

Abstract
Identification of long tracts responsible for the initiation of spontaneous locomotion is critical for spinal cord injury (SCI) repair strategies. Pathways derived from the mesencephalic locomotor region and pontomedullary medial reticular formation responsible for fictive locomotion in decerebrate preparations project to the thoracolumbar levels of the spinal cord via reticulospinal axons in the ventrolateral funiculus (VLF). However, white matter regions critical for spontaneous over-ground locomotion remain unclear because cats, monkeys, and humans display varying degrees of locomotor recovery after ventral SCIs. We studied the contributions of myelinated tracts in the VLF and ventral columns (VC) to spontaneous over-ground locomotion in the adult rat using demyelinating lesions. Animals received ethidium bromide plus photon irradiation producing discrete demyelinating lesions sufficient to stop axonal conduction in the VLF, VC, VLF–VC, or complete ventral white matter (CV). Behavior [open-field Basso, Beattie, and Bresnahan (BBB) scores and grid walking] and transcranial magnetic motor-evoked potentials (tcMMEP) were studied at 1, 2, and 4 weeks after lesion. VLF lesions resulted in complete loss or severe attenuation of tcMMEPs, with mean BBB scores of 18.0, and no grid walking deficits. VC lesions produced behavior similar to VLF-lesioned animals but did not significantly affect tcMMEPs. VC–VLF and CV lesions resulted in complete loss of tcMMEP signals with mean BBB scores of 12.7 and 6.5, respectively. Our data support a diffuse arrangement of axons within the ventral white matter that may comprise a system of multiple descending pathways subserving spontaneous over-ground locomotion in the intact animal.
In frequently studied rat models of thoracic spinal cord injury (SCI), disruption of ascending and descending white matter tracts is the primary cause for the observed locomotor deficits (Magnuson et al., 1999). Dorsal hemisections, even those including one ventral quadrant, do not induce significant deficits unless they extend ventral to the central canal (Imai and Aoki, 1996; Schwab and Brosamle, 1997). Moreover, thoracic contusion injuries that damage ventral white matter produce severe deficits in hindlimb locomotion (Basso et al., 1996). Together, these data suggest that the initiation of hindlimb locomotor activity is not compromised in the rat unless there is bilateral damage to white matter pathways ventral to the central canal. Current hypotheses concerning the origin of descending locomotor command pathways focus on the mesencephalic locomotor region (MLR), an area in the caudal cuneiform nucleus of mammals that produces locomotor movements when stimulated electrically (Shik et al., 1966, 1967). MLR projections synapse in the pontomedullary medial reticular formation (MedRF) before descending as the reticulospinal tract in the ventrolateral funiculus (VLF) of the spinal cord in which they eventually synapse on central pattern generator (CPG) neurons at the lumbar levels (Steeves and Jordan, 1980, 1984; Garcia-Rill et al., 1981; Jordan, 1986, 1991; Noga et al., 1991). Fictive locomotion can also be initiated from the pontomedullary locomotor region (PLR) in the lateral tegmentum of the brainstem via projections to the MedRF, as well as direct projections to the CPG along the dorsolateral funiculus (DLF) white matter tracts (Mori et al., 1977, 1978; Shik and Iagodnitsyn, 1977, 1978; Budakova and Shik, 1980; Kazennikov et al., 1980; Selionov and Shik, 1981). However, lesioning of DLF white matter tracts alone does not prevent MLR- or PLR-evoked fictive locomotion (Sholomenko and Steeves, 1987;Noga et al., 1991).
The pathways identified in studies of MLR-induced fictive locomotion suggest that locomotor dysfunction after thoracic SCI may result from damage to reticulospinal fibers projecting from the MedRF to the thoracolumbar spinal cord. The arrangement of thoracic fibers that subserve locomotion has been difficult to ascertain because conventional surgical approaches cannot reliably produce lesions confined to discrete regions of the thoracic ventral spinal cord. These fibers have long been assumed to lie within a relatively discrete region of the VLF based on ventral quadrant cooling experiments (Noga et al., 1991), surgical lesions in the cervical spinal cord (Steeves and Jordan, 1980), and neonatal in vitro studies (Magnuson and Trinder, 1997). However, a growing body of evidence suggests that these specific white matter regions may not represent the full gamut of descending inputs capable of initiating spontaneous locomotion (Sholomenko and Steeves, 1987; Vilensky et al., 1992; Nathan, 1994;Brustein and Rossignol, 1998). The present study was designed to address the relative contributions of myelinated fiber tracts located in the adult rat thoracic VLF and ventral columns (VC) to locomotion and to provide anatomical, behavioral, and in vivoelectrophysiological correlates for the evaluation of evolving SCI therapies.
MATERIALS AND METHODS
All surgical interventions and both presurgical and postsurgical care were provided in strict accordance with the Public Health Service Policy on Humane Care and Use of Laboratory Animals,Guide for the Care and Use of Laboratory Animals (Institute of Laboratory Animal Resources, National Research Council, 1996), and with the approval of the University of Louisville Institutional Animal Care and Use Committee.
Radiation. One adult female Fischer 344 rat (175 gm) (Harlan Sprague Dawley, Indianapolis IN) was imaged in a Philips Tomoscan 6000 CT scanner to obtain three-dimensional anatomical information for radiation planning. Slices (1.5 mm) were obtained using the maximum resolution afforded by the scanner. Radiation planning was undertaken using ADAC Laboratories (Milpitas, CA) Pinnacle treatment planning software running on a Sun Ultra 2 workstation. Photon irradiation was chosen because it optimized the radiation dose along the spinal cord while sparing skin and internal organs.
