the extent of EP cell migration (Horgan and Copenhaver, 1998), we investigated whether APPL might function as a transmembrane receptor that functionally interacts with Goα in these migratory neurons.
Past studies have shown that insect APPL contains all of the key structural features that typify APP family proteins in other species (Fig. 2A), including conserved E1 and E2 extracellular domains that may interact with potential ligands (Luo et al., 1990; Swanson et al., 2005) and an Aβ domain that has neurotoxic effects when overexpressed as a cleavage fragment (Carmine-Simmen et al., 2009). In addition, the cytoplasmic domain of Manduca APPL shares strong similarity with equivalent domains in other APP-related proteins (Fig. 2B), including 88% sequence identity with Drosophila APPL and 77% identity with human APP695 (Swanson et al., 2005). Of particular note is the TPEER region within the putative Go-binding domain (“Go”; Nishimoto et al., 1993) and the NPXY phosphotyrosine binding domain (Chen et al., 1990; Hoe and Rebeck, 2008; “Y”) that are 100% identical between Manduca and mammalian APP family proteins, suggesting that these domains regulate conserved signaling functions. As with human APP695, APPL undergoes a dynamic sequence of secretase-dependent cleavage to produce soluble ectodomain fragments, C-terminal fragments (CTFs), and APP intracellular domains (AICDs) (Carmine-Simmen et al., 2009; Bolkan et al., 2012; J.M. Ramaker, T.L. Swanson, and P.F. Copenhaver, unpublished observations), each of which might affect different aspects of cell motility (Turner et al., 2003; Gralle and Ferreira, 2007). To investigate whether APPL can act as a Goα-associated receptor in migratory neurons, we first examined whether APPL colocalized with Goα at the plasma membrane of the EP cells, consistent with a potential role in signal transduction.

Goα colocalizes with APP family proteins in the motile regions of developing insect and mammalian neurons. A, Schematic diagram of APPL and APP, indicating the conserved extracellular domains (E1 and E2), Aβ domains, and the cytoplasmic Go-binding domain (Go) and internalization domain (Y). Transmembrane domains (tm) are shown in gray. Epitopes targeted by the different anti-APPL and APP antibodies used in this study are indicated by the labeled black lines. Antibodies targeting APP and APPL extracellular domains are labeled in green and antibodies targeting cytoplasmic domains are labeled in magenta. B, Amino acid alignment of the conserved cytoplasmic domains in human (hu) APP, mouse (m) APP, Manduca (ms) APPL, and Drosophila (dm) APPL. Both the putative Go-binding domain (Go) and the internalization domain (Y) within the cytoplasmic (C-terminal) region of APP family proteins are highly conserved across species. Residues that are identical in at least three species are shaded in black; residues shared by two species are shaded in gray. The dotted black line indicates the putative Go-binding domain identified in APP695 (Nishimoto et al., 1993; Okamoto et al., 1995). C, EP cells at 65 hpf immunostained with anti-nAPPL (green) and anti-cAPPL (magenta); the white box indicates the region highlighted in D–F. Individual channels in E (anti-nAPPL) and F (anti-cAPPL) are shown as monochrome images. Arrowheads indicate colocalized N- and C-terminal immunostaining (which appears white in D) at the plasma membrane, consistent with the presence of full-length transmembrane APPL. Images in C–F show compressed images of three optical sections acquired by confocal imaging. G, Leading processes of EP cells on the dorsal muscle band pathways, immunostained with anti-nAPPL and anti-cAPPL antibodies; the white box indicates the region highlighted in H–J, which includes three to four fasciculated growth cones. Arrowheads indicate colocalized N- and C-terminal immunostaining in the motile growth cones (white regions in H); individual channels in I (anti-nAPPL) and J (anti-cAPPL) are shown as monochrome images. Images in G–J show compressed images of 10 optical sections. K, EP cells coimmunostained with anti-nAPPL (green) and anti-Goα (magenta); white boxes indicate regions highlighted in L–N and O–Q. Arrowheads in L–N indicate the colocalization of APPL and Goα at the plasma membrane of a migrating neuron; arrowheads in O–Q indicate colocalization in the EP cell growth cone. Individual channels in M and P (anti-nAPPL) and N and Q (anti-Goα) are shown as monochrome images. The images in K–Q show single optical sections. R, Rat hippocampal neurons immunostained with antibodies against N-terminal APP (green, 22C11), C-terminal APP (red, pAPP), and Goα (blue); the white box indicates the highlighted region shown in S–T. S, Enlarged view of colocalized N-APP (green) and C-APP (magenta) immunostaining in the neuronal growth cones, indicating the presence of transmembrane APP. T, Corresponding image of anti-Goα immunostaining (shown in monochrome), consistent with the colocalization of Goα with full-length APP in mammalian growth cones. Images R–T show a compressed image of 10 optical sections. Scale bars: (in C,G, K) 7 μm; (in D–F, H–J, L–Q) 3 μm; (in R), 50 μm; (in S, T) 25 μm.
For the current analysis, we used a panel of well characterized antibodies against the extracellular and intracellular domains of APP and APPL (Fig. 2A), including antibodies targeting conserved residues within the putative Go-binding domain. Double immunostaining the developing ENS with anti-nAPPL (against the N-terminal extracellular domain; green) and anti-cAPPL (against the C-terminal intracellular domain; magenta) verified the presence of the full-length holoprotein at the plasma membrane of migrating neurons (Fig. 2C–F, arrowheads) and their leading processes (Fig 2G–J, arrowheads). Also notable in these preparations was the abundance of additional APPL within large perinuclear vesicles, as well as smaller vesicle populations containing either N-terminal (green) or C-terminal fragments (magenta) of the holoprotein (Fig. 2C–F). We also found that the subcellular distributions of APPL and its fragments were markedly altered during periods of active migration and outgrowth, during which transmembrane APPL accumulated in regions of active growth (as manifested by the colocalization of N- and C-terminal epitopes at the membrane). These observations support our previous evidence that APPL undergoes a dynamic sequence of trafficking, processing, and post-translational modifications that correlate with specific phases of EP cell differentiation (Swanson et al., 2005). They also are consistent with recent studies on APP trafficking in neuroblastoma cells (Muresan et al., 2009), suggesting that much of the newly synthesized holoprotein may be proteolytically cleaved and sorted to distinct compartments before their transport out of the cell body. This process may therefore help regulate the bioavailability of APPL as a transmembrane receptor during phases of active motility (J.M. Ramaker, T.L. Swanson, and P.F. Copenhaver, unpublished observations). To determine whether membrane-associated APPL colocalized with Goα in the EP cells, embryos were coimmunostained with antibodies against anti-nAPPL (green) and anti-Goα (magenta). As shown in Figure 2, K–N, robust colocalization was detected at the plasma membrane of the migratory neurons (arrowheads) and within their leading processes (Fig. 2O–Q, arrowheads). Surface labeling of unpermeabilized embryos with anti-nAPPL and subsequent permeabilization and staining for Goα verified that Goα colocalizes with cell-surface APPL in the EP cells (data not shown). To examine the relationship between APP and Goα expression in developing mammalian neurons, embryonic rat hippocampal neurons were isolated and fixed after 5 d in primary culture. The neurons were then triple immunostained with antibodies specific to the extracellular domain (green) and cytoplasmic domain of APP (red/magenta), as well as for Goα (blue/gray scale; Fig. 2R–T). As seen for APPL in the EP cells, full-length APP was found to be abundantly expressed within the growth cones of rat hippocampal neurons, where it colocalized with Goα (yellow/white regions, Fig. 2R–T). These results indicate that transmembrane forms of APP family proteins are closely associated with endogenously expressed Goα in developing neurons, particularly within regions of active growth and motility, and that this association is conserved in both invertebrate and vertebrate preparations.
