

Abstract
So-called 200 Hz ripples occur as transient EEG oscillations superimposed on physiological sharp waves in a number of limbic regions of the rat, either awake or anesthetized. In CA1, ripples have maximum amplitude in stratum pyramidale. Many pyramidal cells fail to fire during a ripple, or fire infrequently, superimposed on the sharp wave-associated depolarization, whereas interneurons can fire at high frequencies, possibly as fast as the ripple itself. Recently, we have predicted that networks of pyramidal cells, interconnected by axon–axon gap junctions and without interconnecting chemical synapses, can generate coherent population oscillations at >100 Hz. Here, we show that such networks, to which interneurons have been added along with chemical synaptic interactions between respective cell types, can generate population ripples superimposed on afferent input-induced intracellular depolarizations. During simulated ripples, interneurons fire at high rates, whereas pyramidal cells fire at lower rates. The model oscillation is generated by the electrically coupled pyramidal cell axons, which then phasically excite interneurons at ripple frequency. The oscillation occurs transiently because rippling can express itself only when axons and cells are sufficiently depolarized. Our model predicts the occurrence of spikelets (fast prepotentials) in some pyramidal cells during sharp waves.
Ripples are high-frequency EEG waves (180–200 Hz in awake rats in CA1) recorded in hippocampus and other limbic structures (Buzsáki et al., 1992; Ylinen et al., 1995;Chrobak and Buzsáki, 1996), superimposed on physiological sharp waves (Buzsáki, 1986). Ripples occur in both awake animals and anesthetized ones, with frequency slower during anesthesia; the gap junction-blocking anesthetic halothane, however, suppresses ripples, without blocking sharp waves themselves (Ylinen et al., 1995). [Halothane, however, has effects on synaptic transmission (Perouansky et al., 1996) and intrinsic membrane properties (Patel et al., 1999), as well as on gap junctions.] In extracellular recordings in CA1, ripple amplitude is highest in stratum pyramidale (Ylinen et al., 1995), consistent with generation of the waves by perisomatic IPSCs and/or action potential-associated currents. Both pyramidal cells and interneurons, on average, increase their firing rates during the ripple, the relative increase being greatest for pyramidal cells (Csicsvári et al., 1998); nevertheless, pyramidal cells fire no or a few spikes during a ripple, whereas at least some interneurons can fire at ripple frequency. Firing of pyramidal cells, on average, leads firing of interneurons by 1–2 msec (Csicsvári et al., 1999b). In intracellular recordings from pyramidal cells, ripple waves are 1–4 mV in amplitude, and the waves in a particular neuron are phase-shifted relative to the extracellular average signal by intracellular hyperpolarization (Ylinen et al., 1995).
In this paper, we present a network model that accounts for the basic experimental observations on ripples in an economical way. The model uses AMPA and GABAA receptor-mediated synaptic interactions between and within populations of pyramidal cells and interneurons; there is also a transient (150 msec) afferent synaptic excitation of both populations, as would be expected to occur with synaptic excitation of CA1, via Schaffer collaterals during a sharp wave. The model incorporates an additional feature, somewhat novel but critical to model function: gap junctions between axons of the principal neurons. This feature allows the pyramidal axons to act as a high-frequency network oscillator, a “signal generator” in effect, that drives the interneurons. Some general properties of gap junctions are reviewed elsewhere (Bennett and Verselis, 1992; Jefferys, 1995; Perez Velazquez and Carlen, 2000).
The hypothesis of axon–axon gap junctions is based on previous physiological and modeling results. Draguhn et al. (1998) described brief, spontaneous 200 Hz multicellular oscillations in hippocampal slices, generated by principal neurons, dependent on gap junctions but not dependent on chemical synapses. Whole-cell recordings of pyramidal cells in low [Ca2+]o media indicated that local population spikes occurred simultaneously with either intracellular action potentials or spikelets [also called fast prepotentials (Spencer and Kandel, 1961) or d-spikes (Schwartzkroin and Prince, 1977)]. These spikelets were presumed to arise as electrotonic coupling potentials between pairs of neurons. Simulations of pairs of pyramidal cells, coupled by gap junctions in soma or dendrites, could not replicate the form of the spikelets, because the soma-dendritic membranes, together with the gap junction, acted as a low-pass filter of a spike in the presynaptic cell; rise times of the coupling potential were too slow. Electrical coupling between axons, however, easily reproduced the form of spikelets if the gap junction conductance was large enough; a presynaptic spike could then evoke a spike in the coupled axon, which would propagate antidromically in the coupled neuron. If this latter spike failed near the axon initial segment, a spikelet would result.
Knowles and Schwartzkroin (1981) showed that stimulation of CA1 alveus, hundreds of microns from a recorded CA1 pyramidal cell in vitro, could evoke an antidromic spike, or a spikelet, after strong or weak stimulation, respectively. Taylor and Dudek (1982a)observed spikelets in antidromically activated CA1 pyramidal cells in low [Ca2+]o media and attributed them to electrotonic coupling at some point between the neurons. Recently, Schmitz and colleagues (D. Schmitz, A. Draguhn, S. Schuchmann, A. Fisahn, E. H. Buhl, R. Dermietzel, U. Heinemann, R. D. Traub, unpublished data) have obtained electrophysiological evidence that axon–axon gap junctions between hippocampal pyramidal cells play a role in spikelet generation, at least in low [Ca2+]o media.
Gap junctions act to synchronize. Why then would oscillations arise in populations of neurons interconnected by axo-axonal gap junctions? This question was addressed by Traub et al. (1999b) using a variety of modeling approaches (with 3072 detailed compartmental neurons and with a cellular automaton model). It was found that (approximately) synchronized population oscillations could arise under the following conditions. (1) Action potentials could propagate across the gap junctions (in detailed models, the propagation took a finite time, typically <0.5 msec). (2) The density of gap junctions was above the “percolation limit” (Erdös and Rényi, 1960), so that each axon connects to more than one other, on average. (3) The density of gap junctions was low enough that the cells did not fire continuously. By trial and error, it was found that densities of less than ∼2.5–3.0 gap junctions per axon were required, an extremely sparse connectivity. (4) There was a background of randomly occurring, spontaneous action potentials. These could occur at low frequency (as low as 0.05 Hz in some cases). The resulting population oscillations could easily occur at 150 Hz or faster. The mechanism appears unique; the oscillation period is not determined by membrane refractoriness or by synaptic time constants, but rather by (1) topological structure of the network and (2) the time for a spike in one axon to induce a spike in a connected axon.