For the spinal cord lesions, animals were anesthetized with sodium pentobarbital (50 mg/kg, i.p.) 48 hr after ethidium bromide (EB) injections, as described below, and secured in the prone position. Animals were then shielded with custom cerrobend collimators that localized the radiation dose to a 3 cm length of cord restricted to the dorsal surface. 40Gy (Blakemore and Patterson, 1978) were delivered to the spinal cord using two Elekta SL-25 linear accelerators delivering 6 MV lateral opposed photon fields at a 100 cm source to axis distance. The CT scanners and linear accelerators are housed at the James Graham Brown Cancer Center (University of Louisville). After irradiation, animals were given gentamycin (0.03 mg/kg, i.m.) and a 10 cc subcutaneous bolus of lactated Ringer's solution, and their cages were placed on a 37°C heating pad overnight.
Surgery. Thirty-two adult female Fischer 344 rats (158–179 gm) were anesthetized with sodium pentobarbital (50 mg/kg, i.p.) and subsequently given prophylactic intramuscular injections of gentamycin (0.03 mg/kg, i.m.). After dorsal laminectomies at the T9 and T10 vertebrae, the dura was opened transversely at three longitudinal sites 2 mm apart. Bilateral injections (0.5 μl per injection) of 0.3 mg/ml EB were placed at stereotaxic coordinates within the VC (0.3 mm lateral to midline at depths of 2.0 mm and then 1.7 mm), VLF (0.7 mm lateral to midline at depths of 1.5 mm and then 1.2 mm), combined VC–VLF, and CV (combined VC–VLF with an additional injection 0.7 mm lateral to midline at 1.1 mm) using custom-pulled glass micropipettes (20 μm internal diameter) attached to a Parker picospritzer (Magnuson et al., 1999; Cao et al., 2001). Each injection was delivered over 10 sec and allowed to diffuse for 3 min before micropipette movement. The surgical site was irrigated copiously with warmed normal saline and closed in layers using Vicryl 5–0 sutures. One control group (n = 4) received injections of normal saline at coordinates identical to the CV lesion.
Spinal cord processing. Control and injured rats were similarly processed for histological examination at 2 and 4 weeks after injury. The histological results were similar at 2 and 4 weeks, and only the 4 week data will be presented. All animals were deeply anesthetized with sodium pentobarbital (60 mg/kg, i.p.) and perfused transcardially with 100 ml of cold 0.1 mPBS, followed by 400 ml of 4% paraformaldehyde in 0.1m phosphate buffer, pH 7.4. Spinal cords were post-fixed at 4°C overnight and then transferred to 30% sucrose at 4°C for cryoprotection (48 hr). Afterward, spinal cords were mounted in TBS compound (Triangle Biomedical Sciences, Durham, NC) and sectioned at 16 μm with a cryostat. Myelin staining was performed with standard luxol fast blue methods. Immunostaining of axons was performed as described previously (Cao et al., 2001) using 160 kDa neurofilament (NF-M) (1:200; Chemicon, Temecula, CA). Images were captured using a Zeiss (Oberkochen, Germany) Axioskop 50 upright microscope equipped with an Optronix three-chip CCD camera connected to a Power Macintosh 9600 computer (Apple Computers, Cupertino, CA) running NIH Image software.
Functional outcome measures. As an in vivoelectrophysiological measure of motor pathway function, transcranial magnetic motor evoked potentials (tcMMEP) were performed at 1, 2, and 4 weeks after lesion, using methods described previously (Magnuson et al., 1999). Briefly, the tcMMEP relies on the activation of subcortical structures with an electromagnetic coil placed over the cranium. Action potentials descend in the ventral spinal cord and synapse onto motoneuron pools in which output signals can be recorded from both of the gastrocnemius muscles. tcMMEP latency measurements have been shown to be very consistent in comparisons made between animals (Linden et al., 1999; Magnuson et al., 1999).
The Basso, Beattie, and Bresnahan (BBB) scale, a 22-point scale (0–21) that is based on the recovery of hindlimb function after contusive thoracic spinal cord injuries in rats (Basso et al., 1995), was used for the evaluation of spontaneous open-field locomotion in all lesioned animals. During the evaluation, animals were allowed to walk freely on the open-field surface for 4 min while being observed by two scorers lacking knowledge of the experimental groups.
Grid walking was also used to assess hindlimb locomotor deficits (Behrmann et al., 1992). Animals underwent 5 d of acclimation on the grid-walking apparatus while also being exposed to food incentives. After acclimation, animals were food deprived for 2 d, followed by a diet restricted to 67% of normal intake. Animals were always allowed access to water ad libitum. Animals were trained for 5 d to cross a 150 cm concourse containing 4 cm square holes for food rewards. The grid-walking surface was covered by a 10 × 10 × 140 cm wooden tunnel with a clear Plexiglas top leading to a darkened box at the end that contained food rewards. Body weights were maintained at 90% of pretest values while animals underwent three prelesion testing sessions with 1 d between each session. Total hindlimb footfalls were counted by two observers unaware of the experimental groups during each trial. A footfall was defined as a misplacement such that the hindpaw to ankle fell completely below the level of the grid. Before surgery, animals were allowed to return to their pretraining weights with ad libitum food. Testing was resumed without food restriction once weekly at 1, 2, and 4 weeks after lesion.
Statistical analyses. Repeated-measures ANOVAs were used to analyze BBB scores for all groups and electrophysiologic measures in the VC group. One-way ANOVAs were used to analyze differences in BBB scores and electrophysiologic measures among groups at each time point. Repeated-measures and one-way ANOVAs were followed by Tukey's or Student–Newman–Keuls post hoc t tests as appropriate for sample size in significance analyses. Student'st tests were used (for equal and unequal variances as applicable) to analyze time points of CV control and VLF groups with respect to VC prelesion values for tcMMEP latencies and amplitudes.
RESULTS
To address the relative contributions of thoracic ventral spinal myelinated tracts to locomotion, well characterized EB combined with x-irradiation (EB-X) methods were used to create discrete bilateral demyelinating lesions in the VLF and VC (Honmou et al., 1996;Jeffery et al., 1999). Unlike previously used mechanical methods, these lesions provided high specificity without significant manipulation of other regions of the spinal cord. tcMMEPs were monitored as an in vivo functional measurement because these signals are severely affected by standardized thoracic contusions that produce locomotor deficits. The goal of localizing this signal was to establish a reliable pathway for therapies that could be monitored for efficacy in vivo.