Both APPL and APP endogenously interact with Goα
In previous investigations, coimmunoprecipitation assays were used to show that both human APP and insect APPL can interact with Goα (Brouillet et al., 1999; Swanson et al., 2005; Shaked et al., 2009), but these studies did not determine whether Goα interacts specifically with full-length APP family proteins or with their C-terminal cleavage products (CTFs and AICDs). Another recent study showed that epitope-tagged CTFs of APP could interact with a different heterotrimeric G protein (Gsα) when overexpressed in cell culture (Deyts et al., 2012). Given that APP contains a conserved BBXXB domain, which in other receptor classes can promote binding to the α-subunits of several different G proteins (Okamoto et al., 1990; Wu et al., 1995; Timossi et al., 2004), these results suggest that APP might function as a promiscuous G protein-associated receptor (Deyts et al., 2012).
To address this issue, we immunoprecipitated APPL from Manduca embryonic lysates with antibodies specific to both the N- and C-terminal domains of the holoprotein, and probed the resultant immunoblots with antibodies against different G protein subunits (Fig. 3A). As shown in Figure 3, B and C, endogenously expressed Goα could be readily coimmunoprecipitated with both N-terminal- and C-terminal-specific APPL antibodies, but not with matched control immunoglobulins (IgG and IgY, respectively). In contrast, APPL did not coimmunoprecipitate with either Giα or Gsα (Fig. 3D,E), despite the robust expression of these G proteins in the embryonic nervous system (Copenhaver et al., 1995). Since APPL expression in insects is restricted to neurons (Martin-Morris and White, 1990), these results indicate that full-length APPL selectively interacts with endogenously expressed Goα but not other G proteins within the developing insect nervous system.

Goα interacts with transmembrane APPL and APP in the nervous system. A, Schematic of the experimental protocol: tissue lysates were immunoprecipitated with antibodies specific for either the N- or C-terminal domains of APPL and APP, then immunoblotted with antibodies specific for different G protein subunits. B–E, Western blots of Manduca embryonic lysates immunoprecipitated with anti-nAPPL or anti-cAPPL and immunoblotted with antibodies against different Gα-subunits. B, Embryonic lysate immunoprecipitated with anti-nAPPL and immunoblotted with anti-Goα. Input lane shows endogenous Goα levels in the lysates before immunoprecipitation; IgG IP shows a matched negative control immunoprecipitation. C, Embryonic lysate immunoprecipitated with anti-cAPPL and immunoblotted with anti-Goα. Input lane shows endogenous Goα levels in the lysates; IgY IP shows a matched negative control immunoprecipitation. The size of Manduca Goα is 41 kDa. D–E, Western blots of Manduca embryonic lysates immunoprecipitated with anti-cAPPL and immunoblotted for either Giα (D) or Gsα (E). Input lanes show endogenous Giα and Gsα levels in the lysates before immunoprecipitation. Neither Giα nor Gsα coimmunoprecipitate with anti-cAPPL or with control IgY. The size of Giα is ∼41 kDa; Gsα is detected as a doublet at 48 and 52 kDa (Copenhaver et al., 1995). F–G, Western blots of immunoprecipitated Drosophila head lysates. F, Drosophila head lysates from wild-type and APPLd flies (which lack APPL expression) immunoprecipitated with anti-cAPPL and immunoblotted with anti-Goα. Inputs show equivalent levels of endogenous Goα in both fly lines. Goα was coimmunoprecipitated with anti-cAPPL antibodies from wild-type lysates but not with control IgY, nor from APPLd lysates. G, Western blot of lysates from Appld flies overexpressing mutant forms of APPL under the control of the GMR promoter. Applsd flies (“secretion-deficient”) express a transmembrane form of APPL lacking the juxtamembrane domain that is normally cleaved by secretases; ApplsdΔCg flies express secretion-deficient transmembrane APPL, which also lacks the putative Go-binding domain. Fly head lysates were immunoprecipitated with anti-Gi/oα and immunoblotted with anti-cAPPL (135 kDa). Input lanes show abundant expression levels of APPLsd and APPLsdΔCg; no APPL-related proteins were detected in APPLd flies. APPLsd but not APPLsdΔCg coimmunoprecipitated with Goα, indicating that the Go domain is necessary for APPL–Goα interactions in vivo. Appld flies served as the negative control. Probing for α-Goα verified that abundant levels of Goα were immunoprecipitated from each of the fly lines. Similar levels of Gβ also coimmunoprecipitated with Gi/o in all three fly lines, indicating equivalent expression of the heterotrimeric G protein complexes. The size of fly Gβ is ∼37 kDa. H–J, Western blot of mouse brain lysates immunoprecipitated with N-terminal- or C-terminal-specific APP antibodies and immunoblotted with antibodies targeting different G protein subunits. H, Immunoprecipitated mouse brain lysates labeled with anti-Goα. Input shows endogenous Goα levels in the lysate before immunoprecipitation. Lane 2: Goα coimmunoprecipitated with anti-nAPP. Lane 3: IgG = matched negative control immunoprecipitation. Lane 4: Goα also coimmunoprecipitated with anti-cAPP (8717; Fig. 2). Lane 5: Matched negative control immunoprecipitation with normal rabbit serum (NRS). I, Immunoprecipitated mouse brain lysates immunoblotted for Gsα. Input shows endogenous Gsα levels in the lysate before immunoprecipitation. Lanes 2–4 show that Gsα did not coimmunoprecipitate with anti-nAPP, control IgG, nor anti-cAPP. J, Mouse brain lysates immunoprecipitated with N-terminal- or C-terminal-specific APP antibodies and immunoblotted with α-Gβ. Input shows Gβ levels in the lysates before immunoprecipitation. Lanes 2–4: Gβ coimmunoprecipitated with both anti-nAPP and anti-cAPP antibodies (8717 and cAPP668). Lane 5: IgG-negative control immunoprecipitation. K, Immunoprecipitated human brain lysates immunoblotted for Goα. Input shows endogenous Goα levels in the lysate before immunoprecipitation. Goα was coimmunoprecipitated with anti-nAPP but not with control IgG.
Previous experiments in Drosophila have shown that flies lacking APPL (Appld) exhibit defects in synaptic differentiation and aberrant adult behavior (Luo et al., 1992; Torroja et al., 1999a), supporting a role for APPL in regulating neuronal growth and target innervation. To complement our studies of APPL-Goα interactions in Manduca, we used fly head lysates to show that Drosophila Goα could be coimmunoprecipitated with anti-cAPPL antibodies from wild-type controls but not Appld flies (Fig. 3F), although both fly strains express equivalent levels of Goα (Fig. 3F, input). As noted above, Nishimoto et al. (1993) identified a candidate Go-binding domain within the cytoplasmic domain of human APP695 (His657-Lys676) that was required for APP-Goα interactions in reconstituted liposomes (Okamoto et al., 1995). To determine whether the conserved Go-binding domain is required for APPL–Goα interactions in the insect nervous system, we used transgenic fly lines expressing modified forms of APPL in the absence of wild-type protein. APPLsd lacks the proteolytic cleavage site in APPL (amino acids 758–791) and consequently is expressed only as a transmembrane protein; APPLsdΔCg also lacks a portion of the putative Go-protein binding site (amino acids 845–855; Torroja et al., 1999b). Using the GMR promoter, UAS constructs of each isoform were expressed in the eyes of Appld flies, and head lysates were immunoprecipitated with anti-Goα. Blotting for α-cAPPL revealed that Goα interacts with APPLsd but not APPLsdΔCg (Fig. 3G), although both constructs were expressed at similar levels (Fig. 3G, input). Appld fly head lysates were used as a negative control. Probing the immunoprecipitates for Goα and Gβ verified that both subunits were abundantly expressed in all of the fly lines used for this analysis (Fig. 3G, IP). These results demonstrate that Goα interacts with transmembrane forms of APPL, consistent with its role as a Goα-associated receptor, and that the conserved Go-binding domain within APPL is necessary for its interactions with Goα in vivo.
We also investigated whether endogenously expressed Goα interacts with full-length APP in the mammalian nervous system. Using similar methods, we found that Goα could be readily coimmunoprecipitated with both nAPP- and cAPP-specific antibodies from lysates prepared from cortical and hippocampal regions of adult mouse brain, compared with control IgG and normal rabbit serum (Fig. 3H), supporting the model that Goα interacts with full-length APP in neurons. In contrast, none of the APP-specific antibodies that we tested were found to coimmunoprecipitate Gsα (Fig. 3I), suggesting that Gsα does not normally interact with full-length APP or its C-terminal cleavage products in the brain. Whether elevated expression of APP or its fragments can promote this interaction under pathological conditions remains to be determined.
