

Abstract
Synaptic signals from retinal bipolar cells were monitored by measuring EPSCs in ganglion cells voltage-clamped at −70 mV. Spontaneous EPSCs were strongly suppressed byl-2-amino-4-phosphonobutyrate (AP-4), an agonist at group III metabotropic glutamate receptors (mGluRs). Agonists of group I or II mGluRs were ineffective. AP-4 also suppressed ganglion cell EPSCs evoked by bipolar cell stimulation using potassium puffs, sucrose puffs, or zaps of current (0.5–1 μA). In addition, AP-4 suppressed Off EPSCs evoked by dim-light stimuli. This indicates that group III mGluRs mediate a direct suppression of bipolar cell transmitter release. An mGluR antagonist, (RS)-α-cyclopropyl-4-phosphonophenylyglycine (CPPG), blocked the action of AP-4. When bipolar cells were weakly stimulated, AP-4 produced a large suppression of the EPSC, but CPPG alone had little effect. Conversely, when bipolar cells were strongly stimulated, CPPG produced an enhancement of the EPSC, but AP-4 alone had little effect. This indicates that endogenous feedback regulates bipolar cell transmitter release and that the dynamic range of the presynaptic metabotropic autoreceptor is similar to that of the postsynaptic ionotropic receptor. Furthermore, the feedback is rapid and intensity-dependent. Hence, concomitant activation of presynaptic and postsynaptic glutamate receptors shapes the responses of ganglion cells.
In a variety of systems, synaptic release is altered by the activation of presynaptic metabotropic glutamate receptors (mGluRs) (for review, see Miller, 1998; Anwyl, 1999). These receptors suppress transmitter release by affecting calcium or potassium channels (Saugstad et al., 1996; Takahashi et al., 1996) or by a direct action on the proteins involved in transmitter release (Scanziani et al., 1995). They can also enhance transmitter release by modulation of calcium release from internal stores (Peng, 1996; Cochilla and Alford, 1998).
To date, eight mGluRs have been cloned and divided into three groups based on sequence homology and pharmacology (Pin and Duvoisin, 1995). Different subcellular distributions of mGluRs have been reported, suggesting they have different functions at the synapse. Group I receptors often appear restricted to the postsynaptic terminal (Martin et al., 1992; Koulen et al., 1997; Lujan et al., 1997), although exceptions have been noted (Cochilla and Alford, 1998). Group II and III receptors are generally presynaptic (Shigemoto et al., 1997). Group II receptors are found to have a diffuse distribution in the periterminal regions of the membrane (Petralia et al., 1996; Shigemoto et al., 1996; Yokoi et al., 1996; Lujan et al., 1997; Shigemoto et al., 1997). Localization outside of the synaptic active zone has led to the hypothesis that these receptors are activated only by prolonged and enhanced synaptic stimulation that produces glutamate “spillover” to perisynaptic sites (Asztely et al., 1997; Scanziani et al., 1997). Consistent with this scheme, group II-mediated inhibition is more apparent when the glutamate concentration in the cleft is artificially raised by pharmacological blockade of glutamate transporters (Maki et al., 1994; Scanziani et al., 1997; Dube and Marshall, 2000).
In contrast, group III receptors are selectively localized near the transmitter release site (Brandstätter et al., 1996; Li et al., 1997; Shigemoto et al., 1997), suggesting that these mGluRs may serve as autoreceptors activated exclusively by synaptically localized glutamate release.
The bipolar to ganglion cell synapse in the retina is well suited to examine the properties of metabotropic glutamate receptors. This synapse can be activated by natural or artificial stimuli, and the release properties of the synapse are graded, robust, and well characterized. We found that the output of this synapse is regulated by mGluR-negative feedback, that the autoreceptor can be rapidly activated (<20 msec), and that the dynamic range of the feedback is similar to that of the feedforward synapse.
MATERIALS AND METHODS
Slice preparation. Larval tiger salamanders (Ambystoma tigrinum) were obtained from Kons Scientific (Germantown, WI) and Charles Sullivan (Nashville, TN) and were kept in tanks maintained at 4°C on a 12 hr light/dark cycle. Retinal slices were prepared as described by Wu (1987). All procedures were performed in accordance with the United States Animal Welfare Act and theNIH Guide for the Care and Use of Laboratory Animals(publication #85–23) and were approved by the Animal Care Committee of the State University of New York. In dim red light, animals were decapitated and double-pithed; eyes were enucleated. The retina was removed from the eyecup, placed on a 0.45 μm pore membrane filter (Millipore, Bedford, MA) with the photoreceptor side down, and subsequently sliced at 150–250 μm intervals using a tissue slicer (Stoelting, Woods Lane, IL). A single slice was then transferred to the recording chamber and viewed under infrared light using a CCD camera attached to an upright Olympus IMT2 fluorescent microscope, equipped with a 40× water-immersion lens (Zeiss, Thornwood, NY).
In a few experiments, a reduced retinal slice preparation was used, containing only cell bodies in the amacrine and ganglion cell layers along with an intact inner plexiform layer (IPL). Slices were made with the vitreal side facing the filter paper, and a perfusion pipette was placed behind the filter paper to direct the flow of Ringer's solution from the ganglion cell layer to the photoreceptor layer. Next, SDS detergent (1–5%) was applied to the outer retina. This dissolved away the outer retinal layers while the perfusion system blew the debris away, protecting the inner retina.