Demyelination
Figure 1B–E shows representative luxol fast blue-stained transverse sections of T10 spinal cord 4 weeks after irradiation and bilateral injections of EB (Blakemore and Patterson, 1978; Honmou et al., 1996) into the VLF (B), VC (C), combined VLF–VC (D), or CV (E). Control animals (n = 4) received injections of 0.9% normal saline into stereotaxic coordinates identical to the CV lesion (Fig.1A). Spinal cords from the control group did not show evidence of demyelination and were structurally intact, aside from one animal that exhibited a slight expansion of the central canal. Histological examination of VLF-lesioned spinal cords (n = 12) revealed well defined areas of demyelination that consistently followed a linear diffusion pattern extending away from each injection point toward the central canal and peripherally to the dura edge (Fig. 1B). The central extent of VLF lesions exhibited varying degrees of ventral gray matter involvement that frequently encompassed laminas VII, VIII, and IX. It should be noted that the gray matter component of thoracic lesions produced in this study should not have contributed to the observed results becauseMagnuson et al. (1999) previously demonstrated no significant effect on locomotor behaviors or tcMMEP recordings after selective thoracic gray matter lesions. Widths of the demyelinated bands ranged from 0.8 to 1.0 mm within the VLF, and sparing of triangular regions of ventral white matter was observed both lateral to the lesion borders and medially within the ventral columns.

Representative luxol fast blue-stained transverse sections of T10 (injury epicenter) spinal cord taken from representative animals in each group 4 weeks after the indicated lesions. A, CV saline control; B, VLF; C, VC; D, VLF–VC; E, CV. Lesions for all animals in the respective groups shared equivalent histopathology. F, Representative parasagittal section taken from the epicenter of a VLF-lesioned animal. Demyelinated lengths of axons (6–8 mm) were sufficient to stop axonal conduction in all lesion types. Arrows indicate the sites of EB injections. Similar histopatholgy was observed in all VLF animals that were similarly sectioned.
Figure 1F shows a parasagittal section taken from a representative VLF spinal cord 4 weeks after lesion induction. It demonstrates the distribution of demyelination reproducibly seen between the three injection sites (arrows). The rostrocaudal extent of demyelination in all lesions ranged from 6 to 8 mm, lengths sufficient to prevent axonal conduction in normally myelinated axons (Honmou et al., 1996), and did not extend rostrally beyond T8 or caudally beyond T11. Animals exhibiting demyelination in the region of the lateral funiculus within the ventral spinal cord normally spared by VLF lesioning did not exhibit significant locomotor deficits (n = 2; data not shown).
VC lesions (n = 7) displayed similarly well demarcated areas of demyelination that extended laterally through the ventral columns from the ventromedial fissure to involve the medial aspects of the ventral gray matter (Fig. 1C). Demyelination of white matter dorsal to the central canal was not observed in VC lesions. Demyelination was restricted to a 0.1–0.2 mm area width immediately adjacent to the lateral gray matter in the largest VLF and combined VLF–VC lesions. VLF–VC lesions (n = 6) spared only the aspects of ventral white matter dorsolateral to VLF lesion borders (Fig. 1D). CV lesions (n = 3) covered an extensive area of ventral white and gray matter because the aim was to create a functional ventral hemisection while leaving the dorsal funiculus and dorsolateral funiculi undisturbed (Fig.1E). The volumes of EB needed to produce CV lesions proved more difficult to control surgically, producing some partial demyelination dorsal to the desired lesion area.
Axonal densities appeared to be reduced within areas of demyelination at 4 weeks as described previously in EB-X adult rat thoracic dorsal column lesions (Jeffery et al., 1999); however, axons that remained were intact because they did not exhibit end-bulb formation (Fig.2). No differentiation between demyelinated and unmyelinated axons was attempted; therefore, damage to unmyelinated axons could not be ruled out. Macrophages were abundant within all lesions at 4 weeks (data not shown), consistent with previous results (Blakemore and Patterson, 1978; Honmou et al., 1996). During the 4 week course of the study, lesions should not have undergone spontaneous remyelination after 40Gy doses of irradiation (Blakemore and Patterson, 1978). Present evidence of the lack of spontaneous remyelination included lesion epicenters devoid of luxol fast blue staining at 4 weeks, persistent absence of tcMMEPs or stable tcMMEP latency measurements in VLF- and VLF–VC-lesioned animals, and the observed lack of recovery of BBB scores in VLF–VC- and CV-lesioned animals (described below).

Axons within the demyelinated VLF lesion epicenter. NF-M immunostaining did not demonstrate end-bulb formation in axons remaining within the lesion epicenter at 4 weeks. Similar data were seen in all VLF animals that were similarly immunostained.