If APP and its orthologs function as authentic Goα-coupled receptors, they would be expected to interact with heterotrimeric Go(αβγ) complexes in their inactive state (Bourne et al., 1991; Galés et al., 2006), whereas activation of APP signaling should induce their dissociation. Conversely, if APP and APPL function as downstream targets for activated Goα, we would not expect them to associate with the αβγ trimer. As shown in Figure 3J, antibodies against both the N- and C-terminal domains of APP coimmunoprecipitated Gβ-subunits as well as Goα, similar to previous reports using extracted membrane preparations (Nishimoto et al., 1993). Last, based on recent studies suggesting that APP–Goα interactions might be perturbed in AD patients (Shaked et al., 2009), we also used human brain lysates prepared from the cortical regions of healthy control subjects (provided by the Oregon Brain Bank) to show that anti-nAPP antibodies also coimmunoprecipitated human Goα, compared with the low background levels of Goα immunoprecipitated by control IgG (Fig. 3K). In combination, these results indicate that both insect APPL and mammalian APP selectively interact with heterotrimeric Go (but not other Gα proteins) under physiological conditions, consistent with an evolutionarily conserved role for APP-Goα signaling in the developing and mature nervous system.
APPL directly binds Goα both in vitro and in vivo
The model that APPL functions as a Goα-associated receptor requires direct contact between the two proteins at the plasma membrane. To address this issue, we used BiFC, a modified “split-GFP” approach whereby complementary fragments of Venus Fluorescent Protein (Vn) are fused to proteins of interest, but they remain nonfluorescent unless brought together by direct interactions between the tagged proteins (Kerppola, 2008; Robida and Kerppola, 2009). For these assays, we fused the N-terminal portion of Venus Fluorescent Protein (Vn1) to the cytoplasmic domain of Manduca APPL (APPL-Vn1) and the complementary C-terminal portion (Vn2) to Goα (Vn2-Goα). As an initial test of these constructs, we transfected them either singly or in combination into COS7 cells, which do not express detectable levels of mammalian Goα or APP (J.M. Ramaker, T.L. Swanson, and P.F. Copenhaver, unpublished observations). After 18–24 h, we then fixed and immunostained the cells with antibodies specific for APPL and Goα to monitor their expression levels (using Alexa Fluor 568- and Alexa Fluor 647-coupled secondary antibodies), independent of BiFC-induced fluorescence (visualized in the green channel). As shown in Figure 4, COS7 cells expressing either APPL-Vn1 (Fig. 4A) or Vn2-Goα alone (Fig. 4B) showed strong immunoreactivity for the individual fusion constructs but produced no detectable BiFC signal (Fig. 4A3,B3). In contrast, cells that coexpressed both APPL-Vn1 and Vn2-Gοα exhibited robust BiFC signals throughout their somata and growing processes (Fig. 4C3). These results demonstrate that APPL can directly bind Goα in exogenous cells.

APPL and Goα directly interact in cell culture. A–D, Representative images of COS7 cells at 18–24 h post-transfection, immunostained with anti-Goα (left columns) and anti-APPL (middle columns) to detect constructs of interest. Right-hand columns show green fluorescent BiFC signals that were produced by direct interactions between Vn1- and Vn2-tagged fusion proteins. A, B, Cell transfected with either APPL-Vn1 alone (A3) or Vn2-Goα alone (B3) did not emit detectable BiFC signals. C, Cell cotransfected with both APPL-Vn1 and Vn2-Goα exhibited BiFC signals at the plasma membrane and throughout growing processes (C3, arrowheads). D, Cell cotransfected with Vn2-Goα and APPLΔGo-Vn1 (APPL lacking the putative Go-binding domain; amino acids 762–791) produced only minimal BiFC signals that were restricted to the Golgi/ER regions. No detectable BiFC signals were present at the plasma membrane or growing processes of the cell (D3, arrowheads), despite the expression of both constructs in these regions (D1, D2). E–G, Lower magnification images of COS7 cells cotransfected with APPL-Vn1 plus equivalent levels of different Vn2-tagged Gα subunits. Forty-eight hours post-transfection, cells were coimmunostained with a Vn2-specific anti-GFP antibody and with anti-APPL (to detect APPL-Vn1). E, Cotransfection of APPL-Vn1 plus Vn2-Goα produced robust BiFC signals (C3), indicating direct APPL-Goα binding (arrowheads). F, Cotransfection of APPL-Vn1 with Vn2-Giα produced only minimal BiFC signals that were primarily localized within the ER/Golgi regions (F3). BiFC signals were not apparent in the plasma membrane or growing processes of the cells, despite the presence of both Vn-tagged constructs in these regions (F1, F2). G, Coexpression of APPL-Vn1 and Vn2-Gsα did not result in any detectable BiFC signals (G3), although both proteins were expressed throughout the cells (G1, G2). Scale bars: 10 μm.
Based on our evidence that the Go-binding domain of APPL is necessary for its association with APPL in vivo (Fig. 3G), we also tested whether this domain was necessary for direct APPL-Goα interactions in our BiFC assays by cotransfecting COS7 cells with Vn2-Goα and APPLΔGo-Vn1 (a form of Manduca APPL lacking the Go-binding domain, amino acids 762–791; Fig. 2B, dotted line). In contrast to the membrane-associated BiFC signals seen in cells that expressed full-length APPL-Vn1 plus Vn2-Goα (Fig. 4C3, arrowheads), cells that coexpressed APPLΔGo-Vn1 plus Vn2-Goα exhibited only minimal BiFC signals that were confined to the Golgi/endoplasmic reticulum (ER) regions (Fig. 4D3), despite the presence of both Vn-tagged constructs in more peripheral regions of the cells (Figs. 4D1,D2, arrowheads). To directly compare BiFC fluorescence in multiple cells transfected with APPL or APPLΔGo, we identified cells with relatively low levels of the Venus-tagged proteins (to avoid saturation in the BiFC channel) and analyzed the BiFC signal as a ratio to the levels of each construct. Quantification of the relative fluorescent intensities in each channel verified that coexpression of Vn2-Goα with full-length APPL-Vn1 produced significantly stronger BiFC signals than coexpression with APPLΔGo-Vn1 (Table 1). These results show that the Go-binding domain previously identified in human APP695 is also required for direct interactions between APPL and Goα.
Coexpression of APPL plus Goα in COS7 cells produces significantly greater BiFC signals than APPLΔGo plus Goα
To test whether APPL selectively interacts with Goα but not other related G proteins, we also cotransfected COS7 cells with APPL-Vn1 and Vn2-tagged constructs of Giα or Gsα. For these assays, we fixed the transfected cells after 48 h in culture, and then immunostained them with anti-APPL and an anti-GFP antibody that only recognizes the Vn2 domain; this approach allowed us to confirm that all of the Vn2-tagged Gα constructs were expressed at comparable levels. As in the previous experiment, cotransfection of APPL-Vn1 with Vn2-Goα produced strong BiFC signals throughout the cells and their growing processes (Fig. 4E3, arrowheads). In contrast, cotransfection of APPL-Vn1 with Vn2-Giα produced only faint BiFC signals that were predominantly restricted to Golgi/ER regions (Fig. 4F3, arrowheads), despite the presence of both proteins at the plasma membrane and in growing processes (Fig. 4F1–F3). Similarly, cells that coexpressed APPL-Vn1 with Vn2-Gsα exhibited no detectable BiFC signals (Fig. 4G3). These results indicate that APPL directly binds Goα but not other related G proteins in cell culture, consistent with the model that APPL selectively interacts with Goα at the plasma membrane.