The slice was continually bathed with control Ringer's solution containing (in mm): 111 NaCl, 2.5 KCl, 1.8 CaCl2, 1 MgCl2, 10 dextrose, and 5 HEPES buffered to pH 7.8. Using a gravity-fed perfusion system, Ringer's solution could be exchanged with drug-containing solutions within 5–10 sec. S-3,5-Dihydrophenylglycine (DHPG), (2S,1′R,2′R,3′R)-2-(2′-3′-dicarboxycyclo-propyl)-glycine (DCG-IV), l-2-amino-4-phosphonobutyrate (AP-4), and (RS)-α-cyclopropyl-4-phosphonophenylglycine (CPPG) were obtained from Tocris Cookson (Ballwin, MO). All other chemicals were obtained from Sigma (St. Louis, MO).
Whole-cell patch-clamp recordings. Recordings were made from neurons in the ganglion cell layer of the retinal slice preparation. All data were from neurons that exhibited transient responses at light onset and/or offset. Neurons were clamped at −70 mV to isolate EPSCs. This was confirmed by testing the effect of GABA, which reversed at this potential (W. Shen, State University of New York, Buffalo, NY personal communication). Whole-cell recordings were made using ∼5 MΩ electrodes containing (in mm): 105 K-gluconate, 5 KCl, 1 MgCl2, 1 BAPTA, 10 HEPES, 4 ATP Na2, 0.5 GTP Na3, and 1% Lucifer yellow. The pH was adjusted to 7.4 with KOH. The voltage- and current-clamp recordings were made with an Axopatch 200B amplifier (Axon Instruments, Foster City, CA). Analog signals were filtered at 1 kHz and sampled at 2 kHz with the Digitadata 1200 analog-to-digital board (Axon Instruments). Clampex8 (pClamp8; Axon instruments) was used to control the voltage command outputs, acquire data, and trigger the light stimulus. The currents and voltages shown are raw data, and electrode junctional potentials and access resistances were not corrected.
Stimulation paradigms. Responses in ganglion cells were evoked by light stimuli or by artificially stimulating bipolar cells. For light stimulation, a red Stanley light-emitting diode (LED) that emitted a range of ∼102-104photons per square micrometer per second at 620 nm was used (Nygaard and Frumkes, 1982). A 1–2 sec light stimulus was presented every 30 sec. In other experiments, bipolar cells were directly stimulated in one of three ways. Puffs (50 msec) of 100 mmK+ in the outer plexiform layer (OPL) or puffs (1 sec) of hyperosmotic (0.5 m) sucrose in the IPL (Yu and Miller, 1995) were delivered through a puff pipette positioned ∼20 μm above the slice preparation. Alternatively, short pulses (1 msec) of current were delivered through an electrode filled with Ringer's solution that was placed directly into the OPL, above the patched ganglion cell. The pulses were generated with a constant-current stimulator (Grass S48 with stimulus isolation unit PSIU6; Grass Instruments, West Warwick, RI).
Analysis. Spontaneous activity in ganglion cells was analyzed using a mini analysis program (Synaptosoft, Leonia, NJ). All other data were analyzed using Clampfit 8 (pClamp8; Axon Instruments).
RESULTS
Group III mGluR activation suppresses spontaneous synaptic activity in ganglion cells
In the dark, spontaneous activity is observed in ganglion cells. This activity has been observed and characterized in third-order cells of the salamander (Taylor et al., 1995; Gao and Wu, 1999), turtle (O'Donovan, 1999), and mouse (Tian et al., 1998) retina. Here we assessed the ability of various mGluRs to modulate spontaneous EPSCs in ganglion cells voltage-clamped at −70 mV. Group I receptors are selectively activated by DHPG (Tauchi et al., 1984; Shen et al., 1995), group II receptors by DCG-IV (Hayashi et al., 1993), and group III receptors by AP-4 (Tanabe et al., 1993).
Application of group I and II mGluR-specific agonists did not significantly affect spontaneous activity. In contrast, group III mGluR activation suppressed spontaneous currents in ganglion cells. Figure1A depicts the spontaneous currents in a transient On–Off ganglion cell recorded in a dark-adapted slice preparation. The spontaneous activity was generated by excitatory glutamatergic input from bipolar cells because the ganglion cell was voltage-clamped close to the chloride reversal potential (−70 mV). Application of 10 μm DHPG or 10 μm DCG-IV, agents that are known to specifically activate groups I and II mGluRs in third-order cells in this preparation (Shen and Slaughter, 1998), did not significantly affect spontaneous activity (Fig.1B,C). In contrast, activating group III mGluRs, using AP-4, dramatically suppressed the spontaneous currents in a reversible manner (Fig.1D,E). In the presence of AP-4, the interval between two events was greatly protracted (mean ± SD interval, 208 ± 210 msec; note change of scale in abscissa) compared with control (25 ± 17 msec). However, the mean amplitude and amplitude distribution of the remaining EPSCs were not significantly changed (mean amplitude was 13 ± 6 pA in AP-4 compared with 15 ± 7 pA in control Ringer's solution). The decrease in frequency and not amplitude are indicative of a presynaptic site of action. Furthermore, AP-4 application did not affect the current responses elicited by pressure injecting glutamate directly onto the ganglion cell (n = 4; data not shown). Together, these observations suggest that group III mGluRs modulate bipolar to ganglion cell communication and that this regulation occurs presynaptic to the ganglion cell.