tcMMEP responses
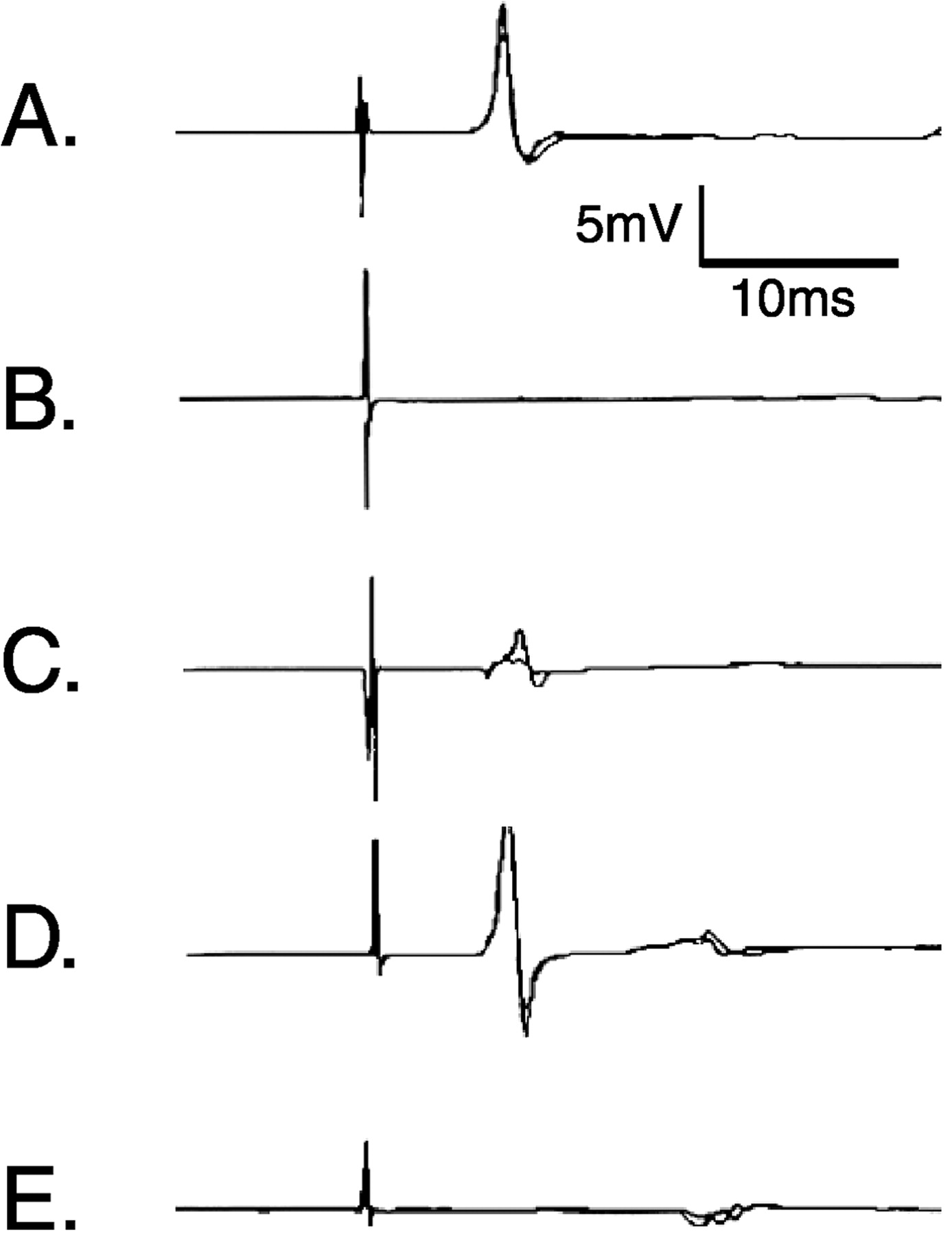
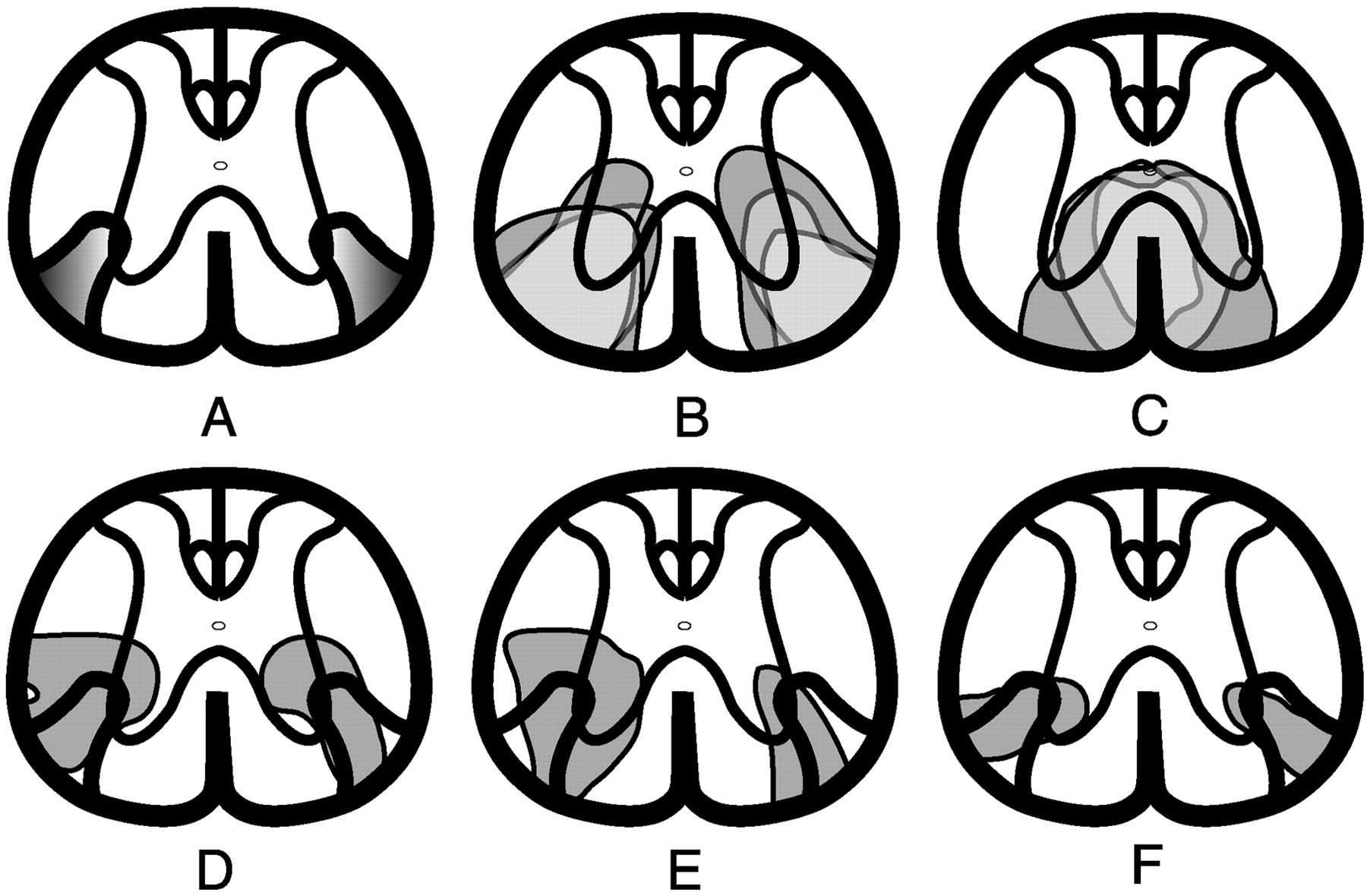
tcMMEP responses were recorded from the left and right gastrocnemius muscles before lesion and 1, 2, and 4 weeks after lesion. Data from control animals are presented in Table1 and Figure3A. Animals receiving VLF lesions either displayed no tcMMEP response (VLFb) or severely impaired responses (VLFa) to transcranial magnetic evoked stimulation at 1 week after lesion (Table 1; Fig. 3B,C). It is worth noting that two of seven VLFb animals that did not have tcMMEP responses at week 1 regained grossly delayed responses by week 2 that remained unchanged to the end of the study despite any histologic evidence of remyelination. In these two animals, small areas of incompletely demyelinated axons may have experienced a transient period of complete dysfunction that recovered by week 2 to a functional level limited by the extent of damage to the myelin sheath. tcMMEP responses with increased latencies and severely reduced amplitudes were seen in VLFa rats (Table 1; Figs. 3C,4D–F). Histologic analyses of these animals consistently demonstrated either slight axial rotational errors in the stereotaxic placement of EB or volumetric errors that left small regions of patchy myelination along the borders of the tcMMEP fiber strip (Fig.4D–F). Figure4A illustrates the hypothetical pathway responsible for tcMMEP conduction in the thoracic ventral white matter. Sections of lesion epicenters from rats with no (Fig. 4B) or severely attenuated tcMMEP responses (Fig.4D–F) were overlaid onto lesions that did not significantly affect tcMMEPs (Fig. 4C) to delineate the minimal bilateral areas involved in tcMMEP conduction.
BBB scores and tcMMEP responses for all groups of rats

Representative examples of tcMMEP responses recorded from gastrocnemius muscles at 4 weeks after bilateral demyelinating lesions. A, CV saline control;B, C, VLF; D, VC;E, VLF–VC or CV. The difference betweenB and C is that C is representative of VLF lesions that severely attenuated tcMMEP responses. The VLF was incompletely lesioned in these animals. tcMMEPs recorded from VC-lesioned animals did not differ significantly from saline controls.