In short, a network of pyramidal axons can act, in principle, as a signal generator at frequencies above 100 Hz. In the added presence of AMPA and sufficiently strong GABAAreceptor-mediated interactions, it has been shown that a system of pyramidal neurons and interneurons can produce sustained γ oscillations dependent on gap junctions (Traub et al., 1999c). Here, we illustrate how ripple oscillations might also be generated.
MATERIALS AND METHODS
The network model used in this paper is a hybrid of the models described by Traub et al. (1999a,b). That is, there is an array of pyramidal cells and interneurons, interconnected (randomly or locally randomly) by chemical synapses. This array, originally developed to study tetanically induced γ and β oscillations, is almost exactly as described in Traub et al. (1999a). (The slight differences are listed below.) To this “chemical synaptic” array, we add a pyramidal axon–axon gap junction connectivity, also defined further on.
Briefly, the network consists of 3072 pyramidal cells and 384 interneurons. Each neuron is multicompartmental and includes five axonal compartments (Traub et al., 1994; Traub and Miles, 1995). The pyramidal cells are arranged in a 96 × 32 array and the interneurons in a 96 × 4 array. The interneurons are divided into four classes of 96 cells each: “basket cells,” inhibiting perisomatic regions; “axo-axonic cells,” inhibiting axon initial segments; and two types of dendrite-contacting interneuron (for more proximal or more distal dendrites, respectively). The time constant of ISPCs induced by basket cells and axo-axonic cells was 10 msec but was 50 msec for dendrite-contacting interneurons. Interneurons other than axo-axonic cells also synaptically contact other interneurons. Synaptic connectivity of pyramidal cells was global, but of interneurons was confined to take place only to cells at most 500 μm away (25 cell diameters along the long axis of the array). Each pyramidal cell receives input from 30 other pyramidal cells and from 80 interneurons. Each interneuron receives input from 60 other interneurons. Only AMPA and GABAA receptor-mediated synaptic interactions were simulated, not NMDA, metabotropic, or GABABmediated. Further structural details can be found by Traub et al. (1999a).
Intrinsic properties of the individual neurons were exactly as described by Traub et al. (1999a), with the exception that gK(AHP) density was uniformly 0.8 mS/cm2 over the soma–dendrites of pyramidal cells, as in the original paper (Traub et al., 1994). The time course of unitary EPSCs in the present study was as follows: 0.75 × t ×e(−t/2) nS on pyramidal cells, and 0.5 × t ×e−t nS on interneurons. Peak unitary IPSC values were 2 nS on pyramidal cells, 1 nS for basket cell-mediated inhibition of interneurons, 0.1 nS for other IPSCs on interneurons. These values are the “default” values used in this study; when modified in the figures, it will be duly noted.
Gap junctions (nonrectifying, voltage-independent) were located between the penultimate axonal compartments (centered 263 μm from the soma) of randomly selected pairs of pyramidal cell axons, subject to the constraint that the respective somata were within 200 μm of each other. The default gap junction resistance (used unless specified otherwise) was 238 MΩ (conductance of 4.2 nS); this resistance allowed action potentials to cross from one axon to the other, at least sometimes (Traub et al., 1999b).
The average number of gap junctions lying on an axon was 1.6. To remind the reader of the structural implications of this density of gap junctions (Erdös and Rényi, 1960; Traub et al., 1999b), this density lies above the percolation limit of one gap junction per cell. This means that a “large cluster” will exist and that all cells not on the large cluster are either isolated or lie on small clusters. [Definitions are as follows: a “cluster” of cells is a set of cells such that if cell A and cell B both are in the set, then a gap junctional path A→ (zero, one, or more intermediate cells, themselves connected by gap junctions) → B exists, with the intermediate cell(s) also lying in the set. With low-connection densities, the number of intermediate cells in the path may be big. A cluster is “large” if its size is of the same order as the whole network; otherwise the cluster is “small.”]
Some structural properties of this particular gap junction network were noted by Traub et al. (1999b). For example, the large cluster has 1990 cells, and the next largest cluster is small, having only 13 cells. The mean path length (which measures the average length of the shortest gap junctional path between randomly selected cell pairs) on the large cluster is ∼17.5. A block diagram of the network model is shown in Figure 1.

Block diagram of network model. The model contains 3072 pyramidal cells and 384 interneurons (basket cells, axo-axonic cells, and dendrite-contacting cells), with intrinsic properties and connectivity described in Materials and Methods. The figure highlights the conceptual distinction we make between the axons of the pyramidal cells and the soma–dendritic membranes. Interactions between components of the model are shown. Afferent inputs in the model are purely excitatory. Only AMPA receptors are simulated for recurrent excitatory connections, and only GABAA receptors for recurrent inhibitory connections.
As in previous studies (Traub et al., 1999a,b), noise was simulated by the generation of random ectopic spikes, originating from small current pulses applied to the most distal compartment of the five-compartment axon. The current pulses had Poisson statistics (as approximated with a pseudorandom number generator), independent between different axons, and occurred once per 5 sec in interneurons; in pyramidal cells, in the figures illustrated here, current pulses were applied on average once per second, but mean intervals of 0.1–10 sec were also tried. In our model of high-frequency oscillations in axonal networks (Traub et al., 1999b), ectopic axonal spikes played a critical role.
Sharp waves were produced by applying a 150 msec excitatory conductance pulse (reversal potential of 60 mV positive to rest) to both pyramidal cells and interneurons: the “afferent input.” The conductance developed in four compartments of the midapical dendrites of pyramidal cells and likewise in four dendritic compartments of the interneurons. The onset of the conductance was spread (randomly and uniformly) over a 1 msec interval for both cell types. The shape of the conductance pulse was a cosine function, peaking at 60 nS for pyramidal cells and 4 nS for interneurons. Further variability was introduced into the system by having different bias currents applied to the somata of the pyramidal cells (−0.15 to −0.05 nA, randomly and uniformly distributed).