Effects of mGluR agonists on spontaneous EPSCs in ganglion cells. The ganglion cell was voltage-clamped at −70 mV ( approximate ECl). The spontaneous EPSCs (left) and a histogram and cumulative distribution of the amplitudes (middle) and of the interevent durations (right) of the spontaneous events are shown in the presence of control Ringer's solution (A), the group I mGluR agonist DHPG (B), the group II receptor agonist DCG-IV (C), the group III receptor agonist AP-4 (D), and washout of drugs (E). The asterisk inD (right) indicates that the cumulative distribution was statistically different from control (*p < 0.005 by the Kolmogorov–Smirnov test).
mGluR-III suppresses output at the axon terminal of Off bipolar cells
An objective was to determine the site of action of AP-4. It is known that AP-4 suppresses the light activity of On bipolar cells and drives these cells close to their dark membrane potential (Shiells et al., 1981; Slaughter and Miller, 1981). AP-4 does not suppress Off bipolar light responses. Although the above results suggest that AP-4 was decreasing the output of bipolar cells, it was not clear (1) whether it was acting on On or Off bipolar cells or both and (2) in the case of On bipolar cells, whether it was suppressing spontaneous activity because of the activation of mGluRs at the dendritic or at the axonal terminals.
Evidence that AP-4 affects the output of Off bipolar cells came from evaluation of dark currents of Off ganglion cells (n = 4). At the onset of light, when Off bipolar cell transmitter release decreases and On bipolar cell release increases, there was a loss of excitatory current and a concomitant cessation of spontaneous activity in these ganglion cells (Fig. 2). At light offset, there was a transient EPSC followed by sustained asynchronous activity. This indicates that Off ganglion cells receive inputs predominantly from Off bipolar cells. AP-4 depressed the Off activity in these ganglion cells. A total of 10 μm AP-4 had little effect at light onset yet decreased the transient EPSC at light offset. Even more striking was the suppression of the asynchronous currents that followed the transient Off EPSC (Fig. 2). Because of the low occurrence of this cell type (<10%), they were not studied more extensively. However, the results demonstrate that AP-4 can modulate the Off pathway. Because AP-4 does not suppress the light responses of Off bipolar cells, this implies that AP-4 is acting at the Off bipolar cell axonal terminals.

Group III mGluRs modulate the Off pathway. The light-evoked EPSC in an Off ganglion cell voltage-clamped at −70 mV (ECl) in control Ringer's solution (A), in the presence of AP-4 (B), and when returned to control Ringer's solution (C). The solid barrepresents a 2 sec red-light stimulus.
mGluR-III suppression of release from bipolar cells is intensity-dependent
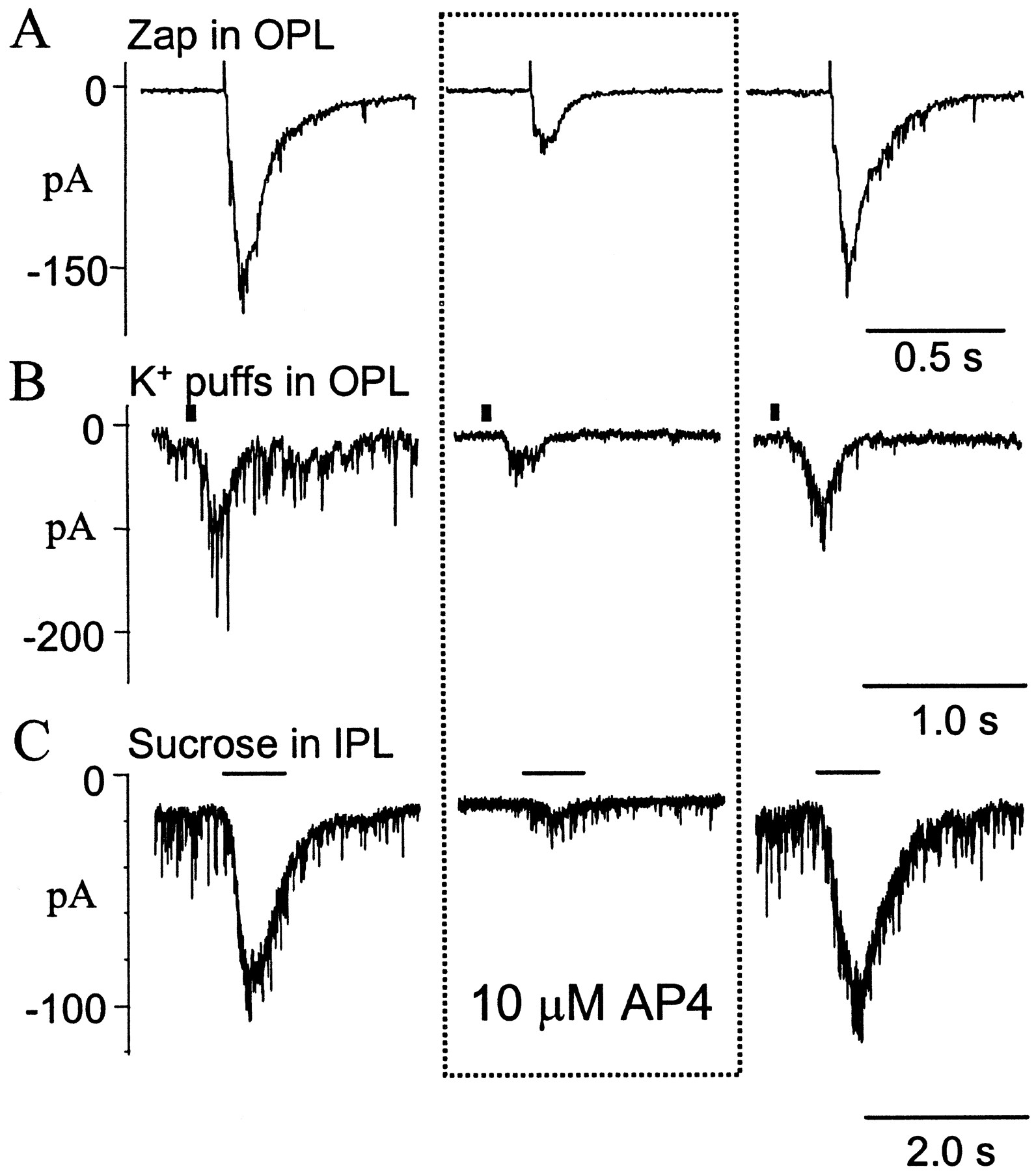