To complement our analysis of APPL–Goα interactions in vitro, we also used the UAS-GAL4 system to express APPL-Vn1 and Vn2-Goα within the fly nervous system. For these experiments, we induced expression of UAS-APPL-Vn1 and UAS-Vn2-Goα using the eye-specific GMR-GAL4 promoter construct. Developing eye discs were then collected at progressive stages of development, fixed, and immunostained with a polyclonal anti-GFP antibody that recognized both Vn1 and Vn2 (visualized with Alexa Fluor 568-conjugated secondary antibodies), allowing us to monitor their expression levels in addition to the BiFC signals that they produced (Fig. 5A–C). When we expressed either APPL-Vn1 (Fig. 5A1) or Vn2-Goα alone (Fig. 5B1), we could readily detect the constructs in developing adult photoreceptors within the eye discs, but (as expected) no BiFC signals were produced (Fig. 5A2,B2). In contrast, coexpression of APPL-Vn1 with Vn2-Goα resulted in robust BiFC signals within the photoreceptor membranes and their axonal projections extending into the developing adult brain (Fig. 5C2). These results demonstrate that APPL and Goα directly bind when coexpressed in vivo, similar to our results in COS7 cells.

APPL and Goα directly interact in Drosophila photoreceptors and developing synapses. Isolated eye discs from third instar larvae expressing UAS-APPL-Vn1, UAS-Vn2-Goα, or both constructs (controlled by GMR-GAL4). Discs were fixed and immunostained with α-GFP (Aves #GFP1020) to label both Vn1 and Vn2 epitopes (visualized with Alexa Fluor 568 secondary antibodies); BiFC signals produced by APPL–Goα interactions were imaged in the green channel. A1, B1, Eye discs expressing either APPL-Vn1 or Vn2-Goα alone did not exhibit detectable BiFC signals (A2, B2). C1, Coexpression of APPL-Vn1 and Vn2-Goα resulted in robust BiFC signals throughout the fly photoreceptors (C2) and their axonal projections extending into the optic stalk (arrowheads). Small white boxes in the low-magnification images indicate highlighted regions shown in the insets of each panel. D, E, Isolated eye disc-brain complexes from mid-third instar Drosophila larvae that expressed Vn2-Goα plus either wild-type APPL-Vn1 or APPLΔGo-Vn1. Magenta outlines demarcate the proximal brain lobe (br) in each preparation. D1, E1, Eye disc–brain complexes from mid-third instar larvae coexpressing Vn2-Goα plus mutated or wild-type forms of APPL-Vn1. Immunostaining for anti-GFP showed that the Venus-tagged constructs were expressed in the developing adult photoreceptors (adjacent to the morphogenetic furrow; arrowheads) and throughout Bolwig's nerve (bn), which carries the axons of 12 larval photoreceptors through the eye disc into the larval optic neuropil of the brain (lon; arrows). Larval photoreceptors are out of the field of view. D2, The combined expression of APPL-Vn1 and Vn2-Goα produced robust BiFC signals in regions where both proteins were coexpressed (indicative of direct APPL–Goα interactions), including the synaptic projections of the larval photoreceptors in the lon. E2, Coexpression of APPLΔGo-Vn1 plus Vn2-Goα produced no detectable BiFC signals, despite comparable expression levels for both APPL constructs in D and E (data not shown). D3, Schematic representation of the late third instar larval eye disc–brain complex (equivalent to dissected preparations in D and E), illustrating the orientation of bn and the lon. Arrowheads indicate the morphogenetic furrow; dotted outline in the br indicates the adult optic anlage (oa), forming adjacent to the lon. F, Eye disc–brain complex from a white pupa expressing APPL-Vn1 plus Vn2-Goα. F1, Immunostaining with anti-GFP revealed the expression of the Venus-tagged constructs in developing adult photoreceptors, their projecting axons in the optic stalk (os), and their differentiating synaptic terminals within the lamina cortex (la) of the brain. F2, Magnified image of the boxed region in F1 (compressed image of 16 optical sections) to highlight the presence of BiFC signals in the photoreceptor axons within the os and their synaptic terminals in the la. F3, Schematic representation of the eye disc–brain complex at the white pupal stage (equivalent to preparation in F1), illustrating the orientation of the os and la regions; small oblong circle (gray) indicates the position of the residual lon (not visible in F1 and F2). Scale bars: (in A–C) 25 μm; (in A–C inset boxes) 15 μm; D–F1, 30 μm; (in F2) 10 μm.
To explore whether APPL–Goα interactions could be visualized in synaptic regions within the brain, we also examined younger third instar larvae, when GMR-induced expression can also be detected in 12 bilaterally paired larval photoreceptors that project into the brain (Steller et al., 1987; Hartenstein, 1988). As shown in Figure 5D, BiFC signals produced by coexpression of APPL-Vn1 with Vn2-Goα could be readily detected in larval photoreceptor axons extending through the developing eye disc via Bolwig's nerve and into the larval optic neuropil (Fig. 5D2), where they make synaptic contacts with several classes of interneurons (Tix et al., 1989; Sprecher et al., 2011). In contrast, when we coexpressed UAS-Vn2-Goα with UAS-APPL ΔGoVn1 (lacking the Go binding domain of APPL) at similar levels, no detectable BiFC signals were produced (Fig. 5E); again demonstrating that this domain is required for APPL-Goα interactions in vivo. At slightly later stages (white pupae), BiFC signals also became apparent within the developing synaptic terminals of adult photoreceptors coexpressing APPL-Vn1 and Vn2-Goα in the lamina cortex (Fig. 5F). These results confirm our immunohistochemical evidence that endogenously expressed Goα colocalizes with APP family proteins in developing neurons, providing additional support for the model that APP-Goα signaling may play a functional role in regulating key aspects of neuronal motility.
APPL–Goα interactions are regulated by Goα activity
A prediction from the model that APPL functions as an authentic G protein-coupled receptor (GPCR) is that APPL should preferentially bind inactive Goα as part of a heterotrimeric complex, whereas activation of Goα should promote its dissociation from APPL (Fig. 6A). To address this issue, we use our coimmunoprecipitation assays to determine whether stimulating G protein activity affected basal APPL-Goα interactions. As an initial test, we prepared lysates from Manduca GV1 cells (which endogenously express both proteins) and verified that APPL and Goα could be readily coimmunoprecipitated with anti-cAPPL antibodies (Fig. 6B, lane 1; histogram values in Fig. 6B were calculated by normalizing coimmunoprecipitated Goα levels relative to untreated controls). In contrast, treating replicate samples with GTPγS, a nonhydrolyzable GTP analog that activates all G proteins (Stryer and Bourne, 1986), reduced APPL-Goα interactions in a concentration-dependent manner (Fig. 6B, lanes 2–4). Treating lysates with GDPβS, which inhibits G protein activation, slightly increased Goα-APPL interactions (Fig. 6B, lane 6). These results indicate that APPL preferentially binds inactive but not active Goα, consistent with the hypothesis that APPL may function as a Goα-associated receptor rather than a downstream target of Goα signaling.