Two sharp waves per simulation were generated, separated by 250 msec. These waves had a similar appearance, although pyramidal cell afterhyperpolarizing conductances were larger in the second wave than in the first. For the sake of consistency, figures were made using data from the second sharp wave of any given simulation.
Simulation programs saved the following types of data: somatic voltages of selected pyramidal cells (some on the large cluster, some not) and interneurons; voltages in the axon (at the site of the gap junction) of selected pyramidal cells; average signals, consisting of somatic voltages of 224 nearby pyramidal cells or of 28 nearby interneurons; and total GABAA synaptic conductance to a pyramidal cell, recurrently generated AMPA conductance to a pyramidal cell, afferent AMPA conductance to a pyramidal cell, and afferent AMPA conductance to an interneuron. Autocorrelations and cross-correlations were computed using the middle 75 msec of data from the simulated sharp wave. Figures that illustrate an “axonal” signal use the voltage in the respective axon at the site of the gap junction.
Programs for simulation and analysis were written in FORTRAN. Simulation programs were written in FORTRAN augmented with instructions for a parallel computer. These latter programs were run on an IBM SP2 with 12 nodes (processors); other programs ran on a single node of the SP2. Some comments on numerical methods are discussed by Traub et al. (1999a). Each simulation was of 450 msec of activity and took ∼2.7 hr. For further details, please contact Roger D. Traub atr.d.traub{at}bham.ac.uk.
RESULTS
The basic properties of simulated ripples in our model (using default parameters) are shown in Figures2 and 3. The afferent input depolarizes the pyramidal cells and, to a lesser extent, the interneurons (Fig.2A,B). An ∼140 Hz oscillation, predominantly subthreshold and ∼2 mV in amplitude, becomes apparent in pyramidal cells, and (as is clearly visible in the average signal) this oscillation is coherent (Fig.2A,B). During the sharp wave-associated depolarization, interneurons begin to fire at high rates. The interneuron in Figure 2A fires at ripple frequency, although not all interneurons do. [Note that the afferent input itself is not enough to induce this high-frequency firing in our model, as shown by a simulation with recurrent excitatory synapses blocked (data not shown).] On average, the mean pyramidal cell signal leads the mean interneuronal signal by 1.3 msec (Fig. 2C), comparable with in vivo experimental data (Csicsvári et al., 1999b).
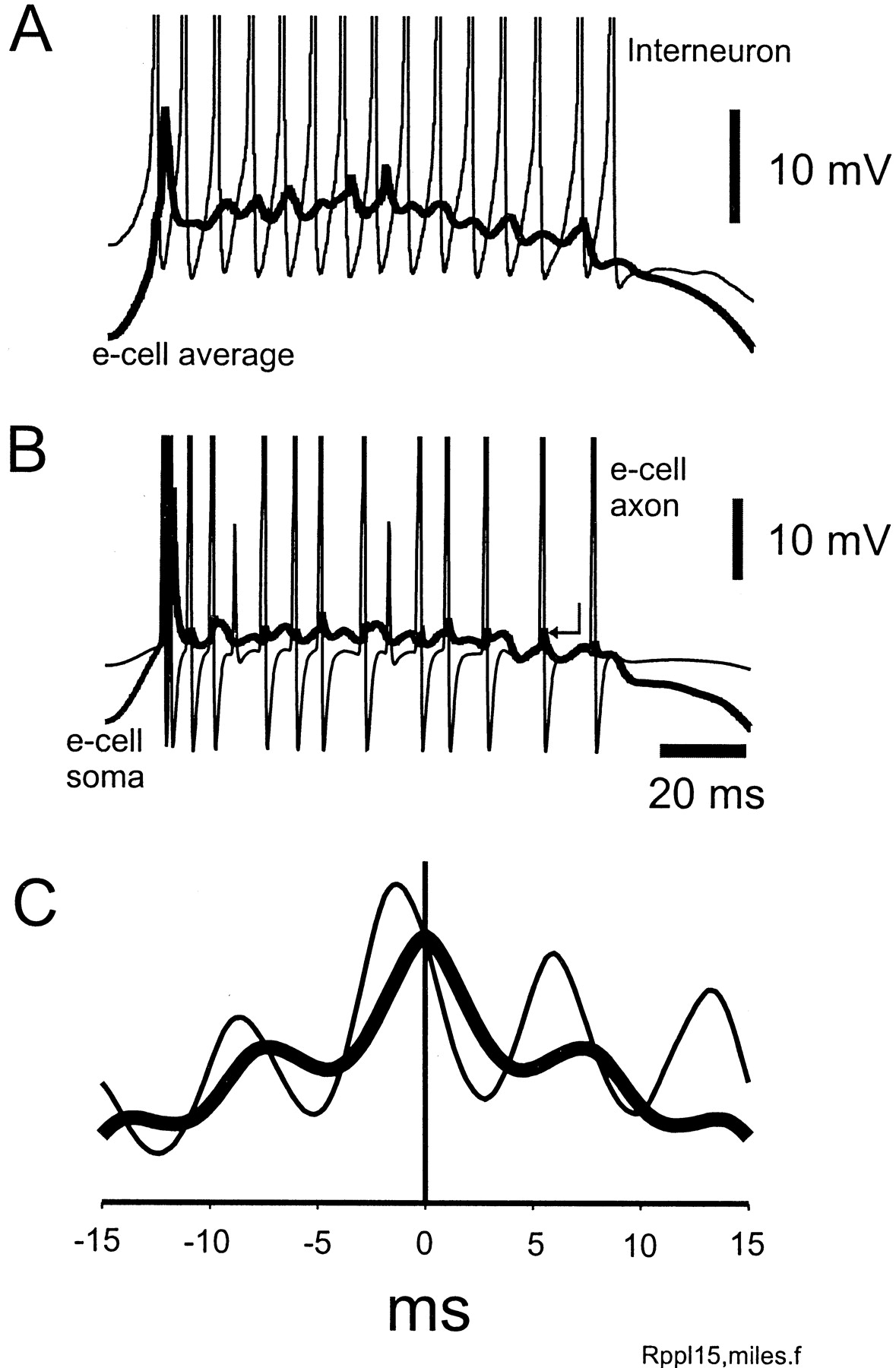
Firing properties of pyramidal cells and interneurons during a simulated ripple. Default parameters were used (see Materials and Methods). A, Average voltage of 224 nearby e-cell somata (thick line) reveals 140 Hz oscillation superimposed on a cellular depolarization. An interneuron (thin line) fires at the same frequency. Spikes truncated. B, Single pyramidal cell (thick line) fires once during the ripple but displays intracellular ripple waves, ∼2 mV in amplitude. The axon of the cell (thin line) fires at more than ripple frequency. Many axonal spikes are followed by ∼1–2 mV spikelets, one of which is indicated by an arrow. [This cell was on the large cluster (see Materials and Methods). Axons of cells off the large cluster do not fire at ripple frequency, and there are virtually no spikelets, although intracellular rippling still occurs (data not shown).] Spikes truncated. Signals in A andB are simultaneous. C, Autocorrelation of local average pyramidal cell signal (thick line) shows rhythmicity at 140 Hz. Cross-correlation of local average pyramidal cells with local average interneurons (thin line) shows that pyramidal cells lead interneurons by 1.3 msec, on average. (Cross-correlation shifted and rescaled on vertical axis.)