Another method of evaluating regulation of the bipolar cell to ganglion cell synapse is to directly stimulate bipolar cells while recording EPSCs in ganglion cells. This avoids the complications produced by the action of AP-4 on other neurons or the effect of AP-4 at the dendritic terminals of On bipolar cells. We used several methods to stimulate bipolar cells. Glutamate release from bipolar cells was stimulated by puffs of 100 mmK+ in the OPL. Although K+ puffs in the OPL depolarize all cells in the vicinity, the direct effects on bipolar cells probably override synaptic input from photoreceptors or horizontal cells. Potassium puffs in the OPL produced EPSCs in ganglion cells. In three of four ganglion cells tested, mGluR activation caused a suppression of the K+ puff-stimulated EPSCs. In the other ganglion cell, there was no significant effect on the K+-evoked EPSCs. Figure3B shows one such recording in which 10 μm AP-4 reversibly reduced the synaptic current evoked.

Effects of AP-4 on the monosynaptic responses in ganglion cells evoked by artificially stimulating bipolar cells. Responses evoked by electrical pulses (2.5 μA, 1 msec) delivered in the OPL (A), 100 mmK+ puffs (50 msec) in the OPL (B), and 0.5 m sucrose puffs (1 sec) in the IPL (C). Responses are recorded (A–C) in control Ringer's solution (left), in the presence of AP-4 (middle), and after the mGluR agonist was washed out (right). The stimulus artifact preceding the EPSC indicates when the zap was delivered (A). The solid barabove the traces (B,C) represents duration of the stimulus. Neurons were voltage-clamped at −70 mV (ECl).
Bipolar cells were also stimulated by puffing hyperosmotic sucrose (0.5m) solution on the inner plexiform layer. The advantage of this method is that photoreceptors are not stimulated and the responses only reflect the properties of bipolar cell axonal terminals (Yu and Miller, 1995). Application of AP-4 reversibly inhibited the sucrose-evoked EPSCs in four of six cells tested (Fig. 3C). In the other two cells tested, AP-4 slightly enhanced the sucrose-evoked responses
To stimulate the bipolar cells in a more controlled manner, an electrical method of stimulation (zap) was used (Higgs and Lukasiewicz, 1999). This method required injecting brief current pulses (1 msec) in the OPL through an electrode filled with Ringer's solution. Using this stimulation protocol, glutamate release from bipolar cell terminals was controlled by varying the amplitude of the stimulus current. Injecting currents incrementally from 0.5 to 5 μA caused a corresponding increase in EPSCs observed in ganglion cells. The increased EPSC amplitude probably arose from a greater depolarization of bipolar cells and from a larger number of bipolar cells being stimulated. Consistent with the data from the K+ and sucrose protocols, AP-4 suppressed the zap-evoked (2.5 μA, 1 msec) currents (Fig. 3A).
In some ganglion cells, AP-4 did not affect the zap-evoked currents. In these cases, the stimulating current was reduced in amplitude. Weaker stimuli produced smaller currents in ganglion cells that were suppressed by AP-4. For example, Figure4A shows a zap-evoked synaptic potential in a ganglion cell that was not suppressed by AP-4. This ganglion cell response was produced by a 5 μA zap in the outer plexiform layer. When the zap was reduced to 1 μA, the smaller EPSC was suppressed by AP-4 (Fig. 4B). In general, AP-4 suppression was inversely related to the strength of the zap stimulus. On average, AP-4 reduced the EPSC peak amplitude evoked by weak stimulation (0.5–1μA, 1 msec) by 81 ± 10% (n = 6; p < 0.005); however, the EPSC peak amplitude evoked by moderate to strong stimuli (3–5 μA, 1 msec) was only reduced by 29 ± 8% (n = 12;p < 0.05). Hence, exogenous activation of mGluRs was much more effective in suppressing weak synaptic inputs to ganglion cells.