tcMMEP fiber strip located within the VLF.A, Sections from the epicenters of animals that showed diminished tcMMEP responses were overlaid with sections taken from the epicenters of lesions that did not significantly affect tcMMEP responses to delineate the minimal anatomical localization of the pathway that carries the tcMMEP. B, Extent of demyelination in three VLF-lesioned animals that exhibited complete loss of tcMMEPs at 4 weeks after lesion. Shading variations indicate areas of overlap between the three lesions.C, Extent of demyelination in three VC-lesioned animals that did not have significantly affected tcMMEP responses.D–F, Extent of demyelination in three VLF-lesioned animals in which tcMMEPs remained but were severely attenuated. In each rat, portions of the tcMMEP fiber strip (A) were spared.
VC-lesioned animals did not display significant changes in tcMMEP responses compared with saline controls, although latencies were slightly delayed in both groups compared with prelesion values (Table1; Fig. 3D). VLF–VC- and CV-lesioned animals never displayed tcMMEP responses after lesion induction (Table1; Fig.3E), because the tcMMEP fiber strip was always located well within the lesion borders (Fig. 4).
Locomotor deficits
BBB open-field locomotor scores for right and left hindlimbs were assessed before lesion and 1, 2, and 4 weeks after lesion. Paired t test analyses did not demonstrate statistical differences (p = 0.35) between the scores for the limbs of each rat, and thus they were averaged. Table 1shows the mean ± SD BBB scores for all groups of rats. Control animals (Fig. 5E) demonstrated BBB scores throughout the 4 weeks, which were not significantly different from those of naı̈ve rats (BBB score of 21). The mean BBB scores for VC-lesioned animals (18.5 ± 1.0 at 4 weeks) (Fig.5C,D) indicate modest but significant reductions in BBB scores compared with pretest values and controls. VLF-lesioned animals (Fig. 5A,B) scored similarly to VC-lesioned animals despite the absence of (Fig.3B; Table 1, VLFb) or severe deficits in (Fig.3C; Table 1, VLFa) tcMMEP responses (BBB scores of 17.0 ± 0 and 18.0 ± 1.2, respectively). Two-way ANOVAs of both VLF and VC groups showed no significant recovery of hindlimb locomotor function by 4 weeks. VLF–VC-lesioned animals (Fig.5F) exhibited weight-supported locomotion with pronounced forelimb–hindlimb coordination deficits (BBB score of 12.7 ± 0.8 at 4 weeks). CV-lesioned animals (Fig. 5G) displayed severe locomotor deficits (BBB score of 6.5 ± 2.1 at 4 weeks) and were unable to support weight with the hindlimbs throughout the course of the study. The paralysis associated with CV-lesioned animals was flaccid, with passive flexion–extension maneuvers revealing no evidence of spasticity in the large hindlimb muscle groups.