During simulated ripple, the pyramidal cell axonal plexus oscillates at high frequency and phasically drives interneurons.A, Interneuron (thin line, spikes truncated) fires at ripple frequency. The AMPA receptor-mediated input to this interneuron (thick line) has a slow (afferent) component lasting >100 msec and a phasic component at ripple frequency. The phasic waves of synaptic excitation lead the action potentials. B, The AMPA input to the interneuron (thick line) lags the oscillatory output axonal network (thin line), plotted as the number of axons depolarized above 70 mV from rest. On average, the local mean interneuron potential lags this axonal signal by 1.5 msec. Data from simulation as in Figure2. Signals in A and B are simultaneous.
Rippling arises in our model as follows. In between the sharp waves, distal axons are hyperpolarized by several millivolts, which suppresses the ectopic spikes (data not shown). As the sharp wave begins, the axonal hyperpolarization is relieved, and ectopic spikes begin and percolate throughout the axonal plexus, generating a high-frequency network oscillation (Fig. 3B). The detailed mechanisms of this oscillation are analyzed by Traub et al. (1999b), but, briefly, the main ideas are these: if gap junctions allow action potentials to cross from one axon to another and if each axon connects (on average) to more than one other, then an ectopic action potential, arising in a single axon, can lead to a wave of activity that spreads through the axonal network. The pattern of this wave does not depend on the detailed kinetics of the active currents in the axon, provided that postspike refractoriness is less than 1 or 2 msec. The period of the oscillation is determined by two main factors: (1) the time it takes for an action potential to cross from one axon to another (expected to be <0.5 msec), and (2) the length of “chains” of gap junctions crossed as the original ectopic spike spreads to the bulk of the axonal population (expected to be in the range of 10–20 or so). The oscillation period is approximately the crossing time multiplied by the expected length of such chains. Our model predicts that small decreases in junctional conductance, by prolonging the time it takes for spikes to cross gap junctions, should slow the period, whereas factors that increase connectivity of the network (i.e., a greater number of gap junctions) should decrease the period. The latter effect is expected because, in a more connected network, an ectopic spike can excite the bulk of the population over relatively shortened chains of gap junctions.
The effects of the axonal network oscillation (Fig. 3) are twofold. First, antidromic propagation into pyramidal cells (lying on the large cluster) can evoke spikes or spikelets within <1 msec (Fig.2B, arrow). With the strength of IPSCs used, however, the probability of full-spike invasion of pyramidal cells is low, and pyramidal cells fire rarely during the ripple (see further on). [Note that for pyramidal cells not lying on the large cluster, spikelets will not be seen; instead, the ripple appears purely as synaptic potentials (data not shown).] Second, the coherent orthodromic output of the axonal plexus induces, after a very short delay, phasic AMPA receptor-mediated EPSPs in interneurons, which entrain their firing (Fig. 3A,B). Note that orthodromic synaptic excitation of interneurons can take place without the parent pyramidal cell soma necessarily firing.