Differential effect of mGluR activation on responses evoked by strong and weak stimuli. A, A 5.0 μA current pulse (1 msec) was delivered to the OPL, and the current responses were recorded in a voltage-clamped ganglion cell.B, Weaker current injections of 1.0 μA elicited smaller responses in the ganglion cell. EPSCs in this cell (forA, B) are shown in control Ringer's solution (left), in the presence of AP-4 (middle), and after the agonist has been washed out (right). Neuron was voltage-clamped at −70 mV (ECl).
Effects of AP-4 in the isolated inner retina
To demonstrate that AP-4 directly acted at the axonal terminals of bipolar cells, we evaluated its actions in a reduced, inner-retinal preparation. After slices were secured in the recording chamber (Fig.5A, left), the outer retina was carefully removed (see Materials and Methods). After the inner retina had been isolated (Fig. 5A,right), the effect of AP-4 on spontaneous and evoked EPSCs in ganglion cells was tested. Spontaneous EPSCs recorded in ganglion cells could be completely blocked by 10 μm2,3-dihydroxy-6-nitro-7-sulfamoylbenzo[f]quinoxaline, a glutamate receptor antagonist (data not shown), suggesting that they originated from bipolar cell axonal terminals. Figure 5B is an example of the spontaneous currents observed in a ganglion cell voltage-clamped at −70 mV. As in the whole-slice preparation, application of 2 μm AP-4 decreased the frequency of these spontaneous currents (Fig. 5B). Similar observations were made in five other ganglion cells.

mGluRs inhibit synaptic transmission in the reduced, inner-retinal preparation. A, Photomicrographs of the retinal slice before (i) and after (ii) the outer retina was removed (see Materials and Methods). B, Voltage-clamp recording of the spontaneous activity from a ganglion cell in a reduced slice preparation in control Ringer's solution (i) and in the presence of 2 μm AP-4 (ii). C, Sucrose-evoked (0.5 m, 1 sec) EPSCs recorded from another ganglion cell in a reduced preparation in control Ringer's solution (i), in the presence of 2 μm AP-4 (ii), and after the drug was washed out (iii). The dark barsabovethe traces show when the sucrose was applied. Neurons were voltage-clamped at −70 mV (ECl).
Next, we examined the effects of AP-4 on evoked synaptic release from bipolar terminals in the reduced, inner-retinal preparation. When hyperosmotic sucrose was puffed on the IPL, robust EPSCs were observed in ganglion cells, as illustrated in Figure 5C. Application of 2 μm AP-4 reversibly reduced the sucrose-evoked EPSCs by 61 ± 15% (n = 4) (Fig.5C). Together, these data confirmed previous results that implied that mGluR activation suppressed synaptic release from bipolar cell axonal terminals.
Endogenous activation of mGluRs
We subsequently examined the role of endogenous mGluR activation in modulating synaptic transmission using an mGluR antagonist, CPPG (Jane et al., 1996; Awatramani and Slaughter, 2000). CPPG antagonized and reversed the effects of exogenously applied AP-4. For example, Figure 6A shows a recording from a ganglion cell in which the application of 2 μm AP-4 diminished the zap-evoked response. A total of 200 μm CPPG blocked the suppression produced by AP-4, enhancing the EPSC relative to control. In a set of eight cells tested, AP-4 reduced the mean EPSC produced by a zap. In six of these eight ganglion cells, 200 μm CPPG not only blocked this action of 2 μm AP-4 but also produced an enhancement of the EPSC (118 ± 22% of control;p < 0.05) (Fig. 6B). In the two other ganglion cells, both AP-4 and CPPG produced a suppression of the zap-evoked EPSC. The finding in six cells that CPPG enhanced the zap-evoked responses in ganglion cells suggests that glutamate released from bipolar cell terminals activates metabotropic autoreceptors, thereby suppressing transmitter release from the axonal terminals.

Endogenous activation of mGluRs suppresses bipolar cell output. A, Zap-evoked (5 μA) EPSCs in ganglion cells voltage-clamped at −70 mV in control Ringer's solution (i), in the presence of 2 μm AP-4 (ii), in the presence of 2 μm AP-4 and 200 μm CPPG (iii), and after the drugs were washed out (iv). B, The mean ± SD peak EPSC amplitude and the total EPSC charge caused by the application of AP-4 are represented by gray columns. The CPPG-mediated enhancements of the peak and the total charge are indicated by white columns (*p < 0.05).