Open-field locomotion 4 weeks after various bilateral lesions within the thoracic ventral spinal cord.A, B, VLF; C,D, VC; E, control; F, VLF–VC; and G, CV. In contrast to control animals (E), note the prominent toe walking in VLF-lesioned animals (A, B).C, D, VC lesions produced BBB scores similar to VLF lesions with less frequent toe walking.F, VLF–VC-lesioned animals displayed severe deficits in coordination but were always capable of weight support.G, CV lesions produced flaccid paralysis of the hindlimbs.
BBB scores were not assigned to animals during the first 6 d after surgery. However, VLF- and VC-lesioned animals did not exhibit loss of weight support during this interval after lesioning when they were removed from their cages and placed on a flat surface during twice daily physical exams. VLF–VC- and CV-lesioned animals exhibited obvious severe locomotor deficits or paraplegia within 48–72 hr after EB injections that did not resolve at any time before the postlesion week 1 BBB scoring session.
It is important to note that the discrete nature of the VLF and VC lesions produced locomotor behavioral changes that were not accurately represented by the BBB scoring system. To evaluate these behavioral changes for both VLF and VC groups, two observers lacking knowledge of the experimental groups recorded spontaneous locomotor behaviors not described in the BBB scoring criteria that are not normally present in naı̈ve animals. Figure 6represents the most frequent hindlimb behaviors. All VLF (VLFa and VLFb)-lesioned animals (n = 12) were observed to always walk on their toes in severe plantar flexion without heel contact on the walking surface (Fig. 5A,B). Toe walking was never seen in control rats. Passive flexion–extension maneuvers of the hindlimbs again did not uncover any evidence of spasticity in the large muscle groups. The base of the tail and ridge of the lumbosacral spine were also noticeably elevated during open-field locomotion in VLF-lesioned animals. Toe walking was also observed in 53% of VC-lesioned animals (n = 7); however, the degree of plantar flexion was not as qualitatively severe as that of the VLF group because their heel frequently made contact with the open-field surface (Fig. 5C,D). Sixty-seven percent of VC-lesioned animals and 43% of VLF-lesioned animals also displayed a phenomenon of heavy stepping during locomotion that can be defined as an audible noise associated with the placement of each hindpaw onto the open-field testing surface. This is in contrast to normal animals that move almost silently, even during brisk locomotion.
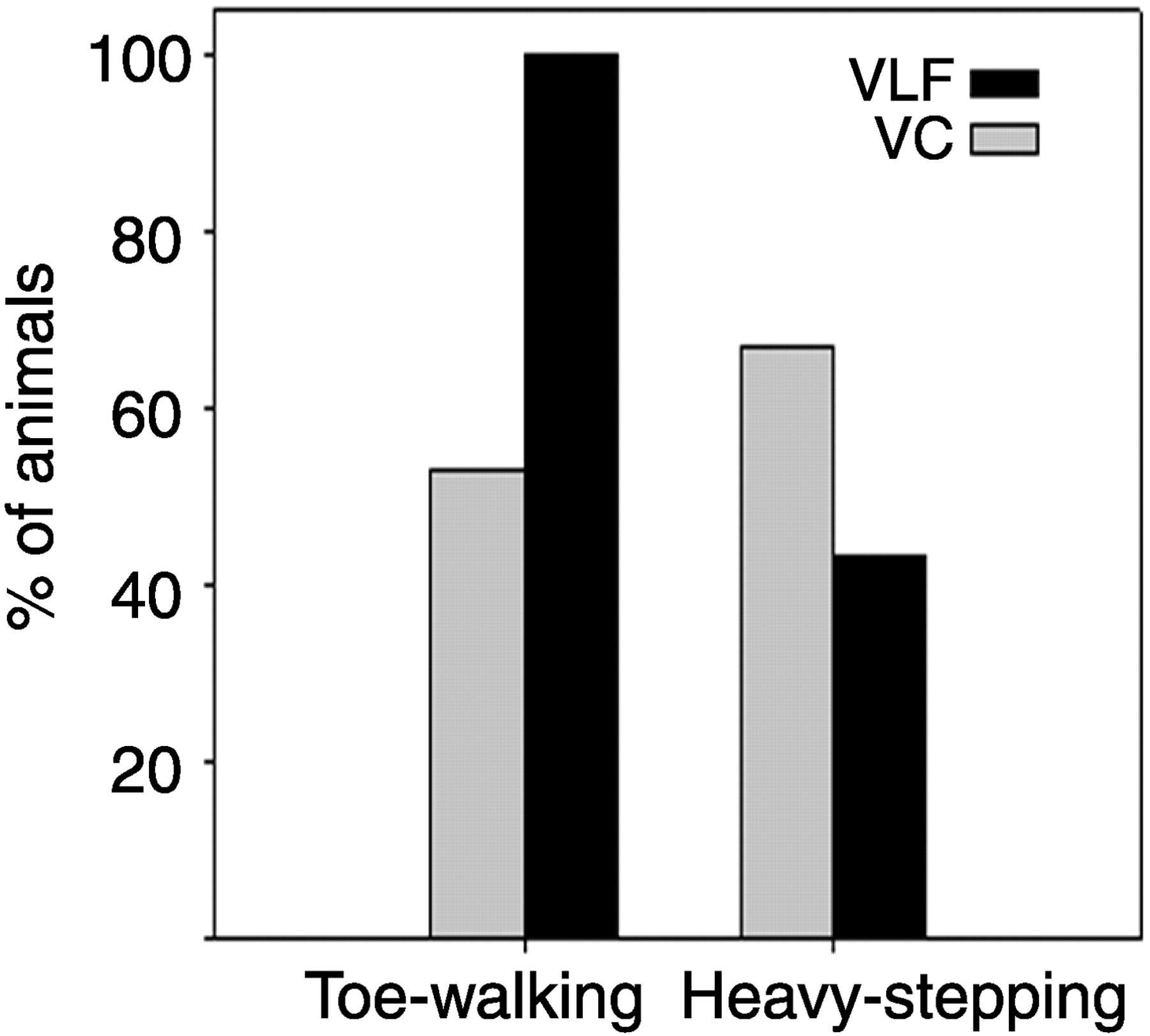
Locomotor deficits exhibited in VC- and VLF-lesioned animals not recognized by the BBB scoring system. All VLF-lesioned animals displayed toe walking during open-field locomotion, and half of VC-lesioned animals exhibited toe walking that was less qualitatively severe. Sixty-seven percent of VC-lesioned animals displayed heavy stepping compared with 43% of VLF-lesioned animals (n = 12 and 7 for VLF and VC, respectively).