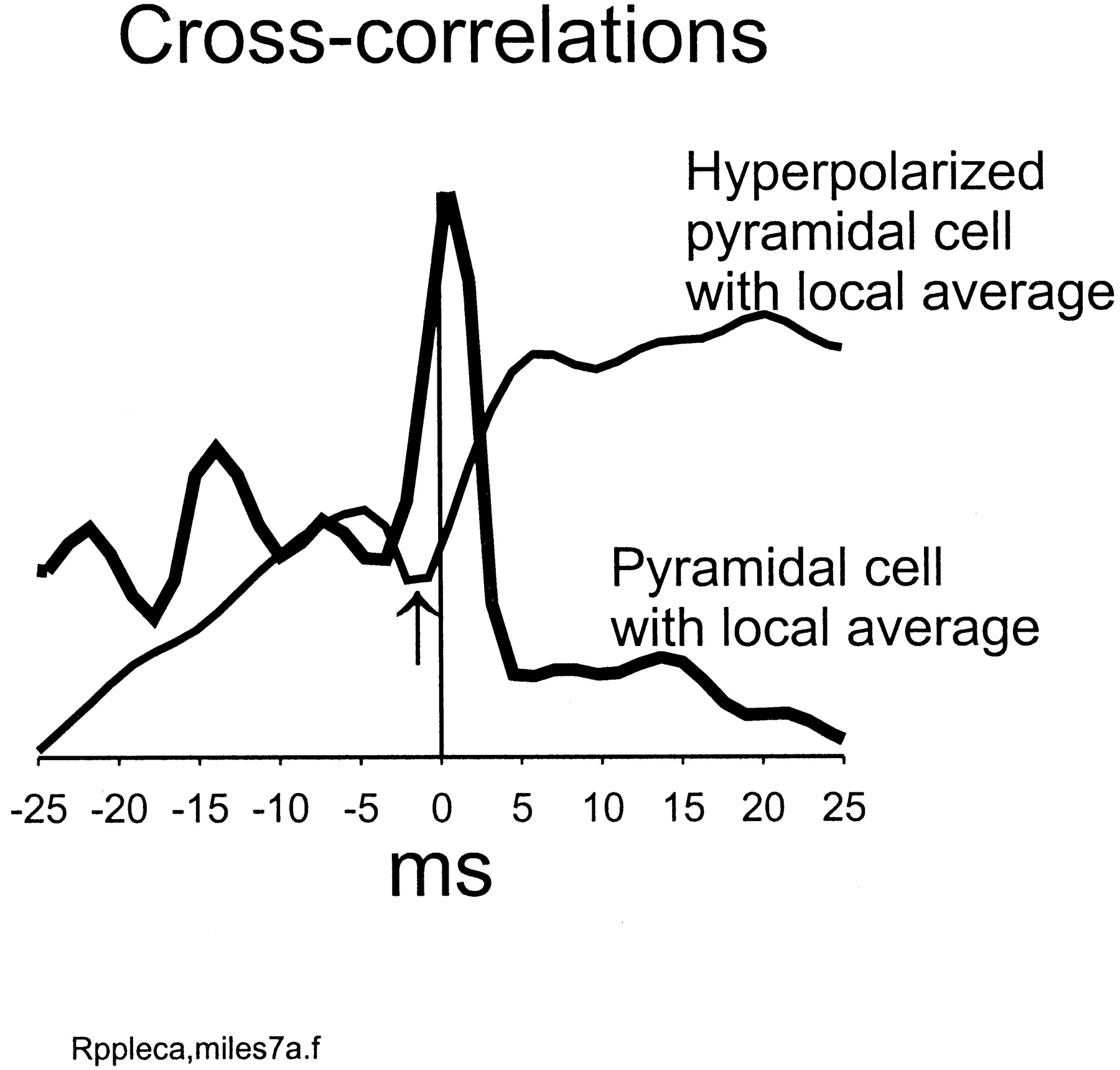
When a pyramidal cell somatic voltage was correlated with the local average pyramidal signal, the peak occurred near 0 msec (Fig.4), not surprisingly. When the same cell was hyperpolarized ∼20 mV during the ripple, it became nearly anticorrelated with the local average signal (Fig. 4), again in agreement with others (Ylinen et al., 1995). This is consistent with the notion that the ripple waves are predominantly IPSPs. This was checked in the model by plotting the actual synaptic conductances “seen” by a pyramidal cell during the rippling (data not shown). Fluctuations in AMPA receptor conductance were ∼5 nS in amplitude, superimposed on the afferent excitatory conductance. In contrast, fluctuations in GABAA receptor conductance were 25–50 nS. There was, additionally, a slower component to the GABAA conductance, produced by the dendrite-contacting interneurons (whose ISPCs are relatively slow; see Materials and Methods).

Hyperpolarizing a pyramidal cell, during a ripple, shifts its phase relative to the local average signal. Using data from the central portion of the ripple in the simulation of Figures 2 and 3, a pyramidal cell somatic signal was cross-correlated with the local average signal (224 pyramidal cells) (thick line). The peak is near 0 msec (0.8 msec). The simulation was repeated, with the index pyramidal cell hyperpolarized ∼20 mV during the ripple, relative to control. The cross-correlation of the index cell with the local average (thin line) now reveals a minimum near 0 msec (−1.5 msec) (arrow).
Rippling in the model requires that gap junctional conductance be sufficiently large, consistent with in vivo data showing that halothane suppresses ripples (Ylinen et al., 1995) (Fig.5). When the gap junctional conductance was reduced 25% from its default value, a γ-frequency rhythm was superimposed on the sharp wave (Fig. 5A), and many of the pyramidal cell action potentials were orthodromic. Partial spikes were seen in the axon, indicative of failed conduction of action potentials across gap junctions. When gap junctional conductance was reduced only 2.5% relative to default (Fig. 5B), ripple waves begin to occur but are not sustained, being broken up at ∼20 msec intervals. Spikelets are also to be noted in the pyramidal cell soma. Finally, for comparison, Figure 5C illustrates rippling, using data from the simulation of Figures 2 and 3. When the gap junctional conductance was increased an additional 12.5% above default value, rippling would still occur but at higher frequency (∼200 Hz; data not shown).

Oscillatory behavior in the model depends on axo-axonal gap junction conductance. Each panel shows the soma of a pyramidal cell lying on the large cluster (thick line) and the axon of the cell (thin line). Spikes are truncated. Gap junctional conductance is expressed relative to the constant c = 10.5 nS and increases in sequenceA→B→C.A, With a small gap junctional conductance, the axonal network cannot sustain high-frequency oscillations. Cellular oscillation occurs at γ frequency (∼50 Hz), and action potentials are orthodromic. B, At a higher gap junction conductance, there is evidence of sustained axonal network activity; the soma fires only once, but the axon fires seven times, and spikelets occur in the soma. C, With a still higher gap junction conductance (as used in Figs. 2 and 3), continuous high-frequency activity can occur in the axonal network. Individual pyramidal cells express a high-frequency oscillation as well, with a synaptic component, and, if the cell is on the large cluster, spikelets occur also.
Traub et al. (1999b) showed that axon–axon gap junctions could produce a high-frequency oscillation superimposed on a spontaneous, synchronized “epileptiform” burst in a disinhibited population of pyramidal cells, interconnected by recurrent excitatory chemical synapses. This is a type of event that occurs in disinhibited CA3 (Wong and Traub, 1983). Figure 6 demonstrates that axo-axonal gap junctions can produce a high-frequency oscillation (∼160 Hz) in an afferently excited, disinhibited population of pyramidal cells, as would occur in CA1, as it responds to excitatory input from CA3 (Schwartzkroin and Prince, 1977; Wong and Traub, 1983). In this case, pyramidal cells (lying on the large cluster) fire full spikes and also 10–20 mV partial spikes during the burst (Fig.6C). This type of activity occurs in a given cell (provided it lies on the large cluster) at the same frequency as the population activity.