APPL–Goα interactions are regulated by Goα activity. A, Schematic of the model that inactive Goα binds APPL as part of a heterotrimeric complex with Gβγ. Activation of Goα (normally due to the exchange of bound GDP for GTP) promotes its dissociation from APPL, whereas inhibiting Goα prevents this dissociation, resulting in an increase in basal APPL–Goα interactions. B, Replicate cultures of Manduca GV1 cells (which endogenously express APPL and Goα) were treated as indicated, then lysed, immunoprecipitated with α-APPL, and immunoblotted with anti-Goα. Relative levels of coimmunoprecipitated Goα were normalized to “no treatment” control run in parallel (lane 1). Stimulating G protein activity with GTPγS (a nonhydrolyzable activator of G proteins) led to a concentration-dependent reduction in the amount of Goα that coimmunoprecipitated with APPL (lanes 2–4). IgY = matched negative control immunoprecipitation (lane 5). Treatment with GDPβS (a nonhydrolyzable G protein inhibitor) caused a modest increase in APPL–Goα interactions (lane 6). C, Lysates from staged groups of embryos (60–65 hpf) were treated as indicated, immunoprecipitated with α-APPL, and immunoblotted with anti-Goα. Gel shows a representative Western blot of coimmunoprecipitated Goα (upper blot) and APPL (lower blot; labeled with anti-nAPPL). Double arrow indicates the immature (smaller) and mature (larger) forms of full-length APPL. Goα levels were calculated as a ratio of immunoprecipitated full-length APPL in the same sample, and ratios were normalized to untreated controls (lane 1). Treatment with 1.0 μm GTPγS decreased APPL–Goα interactions (lane 2). Treatment with Mas 7 (to activate Giα/Goα) decreased APPL–Goα interactions in a concentration-dependent manner (lanes 3–4). In contrast, inhibiting Goα with PTX enhanced APPL–Goα interactions (lanes 5–6). Treatment with 20 μm GDPβS caused only a minor increase in coimmunoprecipitated Goα levels (lane 7). IgY = negative control immunoprecipitation (lane 8). D, Combined analysis of multiple experiments in which Manduca embryonic lysates were treated with G-protein-specific reagents, then immunoprecipitated with anti-cAPPL and immunoblotted with anti-Goα. Each manipulation was repeated in at least three independent experiments and normalized to their respective untreated controls in the same assay. Treatment with both GTPγS and Mas 7 significantly reduced APPL–Goα interactions. Mas 17, an inactive mastoparan analog, had no significant effect, compared with controls. PTX induced a concentration-dependent increase in APPL–Goα interactions. Although treatment with GDPβS caused a slight increase in APPL–Goα interactions in some experiments, overall this effect was not significant at the concentrations tested. IgY represents the species-matched negative control immunoprecipitations performed in each assay. Pairwise statistical analyses were performed between the control and each experimental group using Student's two-tailed t tests, *p < 0.05; **p < 0.01. Error bars indicate SEM.
A potential caveat to the foregoing experiments is that GDP/GTP analogs can modulate the activity of both heterotrimeric and monomeric G proteins (Stryer and Bourne, 1986) and might affect APPL-Goα interactions both directly and indirectly. We therefore performed a similar analysis with reagents that selectively target Goα, using lysates prepared from Manduca embryos at 60–70 hpf (during the active period of EP cell migration and outgrowth). Lysates were immunoprecipitated with anti-cAPPL, and the immunoblots labeled with both anti-Goα (Fig. 6C, upper gel) and anti-nAPPL (Fig. 6C, lower gel); the doublet labeled by the double arrows in Figure 6C indicates the immature and mature forms of full-length APPL, respectively (Swanson et al., 2005). For these experiments, relative levels of coimmunoprecipitated Goα were normalized to APPL levels in each sample, and then quantified as a ratio compared with untreated controls. Because insect APPL is exclusively expressed by neurons (Luo et al., 1990; Swanson et al., 2005), this approach provided a means of testing how Goα activity affects its association with endogenous APPL in the developing nervous system. As shown in Figure 6, C (lanes 1–2), treatment with GTPγS dramatically reduced the amount of Goα that could be coimmunoprecipitated with APPL, compared with untreated controls. Likewise, treatment with the Goα activator Mas 7 (a potent analog of Mastoparan) caused a substantial reduction in APPL–Goα interactions at all concentrations (Fig. 6C, lanes 3–4; and data not shown). Although Mas 7 will also activate Giα (Higashijima et al., 1990), our evidence that APP family proteins associate with Goα but not other G proteins (Fig. 3) suggests that the effects of Mas 7 were specific to Goα in this assay. In contrast, treatment with PTX caused a dramatic increase in the amount of Goα that coimmunoprecipitated with APPL (Fig. 6C, lanes 5–6). Since PTX inhibits insect Goα but not Giα (which lacks the C-terminal cysteine targeted for ADP-ribosylation; Thambi et al., 1989), these results indicate that the inactivation of Goα specifically inhibits its dissociation from APPL. None of these treatments significantly altered APPL levels (Fig. 6C, double arrowheads), indicating that the observed changes in APPL–Goα interactions were not caused by accelerated cleavage or degradation of the holoprotein. Figure 6D summarizes the combined results of multiple experiments using these methods, showing that both GTPγS and Mas 7 caused significant decreases in coimmunoprecipitated Goα. In contrast, treatment with PTX caused a concentration-dependent increase in APPL–Goα interactions, ranging from a slight enhancement at 1 μg/ml to highly significant increases at 2–4 μg/ml (Fig. 6D; and data not shown). In some preparations, GDPβS also caused an increase in the level of coimmunoprecipitated Goα, although this response was more variable and did not reach statistical significance at the concentrations used for these assays. In combination, these experiments support the model that stimulation of APPL signaling within neurons will induce the activation and release of Goα, similar to conventional GPCRs. They also suggest that under normal conditions, a low level of APPL activation coincides with EP cell migration and outgrowth, consistent with a role for APP family proteins in regulating motile responses to endogenous guidance cues.
APPL and Goα signaling regulates common aspects of neuronal migration
To test whether APPL-Goα signaling regulates specific aspects of neuronal motility in vivo, we used our well characterized embryo culture assay for neuronal migration in Manduca (Horgan et al., 1995; Coate et al., 2008). Developmentally synchronous embryos were opened in culture to expose the ENS before the onset of migration (at 52–53 hpf; Fig. 7A), and the EP cells were directly challenged with experimental reagents targeting either Goα or APPL. The embryos were allowed to develop in culture for an additional 24 h, and then fixed and immunostained with anti-Fas II to quantify the full extent of EP cell migration and axon outgrowth. As shown in Figure 7B, embryos treated with control medium during this period exhibited a normal pattern of migration and outgrowth that was predominantly restricted to the midgut muscle bands (Figure 7B, b), with relatively few neurons and processes extending onto the interband regions (Figure 7B, ib). Treating the EP cells with AlF4− to selectively stimulate heterotrimeric G protein activity (Sternweis and Gilman, 1982) curtailed their subsequent migration and outgrowth (Fig. 7C,E), similar to the effects of both GTPγS (which stimulates all G proteins) and mastoparan (specific for Goα and Giα; Horgan et al., 1994, 1995). In contrast, inhibiting Goα with PTX resulted in a distinctive pattern of ectopic migration and outgrowth by the EP cells onto the interband regions (Fig. 7D,F,G), as well as a moderate increase in their average distance traveled along the normal band pathways (Fig. 7E). To confirm the specificity of this effect, we also treated premigratory neurons with the A-Protomer of PTX (which lacks the lectin-like activity of the B-oligomer; Mangmool and Kurose, 2011), using α toxin to enhance cell penetration (as previously described; Horgan et al., 1995). As shown in Figure 7, F and G, this treatment induced a more dramatic increase in the number of ectopic neurons and neurites that extended into the interband regions, as well as a moderate but significant increase in migration and growth along the band pathways (Fig. 7E). Treatment with α toxin alone caused no significant effects (data not shown). These results are consistent with our previous evidence that Goα activity normally restricts the extent of EP cell migration in a Ca2+-dependent manner (Horgan and Copenhaver, 1998).

Inhibiting Goα activity in the EP cells induces ectopic migration and outgrowth. A–D, Representative camera lucida drawings of Manduca embryos in which the EP cells were treated before migration onset with reagents targeting Goα, grown in culture for another 24 h, then fixed and immunostained with anti-Fas II to reveal the full extent of migration and outgrowth in the developing ENS. A, Control embryo immunostained at the onset of an experiment (52 hpf) to show the initial positions of the premigratory EP cells adjacent to the FG/MG boundary. By this stage, subsets of neurons had begun to extend exploratory filopodia onto their future muscle band pathways (b) but had not commenced their migratory dispersal. B, Embryo opened at 52 hpf and treated with control culture medium throughout 24 h of development showed the normal pattern of migration and outgrowth along the muscle bands, with only a few processes growing onto the interband musculature (ib). C, Treatment with 12.5 μm AlF4− (an activator of heterotrimeric G proteins) caused almost complete inhibition of migration and outgrowth, stalling the neurons at the FG/MG boundary. D, Treatment with 100 ng/ml PTX (a specific inhibitor of Goα in insects) induced a distinctive pattern of ectopic migration (open arrows) and outgrowth onto the interband regions (black arrowheads). E, Average distances of neuronal migration and axon outgrowth along band pathways for each treatment group (normalized to matched sets of control embryos in each experiment). F, Quantification of the average number of neurons per embryo that exhibited ectopic migration into each interband region for each treatment condition. G, Quantification of the average extent of ectopic axon outgrowth in the interband regions of embryos treated with various reagents. In E–G, pairwise statistical analyses were performed between each experimental group and their corresponding controls, using Student's two-tailed t tests, *p < 0.05; #p < 0.02; **p < 0.01; ***p < 0.001. N = at least 10 per condition. Error bars indicate SEM.