Other less frequently observed behaviors included the following: 3 of 12 VLF-lesioned animals that had a tendency to take small shuffle-like steps, one VC-lesioned animal of seven that preferentially moved by stepping laterally, and one VC-lesioned animal that converted to an abnormally wide stance after coming to a halt. Histological analysis of the VC-lesioned animal with a widened stance revealed demyelination in the ventral columns that extended into the lumbar enlargement, and tcMMEP responses were comparable with controls. This animal was excluded from the study. No histological or electrophysiological correlates could be determined in the VLF animals exhibiting small steps or the VC animal that stepped laterally.
Figure 7 shows the number of hindlimb footfalls for pretrained animals walking on a grid both before lesion and after VC or VLF (VLFa and VLFb) lesion induction. Prelesion values were 0.1 ± 0.1 and 0.3 ± 0.1 (footfalls per 150 cm trial) for VC and VLF groups, respectively. For postlesion induction, mean footfalls ranged from 0.5 ± 0.4 at 1 week to 0.3 ± 0.6 at 4 weeks for VC- lesioned animals and 1.0 ± 0.6 to 1.0 ± 1.1 for VLF-lesioned animals. No statistical differences were revealed when comparisons were made over time (repeated-measures ANOVA) or between groups (paired t tests). VLF–VC- and CV-lesioned animals did not exhibit locomotion sufficient to warrant grid-walking evaluation.

Grid-walking behaviors. Data represent the mean ± SD of 12 and 7 animals for the VC and VLF lesions, respectively. Neither VC- nor VLF-lesioned animals displayed significant increases in footfalls while walking on the grid (VC, df = 3,15; F = 1.24; p = 0.33; VLF, df = 3,24; F = 1.008;p = 0.42).
DISCUSSION
Lesions
This study examined the role(s) of ventral spinal cord pathways on open-field locomotion in the rat. Bilateral demyelinating VLF lesions at T9–T11, which completely blocked conduction in axons that transmit tcMMEPs, had little impact on BBB locomotor scores. Bilateral demyelinating VC lesions also produced only mild locomotor deficits. Thus, myelinated axons within either the VLF or VC are sufficient for the initiation of open-field locomotion in the rat. However, when VLF and VC lesions were combined or extended to encompass larger amounts of ventral white matter, severe locomotor deficits could be induced.
Historically, evidence supporting a role for the VLF in locomotion comes from studies demonstrating that MLR-evoked hindlimb fictive locomotion is lost after mechanical VLF lesioning (Steeves and Jordan, 1980) or selective VLF cooling (Noga et al., 1991). However, many have suggested that the MLR–MedRF–VLF pathway does not represent a solitary system for the initiation of locomotion in the intact animal. For example, the VLF is not essential or necessary for partial recovery of hindlimb locomotion in birds (Sholomenko and Steeves, 1987), cats (Brustein and Rossignol, 1998), macaque monkeys (Vilensky et al., 1992), and man (Nathan, 1994).
In contrast to several other studies, our animals did not exhibit any significant recovery during the 4 week postlesion period. Commonly, a period of non-weight-supporting behavior is followed by a quick recovery (3–5 d) or functional improvements over an extended period (1–4 weeks). Rapid recovery may represent the return of function in spared reticulospinal fibers, whereas extended periods of recovery may involve complete lesioning of reticulospinal axons (Brustein and Rossignol, 1998) and a reorganization of descending signals for locomotion, possibly via a DLF pathway (Noga et al., 1991). Reorganization did not contribute significantly to our results because CV-lesioned animals displayed loss of weight support within hours after surgery that did not improve. The quick onset of paralysis was likely the result of acute myelin sheath dysfunction secondary to oligodendrocyte intoxication with EB (Graca and Blakemore, 1986). With evidence of severe behavioral deficits acutely after the CV lesion, one would expect profound locomotor deficits within the same time period after VLF or VC lesions if either region contained myelinated axons necessary for activation of the CPG. No loss of weight support was ever observed in the VLF, VC, or VLF–VC groups.
These data suggest that, when demyelinating lesions are produced within the ventral rat spinal cord and the region is subsequently treated with irradiation to prevent remyelination, reorganization sufficient to affect over-ground locomotor scores is not observed. Possible explanations include the following: (1) cues required to initiate reorganization may not be present in the demyelinated spinal cord in which axonal integrity is more widely preserved than in contusive injuries; and (2) subacute administration of high-dose irradiation (>15Gy) may also be a factor in the observed lack of recovery because Ridet et al. (2000) have demonstrated detrimental effects on recovery after compressive SCI in rats. However, irradiation administered in high doses at 17 d after lesion (Kalderon and Fuks, 1996; Kalderon et al., 2001) or at low doses (2Gy) subacutely (Ridet et al., 2000) have been shown to stimulate axonal sprouting and facilitate recovery of function. High-dose irradiation characteristically prevents remyelination (Blakemore and Patterson, 1978); however, effects on other processes including axonal sprouting, glial scar formation, and angiogenesis, may subserve functionally significant roles.
The specificity of present lesions is also important because other studies have relied on surgical techniques that induce inconsistent damage to areas of the ventral spinal cord. These studies must be interpreted cautiously because results demonstrating correlation between lesioned regions of ventral spinal cord and locomotor deficits have not provided clear histological data that can be used for the design of targeted repair strategies.