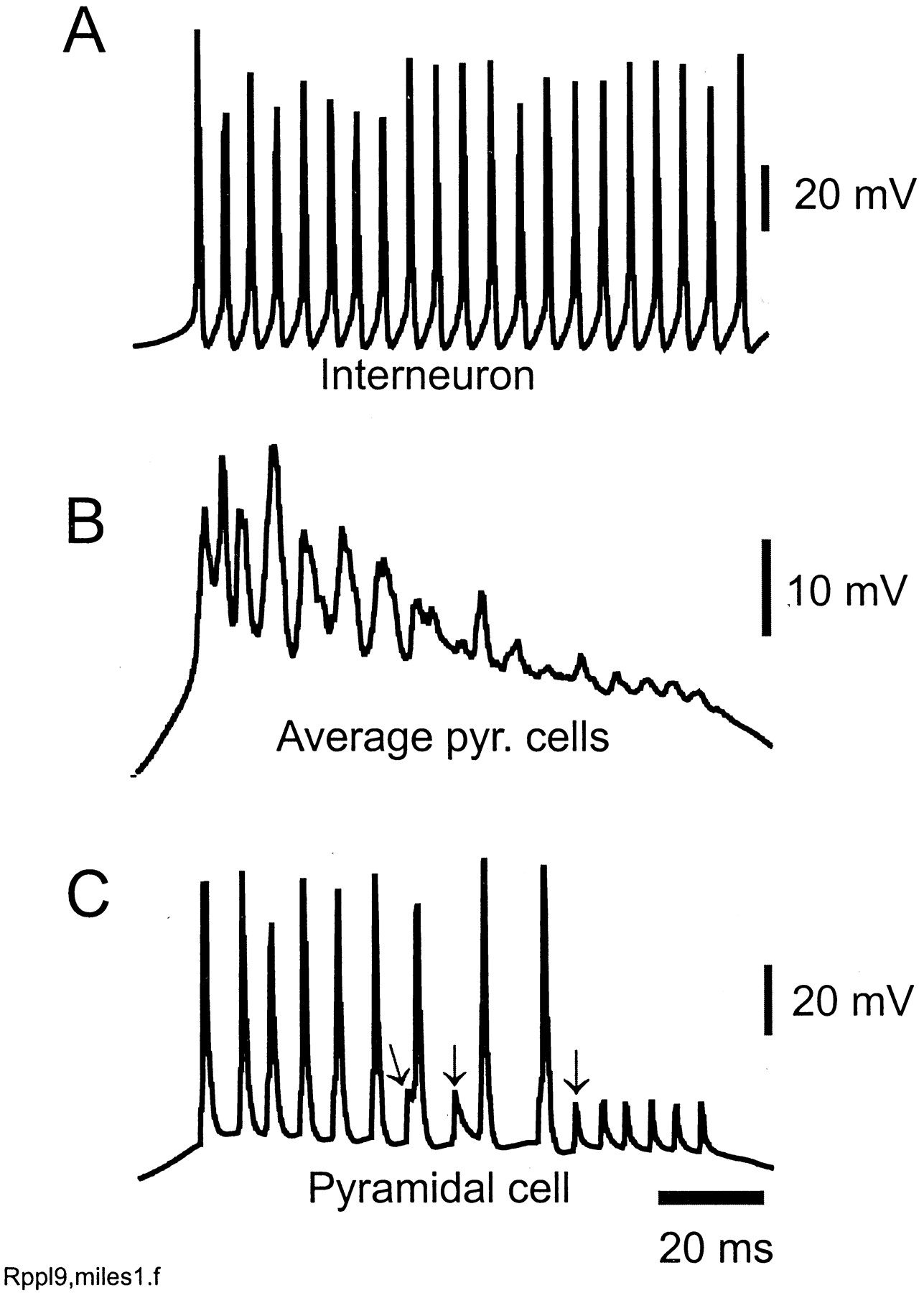
Blocking GABAA receptors allows a ripple frequency oscillation to persist, but pyramidal cell firing increases. Same parameters as in the simulation of Figure 2, but all GABAA conductances set to zero. Traces inA–C are simultaneous. Cells on the large cluster (as inC) exhibit 10–20 mV partial spikes (arrows), deriving from partially blocked antidromic activity. Many of the full action potentials are also antidromic; the axonal network here acts as a high-frequency signal generator, driving pyramidal cells antidromically, as well as orthodromically. Compare the high-frequency oscillation superimposed on CA1 epileptiform field potentials (Schwartzkroin and Prince, 1977) (see also Traub et al., 1999b).