Based on reports that APP family proteins can regulate neuronal migration in other systems (Herms et al., 2004; Young-Pearse et al., 2007) and our evidence that APPL endogenously interacts with Goα in the developing ENS, we tested whether altering APPL expression in the EP cells induced similar defects in their migratory behavior. We first evaluated a panel of morpholinos targeting the coding domain of Manduca APPL mRNA for their effectiveness in knocking down APPL expression in GV1 cells. As shown in Figure 8A, one of these morpholinos (MO-APPL2) caused significant reductions in APPL levels that were both concentration-dependent and specific, almost completely eliminating APPL protein levels (Fig. 8B) without affecting Goα or tubulin expression (used as off-target controls; Fig. 8A,B). Staged embryos were then opened in culture at 48 hpf (7 h before migration onset) and treated with either MO-APPL2 or control morpholinos for 24 h, using Endo-Porter to facilitate their transport into developing neurons (Coate et al., 2008). Compared with the infrequent ectopic events seen in embryos treated with control morpholinos (Fig. 8G), knocking down APPL expression in the EP cells resulted in a pattern of excessive migration and outgrowth onto the interband regions (Fig. 8D,G,H), a phenotype strikingly similar to the ectopic growth induced by PTX treatment (Fig. 7D). Because our methods for visualizing the EP cells (with anti-Fas II immunostaining) precluded a simultaneous analysis of APPL expression, it was not possible to directly correlate the extent of ectopic growth by individual neurons and their residual APPL levels. Nevertheless, quantifying these effects for the entire population of EP cells indicated that inhibiting APPL expression resulted in the same distinctive pattern of ectopic growth and migration caused by inhibiting Goα activity.

Disrupting APPL signaling in the EP cells induces ectopic migration and outgrowth. A, Western blot of Manduca GV1 cells treated with APPL-specific morpholinos (MOs) for 48 h, then lysed and immunoblotted with anti-APPL to demonstrate the selective knock down of APPL expression. Immunoblotting the same gel with anti-tubulin served as a control for nonspecific effects of the MOs. Lanes 1–2: Replicate GV1 cell lysates treated with 20 μm standard control MOs (plus 0.6% Endo-Porter). Lanes 3–8: Replicate lysates treated with 0.6% Endo-Porter plus increasing concentrations of APPL MOs. B, Quantification of replicate experiments demonstrating the effectiveness of the APPL MOs at different concentrations (normalized to tubulin levels); treatment with 5 and 20 μm APPL significantly inhibited APPL protein levels. Quantification of Goα levels in the same samples (also normalized to tubulin) showed that Goα was unaffected by the MO treatments. C–H, Inhibiting APPL signaling in the EP cells phenocopies the effect of inhibiting Goα activity on ectopic migration and outgrowth. C–E, Camera lucida drawings of Manduca embryos in which the EP cells were treated before migration onset with reagents targeting APPL signaling, allowed to develop in culture for another 24–48 h, then fixed and immunostained with anti-Fas II to reveal the full extent of migration and outgrowth. C, Control embryo treated with culture medium exhibited a normal pattern of migration and outgrowth (same preparation shown in Fig. 7B). D, Treatment with APPL-specific MOs (50 μg/ml) induced the same overall pattern of ectopic migration and outgrowth caused by inhibiting Goα activity. E, Treatment with anti-nAPPL (1 μg/ml) also induced a similar phenotype. Open arrows in D and E indicate ectopic neurons in the interband regions; black arrowheads indicate ectopic processes. F, Average distances of neuronal migration and axon outgrowth along the band pathways for each treatment group (normalized to matched sets of control embryos in each experiment). G, Quantification of the average number of neurons that exhibited ectopic migration onto the interband regions for each treatment condition. H, Quantification of the average extent of ectopic axon outgrowth per interband region for each treatment condition. *p < 0.05; #p < 0.02; **p < 0.01; ***p < 0.001; Student's two-tailed t tests. N = at least 10 per condition. Error bars indicate SEM.
Although several candidate proteins have now been shown to interact with APP family proteins in different contexts (Ho and Südhof, 2004; Osterfield et al., 2008; Rice et al., 2012), authentic ligands that regulate their intrinsic signaling activity have yet to be identified. As an alternative strategy, we used our anti-nAPPL antibody (targeting the extracellular domain of APPL) to block endogenous interactions between APPL and its putative binding partners, as a means of testing whether APPL-dependent signaling is required for their normal pattern of migration and outgrowth. Accordingly, staged embryos were opened in culture shortly before the onset of migration (52–23 hpf), and the EP cells were directly treated with medium containing either anti-nAPPL or control IgG. Imaging these preparations after 24 h revealed the same pattern of ectopic growth and migration caused by PTX treatment and APPL-specific morpholinos (Figs. 8E,G,H), whereas none of these treatments had a significant effect on the extent of migration and axon growth along the band pathways (Fig. 8F). Thus, inhibition of either APPL or Goα activity produced an identical phenotype within the developing ENS, in which the migratory neurons and their processes traveled inappropriately into regions that were normally inhibitory to neuronal growth.
The dramatic effects caused by perturbing APPL signaling in our embryonic culture assays were particularly striking, compared with the relatively subtle phenotypes induced by the genetic deletion of APPL. In flies lacking APPL, the initial formation of the nervous system was surprisingly normal (Luo et al., 1992), although these animals displayed more subtle defects in axonal targeting and postsynaptic synaptic growth (Ashley et al., 2005; Mora et al., 2013), and they exhibited aberrant responses to both neurodegenerative insults and traumatic brain injury (Leyssen et al., 2005; Wentzell et al., 2012). Flies lacking APPL also exhibited behavioral and learning deficits that could be rescued by re-expression of APPL or human APP (Luo et al., 1992). These results are consistent with the model that APPL-dependent interactions modulate multiple aspects of neuronal motility throughout life. In comparison, genetic deletion of Goα in Drosophila caused more extensive abnormalities in neuronal growth and guidance, including errors in axonal guidance, fasciculation, and target innervation (Frémion et al., 1999), suggesting that additional guidance factors in addition to APPL normally contribute to the homeostatic control of Goα in the nervous system. In this manner, the local control of Goα activity within motile neurons may provide a convergent mechanism for integrating their responses to a range of attractive and repulsive stimuli (He et al., 2006; Bromberg et al., 2008). The dramatic patterns of ectopic growth caused by our acute manipulations of APPL in the ENS may therefore reflect a lack of compensatory modulation of Goα signaling over the relatively short time course of our embryonic culture assays.
In combination with previous evidence that local activation of Goα in the EP cells induces Ca2+-dependent filopodial retraction (Horgan and Copenhaver, 1998), we propose that APPL (and its vertebrate orthologs) can function as unconventional Goα-associated receptors that regulate the motile behavior of developing neurons (Fig. 9). By this model, transmembrane APPL normally trafficks to the leading processes of developing neurons, where it assembles into a signaling complex with Goα. Interactions between APPL on exploratory filopodia and unidentified ligands on adjacent cells induce the local activation and release of Goα, which in turn promotes Ca2-dependent filopodial retraction. Global stimulation of this pathway should therefore induce a collapse/stall response in migration (as caused by activators of Goα), whereas inhibition of either APPL or Goα should permit ectopic, inappropriate migration and outgrowth (as illustrated by the foregoing manipulations). In this manner, APPL and its vertebrate orthologs might contribute to the dynamic positioning of neurons and their processes within the developing nervous system and in regions of synaptic plasticity in the adult brain.