Species differences may also play a role in the various results. Extensive ventral white matter lesions in the cat cause only moderate changes in locomotion (Brustein and Rossignol, 1998), whereas in birds (Sholomenko and Steeves, 1987) and rats (present data), similar lesions induce severe locomotor deficits or lasting paraplegia. Complete bilateral ventral hemisections in humans do not result in measurable locomotor deficits (Nathan, 1994). Collectively, these data suggest a considerable degree of interspecies variability in the functional redundancy of locomotor pathways.
Our lesions should not have interrupted action potential conduction in unmyelinated axons, in contrast to the previously used mechanical and selective cooling methods. Although the possibility that unmyelinated axons carried signals for activation of the CPG through the lesioned VLF or VC cannot be ruled out, the latencies of intraspinal field potentials recorded from the spinal cord in cats during MLR-evoked fictive locomotion are consistent with myelination (Noga et al., 1995). However, administration of noradrenergic and serotonergic drugs significantly enhances locomotor function after SCI, suggesting a significant role for unmyelinated ceruleospinal and raphespinal axons in the recovery of locomotor function (Barbeau et al., 1993; Barbeau and Rossignol, 1994; Chau et al., 1998a,b; Brustein and Rossignol, 1999). Based on previous data demonstrating the VLF as essential for MLR-evoked fictive locomotion and the present data, it is likely that the MLR–MedRF–VLF pathway may represent a primary locomotor pathway within a comprehensive locomotor system capable of CPG activation via alternative pathways after VLF lesions.
When the VLF is evaluated in the context of a larger locomotor system, there are several explanations for the lack of locomotor deficits after discrete ventral cord lesions. Local interneuron cell columns, such as the propriospinal tract, may bypass the VLF lesions (Kazennikov et al., 1979; Budakova and Shik, 1980). However, functionally significant propriospinal contributions in this study, if present, must be diffusely arranged in the ventral half of the spinal cord (for review, see Ritter et al., 1999) because VC–VLF and CV lesions produced significant locomotor deficits and flaccid paralysis, respectively, without significant bilateral damage to dorsal horn propriospinal cells of origin. Despite the clear histologic confirmation afforded by the methods used here, each lesion type is a partial interruption of multiple pathways. Pathways likely involved in ventral spinal cord lesions include reticulospinal, raphespinal, ceruleospinal, vestibulospinal, and spinocerebellar. The current status of pathways, including PLR–DLF, ceruleospinal, and raphespinal projections, must be interpreted with caution because redundancy within the locomotor system may exist such that lesioning of the VLF may unmask roles for these pathways in spontaneous locomotion. If multiple tracts carry the signal to initiate locomotion simultaneously, selective lesioning of any one of these tracts would not be expected to remove input to the CPG. Alternatively, partial lesioning of one diffusely arranged tract, as may also be represented here, may not be sufficient to impart large locomotor deficits.
tcMMEP fiber strip
The tcMMEP has been used as a functional measurement of motor pathway damage in common models of SCI (Magnuson et al., 1999) and likely represents a bilateral activation of multiple subcortical structures with axons that descend in the VLF and synapse onto lumbar motoneurons monosynaptically. Previous studies demonstrated a monosynaptic pathway after stimulation of the surgically isolated VLF in neonatal rat spinal cord (Pinco and Lev-Tov, 1994). Although the location of this tcMMEP pathway also corresponds to the region proposed to carry fibers important for MLR-evoked locomotion (Steeves and Jordan, 1980; Jordan, 1986, 1991; Noga et al., 1991), stimulation of the MLR in cats elicited disynaptically and trisynaptically evoked field potentials in lumbar motor nuclei (Noga et al., 1995). Thus, although no conclusions can be made concerning any specific pathway represented by the tcMMEP signal, present evidence suggests that the tcMMEP does not represent a pathway involved in CPG activation. However, the tcMMEP will remain an important tool for the in vivo functional evaluation of SCI therapies as correlations with other voluntary movements are elucidated.
Behavior
VLF and VC lesions resulted in open-field locomotor deficits not fully indicated by the BBB scoring system. VLF- and VC-lesioned animals exhibited toe walking and heavy stepping, respectively. This behavior may represent an extensor bias derived from the lesioning of descending axons within the ventral white matter that act to modulate afferent hindlimb input onto lumbar spinal cord interneurons (Barbeau et al., 1999; Gosgnach et al., 2000). Neither VC- nor VLF-lesioned animals exhibited significant deficits on grid-walking tasks, a behavioral test chosen for its sensitivity in detecting hindlimb sensorimotor deficits (Behrmann et al., 1992). These data suggest that pathways within the lesioned areas of the VC and VLF do not contribute significantly to the sensorimotor control mechanisms needed to accomplish grid-walking tasks.
Implications for spinal cord repair
This study demonstrates that pathways within either the VLF or VC are sufficient for spontaneous open-field locomotion, despite the existence of a discrete fiber strip within the VLF that carries the tcMMEP. Fibers carrying the signals for the initiation of locomotion are likely redundant within the ventral half of the spinal cord because locomotor deficits are proportional to the amount of lesioned ventral white matter. Therapies aimed at the restoration of locomotor function in rat models of SCI should focus on generalized increases in functional white matter within the entire ventral half of the spinal cord because complete repair of all of these pathways may not be necessary.
Footnotes
This work was supported by National Institutes of Health Grant NS38665, Norton Healthcare, and the Commonwealth of Kentucky Research Challenge Trust Fund (S.R.W.), the Kentucky Spinal Cord and Head Injury Research Trust (D.S.K.M.), and the Paralysis Project of America (Q.L.C.). We thank Dr. Larry M. Jordan for his critical comments on this manuscript.
Correspondence should be addressed to Dr. Scott R. Whittemore, Kentucky Spinal Cord Injury Research Center, Department of Neurological Surgery, University of Louisville School of Medicine, 511 South Floyd Street, Medical Dental Research Building 616, Louisville, KY 40292. E-mail: swhittemore{at}louisville.edu.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}