Modulation of the Off response in ganglion cells by group III mGluRs
Because the activation of mGluRs was found to influence bipolar cell output, we subsequently examined the ramifications on the light-driven synaptic responses of ganglion cells. When examining the light-evoked EPSC in ganglion cells, the action of bipolar cells cannot be dissociated from the rest of the retinal circuitry. The effects of applying mGluR agonists and antagonists include their actions in both the inner and outer retina. In the outer retina, applying AP-4 completely eliminates the On bipolar cell response to light but spares the responses of Off bipolar cells. As a result, the study of mGluR regulation of light responses is limited to the Off system. Because mGluR activation differentially suppressed EPSCs to weak and strong zaps, we compared the effects of AP-4 on Off responses evoked by dim- and bright-light stimuli. Figure7A shows a voltage-clamp recording of light-evoked EPSCs in an On–Off ganglion cell held near the chloride reversal potential (−70 mV). The On and Off EPSCs were elicited by 1 sec light stimuli of increasing intensity. When AP-4 was applied, the On response was blocked at all light intensities. This is because of the suppression of the On bipolar light response produced by activation of dendritic mGluRs. The Off responses to the dimmer stimuli were suppressed. As the stimulus was made brighter, the Off response became less suppressed by AP-4 application. With a still brighter light stimulus, the Off response in the presence of AP-4 was slightly enhanced compared with control. The AP-4 suppression of weak Off responses is similar to its effect on zap-induced EPSCs in ganglion cells. However, the enhancement of Off responses to bright-light stimuli does not have a correlate in the zap experiments (see Discussion).

Intensity-dependent modulation of Off responses in ganglion cells. A, Light responses evoked by stimuli of increasing intensity in a ganglion cell voltage-clamped at −70 mV in control Ringer's solution (top) and in the presence of AP-4 (bottom). B, Responses to the same stimulus paradigm recorded in current-clamp mode (zero current), in AP-4 (top), and after the drug was washed out (bottom). C, The mean ± SE of the peak responses from eight cells measured by voltage clamp is plotted as a function of light intensity. The filled squares represent responses measured in control Ringer's solution; the open circles represent responses measured in AP-4. D, Mean AP-4-mediated inhibition as a function of light intensity. The asterisks in Cand D represent mean responses that are statistically different from control (p < 0.01).
In current-clamp mode, effects of AP-4 on the Off responses were similar, but the inhibition was less pronounced. Figure 7Bshows voltage responses in the same cell. Once again, application of AP-4 completely blocked the On responses at all light intensities and suppressed the Off EPSPs to dimmer light stimuli. However, in dim light, the AP-4 suppression of the Off EPSP was less evident than the suppression of the Off EPSC. Voltage-gated conductances play an important role in shaping ganglion cell responses (Diamond and Copenhagen, 1995) and may mask changes in inputs. This might also explain why previous intracellular recordings revealed virtually no effect of AP-4 on Off responses in ganglion cells. Thus, the EPSC seems to be a better gauge of the actions of AP-4.
The effects of AP-4 on Off EPSCs from 10 cells are summarized in Figure7C. The peak Off EPSC amplitudes are plotted as a function of light intensity in control conditions (Fig. 7C,filled squares) and in the presence of AP-4 (Fig.7C, open circles). With the dimmest light stimulus, AP-4 suppressed the responses by an average of 86 ± 10%. Response amplitudes were much less suppressed when the stimulus was 1 log unit brighter. In six ganglion cells, AP-4 enhanced the Off EPSCs by 25 ± 14%; in the other four cells, the EPSCs were suppressed by 35 ± 13% compared with control. Figure7D shows the inverse relationship between light intensity and the suppression caused by AP-4.
To test the possibility that mGluRs acted by regulating inhibitory input to bipolar cells, we examined the effects of AP-4 on ganglion cell responses in the presence of ionotropic GABA and glycine receptor antagonists (100 μm picrotoxin and 10 μmstrychnine). Figure 8 shows the effects of AP-4, in the presence of picrotoxin and strychnine, on ganglion cell On and Off EPSCs evoked by a series of light stimuli of increasing intensity. As expected, AP-4 suppressed all On responses because of its action at On bipolar dendrites. AP-4 suppressed the Off EPSCs evoked by dim-light stimulation. In response to moderate light stimuli, Off responses in the presence of AP-4 were similar to control Off responses. As the stimulus was made brighter, Off responses were enhanced in the presence of AP-4. Similar effects were observed in eight other ganglion cells. These experiments demonstrate that the effects of mGluRs on ganglion cell Off responses are independent of this inhibitory circuitry.

The actions of AP-4 do not depend on the inhibitory circuitry of the retina. All responses were recorded in the presence of 100 μm picrotoxin and 10 μmstrychnine (P + S). A, A series of EPSCs in a ganglion cell evoked by increasing intensities of light stimuli. Responses to the same stimuli were also recorded in the added presence of AP-4 (B) and after AP-4 had been washed out (C). Neurons were voltage-clamped at −70 mV (ECl).