A model for the role of APPL-Goα signaling in the control of neuronal migration within the developing ENS. Transmembrane APPL is expressed in the motile processes of migrating EP cells, where it colocalizes with Goα. When exploratory filopodia extend off the band pathway (b) onto the adjacent interband regions (ib), they encounter ligands (as yet unidentified; black octagons) that induce APPL-dependent activation of Goα. In turn, local activation of Goα within the leading process induces Ca2+ influx via voltage-independent channels (Horgan and Copenhaver, 1998), resulting in filopodial retraction, thereby restricting inappropriate outgrowth and migration into these regions.
Discussion
APP family proteins as unconventional Goα-coupled receptors
Evidence that APP can function as a G protein-associated receptor originated from studies by Nishimoto et al. (Nishimoto et al., 1993; Okamoto et al., 1995), who used reconstituted phospholipid vesicles to show that clustered APP can activate Goα via a 20 aa sequence within its cytoplasmic domain. However, the physiological role of APP as an authentic regulator of Goα activity has remained unclear. We have now used several different approaches to show that endogenously expressed APPL (in insects) and APP (in mammals) interact with Goα in the nervous system. Simultaneous immunolabeling with antibodies against both the N- and C-terminal domains of APP and APPL indicated that the full-length forms of these proteins colocalize with Goα in regions of active motility, including the leading processes of migrating neurons and the growth cones of elongating neurites (Figs. 1, 2). Likewise, our coimmunoprecipitation analysis showed that both full-length APPL and APP interact with endogenously expressed Goα, in contrast to other related Gα-subunits (including Giα and Gsα; Fig. 3). Using genetically modified Drosophila lines, we demonstrated that the conserved Go-binding domain within APPL is required for this interaction (Fig. 3G), providing new support for the model proposed by Nishimoto et al (1993). A pharmacological analysis of APPL–Goα interactions in lysates from Manduca cells and embryos demonstrated that activation of Goα induced its dissociation from APPL, whereas inhibiting Goα enhanced APPL–Goα interactions (Fig. 6), paralleling the effects of similar manipulations targeting conventional GPCRs (Mangmool and Kurose, 2011).
Last, we adapted BiFC protocols to show that APPL directly interacts with Goα (but not other Gα-subunits) in transfected COS7 cells (Fig. 4) and within the fly nervous system (Fig. 5). Consistent with our coimmunoprecipitation assays, we found that the production of BiFC signals by APPL–Goα interactions required the conserved Go-binding domain in APPL (Figs. 4, 5). These results provide the first demonstration that APPL and Goα can directly bind in vivo, and they support the model that APP family proteins represent a distinct class of unconventional (single-transmembrane) Goα-coupled receptors (Patel, 2004).
APP family proteins and the control of neuronal migration
Although the normal functions of APP and its orthologs remain controversial, considerable evidence indicates that they can regulate multiple aspects of neuronal motility. Developmentally expressed APP (in mammals) and APPL (in insects) are upregulated in regions of active growth and migration (Luo et al., 1990; Clarris et al., 1995; Torroja et al., 1999a; Sabo et al., 2003; Swanson et al., 2005; Young-Pearse et al., 2007), while both the transmembrane holoproteins and their cleaved ectodomain fragments are capable of modulating cell adhesion responses, neurite arborization, and synaptogenesis (Qiu et al., 1995; Torroja et al., 1999a; Leyssen et al., 2005; Soba et al., 2005). In particular, several recent reports have indicated that APP may play an important role in controlling neuronal migration within the mammalian brain. However, different experimental strategies have produced conflicting results, possibly due in part to the overlapping activities of the related proteins APLP1 and APLP2 (Heber et al., 2000). Analogous to the rather subtle defects seen in flies lacking APPL, genetic deletion of APP alone produced no major abnormalities in mouse brain development, although these animals subsequently exhibited a variety of postnatal defects in dendritic growth, synaptic plasticity, and spatial learning (Sugaya et al., 1996; Dawson et al., 1999; Phinney et al., 1999). Even in mice lacking all three APP family proteins, many aspects of brain development were found to be essentially normal, except for a striking pattern of heterotopias resembling cobblestone lissencephaly (Herms et al., 2004), caused by excessive, inappropriate neuronal migration in the developing cortex (Devisme et al., 2012). Similar defects were also reported in mice lacking both Fe65 and Fe65L1 (Guénette et al., 2006), cytoplasmic adapter proteins that regulate APP trafficking and processing and may control the bioavailability of the holoprotein (McLoughlin and Miller, 2008; Dumanis et al., 2012). These results support the model that signaling by APP and its orthologs normally restricts the extent of neuronal migration, analogous to our results in Manduca. As with APPL in the insect nervous system, we postulate that other guidance cues in addition to APP regulate Goα activity within motile neurons in the developing mammalian brain, providing a mechanism for integrating local growth responses to a variety of environmental stimuli. Given our evidence that endogenously expressed APP and Goα interact in both murine and human neural tissue, it will be interesting to test whether the synaptic defects caused by the loss of APP family proteins can be linked to alterations in Goα-dependent signaling pathways.
In contrast to the heterotopias induced by genetic deletion of APP and its orthologs, interfering with APP expression in neuronal precursors by RNA interference resulted in the premature arrest of migration by their progeny (Young-Pearse et al., 2007), suggesting that APP normally promotes migration in response to permissive guidance cues in the developing cortical plate (Young-Pearse et al., 2008; Rice et al., 2012). Recent evidence demonstrating that members of the APP family also regulate the mitotic behavior of cortical progenitors (independent of their role in migration) may provide an explanation for these disparate results (Shariati et al., 2013). Since insect APPL is not expressed by developing neurons until after their terminal mitosis (Luo et al., 1990; Swanson et al., 2005), our manipulations in the embryonic ENS have specifically addressed its function during the migratory period of development.
By exploiting the comparative simplicity of Manduca as a model system, we have shown that APPL plays an important role in regulating the directionality of motile neurons within the nervous system. Knocking down APPL expression in the EP cells with morpholinos eliminated their normal inhibitory response to the interband regions of the ENS, resulting in a distinctive pattern of inappropriate migration and outgrowth (Fig. 8D,G,H). Similarly, treating the migratory neurons with antibodies against the extracellular domain of APPL induced the same ectopic phenotype (Fig. 8E), presumably by interfering with endogenous interactions between APPL and its functional ligands (as yet unidentified). An alternative explanation is that treatment with anti-APPL antibodies might have accelerated the internalization or clearance of APPL from the surface of the migrating neurons, which would produce the same effect. In comparison, EP cells that remained on the muscle bands in these preparations migrated correctly (Fig. 8F), indicating that APPL is not required for their response to permissive cues associated with their normal pathways (including Fas II; Wright et al., 1999). Intriguingly, APPL has also been postulated to restrict inappropriate nuclear migration by developing photoreceptor neurons in Drosophila, albeit via a more indirect signaling mechanism (Pramatarova et al., 2006). Whether overstimulation of APPL signaling in the EP cells will induce premature termination of migration (similar to Goα hyperactivation) remains to be determined. Nevertheless, these results support the model that APPL functions as a guidance receptor that transduces neuronal responses to local inhibitory cues, preventing developing neurons from growing into inappropriate regions of the nervous system.
The role of APP–Goα interactions in neural development and disease
Work on a number of model systems has demonstrated an important role for Goα in the control of neuronal growth and motility. Goα is the most abundant heterotrimeric G protein in the brain (Strathmann and Simon, 1990; Chen et al., 1999); it is enriched in growth cone membranes and migrating neurons (Chang et al., 1988; Strittmatter, 1992; Horgan et al., 1994; Bates and Meyer, 1996), and it regulates neurite outgrowth (He et al., 2006; Bromberg et al., 2008). In cultured chick neurons, PTX-sensitive Go/i proteins have been shown to transduce “collapse” responses in growth cones that encounter certain inhibitory cues, in part by promoting Ca2+-dependent filopodial retraction (Igarashi et al., 1993; Nakayama et al., 1999). The misregulation of this pathway may also affect regeneration responses within the mammalian CNS (Bates and Meyer, 1996). Previously, we showed that Goα stimulation in the EP cells inhibits their motility and causes premature stalling within the developing ENS (Fig. 7C), a response that is transduced via activation of voltage-independent Ca2+ currents (Horgan and Copenhaver, 1998). In contrast, inhibiting Goα resulted in the same characteristic pattern of inappropriate migration and outgrowth caused by interference with APPL-dependent responses (Fig. 8D,E).