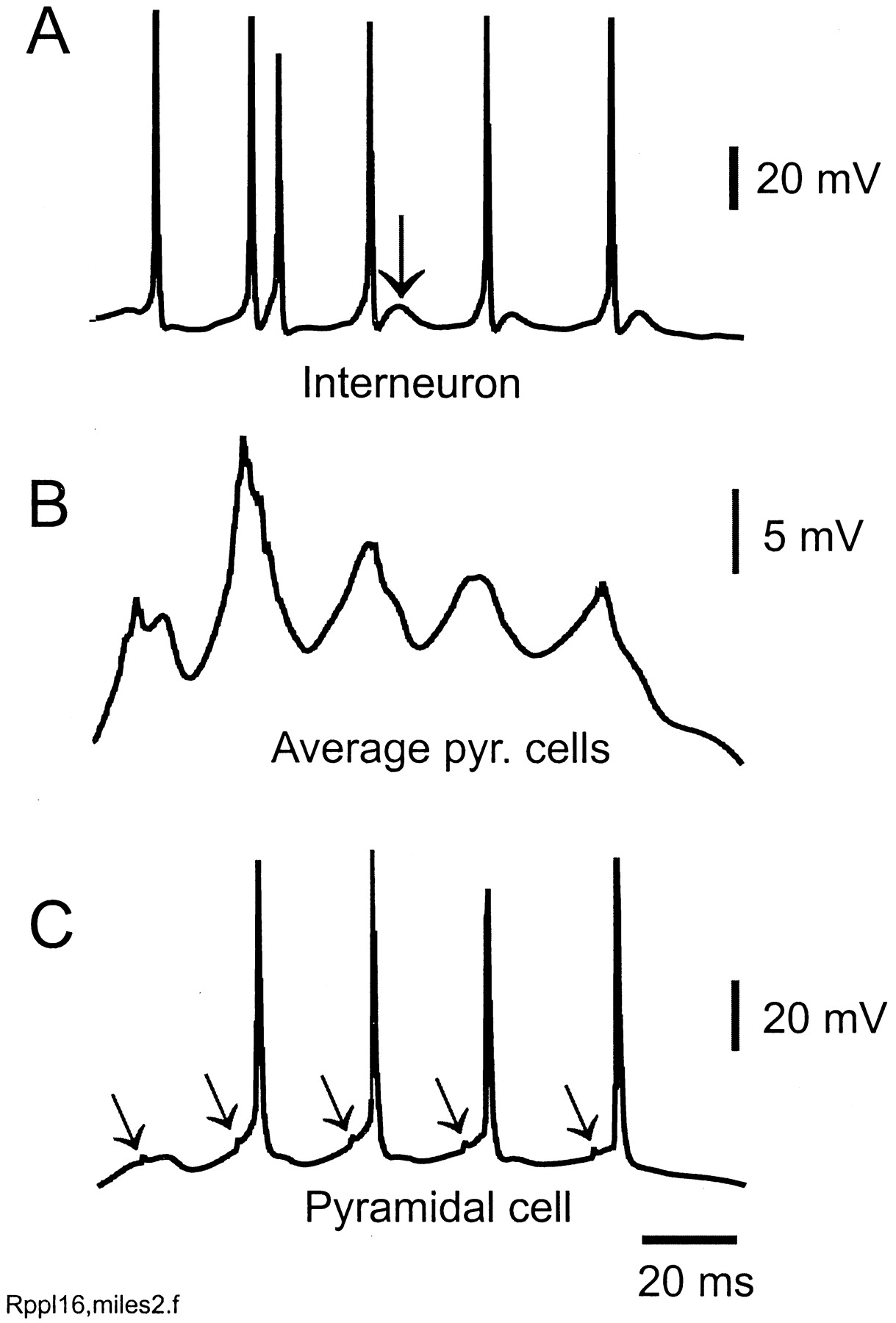
Rippling in the model also requires that IPSCs on pyramidal cells not be excessively large. Figure 7illustrates a simulation with parameters identical to the simulation of Figures 2 and 3, except that unitary IPSCs on pyramidal cells are 50% larger. In this case, a γ-frequency oscillation occurs instead of rippling. [It is interesting that transient runs of γ-frequency oscillations have been recorded in CA1 in vivo(Csicsvári et al., 1999a).] IPSCs interrupt the firing of interneurons (Fig. 7A), the axonal population (data not shown), and pyramidal cells (Fig.7B,C). That axonal activity still can take place is shown by the occurrence of spikelets in at least some of the pyramidal cells (Fig. 7C, arrows).

Increasing GABAA IPSCs on pyramidal cells converts the ripples to a γ-frequency oscillation (∼50 Hz in this case). Same parameters as in the simulation of Figure 2, but unitary IPSCs on pyramidal cells are 50% larger. In this case, the axonal network is spontaneously active (Fig. 8A), but the activity is not continuous, being broken up by IPSPs. The network of depolarized pyramidal cells and interneurons then produces γ oscillations, reminiscent of models of tetanically induced γ (Traub et al., 1999a) and of glutamate-induced γ (Burchell et al., 1998), but with the difference that spontaneous axonal activity is prominent here. Traces in A–C are simultaneous. A, Interneuron showing doublets and spikes followed by EPSPs (arrow) (cf. Whittington et al., 1997). Average pyramidal signal (B) and pyramidal cell (C) do not exhibit ripple waves. Note the spikelets in the pyramidal cell (oblique arrows).
Figure 8 illustrates the transition from γ-frequency oscillation to rippling as the conductance of pyramidal IPSCs is reduced. (Other parameters are as in the simulation of Fig.2.) Here, one can see how sufficiently large IPSCs break up the high-frequency firing of the axon, either repeatedly (Fig.8A, arrows) or intermittently (Fig.8B, arrow).

Transition from γ to rippling as synaptic inhibition is reduced. The simulation of Figure 2 (same as Fig.8C) was repeated with identical parameters, except for the conductance of IPSCs on pyramidal cells. Each panel shows the potential of a pyramidal cell soma and its axon (same cell in all of the panels). A, γ rhythm. B, Rippling, although axonal activity is not continuous. C, Rippling, with continuous axonal activity. Note in A andB the IPSP-induced hyperpolarizations in the axonal potential (some marked with arrows).
DISCUSSION
To place our model results in perspective, we shall consider briefly four possible models of rippling and attempt to identify experiments that could distinguish between the different models. Of course, these models could possibly work in conjunction with one another in various combinations, but it is premature to enter into the resulting complexities. We suspect that ephaptic interactions, as appear to contribute to field burst synchronization (Jefferys and Haas, 1982; Taylor and Dudek, 1982b; Konnerth et al., 1984), are not of primary importance during ripples, because the extracellular potentials are a fraction of a millivolt (Ylinen et al., 1995); hence, we consider below only mechanisms based on chemical synapses and/or gap junctions.