To examine the role of endogenous activation of mGluRs on synaptic inputs, we measured light responses of ganglion cells in the presence of the antagonist CPPG. In these experiments, a more extensive cocktail of inhibitory antagonists was used. A combination of picrotoxin and imidazole-4-acetic acid blocked GABAA and GABAC receptors, CGP35348 blocked GABAB receptors, and strychnine blocked glycine receptors. Under these conditions, AP-4 still strongly suppressed the Off EPSCs evoked by dim-light stimuli (Fig.9A,B,first row). In all six cells tested, the addition of CPPG blocked the effects of AP-4 on the Off EPSCs. In the presence of CPPG, responses to dim lights were not significantly affected in these cells (96 ± 12% of control) (Fig.9A,B, first row). Thus, endogenous feedback was not significantly activated by weak stimuli. In two of six cells, AP-4 decreased the Off response to bright-light stimuli and CPPG reversed and enhanced the EPSC (22 ± 6% enhancement compared with control) (Fig. 9A, second row). This was similar to what was found in the zap experiments (Fig. 6A). However, in four of the six cells, AP-4 either enhanced or did not affect the Off response to bright light (Fig. 9B, second row). In these cells, CPPG reduced the Off EPSCs by 36 ± 9% (Fig. 9B,second row). Thus, CPPG always enhanced ganglion cell EPSCs because of strong zap stimulation of bipolar cells, but CPPG often suppressed Off EPSCs evoked by strong light stimuli. This suggests that bright-light stimuli recruit another circuit besides the bipolar to ganglion cell connection stimulated by the zap. Nonetheless, CPPG experiments using both zap and light stimuli reveal that endogenous activation of mGluRs was only apparent with stronger stimulations.

Bright-light stimuli activate endogenous mGluRs, but weak stimuli do not. For the two types of responses seen in ganglion cells (A, B), responses to dim light are shown in the top row (gray bars) of A and B, whereas responses to bright light (black bars) are shown in thebottom row of A and B. Responses in control (left), in 2 μm AP-4 (middle), and in 2 μm AP-4 plus 200 μm CPPG (right) are illustrated inA and B. The solid gray bars and black bars represent 1 sec (A) and 2 sec (B) of red-light stimuli. Experiments were performed in the presence of 100 μm picrotoxin, 10 μm strychnine, and 500 μm CGP35348. Neurons were voltage-clamped at −70 mV (ECl).
DISCUSSION
Autoreceptor feedback
These experiments demonstrate that group III mGluRs shape bipolar cell outputs to ganglion cells. The effects of agonists and antagonists on monosynaptically evoked EPSCs recorded in ganglion cells suggest that the endogenous activation of mGluRs suppresses bipolar cell output. It is likely that these receptors function as autoreceptors and regulate bipolar cell transmitter release.
Stimuli that produced small postsynaptic responses were strongly suppressed by AP-4 and largely unaffected by CPPG. Conversely, zap stimuli that produced large EPSCs in ganglion cells were relatively unaffected by AP-4 but enhanced by CPPG. The fact that CPPG had little effect during weak stimulation suggests that few autoreceptors were activated under these conditions. Because AP-4 had little effect during strong stimulation, mGluRs were probably saturated under these conditions. Thus, the dynamic range of the postsynaptic response is well matched to the activation of the presynaptic autoreceptors. These experiments were performed on transiently responding ganglion cells; the autoreceptor feedback may differ in sustained ganglion cells.
The correlated activation of presynaptic and postsynaptic receptors argues for a localized action at the synapse. This scenario is distinct from the proposed heterosynaptic regulation mediated by mGluR2 (Scanziani et al., 1997) at mossy fiber synapses in the hippocampus. At these synapses, mGluR2s are located far from the release site (Yokoi et al., 1996) and are stimulated by activity-dependent glutamate spillover (Scanziani et al., 1997). In contrast, group III receptors are often found near synaptic release sites. The feedback receptors at the bipolar cell synapse seem to fit this group III model.
Properties of the autoreceptor
Several group III receptors have been identified in the retina. mGluR6 is localized exclusively to dendritic terminals of On bipolar cells (Nomura et al., 1994; Vardi and Morigiwa, 1997). The mGluR4 receptor has not been observed in bipolar cells of the retina (Hartveit et al., 1995; Koulen et al., 1996). The mGluR7 receptor has been localized to the bipolar cell terminal in rat retina, in which it is found at only one side of the presynaptic dyad (Brandstätter et al., 1996). These receptors have also been found at or near the bipolar cell terminals in salamander retina (Higgs and Lukasiewicz, 2000). However, mGluR7 has a low affinity (EC50 is 1 mm for glutamate and 160 μm for AP-4) (Anwyl, 1999) and is thus not likely to account for the effects we have observed. mGluR8 has been localized to the inner plexiform layer and does have high affinity for AP-4. Therefore, this receptor is a potential candidate for the bipolar cell autoreceptor characterized in this study. This leaves the possibility that another, perhaps more selective, autoreceptor function is mediated by mGluR7.
Application of CPPG increased the peak of an EPSC evoked by a single test pulse. Because the EPSC peaked in <20 msec, the endogenous autoreceptor feedback system must act within this time scale. Thus, it appears that postsynaptic and presynaptic responses are well correlated in time as well as in dynamic range. Rapid regulation by mGluRs on this time scale has been observed in neurons in the hypothalamus (Schrader and Tasker, 1997) and in the lamprey brainstem (Cochilla and Alford, 1998). It suggests a truncated transduction cascade, possibly a direct G-protein action.