Based on our evidence that Goα and APPL colocalize within the leading processes of the EP cells, that they are functionally coupled and directly interact via the Go-binding domain in APPL, and that inhibiting either Goα or APPL signaling produces the same distinctive phenotype of ectopic migration and outgrowth, we hypothesize that APPL functions as a Goα-coupled receptor that regulates neuronal motility in a variety of contexts. Moreover, our evidence that transmembrane APP selectively associates with Goα in mammalian neurons (Figs. 2R–T, 3H–K) suggests that this interaction is evolutionarily conserved. In the developing nervous system, this signaling pathway would provide a previously unrecognized mechanism for restricting inappropriate migration and outgrowth, a response that might also play a role in synaptic remodeling (complementing the model proposed by Ashley et al., 2005). In the context of AD, several intriguing studies have suggested that the misregulation of APP-Goα signaling may also play a role in progressive neurodegeneration. The neurotoxic effects of FAD-associated mutations in APP were previously shown to hyperactivate Goα in cell culture, whereas blocking Goα signaling could prevent the apoptotic effects of disease-associated APP isoforms (Okamoto et al., 1996; Yamatsuji et al., 1996). More recently, studies using human brain samples revealed that both elevated G protein activity and decreased APP-Goα interactions correlated with the severity of AD symptoms (Reis et al., 2007; Shaked et al., 2009; Sola Vigo et al., 2009), while in vitro assays showed that Aβ peptides could disrupt APP–Goα interactions, resulting in the hyperactivation of Goα and aberrant Ca2+ influx (Reis et al., 2007; Shaked et al., 2009; Sola Vigo et al., 2009). These results are also consistent with evidence that the dysregulation of neuronal Ca2+ may be an important factor in initiating the neurodegenerative responses that typify AD (Khachaturian, 1987; LaFerla, 2002). Further investigation of the normal processes controlled by APP-Goα signaling (including Ca2+-dependent synaptic growth and remodeling) may therefore provide the framework for testing whether the misregulation of this pathway contributes to a variety of age-related neurological conditions.
Footnotes
Support for this work was provided by National Institutes of Health (NIH) RO1 AG025525 and R21 NS080036 (P.F.C.), and by a grant from the Oregon Partners for Alzheimer's Research to J.M.R., who is a Vertex Scholar. The Oregon Brain Bank is supported by the Layton Aging and Alzheimer's Disease Center, NIH P30 AG8017. Confocal imaging was supported by NIH P30 NS061800. We thank Drs. Doris Kretzschmar, David Morton, and Larry S. Sherman for critical input on this manuscript. We also thank Drs. Vivian Budnik, Michael Gorczyca, and Kalpana White for generously sharing their APPL antibodies, fly lines, and helpful suggestions. Primary cultures of embryonic rat hippocampal neurons were provided by Dr. Gary Banker and Ms. Barbara Smoody, Jungers Center, Oregon Health and Science University (OHSU), Portland, OR. Mouse brain samples were kindly provided by Dr. Joseph Quinn and Christopher Harris (Department of Neurology, Portland Veterans Administration Medical Center, and Layton Center for Aging and Alzheimer's Disease Research, OHSU). Frozen samples of human brain tissue were made available by the Oregon Brain Bank (following established protocols), with the generous assistance of Dr. Randall L. Woltjer (Department of Pathology, OHSU). We thank Dr. Stephen Michnick (University of Montreal) for providing plasmids containing the coding domains for complementary portions of Venus Fluorescent Protein (Vn1 and Vn2). We Are grateful to Dr. Stefanie Kaech and Aurelie Snyder for assistance with the confocal imaging analysis that was performed in the Advanced Light Microscopy Core, Jungers Center, OHSU.
The authors declare no competing financial interests.
- Correspondence should be addressed to Dr. Philip F. Copenhaver, Cell and Developmental Biology L-215; Oregon Health and Science University; 3181 SW Sam Jackson Park Road; Portland, OR, 97239. copenhav{at}ohsu.edu





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
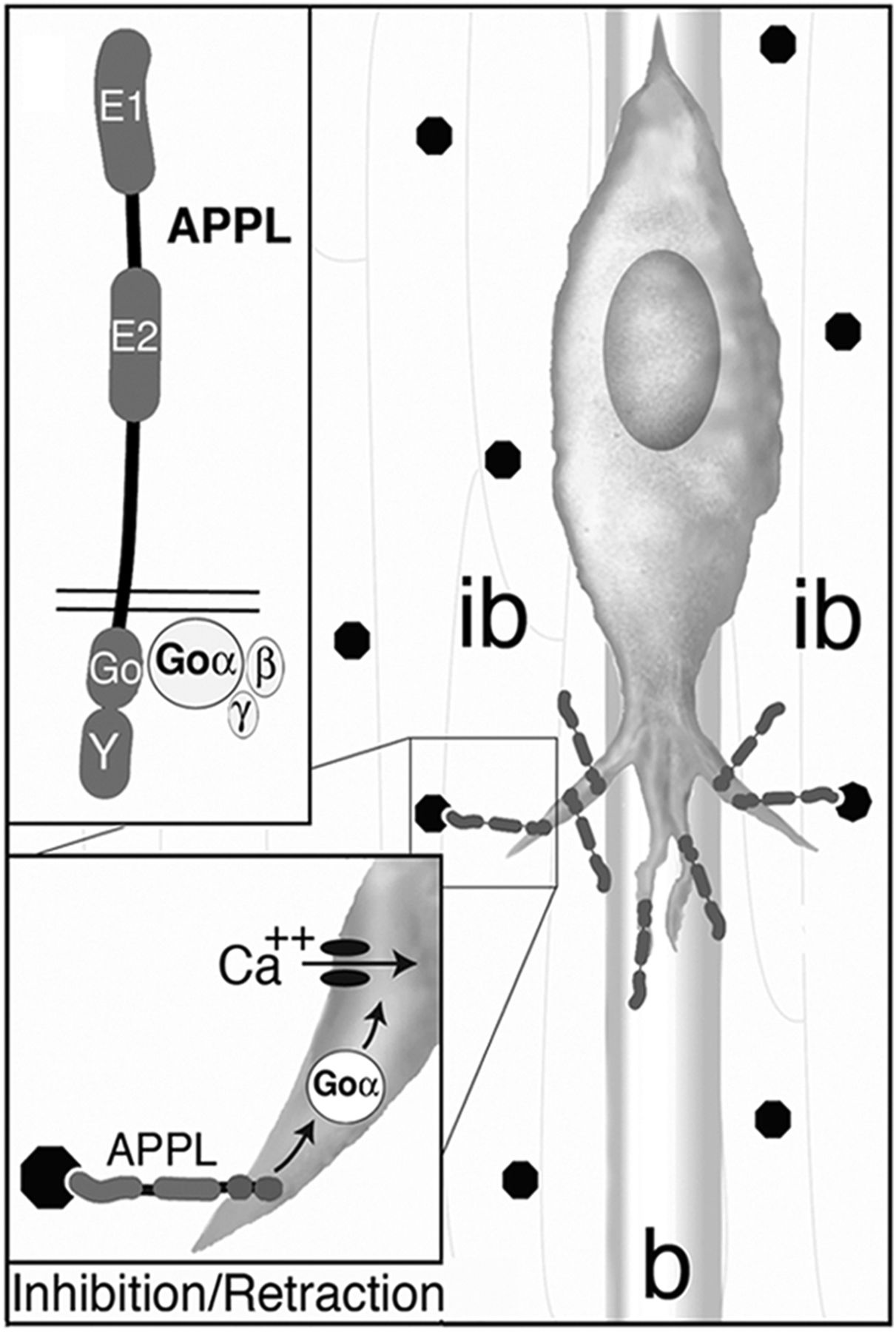{kind=link}