Could rippling result from slow excitation of pyramidal cell and interneuron populations, together with purely chemical synaptic interactions? The closest experimental paradigms in vitrowould be the oscillations induced in CA1 or CA3 by tetanic stimulation (Whittington et al., 1997), by glutamate application (Burchell et al., 1998), or by carbachol (Fisahn et al., 1998). Rippling is not apparent in any of these experimental protocols, although it is conceivable that IPSPs are large enough to suppress rippling (Figs. 7, 8). However, tetanically induced oscillations in CA1 in vitro, in the presence of bath-applied morphine (which suppresses GABA release), also do not show rippling (Whittington et al., 1998, their Fig. 1). In addition, tetanic oscillations in morphine exhibit population spikes, with pyramidal cells firing at γ frequencies (Whittington et al., 1998; Faulkner et al., 1999), a cellular behavior not seen during ripples. This hypothesis also would not explain the suppression of ripples by halothane. Nevertheless, in vivo, it is conceivable that there is relatively more excitation of interneurons than of pyramidal cells during a sharp wave, occurring in some manner not captured by the slice protocols.
Could rippling result from GABAergic interactions between interneurons, in effect, ultrafast “interneuron network γ?” In experimental studies of interneuron network γ, in CA1 in vitro, application of l-glutamate produced frequencies only as high as 49 Hz (Traub et al., 1996, their Fig. 1). Again, one might consider that synaptic inhibition in the experimental situation is more effective than during a sharp wave. Note, however, that even in bicuculline (up to 5.0 μm, just short of blocking the oscillations), frequencies only as high as 65 Hz occurred (Traub et al., 1996, their Fig. 3). In morphine, a drug that reduces GABA release, epochs with frequencies as high as ∼90 Hz could be observed in pharmacologically isolated interneuron networks (Whittington et al., 1998, their Fig. 2); therefore, this hypothesis cannot be dismissed too lightly.
Theoretical arguments weigh against ultrafast interneuron network γ; as the oscillatory period becomes short relative to the time constant of GABAA IPSC relaxation, then the effects of heterogeneity in cellular excitability become more and more pronounced, and the oscillation breaks up (Wang and Buzsáki, 1996; Chow et al., 1998; White et al., 1998). If this time constant is ∼10 msec (Traub et al., 1996), then it has been argued (Chow et al., 1998) that 200 Hz oscillations in interneuron networks cannot be stable. One must be open to the possibility, however, that some subset of interneurons could exist, mutually inhibitory with extremely fast IPSCs, faster than those normally encountered. Even then, this hypothesis would not explain the halothane suppression of ripples.
Could ripples result from purely interneuron networks, interconnected by gap junctions (Galarreta and Hestrin, 1999; Gibson et al., 1999)? Such gap junctions might, in principle, occur between dendrites (as recognized ultrastructurally; Kosaka, 1983) or, hypothetically, between axons. Networks of interneurons interconnected by dendritic gap junctions have been shown, using simulations, to be capable of generating autonomous network bursts (Traub, 1995), but only under conditions in which the dendrites are excitable enough to support action potential initiation. Interneuron networks interconnected by axon–axon and dendrite–dendrite gap junctions can also (in simulations) generate fast population oscillations (R. D. Traub, unpublished data) by the same mechanisms as could work for pyramidal cell networks, provided that mutual GABAergic inhibition between the interneurons is suppressed. The hypothesis of gap junctionally coupled interneuron networks is consistent with the halothane data. If the hypothesis is correct, one would expect to find evidence of axonal activity in interneuronal recordings in vivo: antidromic spikes, partial spikes, or spikelets.
Could ripples result primarily from network oscillations generated in pyramidal cell axon networks, with gap junctional interconnections? This is the hypothesis explored in the present paper. The “core” of the ripple generator is as shown, in block diagram form, in Figure9A. The pyramidal cell axonal network acts as a high-frequency signal generator, working when (1) gap junctional conductance is large enough for spikes to cross gap junctions (Traub et al., 1999b) (Fig. 5), and (2) the axons are not too hyperpolarized. Afferent excitation of pyramidal cells can overcome axonal hyperpolarization and allow the signal generator to operate (Figs. 2, 3). The output of the axonal signal generator phasically excites interneurons and induces many of them to fire at high frequency (Figs. 2, 3). (In CA1, recurrent excitation of pyramidal cells is of less importance than the excitation of interneurons, especially with IPSPs present.) The degree of pyramidal cell firing is regulated by the strength of inhibition on pyramidal cells (Fig. 6); our proposed mechanism can in principle work, however, without any firing by the pyramidal cell somata. If, on the other hand, IPSPs are too large, then axonal activity is broken up and a γ-frequency oscillation results. This latter behavior is similar to what we propose happens during carbachol- and kainate-induced oscillations in CA3 in vitro(Traub et al., 1999c). This hypothesis is consistent with the disruption of ripples by halothane.

Block diagram of subnetworks believed critical for generating high-frequency (>100 Hz) oscillations in interneuronal populations and for gap junction-sensitive γ oscillations.A, The pyramidal cell axons, interconnected by gap junctions, act as a high-frequency signal generator, the output of which phasically excites interneurons via AMPA receptors. (Each interneuron will, of course, be excited by many axons.) Some of the interneurons follow the phasic input at high frequency.B, If the interneurons also inhibit the axon initial segments powerfully enough, the high-frequency output of the axon network can be modulated at a frequency determined by GABAAIPSC time course; a γ-frequency rhythm could thereby arise (Traub et al., 1999c).
If our proposed mechanism is correct, there are two key experimental implications. First, there should be evidence of antidromic activity in some (not all) CA1 pyramidal cells during ripples: antidromic spikes, partial spikes, or spikelets. Second, there should be evidence of ripple frequency EPSPs in hyperpolarized CA1 interneurons during sharp waves. Because of afferent excitation of CA1 via Schaffer collaterals during the sharp waves, recordings would optimally be made in interneurons receiving minimal Schaffer input but which are recurrently excited by CA1 pyramidal cell axons (Sik et al., 1995; Freund and Buzsáki, 1996; G. Buzsáki, personal communication).
Footnotes
This work was supported by the Wellcome Trust. R.D.T. is a Wellcome Principal Research Fellow. We thank Eberhard Buhl and György Buzsáki for helpful discussions.
Correspondence should be addressed to Roger D. Traub, Division of Neuroscience, University of Birmingham School of Medicine, Vincent Drive, Edgbaston, Birmingham B15 2TT, UK. E-mail:r.d.traub{at}bham.ac.uk.





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}