Metabotropic glutamate receptors regulate Off bipolar cell output
In the dark, a continuous train of spontaneous excitatory synaptic currents is observed in ganglion cells (Tian et al., 1998; Gao and Wu, 1999). These spontaneous EPSCs, generated by glutamatergic input from bipolar cells, are more likely to originate from Off bipolar cells because these neurons are depolarized and exhibit prominent spontaneous activity in darkness, whereas On bipolar cells are hyperpolarized and are relatively quiet (Wu et al., 2000). This spontaneous activity observed in ganglion cells is dramatically reduced by AP-4, indicating that mGluRs regulate output of Off bipolar cells. Agonists of group I or group II mGluRs do not appreciably influence spontaneous EPSCs, indicating that this regulation is mediated by a group III mGluR. Consistent with this finding, AP-4 was also found to reduce the dark currents in Off ganglion cells (cells that received their inputs primarily from Off bipolar cells). Because AP-4 does not suppress the light-evoked response of Off bipolar cells, it is most likely acting at the bipolar cell axon terminal.
Metabotropic glutamate receptors act at On bipolar cell axonal terminals
To demonstrate that AP-4 regulates the output of On bipolar cells, exogenous stimulation was used. Using artificial stimulation, it was difficult to ensure that only On bipolar cells were stimulated. However, it is likely that these protocols preferentially stimulated transmitter release from On bipolar cells. In the dark, Off bipolar cells are depolarized and release transmitter. On bipolar cells are hyperpolarized and release very little transmitter. The zaps and K+ puffs used depolarize both types of bipolar cells, but the incremental transmitter release should be greater for the On bipolar cell. Consistent with this proposition, exogenous stimulation of bipolar cells just before a light stimulus depressed On more than Off responses (Yu and Miller, 1995). Direct depolarization of bipolar cells, using puffs of high potassium in the outer plexiform layer, initiated EPSCs in ganglion cells. These EPSCs were suppressed by AP-4, indicating that AP-4 acted directly on the bipolar cell to reduce its output to ganglion cells. Similarly, sucrose puffs in the inner plexiform layer produced EPSCs in ganglion cells, and these responses could also be suppressed by AP-4. Together, these data suggest that AP-4 is acting at the On bipolar cell synaptic terminals.
Other mGluR pathways may influence the light responses
Because AP-4 suppressed spontaneous activity, it was anticipated that it would also reduce the light-evoked EPSCs in ganglion cells. AP-4 blocks On light responses because of an action at On bipolar dendrites, so only the Off bipolar input to ganglion cells could be analyzed. The Off EPSC was consistently suppressed when weak to moderate (nonsaturating) light stimuli were used. The effects of AP-4 on responses to nonsaturating light stimuli were similar to our other experimental protocols (potassium, sucrose, current stimuli, and spontaneous activity) and could be interpreted as a simple suppression at the Off bipolar cell axon terminal. However, an additional effect of AP-4 at the photoreceptors (Koulen et al., 1999) cannot be ruled out. The effects of AP-4 on responses to bright light were more variable. In some neurons, the EPSC was decreased by AP-4. In these cells, CPPG enhanced the responses to strong light stimulation. However, in many neurons, AP-4 did not alter or even enhanced the Off responses, as reported previously (Arkin and Miller, 1987). In these cases, CPPG suppressed the Off EPSC. In every case, the action of CPPG was opposite to that of AP-4, indicating that these varied effects of AP-4 represent a specific action on mGluRs. Because AP-4 consistently suppressed the ganglion cell EPSCs produced by direct bipolar cell stimulation, spontaneous EPSCs, and dim light, it is likely that the AP-4 enhancement during bright-light stimuli activates another part of the retinal network.
A likely candidate is the rod input to the Off bipolar cell. Hare and Owen (1992) found that AP-4 enhanced rod input to Off bipolar cells. A red LED stimulus was used in our experiments. This preferentially stimulates cones, such that dim-light intensities had a very small effect on rods. Strong red LED stimulation was required to vigorously stimulate rods. Therefore, the AP-4 enhancement of Off responses is likely the result of an augmented rod component that is only evident during strong light stimulation.
Physiological implications
Functionally, the autoreceptor feedback may extend the dynamic range of the bipolar cell output synapse. Weak stimuli evoke little feedback (based on the small CPPG effect). Strong signals that might saturate the bipolar cell output do not do so because of the negative feedback (based on enhancement of strong zaps by CPPG). This permits an increase in output range. An interesting extension of this concept is that the mGluR may allow for potentiation of bipolar cell output. If the mGluR is suppressed, then the output of the bipolar cell will be enhanced, particularly to moderate and strong light stimuli. The bipolar cell output can be potentiated by protein kinase C (Minami et al., 1998). Protein kinase C can suppress group III mGluRs (Macek et al., 1999). This suggests a possible mechanism in which protein kinase C inhibits the mGluR, thereby potentiating the output of bipolar cells. Thus, the gain of the bipolar to ganglion cell synapse might be controlled by factors in the inner retina that regulate autoreceptor feedback.
Footnotes
This work was supported by National Eye Institute Grant EY05725. We thank Dr. Asbed Keleshian for his critical comments on this manuscript.
Correspondence should be addressed to Malcolm M. Slaughter, Department of Physiology and Biophysics, 124 Sherman Hall, 3435 Main Street, Buffalo, NY 14214. E-mail: mslaught{at}buffalo.edu.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}